Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Animals, Experimental Design and Treatments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Treatment | |||
|---|---|---|---|---|
| Control | FO | FO Plus SeE | SeE | |
| Ground corn | 77.00 | 76.95 | 76.93 | 76.98 |
| Soybean meal | 13.00 | 13.00 | 13.00 | 13.00 |
| Coastcross hay | 6.00 | 6.00 | 6.00 | 6.00 |
| Limestone 1 | 2.00 | 2.00 | 2.00 | 2.00 |
| Mineral premix 2 | 0.60 | 0.60 | 0.60 | 0.60 |
| Urea | 0.90 | 0.90 | 0.90 | 0.90 |
| Ammonium chloride | 0.50 | 0.50 | 0.50 | 0.50 |
| Functional oil 3 | 0.00 | 0.05 | 0.05 | 0.00 |
| Hydroxy-selenomethionine 4 | - | - | 0.0015 | 0.0015 |
| Vitamin E supplement 5 | - | - | 0.015 | 0.015 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutrients | ||||
| Metabolizable energy, Mcal/kg 6 | 2.94 | 2.94 | 2.94 | 2.94 |
| Crude protein, % | 17.40 | 17.39 | 17.39 | 17.40 |
| Rumen degradable protein, % | 10.21 | 10.20 | 10.20 | 10.20 |
| Neutral detergent fiber (NDF), % | 16.68 | 16.67 | 16.67 | 16.67 |
| NDF effective, % | 6.92 | 6.91 | 6.92 | 6.91 |
| Acid detergent fiber (ADF), % | 13.23 | 11.43 | 11.07 | 12.33 |
| N bound to NDF, % | 0.26 | 0.26 | 0.25 | 0.31 |
| N bound to ADF, % | 0.52 | 0.52 | 0.52 | 0.52 |
| Acid detergent lignin, % | 2.48 | 1.65 | 1.69 | 2.77 |
| Calcium, % | 0.87 | 0.87 | 0.87 | 0.87 |
| Phosphorus, % | 0.42 | 0.42 | 0.42 | 0.42 |
| Selenium, mg/kg | 0.13 | 0.15 | 0.45 | 0.49 |
2.3. Feed Intake and Chemical Analysis
2.4. Enzyme Activity and Blood Parameters
2.5. Slaughter, Sampling and Carcass Traits Assessment
2.6. Rumenitis and Rumen Morphology Analysis
2.7. Physical Analysis of Meat
2.8. Assessment of Meat Shelf Life Based on Lipid Peroxidation and Microbiological Quality
2.9. Statistical Analysis
3. Results
3.1. Performance, Carcass, Meat Quality Traits and Rumen Health
3.2. Oxidative Stress and Blood Parameters
3.3. Meat Shelf Life, Lipid Peroxidation (TBARS) and Microbiological Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asadollahi, S.; Sari, M.; Erafanimajd, N.; Kiani, A.; Ponnampalam, E.N. Supplementation of sugar beet pulp and roasted canola seed in a concentrate diet altered carcass traits, muscle (longissimus dorsi) composition and meat sensory properties of Arabian fattening lambs. Small Rumin. Res. 2017. [Google Scholar] [CrossRef]
- Gallo, S.B.; de Almeida Merlin, F.; de Macedo, C.M.; de Oliveira Silveira, R.D. Whole grain diet for Feedlot Lambs. Small Rumin. Res. 2014, 120, 185–188. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Small Ruminats: Sheep, Goats, Cervids and New World Camelids, 6th ed.; National Academy Press: Washington, DC, USA, 2007. [Google Scholar]
- Ametaj, B.N.; Emmanuel, D.G.V.; Zebeli, Q.; Dunn, S.M. Feeding high proportions of barley grain in a total mixed ration perturbs diurnal patterns of plasma metabolites in lactating dairy cows. J. Dairy Sci. 2009, 92, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. The role of oxidative stress in small ruminants’ health and production. Rev. Bras. Zootec. 2010, 39, 348–363. [Google Scholar] [CrossRef]
