

Intensified Influenza Virus Production in Suspension HEK293SF Cell Cultures Operated in Fed-Batch or Perfusion with Continuous Harvest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line and Culture Conditions
2.2. Virus Strains and Infection Conditions
2.3. Feeding Strategies and Process Operation
2.4. Small-Scale Experiments
2.5. Bioreactor Set-Up and Operation
2.6. Virus Quantification and Productivity Evaluation
3. Results
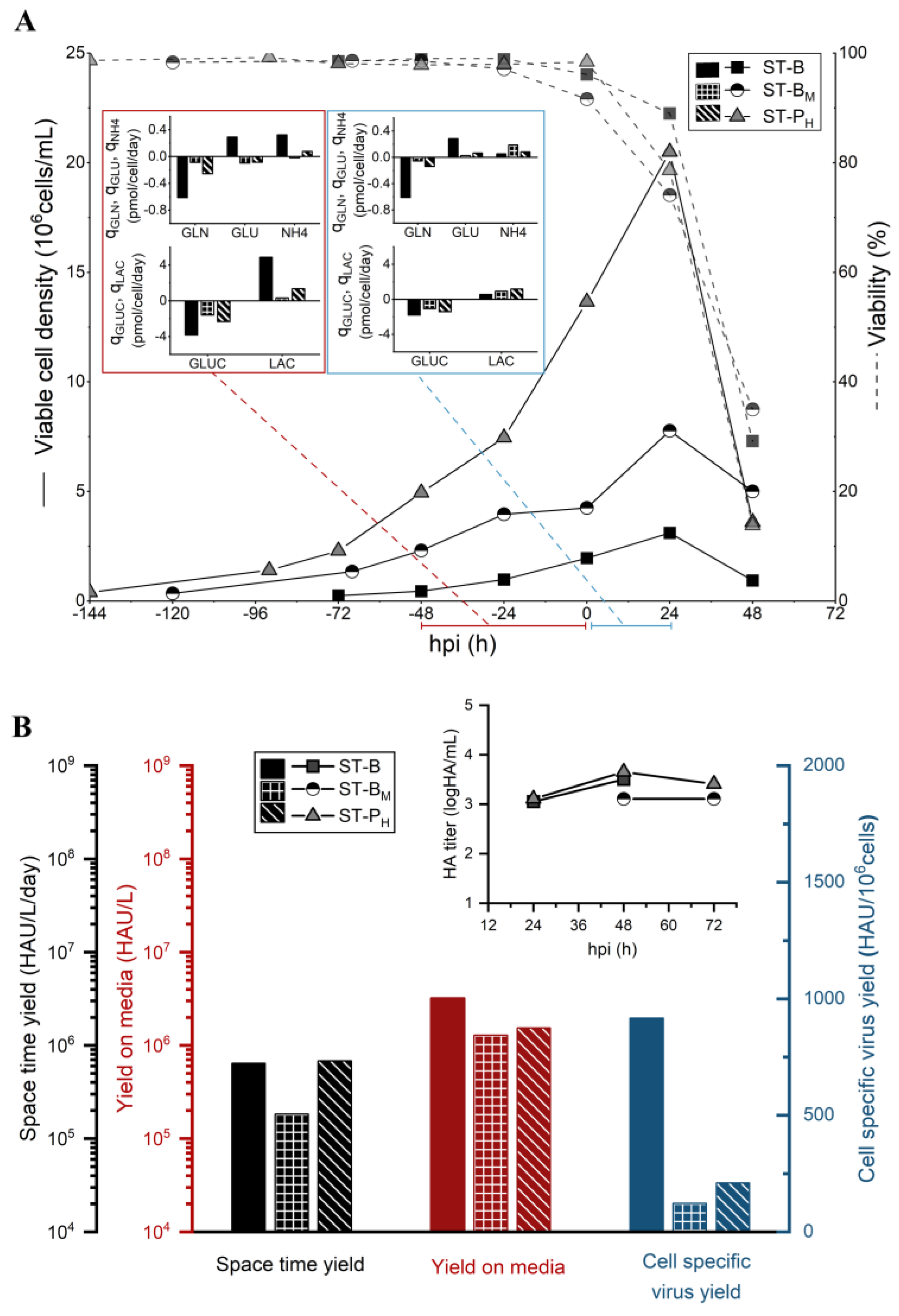
3.1. Small-Scale Characterization of the CDE
3.2. Small-Scale Development of HCD Feeding Strategies
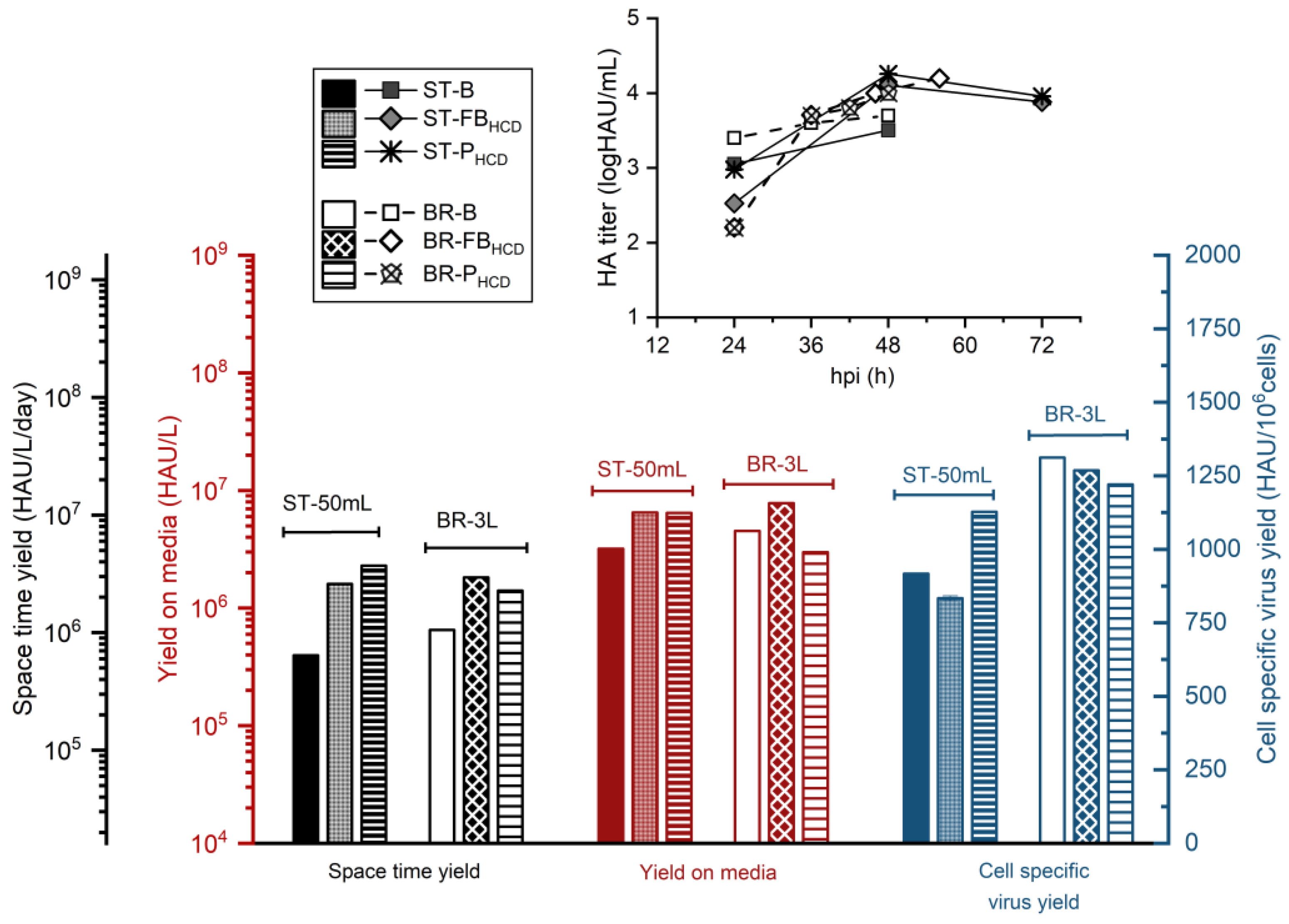
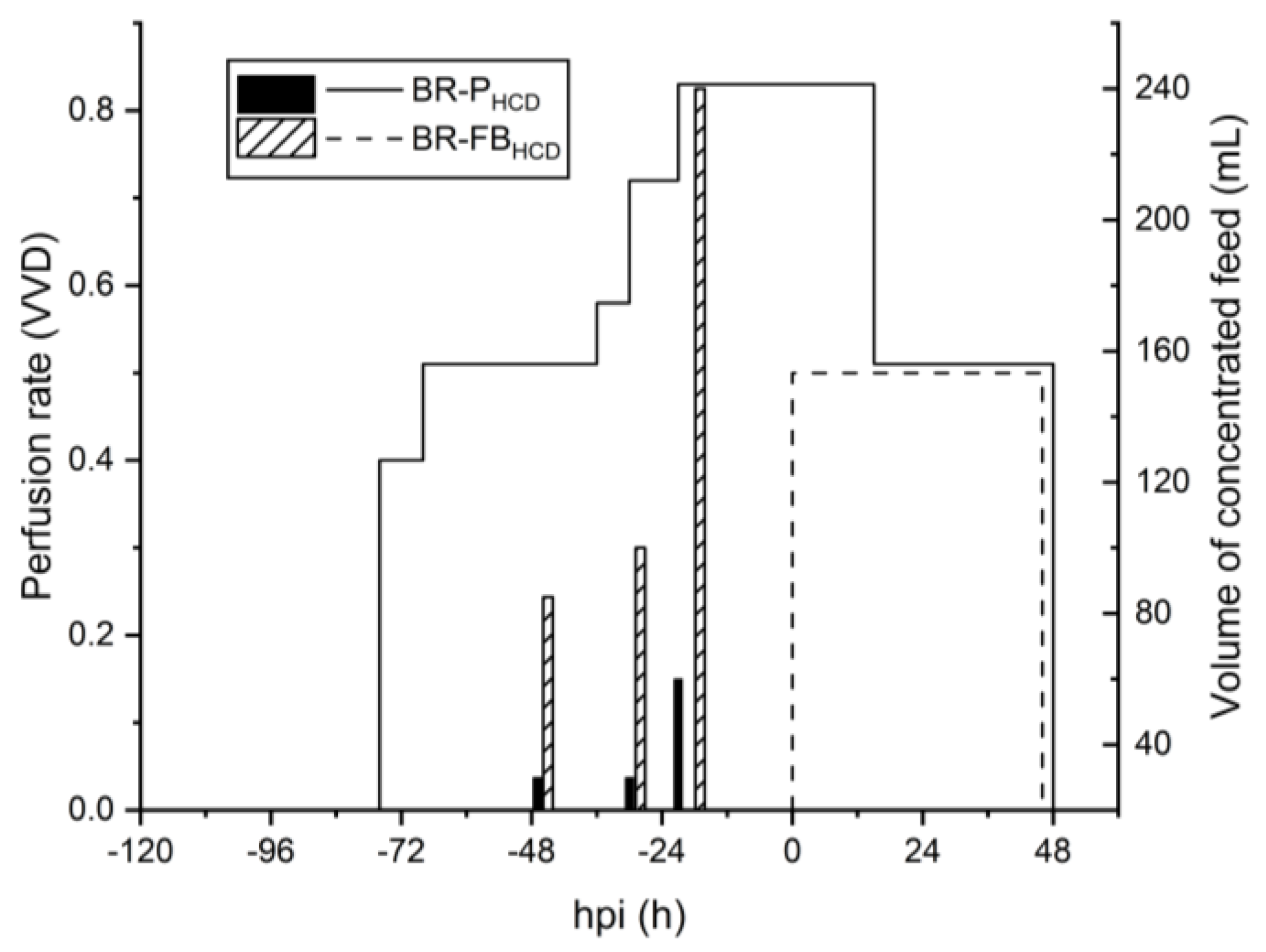
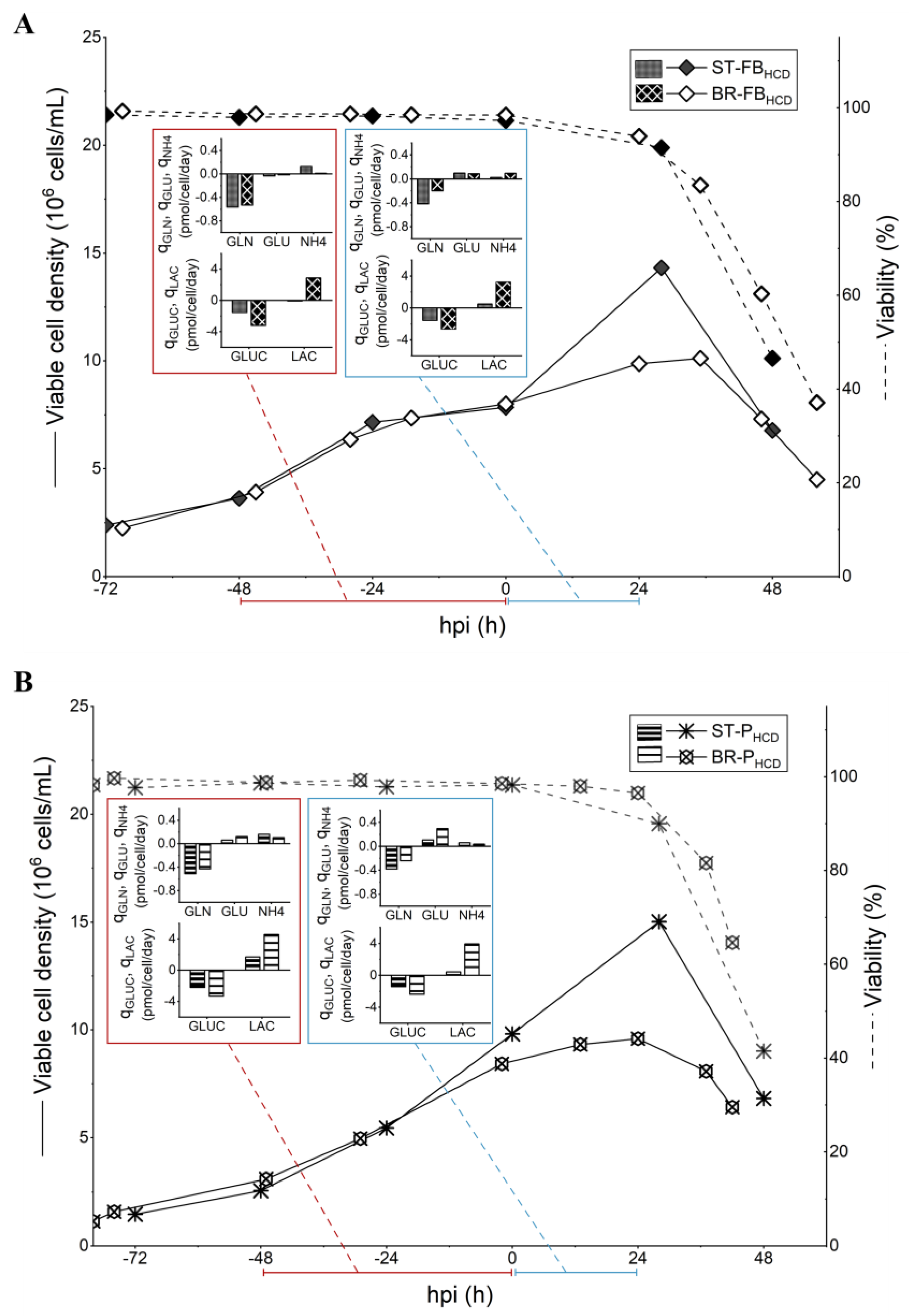
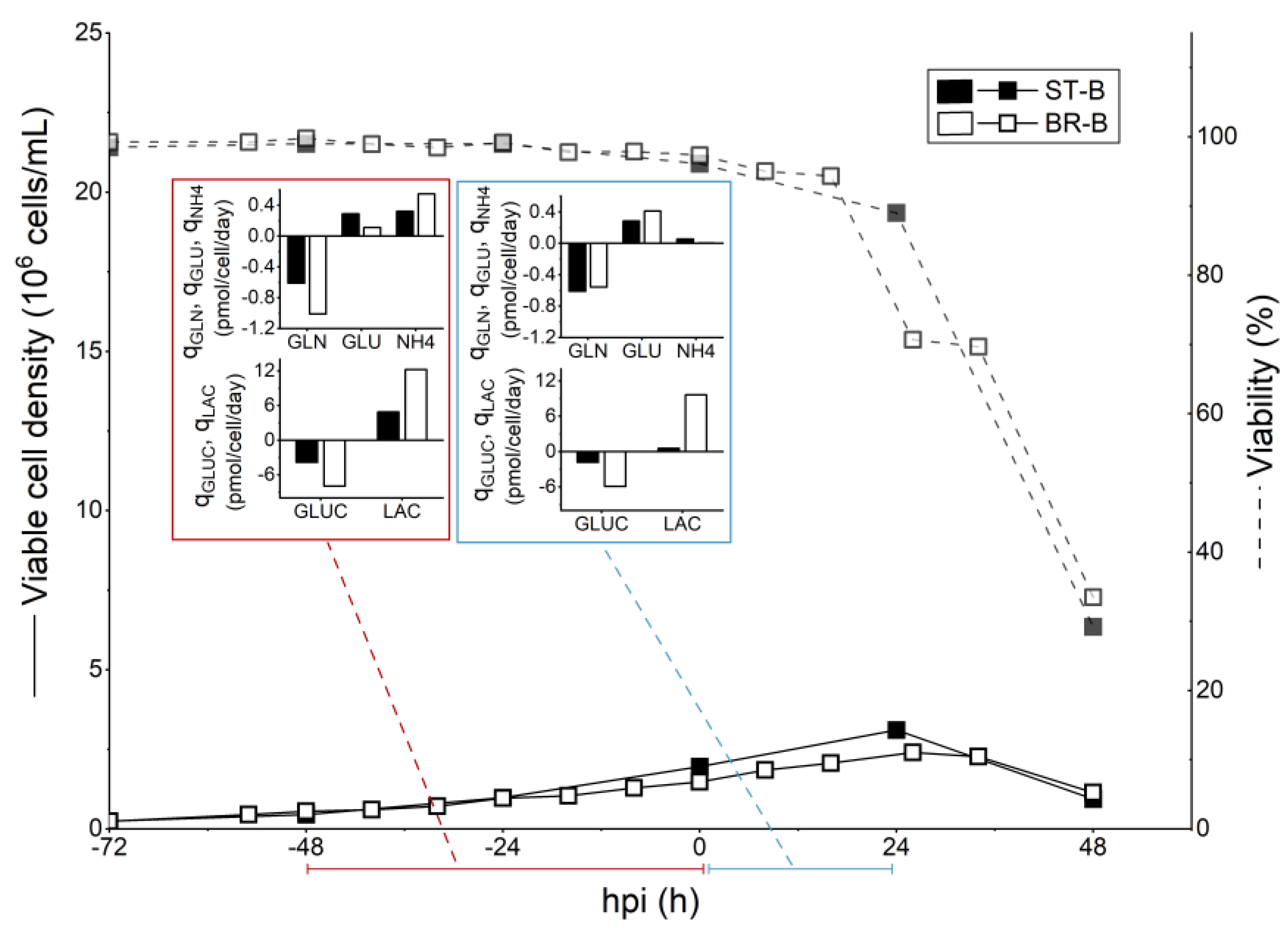
3.3. Scale Up to 3 L STR Bioreactor
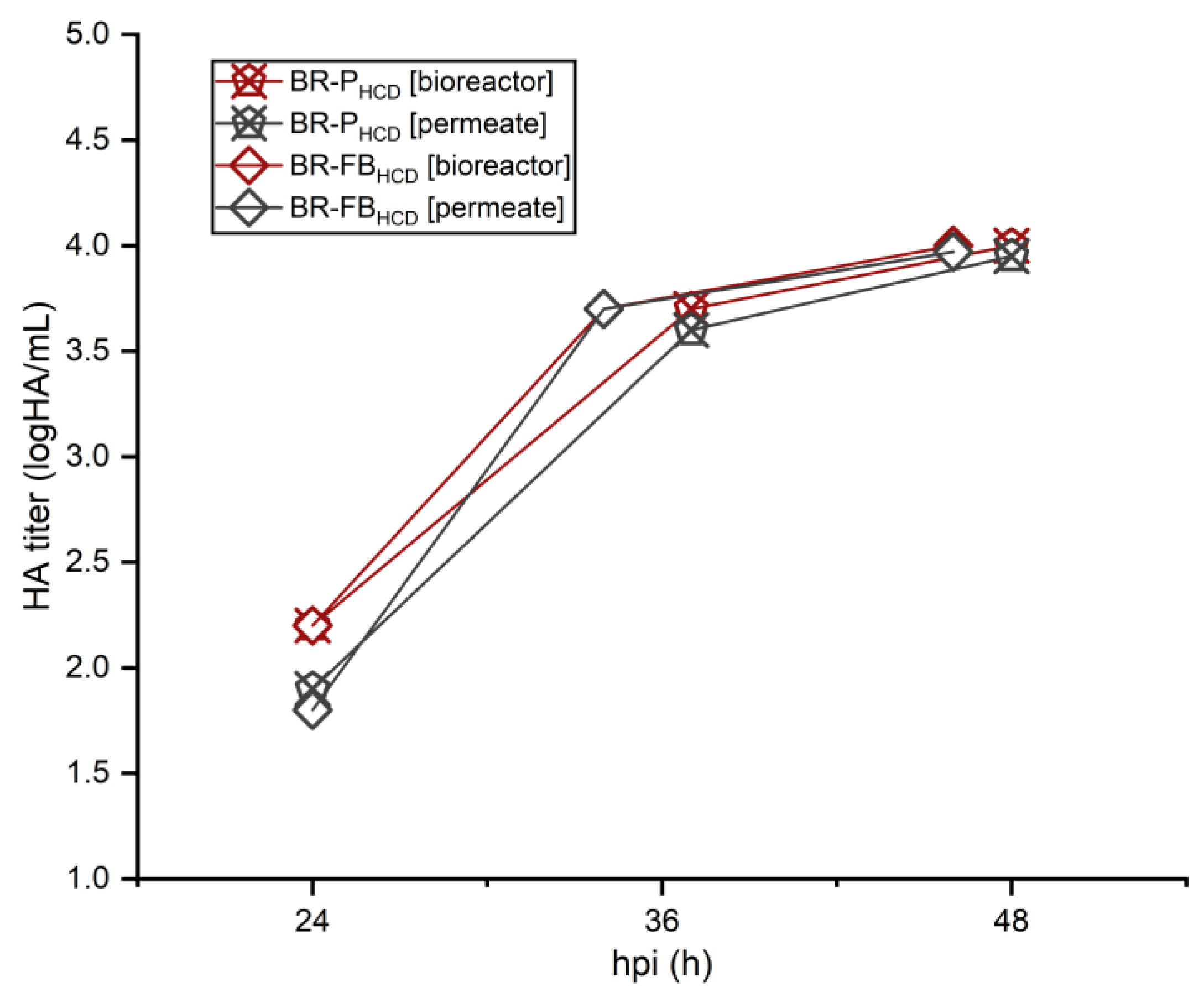
3.4. IAV Production and Harvest Using the TFDF System
4. Discussion
4.1. Small-Scale Characterization of the CDE
4.2. Small-Scale Development of HCD Feeding Strategies
4.3. Scale Up to 3 L STR Bioreactor
4.4. IAV Production and Harvest Using the TFDF System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, E.; Wood, J.G.; Chadwick, C.; Newall, A.T.; Torvaldsen, S.; Moen, A.; Torelli, G. Global production capacity of seasonal and pandemic influenza vaccines in 2019. Vaccine 2021, 39, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Kash, J.C. Influenza virus evolution, host adaptation, and pandemic formation. Cell Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.A.T.; Kamen, A.A.; Henry, O. Recent advances and current challenges in process intensification of cell culture-based influenza virus vaccine manufacturing. Can. J. Chem. Eng. 2021, 99, 2525–2535. [Google Scholar] [CrossRef]
- Pohlscheidt, M.; Jacobs, M.; Wolf, S.; Thiele, J.; Jockwer, A.; Gabelsberger, J.; Jenzsch, M.; Tebbe, H.; Burg, J. Optimizing capacity utilization by large scale 3000 L perfusion in seed train bioreactors. Biotechnol. Prog. 2013, 29, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Hardy, C.T.; Young, S.A.; Webster, R.G.; Naeve, C.W.; Owens, R.J. Egg