
Adjuvanted Vaccine Induces Functional Antibodies against Pseudomonas aeruginosa Filamentous Bacteriophages


Abstract
1. Introduction
2. Materials and Methods
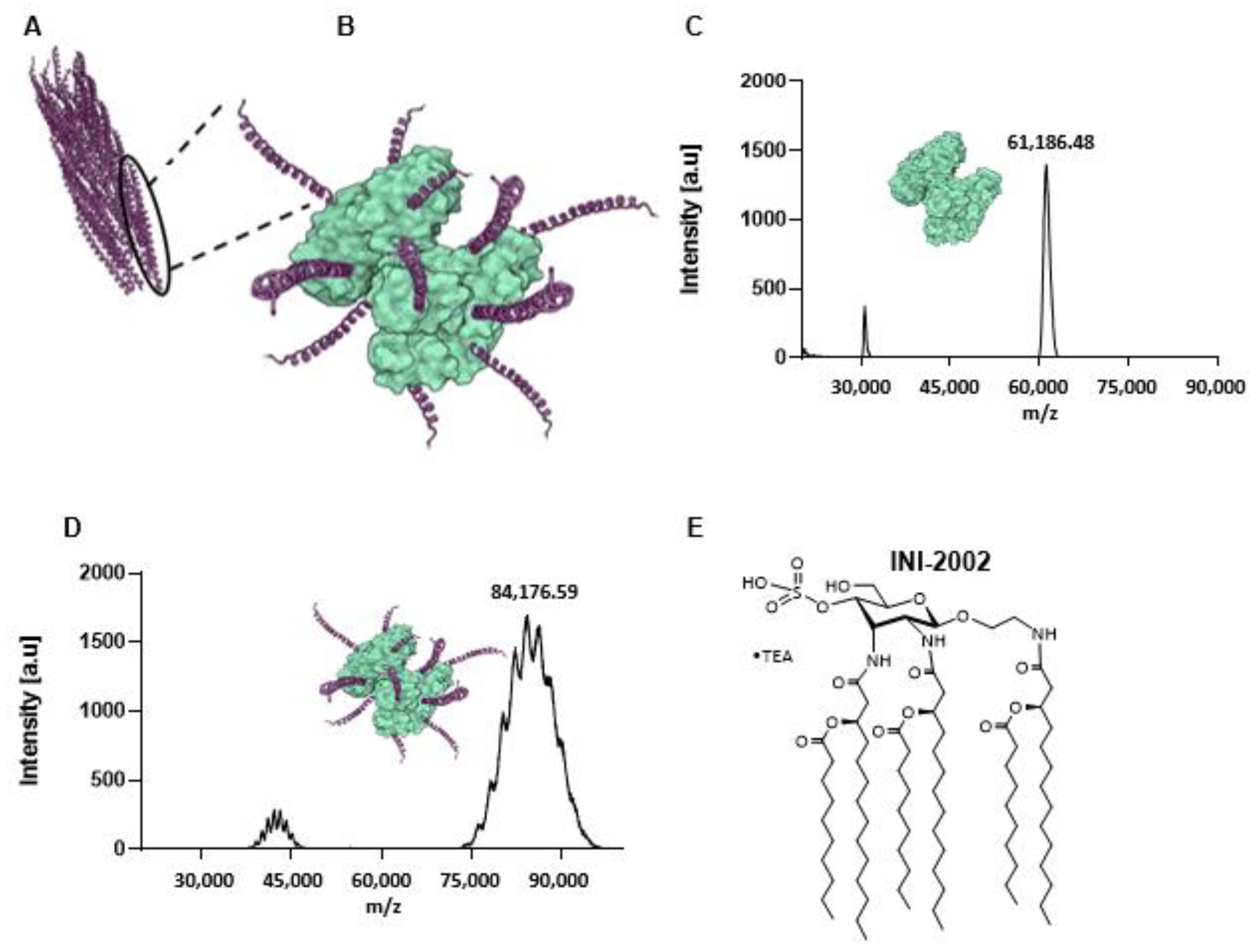
2.1. INI-2002 Compound Synthesis
2.2. CoaB Conjugate to CRM
2.3. Bacterial Strains, Plasmids, and Growth Conditions
2.4. P. aeruginosa and Pf4 Propagation
2.5. In Vivo Experiments
2.6. Pf4 Serum Inhibition Assay
2.7. ELISA for Anti-CoaB Peptide Antibody Quantification
2.8. Phage ELISA
2.9. Cell-Mediated Immunity Analysis
3. Results
3.1. Production and Characterization of INI-2002 Adjuvanted CoaB Peptide-CRM Conjugate Vaccine
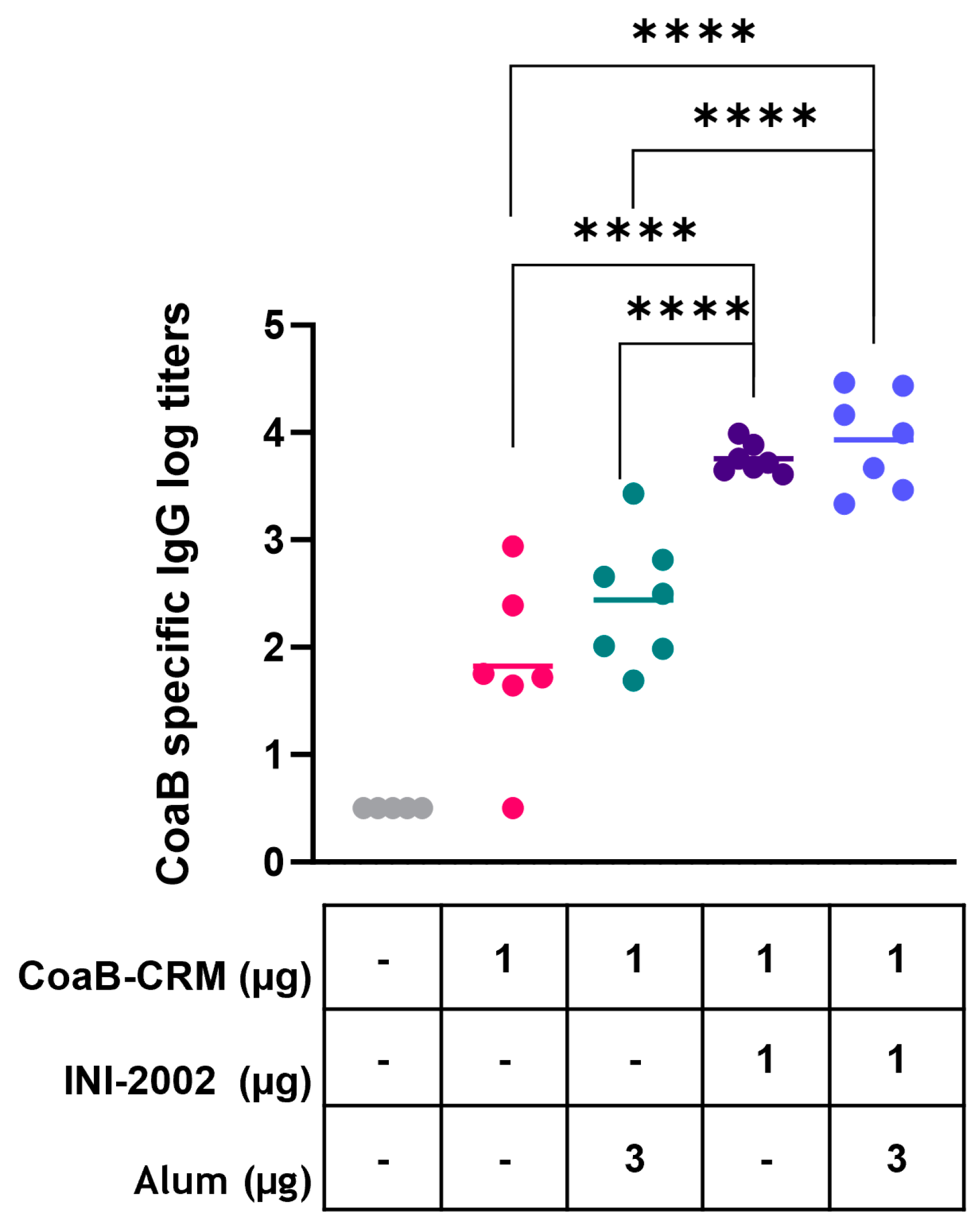
3.2. INI-2002 Adjuvanted CoaB-CRM Vaccine-Enhanced CoaB-Specific Antibody Titers