- Sgorlon, S.; Stradaioli, G.; Gabai, G.; Stefanon, B. Variation of starch and fat in the diet affects metabolic status and oxidative stress in ewes. Small Rumin. Res. 2008, 74, 123–129. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef]
- Stevanović, Z.D.; Bošnjak-Neumüller, J.; Pajić-Lijaković, I.; Raj, J.; Vasiljević, M. Essential Oils as Feed Additives-Future Perspectives. Molecules 2018, 23, 1717. [Google Scholar] [CrossRef]
- Murakami, A.E.; Eyng, C.; Torrent, J. Effects of functional oils on coccidiosis and apparent metabolizable energy in broiler chickens. Asian-Australasian J. Anim. Sci. 2014, 27, 981–989. [Google Scholar] [CrossRef]
- Osmari, M.P.; De Matos, L.F.; Salab, B.L.; Diaz, T.G. Líquido da casca da castanha de caju: Características e aplicabilidades na produção animal Cashew nut shell liquid: Characteristics and applicability in animal production. Maringá 2015, 9, 143–149. [Google Scholar]
- Zotti, C.A.; Silva, A.P.; Carvalho, R.; Marino, C.T.; Rodrigues, P.H.M.; Silva, L.F.P.; McAllister, T.A.; Leme, P.R. Monensin and a blend of castor oil and cashew nut shell liquid used in a high-concentrate diet abruptly fed to Nellore cattle. J. Anim. Sci. 2017, 95, 4124. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.D.J.A.D.S.; Araújo, B.Q.; Citó, A.M.D.G.L.; Da Silva, J.; Saffi, J.; Richter, M.F.; Ferraz, A.D.B.F. Antioxidant properties and chemical composition of technical Cashew Nut Shell Liquid (tCNSL). Food Chem. 2011, 126, 1044–1048. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Ponnampalam, E.N.; Leury, B.J.; Liu, F.; Dunshea, F.R. Antioxidant dynamics in the live animal and implications for ruminant health and product (meat/milk) quality: Role of vitamin E and selenium. Anim. Prod. Sci. 2014, 54, 1525–1536. [Google Scholar] [CrossRef]
- Halliwell, B.; Chirico, S. Lipid peroxidation: Significance and its mechanism. Am. J. Clin. Nutr. 1993, 57, 715S–725S. [Google Scholar] [CrossRef]
- Bezerra, H.V.A.; Gallo, S.B.; Rosa, A.F.; Fernandes, A.C.; da Le Silva, S.; Leme, P.R. Impact of purified lignin on performance, rumen health, oxidative stress control and meat quality of lambs fed a high-concentrate diet. Livest. Sci. 2020, 231, 103882. [Google Scholar] [CrossRef]
- Spears, J.W. Comparative Trace Element Nutrition Trace Mineral Bioavailability in Ruminants. J. Nutr. 2003, 133, 1506–1509. [Google Scholar] [CrossRef]
- Singh, U.; Devaraj, S.; Jialal, I. Vitamin E, oxidative stress, and inflammation. Annu. Rev. Nutr. 2005, 25, 151–174. [Google Scholar] [CrossRef]
- Venkateswaran, V.; Fleshner, N.E.; Klotz, L.H. Synergistic effect of vitamin E and selenium in human prostate cancer cell lines. Prostate Cancer Prostatic Dis. 2004, 7, 54–56. [Google Scholar] [CrossRef]
- Hogan, J.S.; Smith, K.L.; Weiss, W.P.; Todhunter, D.A.; Schockey, W.L. Relationships Among Vitamin E, Selenium, and Bovine Blood Neutrophils. J. Dairy Sci. 1990, 73, 2372–2378. [Google Scholar] [CrossRef]
- Coneglian, S.M.; Castañeda-Serrano, R.D.; Cruz, O.T.B.; Branco, A.F. Effects of essential oils of Cashew and Castor on intake, digestibility, ruminal fermentation and purine derivatives in beef cattle fed high grain diets. Semin. Agrar. 2019, 40, 2057–2069. [Google Scholar] [CrossRef]