fluids and cells of the chorioallantoic membrane of embryonated chicken eggs can select different variants of influenza A (H3N2) viruses. Virology 1995, 211, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Zost, S.J.; Parkhouse, K.; Gumina, M.E.; Kim, K.; Diaz Perez, S.; Wilson, P.C.; Treanor, J.J.; Sant, A.J.; Cobey, S.; Hensley, S.E. Contemporary H3N2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc. Natl. Acad. Sci. USA 2017, 114, 12578–12583. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shih, J.; Sinacore, M.; Ryll, T.; Yusuf-Makagiansar, H. Development and implementation of a perfusion-based high cell density cell banking process. Biotechnol. Prog. 2011, 27, 824–829. [Google Scholar] [CrossRef]
- Müller, J.; Ott, V.; Eibl, D.; Eibl, R. Seed Train Intensification Using an Ultra-High Cell Density Cell Banking Process. Processes 2022, 10, 911. [Google Scholar] [CrossRef]
- Frank, G.T. Transformation of biomanufacturing by single-use systems and technology. Curr. Opin. Chem. Eng. 2018, 22, 62–70. [Google Scholar] [CrossRef]
- Dumont, J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, H.; Zhang, Y.; Zhan, C.; Malm, M.; Field, R.; Turner, R.; Sellick, C.; Varley, P.; Rockberg, J.; Chotteau, V. Small-scale bioreactor supports high density HEK293 cell perfusion culture for the production of recombinant Erythropoietin. J. Biotechnol. 2020, 309, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Pulix, M.; Lukashchuk, V.; Smith, D.C.; Dickson, A.J. Molecular characterization of HEK293 cells as emerging versatile cell factories. Curr. Opin. Biotechnol. 2021, 71, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Manceur, A.P.; Kim, H.; Misic, V.; Andreev, N.; Dorion-Thibaudeau, J.; Lanthier, S.; Bernier, A.; Tremblay, S.; Gelinas, A.M.; Broussau, S.; et al. Scalable Lentiviral Vector Production Using Stable HEK293SF Producer Cell Lines. Hum. Gene Ther. Methods 2017, 28, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: Knowns and