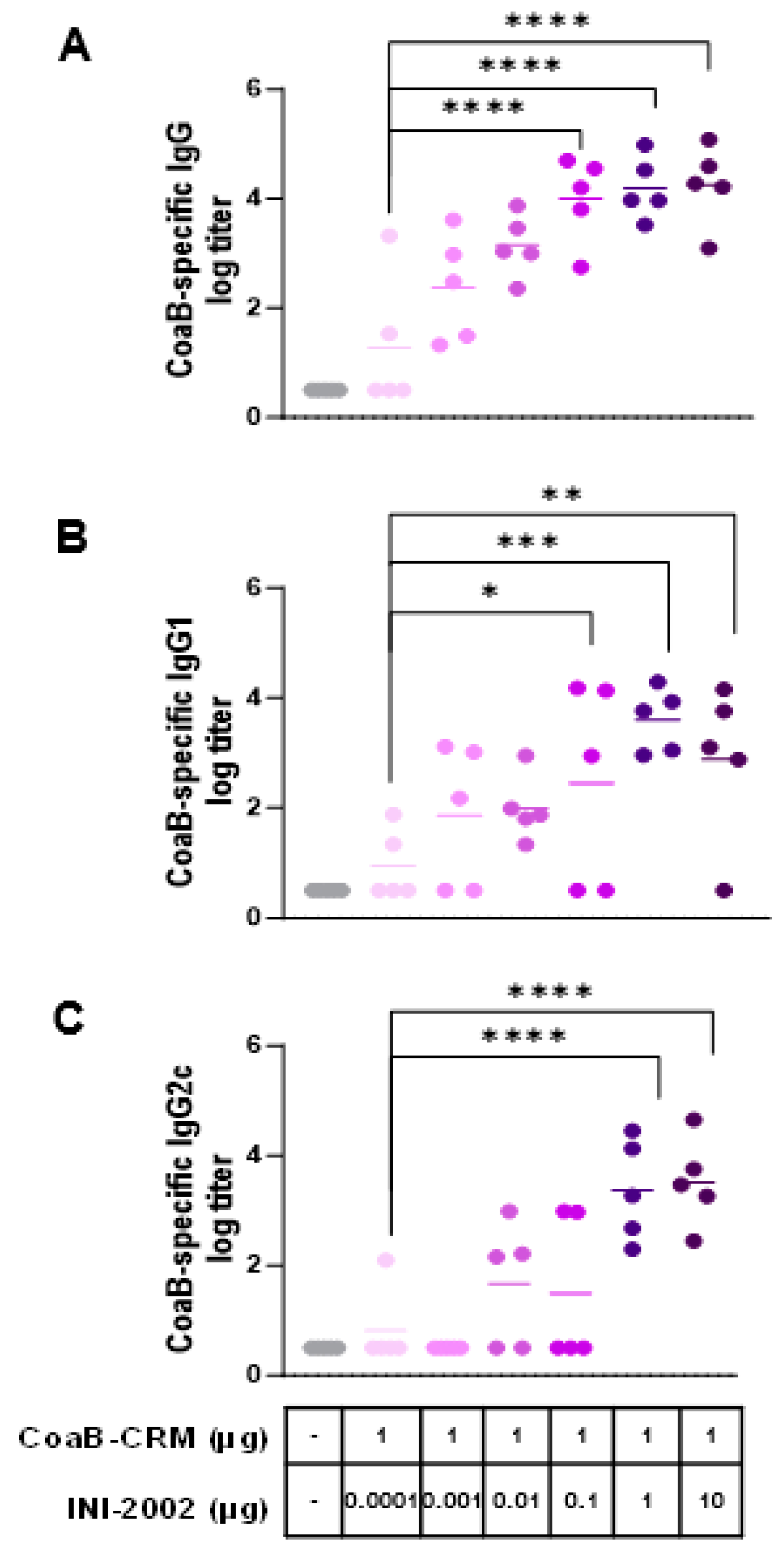
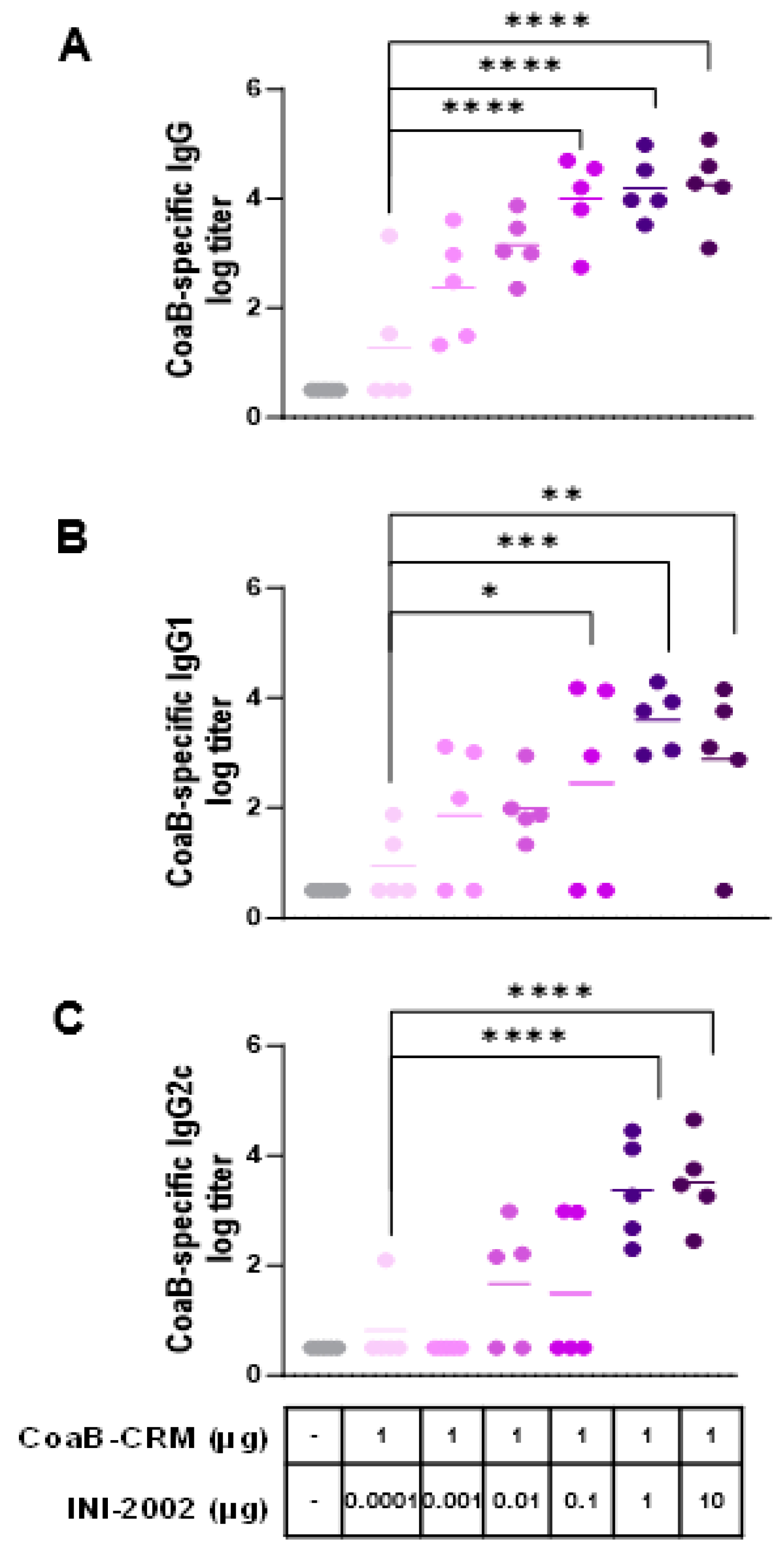
3.3. INI-2002 Enhances Humoral Immunity to CoaB Peptide in a Dose-Dependent Fashion
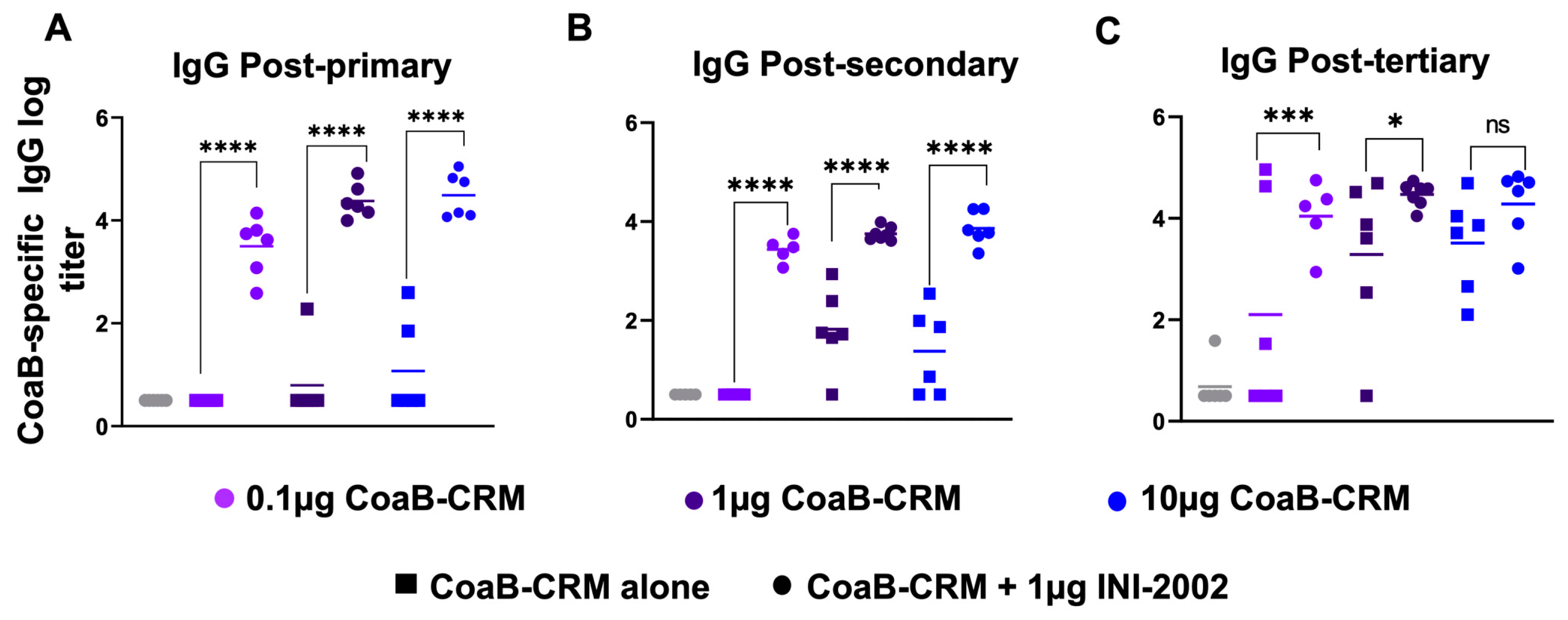
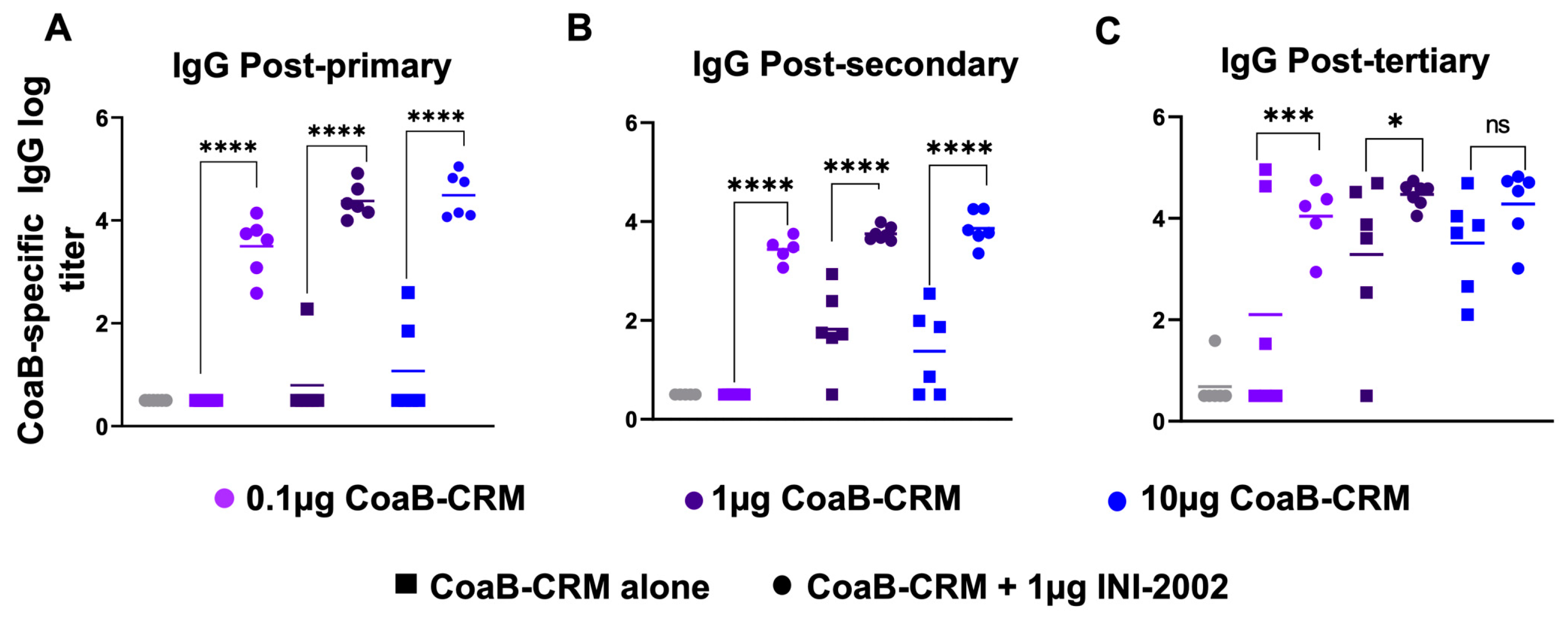
3.4. INI-2002 Adjuvant Enhances Humoral Immunity across the Antigen Dose Range and Is Antigen-Dose Sparing
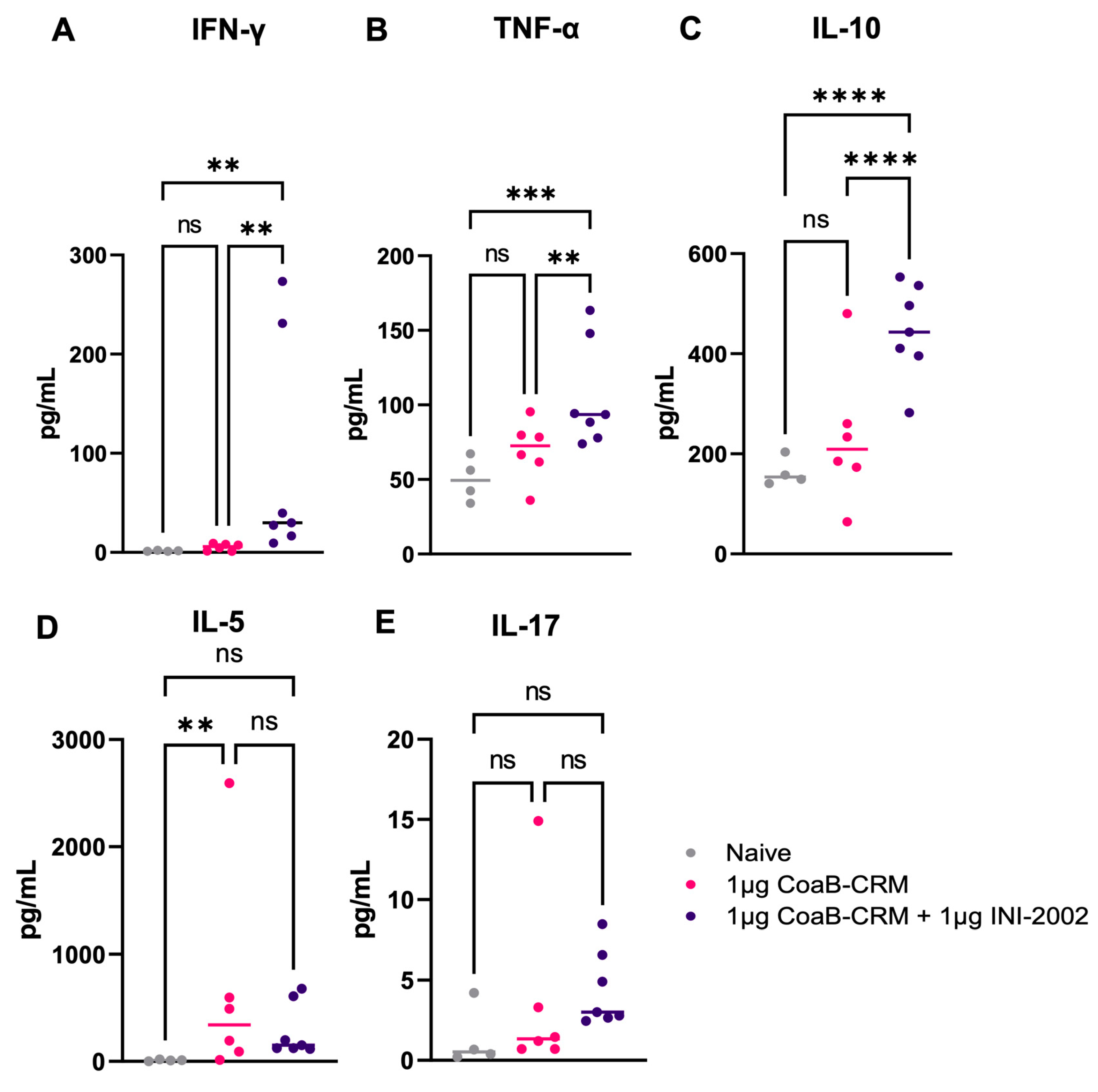
3.5. INI-2002 Induces a Balanced T-Helper Cell Response with CRM
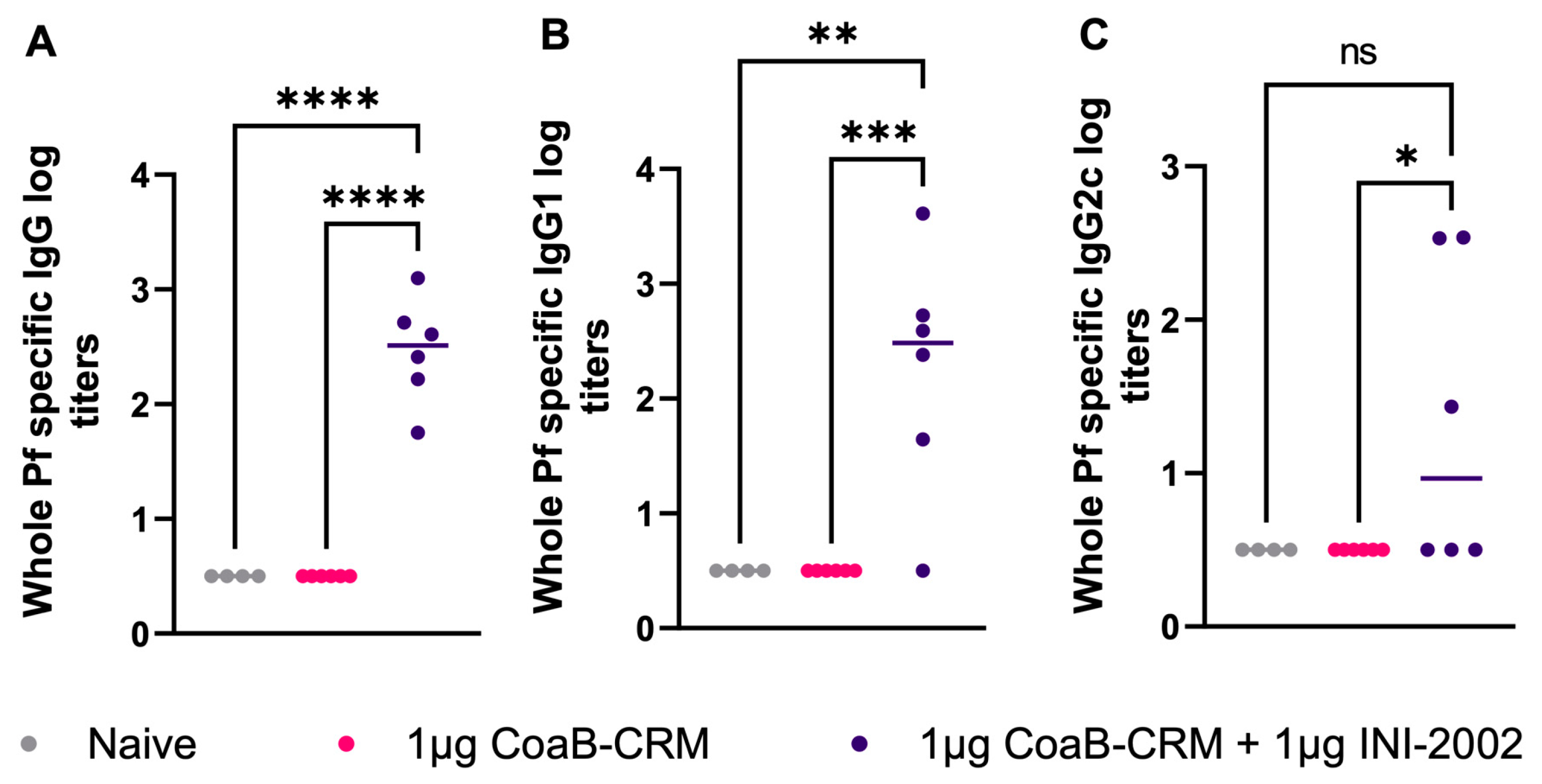
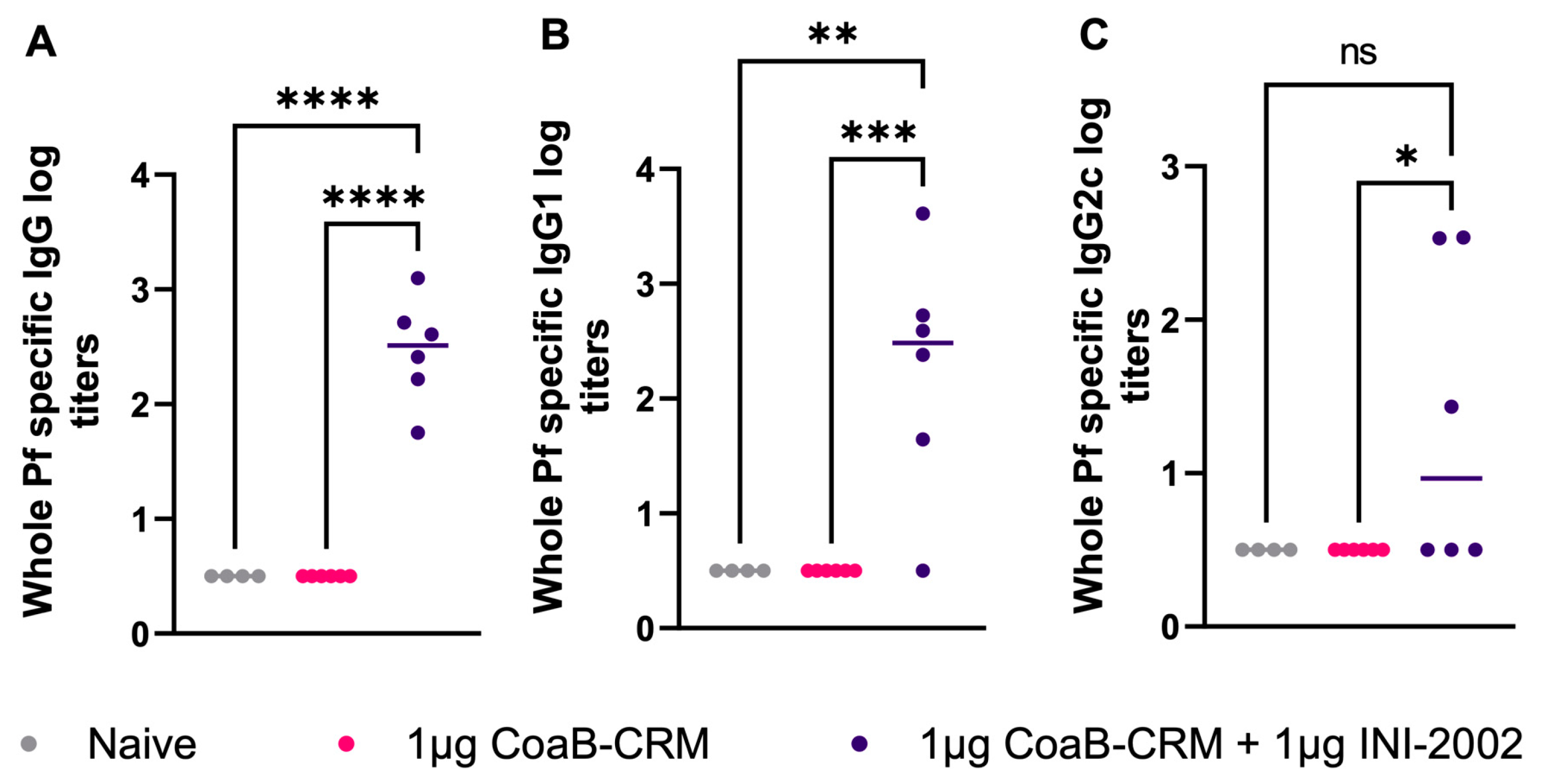
3.6. Antibodies Generated from CoaB-CRM-Vaccinated Animals Recognize Native Pf4 Virions
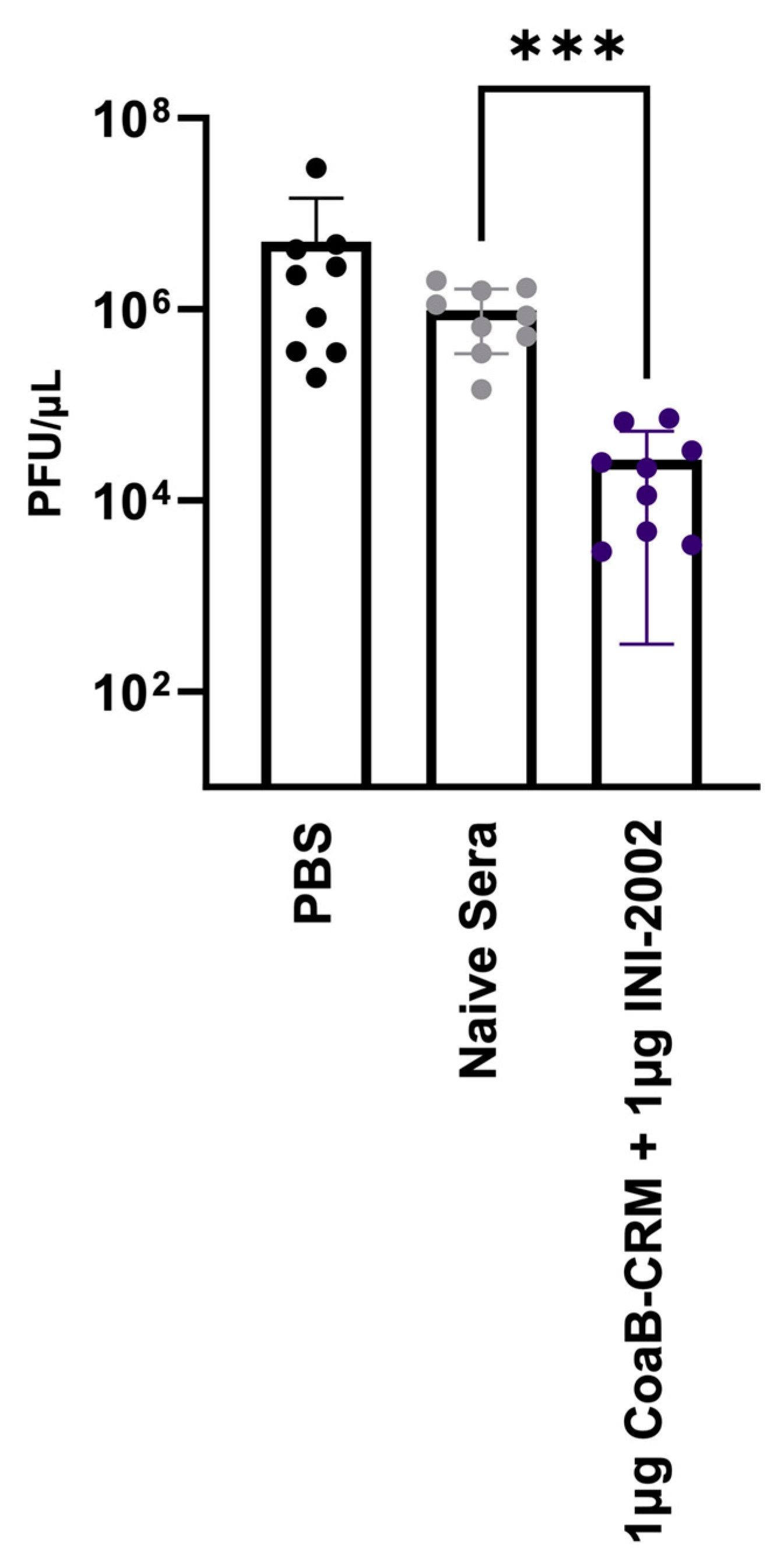
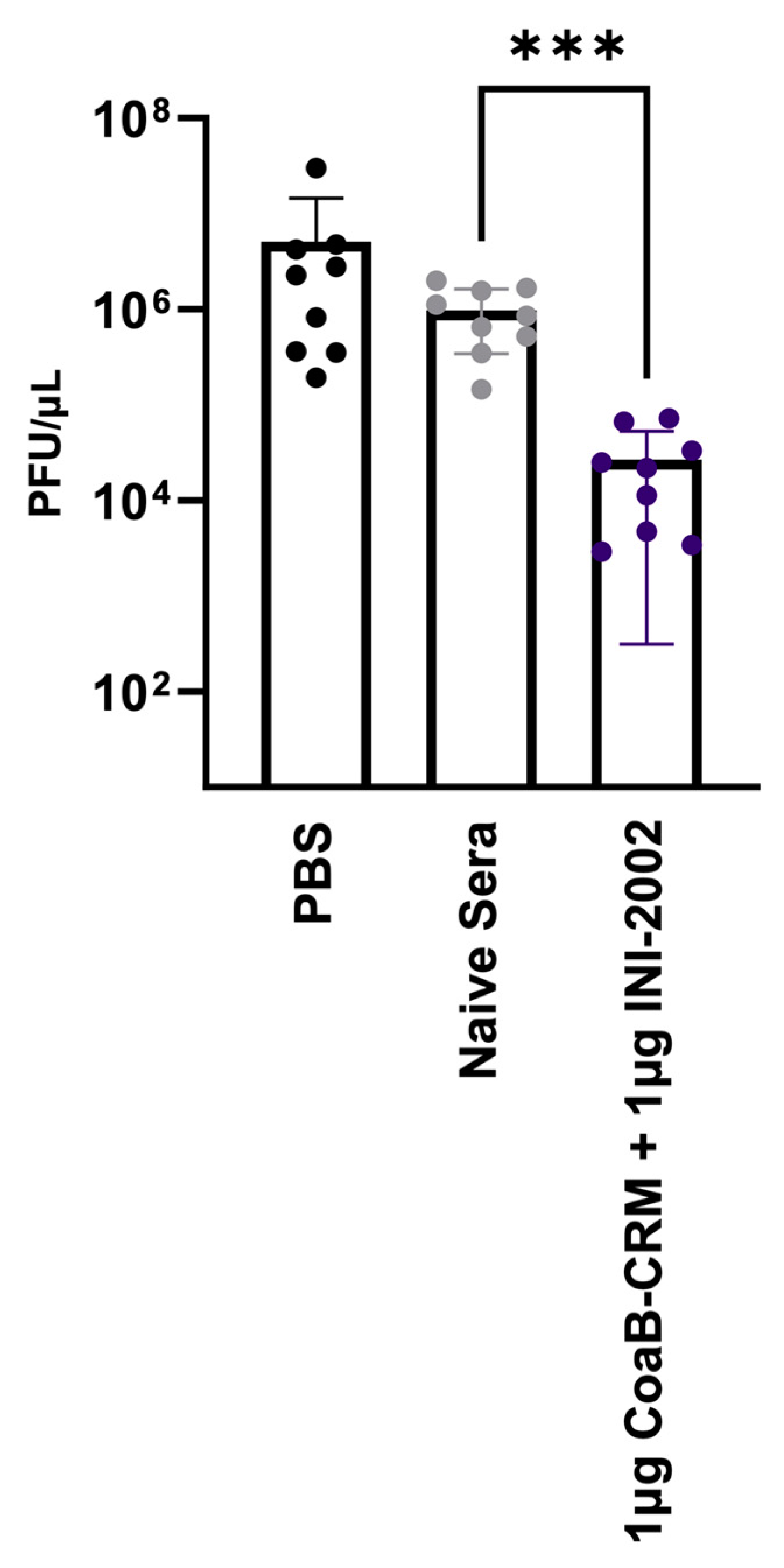