- Weiss, W.P.; Conrad, H.R.; St. Pierre, N.R. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- AOAC-Association of Official Agricultural Chemistis. Proceedings of the Official Methods of Analysis of AOAC International, 16th ed.; AOAC-Association of Official Agricultural Chemistis: Arlington, TX, USA, 1995; pp. 10–11. [Google Scholar]
- Olson, O.E.; Cary, I.S.; Palmer, E.E. Modification of the official fluorimetric method for selenium in plants. Assoc. Off. Anal. Chem. 1975, 58, 117–121. [Google Scholar]
- Bigham, M.L.; McManaus, W.R. Whole wheat grain feeding of lambs. Effects of roughage and wheat grain mixtures. Aust. J. Agric. Res. 1975, 26, 1053–1062. [Google Scholar] [CrossRef]
- Resende-Junior, J.C.; Alonso, L.D.S.; Pereira, M.N.; Roca, M.G.; Duboc, M.V.; De Oliveira, E.C.; De Melo, L.Q. Effect of the feeding pattern on rumen wall morphology of cows and sheep. Brazilian J. Vet. Res. Anim. Sci. 2006, 43, 526–536. [Google Scholar] [CrossRef][Green Version]
- CIE. Recommendations on Uniform Color Spaces-Color Equations, Psychometric Color Terms. In Proceedings of the Commission Internationale de l’eclairage (CIE); CIE: Paris, France, 1986. [Google Scholar]
- AMSA-American Meat Science Association Research Guidelines for Cookery, Sensory Evaluation and Instrumental Tenderness Measurements of Fresh Meat. In Proceedings of the American Meat Science Association; AMSA: Chicago, IL, USA, 1995.
- Vyncke, W. Evaluation of the Direct Thiobarbituric Acid Extraction Method for Determining Oxidative Rancidity in Mackerel (Scomber scombrus) L. Fette Seifen Anstrichm. 1975, 77, 239–240. [Google Scholar] [CrossRef]
- APHA. Compendium of Methods for the Microbiological Examination of Foods, 3rd ed.; APHA: Washington, DC, USA, 2001. [Google Scholar]
- Fairweather-Tait, S.; Hurrel, R.F. Bioavailability of minerals and trace elements. Members of EC Flair Concerted Action No. 10: Measurement of micronutrient Swiss Federal Institute of Technology Zurich, Laboratory for Human Nutrition. Measurement 2018, 9, 295–324. [Google Scholar]
- Paiva, F.A.; Netto, A.S.; Corrêa, L.B.; Silva, T.H.; Guimarães, I.C.S.B.; Del Claro, G.R.; Cunha, J.A.; Zanetti, M.A. Organic selenium supplementation increases muscle selenium content in growing lambs compared to inorganic source. Small Rumin. Res. 2019, 175, 57–64. [Google Scholar] [CrossRef]
- Bodnar, M.; Szczyglowska, M.; Konieczka, P.; Namiesnik, J. Methods of Selenium Supplementation: Bioavailability and Determination of Selenium Compounds. Crit. Rev. Food Sci. Nutr. 2016, 56, 36–55. [Google Scholar] [CrossRef]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H.E. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Bleys, J.; Navas-Acien, A.; Guallar, E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch. Intern. Med. 2008, 168, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.R.; Berry, M.J. The influence of selenium on immune responses. Mol. Nutr. Food Res. 2008, 52, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein P-Expression, functions, and roles in mammals. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 1441–1447. [Google Scholar] [CrossRef]