unknowns of antigen presentation and modes of action. npj Vaccines 2021, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Hein, M.D.; Kazenmaier, D.; van Heuvel, Y.; Dogra, T.; Cattaneo, M.; Kupke, S.Y.; Stitz, J.; Genzel, Y.; Reichl, U. Production of retroviral vectors in continuous high cell density culture. Appl. Microbiol. Biotechnol. 2023, 107, 5947–5961. [Google Scholar] [CrossRef] [PubMed]
- Henry, O.; Dormond, E.; Perrier, M.; Kamen, A. Insights into adenoviral vector production kinetics in acoustic filter-based perfusion cultures. Biotechnol. Bioeng. 2004, 86, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Henry, O.; Perrier, M.; Kamen, A. Metabolic flux analysis of HEK-293 cells in perfusion cultures for the production of adenoviral vectors. Metab. Eng. 2005, 7, 467–476. [Google Scholar] [CrossRef]
- Bock, A.; Schulze-Horsel, J.; Schwarzer, J.; Rapp, E.; Genzel, Y.; Reichl, U. High-density microcarrier cell cultures for influenza virus production. Biotechnol. Prog. 2011, 27, 241–250. [Google Scholar] [CrossRef]
- Petiot, E.; Jacob, D.; Lanthier, S.; Lohr, V.; Ansorge, S.; Kamen, A.A. Metabolic and kinetic analyses of influenza production in perfusion HEK293 cell culture. BMC Biotechnol. 2011, 11, 84. [Google Scholar] [CrossRef]
- Xu, S.; Gavin, J.; Jiang, R.; Chen, H. Bioreactor productivity and media cost comparison for different intensified cell culture processes. Biotechnol. Prog. 2017, 33, 867–878. [Google Scholar] [CrossRef]
- Bausch, M.; Schultheiss, C.; Sieck, J.B. Recommendations for Comparison of Productivity Between Fed-Batch and Perfusion Processes. Biotechnol. J. 2019, 14, e1700721. [Google Scholar] [CrossRef]
- Tapia, F.; Vazquez-Ramirez, D.; Genzel, Y.; Reichl, U. Bioreactors for high cell density and continuous multi-stage cultivations: Options for process intensification in cell culture-based viral vaccine production. Appl. Microbiol. Biotechnol. 