3.7. Anti-Pf4 Antibodies Disrupt the Pf4 Replication Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Multiple Mechanisms of Antimicrobial Resistance in Pseudomonas aeruginosa: Our Worst Nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Goossens, H. Susceptibility of multi-drug-resistant Pseudomonas aeruginosa in intensive care units: Results from the European MYSTIC study group. Clin. Microbiol. Infect. 2003, 9, 980–983. [Google Scholar] [CrossRef]
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef]
- Sainz-Mejías, M.; Jurado-Martín, I.; McClean, S. Understanding Pseudomonas aeruginosa-Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells 2020, 9, 2617. [Google Scholar] [CrossRef]
- Sweere, J.M.; Van Belleghem, J.D.; Ishak, H.; Bach, M.S.; Popescu, M.; Sunkari, V.; Kaber, G.; Manasherob, R.; Suh, G.A.; Cao, X.; et al. Bacteriophage trigger antiviral immunity and prevent clearance of bacterial infection. Science 2019, 363, eaat9691. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Jensen, P.Ø.; Fiandaca, M.J.; Pedersen, J.; Hansen, C.R.; Andersen, C.B.; Pressler, T.; Givskov, M.; Høiby, N. Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatr. Pulmonol. 2009, 44, 547–558. [Google Scholar] [CrossRef]
- Secor, P.R.; Burgener, E.B.; Kinnersley, M.; Jennings, L.K.; Roman-Cruz, V.; Popescu, M.; Van Belleghem, J.D.; Haddock, N.; Copeland, C.; Michaels, L.A.; et al. Pf Bacteriophage and Their Impact on Pseudomonas Virulence, Mammalian Immunity, and Chronic Infections. Front. Immunol. 2020, 11, 244. [Google Scholar] [CrossRef]