- Köhrle, J.; Gärtner, R. Selenium and thyroid. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 815–827. [Google Scholar] [CrossRef]
- Fisinin, V.I.; Papazyan, T.T.; Surai, P.F. Producing selenium-enriched eggs and meat to improve the selenium status of the general population. Crit. Rev. Biotechnol. 2009, 29, 18–28. [Google Scholar] [CrossRef]
- Pardi, M.C.; Dos Santos, I.F.; De Souza, E.R.; Pardi, H.S. Ciência, Higiene e Tecnologia da Carne, 2nd ed.; CEGRAF-UFG, Ed.; EDUFF: Niterói, Brazil, 1993. [Google Scholar]
- Van Oeckel, M.J.; Warnants, N.; Boucqué, C.V. Comparison of different methods for measuring water holding capacity and juiciness of pork versus on-line screening methods. Meat Sci. 1999, 51, 313–320. [Google Scholar] [CrossRef]
- Warner, R.D. The Eating Quality of Meat-IV Water-Holding Capacity and Juiciness; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; ISBN 9780081006979. [Google Scholar]
- Huff-Lonergan, E.; Lonergan, S.M. Mechanisms of water-holding capacity of meat: The role of postmortem biochemical and structural changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef]
- Calvo, L.; Toldrá, F.; Aristoy, M.C.; López-Bote, C.J.; Rey, A.I. Effect of dietary organic selenium on muscle proteolytic activity and water-holding capacity in pork. Meat Sci. 2016, 121, 1–11. [Google Scholar] [CrossRef]
- Hollands, A.; Corriden, R.; Gysler, G.; Dahesh, S.; Olson, J.; Ali, S.R.; Kunkel, M.T.; Lin, A.E.; Forli, S.; Newton, A.C.; et al. Natural product anacardic acid from cashew nut shells stimulates neutrophil extracellular trap production and bactericidal activity. J. Biol. Chem. 2016, 291, 13964–13973. [Google Scholar] [CrossRef]
- Schmidt, R.L.; Simonović, M. Synthesis and decoding of selenocysteine and human health. Croat. Med. J. 2012, 53, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Tüzün, A.; Erdil, A.; Inal, V.; Aydin, A.; Baǧci, S.; Yeşilova, Z.; Sayal, A.; Karaeren, N.; Daǧalp, K. Oxidative stress and antioxidant capacity in patients with inflammatory bowel disease. Clin. Biochem. 2002, 35, 569–572. [Google Scholar] [CrossRef]
- Melo Cavalcante, A.A.; Rubensam, G.; Picada, J.N.; Gomes da Silva, E.G.; Fonseca Moreira, J.C.; Henriques, J.A.P. Mutagenicity, antioxidant potential, and antimutagenic activity against hydrogen peroxide of cashew (Anacardium occidentale) apple juice and cajuina. Environ. Mol. Mutagen. 2003, 41, 360–369. [Google Scholar] [CrossRef]
- Pandey, D.; Chen, F.; Patel, A.; Wang, C.; Dimitropoulou, C.; Patel, V.; Rudic, R.D.; Stepp, D.; Fulton, D. SUMO1 negatively regulates ROS production from NADPH oxidases. Arter. Thromb Vasc Biol. 2011, 31, 1634–1642. [Google Scholar] [CrossRef]
- Esterbauer, H.; Striegl, G.; Puhl, H.; Oberreither, S.; Rotheneder, M.; El-Saadani, M.; Jurgens, G. The Role of Vitamin E and Carotenoids in Preventing Oxidation of Low Density Lipoproteins. Ann. N. Y. Acad. Sci. 1989, 570, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Pütter, J. Methods of Enzymatic Analysis V4, 2nd ed.; Bergmeyer, H.-U., Ed.; Academic Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Pratt, D.S.; Kaplan, M.M. Evaluation of abnormal liver-enzyme results in asymptomatic patients. N. Engl. J. Med. 2000, 342, 1266–1271. [Google Scholar] [CrossRef]