2016, 100, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Konakovsky, V.; Clemens, C.; Müller, M.; Bechmann, J.; Berger, M.; Schlatter, S.; Herwig, C. Metabolic Control in Mammalian Fed-Batch Cell Cultures for Reduced Lactic Acid Accumulation and Improved Process Robustness. Bioengineering 2016, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.; Hiller, G.; Luan, Y.T.; Kittredge, A.; DeFelice, J.; Drapeau, D. High-end pH-controlled delivery of glucose effectively suppresses lactate accumulation in CHO fed-batch cultures. Biotechnol. Bioeng. 2011, 108, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Hiller, G.W.; Ovalle, A.M.; Gagnon, M.P.; Curran, M.L.; Wang, W. Cell-controlled hybrid perfusion fed-batch CHO cell process provides significant productivity improvement over conventional fed-batch cultures. Biotechnol. Bioeng. 2017, 114, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- Genzel, Y.; Vogel, T.; Buck, J.; Behrendt, I.; Ramirez, D.V.; Schiedner, G.; Jordan, I.; Reichl, U. High cell density cultivations by alternating tangential flow (ATF) perfusion for influenza A virus production using suspension cells. Vaccine 2014, 32, 2770–2781. [Google Scholar] [CrossRef]
- Vazquez-Ramirez, D.; Jordan, I.; Sandig, V.; Genzel, Y.; Reichl, U. High titer MVA and influenza A virus production using a hybrid fed-batch/perfusion strategy with an ATF system. Appl. Microbiol. Biotechnol. 2019, 103, 3025–3035. [Google Scholar] [CrossRef] [PubMed]
- Coronel, J.; Granicher, G.; Sandig, V.; Noll, T.; Genzel, Y.; Reichl, U. Application of an Inclined Settler for Cell Culture-Based Influenza A Virus Production in Perfusion Mode. Front. Bioeng. Biotechnol. 2020, 8, 672. [Google Scholar] [CrossRef]
- Granicher, G.; Coronel, J.; Trampler, F.; Jordan, I.; Genzel, Y.; Reichl, U. Performance of an acoustic settler versus a hollow fiber-based ATF technology for influenza virus production in perfusion. Appl. Microbiol. Biotechnol. 2020, 104, 4877–4888. [Google Scholar] [CrossRef]