- Knezevic, P.; Voet, M.; Lavigne, R. Prevalence of Pf1-like (pro) phage genetic elements among Pseudomonas aeruginosa isolates $. Virology 2015, 483, 64–71. [Google Scholar] [CrossRef]
- Tarafder, A.K.; von Kügelgen, A.; Mellul, A.J.; Schulze, U.; Aarts, D.G.; Bharat, T.A. Phage liquid crystalline droplets form occlusive sheaths that encapsulate and protect infectious rod-shaped bacteria. Proc. Natl. Acad. Sci. USA 2020, 117, 4724–4731. [Google Scholar] [CrossRef]
- Coffman, R.L.; Sher, A.; Seder, R.A. NIH Public Access Vaccine Adjuvants: Putting Innate Immunity to Work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, J.K.; Bess, L.S.; Walsh, L.M.; Ward, J.M.; Johnson, C.L.; Livesay, M.T.; Jackson, K.J.; Evans, J.T.; Ryter, K.T.; Bazin-Lee, H.G. Diamino Allose Phosphates: Novel, Potent, and Highly Stable Toll-like Receptor 4 Agonists. J. Med. Chem. 2023, 66, 13900–13917. [Google Scholar] [CrossRef] [PubMed]
- Crouse, B.; Miller, S.M.; Muelken, P.; Hicks, L.; Vigliaturo, J.R.; Marker, C.L.; Guedes, A.G.; Pentel, P.R.; Evans, J.T.; LeSage, M.G.; et al. A TLR7/8 agonist increases efficacy of anti-fentanyl vaccines in rodent and porcine models. NPJ Vaccines 2023, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Holloway, B.W.; Krishnapillai, V.; Morgan, A.F. Chromosomal Genetics of Pseudomonas. 1979. Available online: https://journals.asm.org/journal/mr (accessed on 1 June 2020).
- Rice, S.A.; Tan, C.H.; Mikkelsen, P.J.; Kung, V.; Woo, J.; Tay, M.; Hauser, A.; McDougald, D.; Webb, J.S.; Kjelleberg, S. The biofilm life cycle and virulence of Pseudomonas aeruginosa are dependent on a filamentous prophage. ISME J. 2009, 3, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.K.; Fitzpatrick, A.D.; Schwartzkopf, C.M.; Faith, D.R.; Jennings, L.K.; Coluccio, A.; Hunt, D.J.; Michaels, L.A.; Hargil, A.; Chen, Q.; et al. A Filamentous Bacteriophage Protein Inhibits Type IV Pili To Prevent Superinfection of Pseudomonas aeruginosa. mBio 2022, 13, e02441-21. [Google Scholar] [CrossRef]
- FDA. Pneumococcal 13-Valent Conjugate Vaccine (Diphtheria CRM197 Protein) Package Insert. 2022. Available online: http://www.fda.gov/BiologicsBloodVaccines/Vaccines/ApprovedProducts/ucm201667.htm (accessed on 10 April 2023).