- Smilin Bell Aseervatham, G.; Arul Ananth, D.; Sivasudha, T. The Liver: Oxidative Stress and Dietary Antioxidants; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 149, ISBN 9780128039519. [Google Scholar]
- Schwarz, K. Essentiality and metabolic functions of selenium. Med. Clin. N. Am. 1976, 60, 745–758. [Google Scholar] [CrossRef]
- Toyomizu, M.; Okamoto, K.; Ishibashi, T.; Nakatsu, T.; Akiba, Y. Reducing effect of dietary anacardic acid on body fat pads in rats. Anim. Sci. J. 2003, 74, 499–504. [Google Scholar] [CrossRef]
- Braz, N.M.; Freitas, E.R.; Trevisan, M.T.S.; do Nascimento, G.A.J.; Salles, R.P.R.; Cruz, C.E.B.; Farias, N.N.P.; da Silva, I.N.G.; Watanabe, P.H. Serum biochemical profile, enzymatic activity and lipid peroxidation in organs of laying hens fed diets containing cashew nut shell liquid. J. Anim. Physiol. Anim. Nutr. 2018, 102, 67–74. [Google Scholar] [CrossRef]
- Lauridsen, C.; Buckley, D.J.; Morrissey, P.A. Influence of dietary fat and vitamin E supplementation on α-tocopherol levels and fatty acid profiles in chicken muscle membranal fractions and on susceptibility to lipid peroxidation. Meat Sci. 1997, 46, 9–22. [Google Scholar] [CrossRef]
- Rey, A.I.; De Cara, A.; Segura, J.F.; Martí, P.; Hechavarría, T.; Calvo, L. Dietary oleuropein extract supplementation and its combination with α-tocopheryl acetate and selenium modifies the free fatty acid profile of pork and improves its stability. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ferket, P.R.; Malheiros, R.D.; Moraes, V.M.B.; Ayoola, A.A.; Barasch, I.; Toomer, O.T.; Torrent, J. Effects of functional oils on the growth, carcass and meat characteristics, and intestinal morphology of commercial turkey toms. Poult. Sci. 2020, 99, 3752–3760. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.R.; Kethireddipalli, P.; Edition, T. Psychrotrophic Bacteria Milk and Milk Products|Micro-Biology of Liquid Milk; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Iulietto, M.F.; Sechi, P.; Borgogni, E.; Cenci-Goga, B.T. Meat spoilage: A critical review of a neglected alteration due to ropy slime producing bacteria. Ital. J. Anim. Sci. 2015, 14, 316–326. [Google Scholar] [CrossRef]
- Naguib, M.M.; Valvano, M.A. Vitamin E Increases Antimicrobial Sensitivity by Inhibiting Bacterial Lipocalin Antibiotic Binding. mSphere 2018, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.; Wunderlich, F.; Sies, H. Dietary Selenium in Adjuvant Therapy of Viral. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Pittman, C.I.; Pendleton, S.; Bisha, B.; O’Bryan, C.A.; Belk, K.E.; Goodridge, L.; Crandall, P.G.; Ricke, S.C. Activity of Citrus Essential Oils againstEscherichia coli O157:H7 and Salmonella spp. and Effects on Beef Subprimal Cuts under Refrigeration. J. Food Sci. 2011, 76, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods-A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Moraes, P.O.; Cardinal, K.M.; Gouvêa, F.L.; Schroeder, B.; Ceron, M.S.; Lunedo, R.; Frazzon, A.P.G.; Frazzon, J.; Ribeiro, A.M.L. Comparison between a commercial blend of functional oils and monensin on the performance and microbiota of coccidiosis-challenged broilers. Poult. Sci. 2019, 98, 5456–5464. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; Van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef]