- Padawer, I.; Ling, W.L.; Bai, Y. Case Study: An accelerated 8-day monoclonal antibody production process based on high seeding densities. Biotechnol. Prog. 2013, 29, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Lu, J.; Kwiatkowski, C.; Yuan, H.; Kshirsagar, R.; Ryll, T.; Huang, Y.M. Perfusion seed cultures improve biopharmaceutical fed-batch production capacity and product quality. Biotechnol. Prog. 2014, 30, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Karst, D.J.; Steinebach, F.; Soos, M.; Morbidelli, M. Process performance and product quality in an integrated continuous antibody production process. Biotechnol. Bioeng. 2017, 114, 298–307. [Google Scholar] [CrossRef]
- Nikolay, A.; Leon, A.; Schwamborn, K.; Genzel, Y.; Reichl, U. Process intensification of EB66(R) cell cultivations leads to high-yield yellow fever and Zika virus production. Appl. Microbiol. Biotechnol. 2018, 102, 8725–8737. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bissinger, T.; Genzel, Y.; Liu, X.; Reichl, U.; Tan, W.S. High cell density perfusion process for high yield of influenza A virus production using MDCK suspension cells. Appl. Microbiol. Biotechnol. 2021, 105, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Hein, M.D.; Chawla, A.; Cattaneo, M.; Kupke, S.Y.; Genzel, Y.; Reichl, U. Cell culture-based production of defective interfering influenza A virus particles in perfusion mode using an alternating tangential flow filtration system. Appl. Microbiol. Biotechnol. 2021, 105, 7251–7264. [Google Scholar] [CrossRef] [PubMed]
- Hadpe, S.R.; Sharma, A.K.; Mohite, V.V.; Rathore, A.S. ATF for cell culture harvest clarification: Mechanistic modelling and comparison with TFF. J. Chem. Technol. Biotechnol. 2017, 92, 732–740. [Google Scholar] [CrossRef]
- Nikolay, A.; de Grooth, J.; Genzel, Y.; Wood, J.A.; Reichl, U. Virus harvesting in perfusion culture: Choosing the right type of hollow fiber membrane. Biotechnol. Bioeng. 2020, 117, 3040–3052. [Google Scholar] [CrossRef]
- Tran, M.Y.; Kamen, A.A. Production of Lentiviral Vectors Using a HEK-293 Producer Cell Line and Advanced Perfusion Processing. Front. Bioeng. Biotechnol. 2022, 10, 887716. [Google Scholar] [CrossRef]