- FDA. Meningococcal (Groups A, C Y and W-135) Oligosaccharide Diphtheria CRM197 Conjugate Vaccine Package Insert. 2022. Available online: http://www.fda.gov/BiologicsBloodVaccines/Vaccines/ApprovedProducts/ucm201342.htm (accessed on 10 April 2023).
- Bröker, M.; Costantino, P.; DeTora, L.; McIntosh, E.D.; Rappuoli, R. Biochemical and biological characteristics of cross-reacting material 197 (CRM197), a non-toxic mutant of diphtheria toxin: Use as a conjugation protein in vaccines and other potential clinical applications. Biologicals 2011, 39, 195–204. [Google Scholar] [CrossRef]
- Faure, K.; Sawa, T.; Ajayi, T.; Fujimoto, J.; Moriyama, K.; Shime, N.; Wiener-Kronish, J.P. TLR4 signaling is essential for survival in acute lung injury induced by virulent Pseudomas aeruginosa secreting type III secretory toxins. Respir. Res. 2004, 5, 1. [Google Scholar] [CrossRef]
- Hernandez, A.; Patil, N.K.; Stothers, C.L.; Luan, L.; McBride, M.A.; Owen, A.M.; Burelbach, K.R.; Williams, D.L.; Sherwood, E.R.; Bohannon, J.K. Immunobiology and application of toll-like receptor 4 agonists to augment host resistance to infection. Pharmacol. Res. 2019, 150, 104502. [Google Scholar] [CrossRef]
- Snapper, C.M.; Paul, W.E. Interferon-gamma and B Cell Stimulatory Factor-1 Reciprocally Regulate Ig Isotype Production. Science 1987, 236, 944–947. [Google Scholar] [CrossRef]
- Zuercher, A.W.; Horn, M.P.; Wu, H.; Song, Z.; Bundgaard, C.J.; Johansen, H.K.; Høiby, N.; Marcus, P.; Lang, A.B. Intranasal immunisation with conjugate vaccine protects mice from systemic and respiratory tract infection with Pseudomonas aeruginosa. Vaccine 2006, 24, 4333–4342. [Google Scholar] [CrossRef] [PubMed]
- Döring, G. International Journal of Medical Microbiology Prevention of Pseudomonas aeruginosa infection in cystic fibrosis patients. Int. J. Med. Microbiol. 2010, 300, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Billeskov, R.; Lindenstrøm, T.; Woodworth, J.; Vilaplana, C.; Cardona, P.J.; Cassidy, J.P.; Mortensen, R.; Agger, E.M.; Andersen, P. High antigen dose is detrimental to post-exposure vaccine protection against tuberculosis. Front. Immunol. 2018, 8, 1973. [Google Scholar] [CrossRef] [PubMed]
- Birger, A. Studies on the regulation of avidity at the level of the single antibody-forming cell-The Effect of Antigen Dose and the Time After Immunization. J. Exp. Med. 1970, 132, 77–88. [Google Scholar] [CrossRef]
- Buret, A.; Dunkley, M.L.; Pang, G.; Clancy, R.L.; Cripps, A.W. Pulmonary immunity to Pseudomonas aeruginosa in intestinally immunized rats: Roles of alveolar macrophages, tumor necrosis factor alpha, and interleukin-1α. Infect. Immun. 1994, 62, 5335–5343. [Google Scholar] [CrossRef]
- Chmiel, J.F.; Konstan, M.W.; Knesebeck, J.E.; Hilliard, J.B.; Bonfield, T.L.; Dawson, D.V.; Berger, M. IL-10 attenuates excessive inflammation in chronic Pseudomonas infection in mice. Am. J. Respir. Crit. Care Med. 1999, 160, 2040–2047. [Google Scholar] [CrossRef]
- Sokol, P.A.; Kooi, C.; Hodges, R.S.; Cachia, P.; Woods, D.E. Immunization with a Pseudomonas aeruginosa elastase peptide reduces severity of experimental lung infections due to P. aeruginosa or Burkholderia cepacian. J. Infect. Dis. 2000, 181, 1682–1692. [Google Scholar] [CrossRef]
- Cachia, P.J.; Glasier, L.M.G.; Hodgins, R.R.W.; Wong, W.Y.; Irvin, R.T.; Hodges, R.S. The use of synthetic peptides in the design of a consensus sequence vaccine for Pseudomonas aeruginosa. J. Pept. Res. 1998, 52, 289–299. [Google Scholar] [CrossRef]
- Fiedoruk, K.; Zakrzewska, M.; Daniluk, T.; Piktel, E.; Chmielewska, S.; Bucki, R. Two lineages of Pseudomonas aeruginosa filamentous phages: Structural uniformity over integration preferences. Genome Biol. Evol. 2020, 12, 1765–1781. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Román-Cruz, V.C.; Miller, S.M.; Schoener, R.A.; Lukasiewicz, C.; Schmidt, A.K.; DeBuysscher, B.L.; Burkhart, D.; Secor, P.R.; Evans, J.T. Adjuvanted Vaccine Induces Functional Antibodies against Pseudomonas aeruginosa Filamentous Bacteriophages. Vaccines 2024, 12, 115. https://doi.org/10.3390/vaccines12020115
Román-Cruz VC, Miller SM, Schoener RA, Lukasiewicz C, Schmidt AK, DeBuysscher BL, Burkhart D, Secor PR, Evans JT. Adjuvanted Vaccine Induces Functional Antibodies against Pseudomonas aeruginosa Filamentous Bacteriophages. Vaccines. 2024; 12(2):115. https://doi.org/10.3390/vaccines12020115
Chicago/Turabian StyleRomán-Cruz, Valery C., Shannon M. Miller, Roman A. Schoener, Chase Lukasiewicz, Amelia K. Schmidt, Blair L. DeBuysscher, David Burkhart, Patrick R. Secor, and Jay T. Evans. 2024. "Adjuvanted Vaccine Induces Functional Antibodies against Pseudomonas aeruginosa Filamentous Bacteriophages" Vaccines 12, no. 2: 115. https://doi.org/10.3390/vaccines12020115
APA StyleRomán-Cruz, V. C., Miller, S. M., Schoener, R. A., Lukasiewicz, C., Schmidt, A. K., DeBuysscher, B. L., Burkhart, D., Secor, P. R., & Evans, J. T. (2024). Adjuvanted Vaccine Induces Functional Antibodies against Pseudomonas aeruginosa Filamentous Bacteriophages. Vaccines, 12(2), 115. https://doi.org/10.3390/vaccines12020115