- Mendes, J.P.; Fernandes, B.; Pineda, E.; Kudugunti, S.; Bransby, M.; Gantier, R.; Peixoto, C.; Alves, P.M.; Roldao, A.; Silva, R.J.S. AAV process intensification by perfusion bioreaction and integrated clarification. Front. Bioeng. Biotechnol. 2022, 10, 1020174. [Google Scholar] [CrossRef]
- Tona, R.M.; Shah, R.; Middaugh, K.; Steve, J.; Marques, J.; Roszell, B.R.; Jung, C. Process intensification for lentiviral vector manufacturing using tangential flow depth filtration. Mol. Ther. Methods Clin. Dev. 2023, 29, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Le Ru, A.; Jacob, D.; Transfiguracion, J.; Ansorge, S.; Henry, O.; Kamen, A.A. Scalable production of influenza virus in HEK-293 cells for efficient vaccine manufacturing. Vaccine 2010, 28, 3661–3671. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID(50) for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Sanfeliu, A.; Paredes, C.; Cairó, J.J.; Gòdia, F. Identification of key patterns in the metabolism of hybridoma cells in culture. Enzym. Microb. Technol. 1997, 21, 421–428. [Google Scholar] [CrossRef]
- Zupke, C.; Sinskey, A.J.; Stephanopoulos, G. Intracellular flux analysis applied to the effect of dissolved oxygen on hybridomas. Appl. Microbiol. Biotechnol. 1995, 44, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Mulukutla, B.C.; Gramer, M.; Hu, W.S. On metabolic shift to lactate consumption in fed-batch culture of mammalian cells. Metab. Eng. 2012, 14, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Cortin, V.; Thibault, J.; Jacob, D.; Garnier, A. High-titer adenovirus vector production in 293S cell perfusion culture. Biotechnol. Prog. 2004, 20, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Bissinger, T.; Fritsch, J.; Mihut, A.; Wu, Y.; Liu, X.; Genzel, Y.; Tan, W.S.; Reichl, U. Semi-perfusion cultures of suspension MDCK cells enable high cell concentrations and efficient influenza A virus production. Vaccine 2019, 37, 7003–7010. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, X.; Silva, C.A.T.; Farnos, O.; Venereo-Sanchez, A.; Toussaint, C.; Dash, S.; Gonzalez-Dominguez, I.; Bernier, A.; Henry, O.; et al. Membrane Chromatography-Based Downstream Processing for Cell-Culture Produced Influenza Vaccines. Vaccines 2022, 10, 1310. [Google Scholar] [CrossRef]
- Repligen Corporation. TFDF Technology. Available online: https://www.repligen.com/technologies/tfdf/tfdf-technology (accessed on 22 February 2023).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run Code | ST-B | ST-BM | ST-PH | ST-FBHCD | ST-PHCD | BR-B | BR-FBHCD | BR-PHCD |
|---|---|---|---|---|---|---|---|---|
| VR (mL) | 10 | 10 | 10 | 10 | 10 | 1800 | 2000 | 2000 |
| Growth phase | Batch | Batch | Perfusion c (0.5 VVD) | Fed-batch | Perfusion c (0.5 VVD) | Batch | Fed-batch | Perfusion (0.4–0.8 VVD) |
| Infection phase | Batch | Batch | Perfusion c (0.5 VVD) | Perfusion c (0.5 VVD) | Perfusion c (0.5 VVD) | Batch | Perfusion (0.5 VVD) | Perfusion (0.5 VVD) |
| VCD0 (106 cells/mL) | 0.35 | 0.35 | 0.35 | 2 | 1.5 | 0.5 | 2 | 1.5 |
| VCDTOI (106 cells/mL) | 2 | 4 | 13 | 8–10 | 8–10 | 1.6 | 8.5 | 8.6 |
| VCDMAX (106 cells/mL) a | 3.2 ± 0.16 | 8.1 ± 0.4 | 20.5 ± 2.16 | 14.3 ± 0.08 | 15.0 ± 0.05 | 2.4 ± 0.16 | 10.1 ± 0.42 | 10 ± 0.5 |
| HATOH (logHAU/mL) | 3.5 | 3.1 | 3.65 | 4.11 | 4.26 | 3.7 | 4.2 | 4 |
| TCIDTOH (108 IVP/mL) a | 0.86 ± 0.15 | - | - | 5.28 ± 0.07 | 6.75 ± 1.5 | 0.53 ± 0.1 | 1.01 | 1.14 ± 0.21 |
| HATot (HAU) b | (3.20 ± 0.2) × 104 | (1.28 ± 0.3) × 104 | (5.44 ± 0.1) × 104 | (1.30 ± 0.01) × 105 | (1.86 ± 0.01) × 105 | 8.1 × 106 | 3.4 × 107 | 2.4 × 107 |
| Y/M (106 HAU/L) b | 3.20 ± 0.06 | 1.28 ± 0.2 | 1.53 ± 0.08 | 6.51 ± 0.04 | 6.45 ± 0.06 | 4.53 | 7.81 | 2.98 |
| CSVY (HAU/106 cell) b | 917 ± 12 | 122 ± 17 | 209 ± 35 | 834 ± 2 | 1129 ± 7 | 1311 | 1268 | 1220 |
| CSVYTCID (IVP/cell) b | 24.7 ± 4 | - | - | - | - | 15 | - | 14 |
| STY (106 HAU/L/day) b | 0.64 ± 0.01 | 0.18 ± 0.01 | 0.68 ± 0.01 | 2.60 ± 0.03 | 3.72 ± 0.02 | 1.1 | 3 | 2.3 |
| Total media | 1 × VR | 1 × VR | 4 × VR | 2 × VR | 2.9 × VR | 1 × VR | 2.2 × VR | 4 × VR |
| ST-B | ST-BM | ST-PH | ST-FBHCD | ST-PHCD | BR-B | BR-FBHCD | BR-PHCD | ||
|---|---|---|---|---|---|---|---|---|---|
| YLAC/GLUC | −48 hpi | 1.75 ± 0.4 | 0.2 ± 0.04 | 0.57± 0.002 | 0.06 ± 0.00 | 0.73 ± 0.02 | 1.58 | 0.78 | 1.29 |
| 24 hpi | 0.31 ± 0.04 | 0.87 ± 0.12 | 0.59 ± 0.15 | 0.32 ± 0.05 | 0.31 ± 0.01 | 1.64 | 1.24 | 1.46 | |
| YNH4/GLN | −48 hpi | 0.51 ± 0.05 | 0.07 ± 0.00 | 0.29 ± 0.1 | 0.22 ± 0.01 | 0.31 ± 0.001 | 0.53 | 0.01 | 0.2 |
| 24 hpi | 0.09 ± 0.03 | 2.97 ± 0.44 | 0.83 ± 0.01 | 0.06 ± 0.01 | 0.16 ± 0.001 | 0.06 | 0.47 | 0.12 |
| Volume (mL) | Virus Yield (logHAU) | Virus Yield (IVP) | ||||
|---|---|---|---|---|---|---|
| Bioreactor (TOH) | TFDF Harvest | Bioreactor (TOH) | TFDF Harvest | Bioreactor (TOH) | TFDF Harvest | |
| BR-B | 1720 | 1950 | 6.89 | 6.95 | 7.9 × 1010 | 5.66 × 1010 |
| BR-FBHCD | 2100 | 2510 | 7.48 | 7.49 | 2.1 × 1011 | 2.3 × 1011 |
| BR-PHCD | 1900 | 2150 | 7.29 | 7.18 | 2.2 × 1011 | 1.7 × 1011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, C.A.T.; Kamen, A.A.; Henry, O. Intensified Influenza Virus Production in Suspension HEK293SF Cell Cultures Operated in Fed-Batch or Perfusion with Continuous Harvest. Vaccines 2023, 11, 1819. https://doi.org/10.3390/vaccines11121819
Silva CAT, Kamen AA, Henry O. Intensified Influenza Virus Production in Suspension HEK293SF Cell Cultures Operated in Fed-Batch or Perfusion with Continuous Harvest. Vaccines. 2023; 11(12):1819. https://doi.org/10.3390/vaccines11121819
Chicago/Turabian StyleSilva, Cristina A. T., Amine A. Kamen, and Olivier Henry. 2023. "Intensified Influenza Virus Production in Suspension HEK293SF Cell Cultures Operated in Fed-Batch or Perfusion with Continuous Harvest" Vaccines 11, no. 12: 1819. https://doi.org/10.3390/vaccines11121819
APA StyleSilva, C. A. T., Kamen, A. A., & Henry, O. (2023). Intensified Influenza Virus Production in Suspension HEK293SF Cell Cultures Operated in Fed-Batch or Perfusion with Continuous Harvest. Vaccines, 11(12), 1819. https://doi.org/10.3390/vaccines11121819