| Traits | Treatment | p-Value | |||
|---|---|---|---|---|---|
| Control | FO | FO Plus SeE | SeE | ||
| Initial BW, kg | 22.26 ± 2.41 | 22.32 ± 2.50 | 22.79 ± 1.97 | 22.32 ± 2.18 | 0.9621 |
| Final BW, kg | 40.21 ± 2.06 | 38.79 ± 3.32 | 41.71 ± 3.24 | 39.88 ± 3.04 | 0.234 |
| ADG, kg | 0.35 ± 0.05 | 0.32 ± 0.03 | 0.36 ± 0.04 | 0.34 ± 0.06 | 0.359 |
| DM intake, kg/day | 1.49 ± 0.09 | 1.50 ± 0.05 | 1.46 ± 0.22 | 1.41 ± 0.20 | 0.817 |
| Feed efficiency, g/kg DMI | 0.23 ± 0.04 | 0.21 ± 0.03 | 0.25± 0.02 | 0.24± 0.04 | 0.229 |
| Hot carcass weight, kg | 19.46 ± 0.99 | 18.94 ± 1.97 | 20.28 ± 0.04 | 19.58 ± 1.07 | 0.252 |
| Hot carcass yield, % | 48.42 ± 1.46 | 48.78 ± 1.67 | 48.61 ± 1.07 | 49.16 ± 1.67 | 0.791 |
| pH 1 h | 6.45 ± 0.19 | 6.58 ± 0.14 | 6.63 ± 0.12 | 6.49 ± 0.20 | 0.144 |
| Temperature 1 h, °C | 30.11 ± 2.82 | 29.35 ± 1.34 | 29.59 ± 2.51 | 29.49 ± 1.81 | 0.915 |
| Cold carcass weight, kg | 18.94 ± 1.05 | 18.41 ± 2.06 | 19.71 ± 1.59 | 19.08 ± 1.06 | 0.298 |
| Cold carcass yield, % | 47.10 ± 1.48 | 47.39 ± 1.79 | 47.25 ± 1.14 | 47.91 ± 1.61 | 0.741 |
| pH 24 h | 5.87 ± 0.42 | 5.82 ± 0.34 | 5.68 ± 0.06 | 5.69 ± 0.09 | 0.423 |
| Temperature 24 h, °C | 11.59 ± 3.25 | 12.99 ± 3.49 | 12.98 ± 2.48 | 13.26± 2.95 | 0.699 |
| Loin eye area, cm2 | 28.00 ± 5.13 | 30.25 ± 7.18 | 30.38 ± 6.05 | 32.38 ± 4.65 | 0.560 |
| Subcutaneous fat thickness, mm | 7.75 ± 4.40 | 5.75 ± 2.18 | 7.88 ± 2.35 | 6.50 ± 2.56 | 0.447 |
| Cooking loss, % | 24.58 ± 4.30 ab | 26.66 ± 3.34 ab | 29.91 ± 4.32 a | 23.81 ± 4.35 b | 0.028 |
| Shear force, N | 39.66 ± 11.77 | 42.34 ± 12.56 | 50.85 ± 8.41 | 47.33 ± 11.40 | 0.231 |
| Muscle selenium, mg/kg | 0.07 ± 0.01 b | 0.05 ± 0.007 b | 0.10 ± 0.01 a | 0.11 ± 0.01 a | <0.0001 |
| Serum selenium, mg/kg | 0.11 ± 0.001 b | 0.11 ± 0.01 b | 0.15 ± 0.009 a | 0.13 ± 0.01 a | <0.0001 |
| Parameters | Treatment | ||||
|---|---|---|---|---|---|
| Control | FO | FO Plus SeE | SeE | p-Value | |
| Rumenitis score | 0.88 ± 0.64 | 1.63 ± 1.84 | 1.13 ± 0.99 | 1.50 ± 1.06 | 0.551 |
| Papillary area, cm2 | 0.24 ± 0.04 | 0.26 ± 0.08 | 0.31 ± 0.09 | 0.30 ± 0.10 | 0.378 |
| Absorption surface, cm2 of wall | 14.68 ± 2.89 | 16.44 ± 5.45 | 15.88 ± 4.31 | 15.13 ± 5.83 | 0.899 |
| Absorption surface, % | 93.67 ± 1.91 | 94.04 ± 2.15 | 93.88 ± 1.96 | 93.18 ± 2.71 | 0.890 |
| Papilla number/cm2 of wall | 59.15 ± 18.38 | 59.41 ± 9.72 | 49.50 ± 10.24 | 47.51 ± 12.49 | 0.172 |
| Parameters | Treatment | Days | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | FO | FO Plus SeE | SeE | 0 | 30 | 53 | Treatment | Time | Treatment * Time | |
| SOD | 0.261 ± 0.42 | 0.278 ± 0.34 | 0.375 ± 0.44 | 0.438 ± 0.47 | 0.410 ± 0.29 | 0.407 ± 0.37 | 0.197 ± 0.38 | 0.412 | 0.072 | 0.369 |
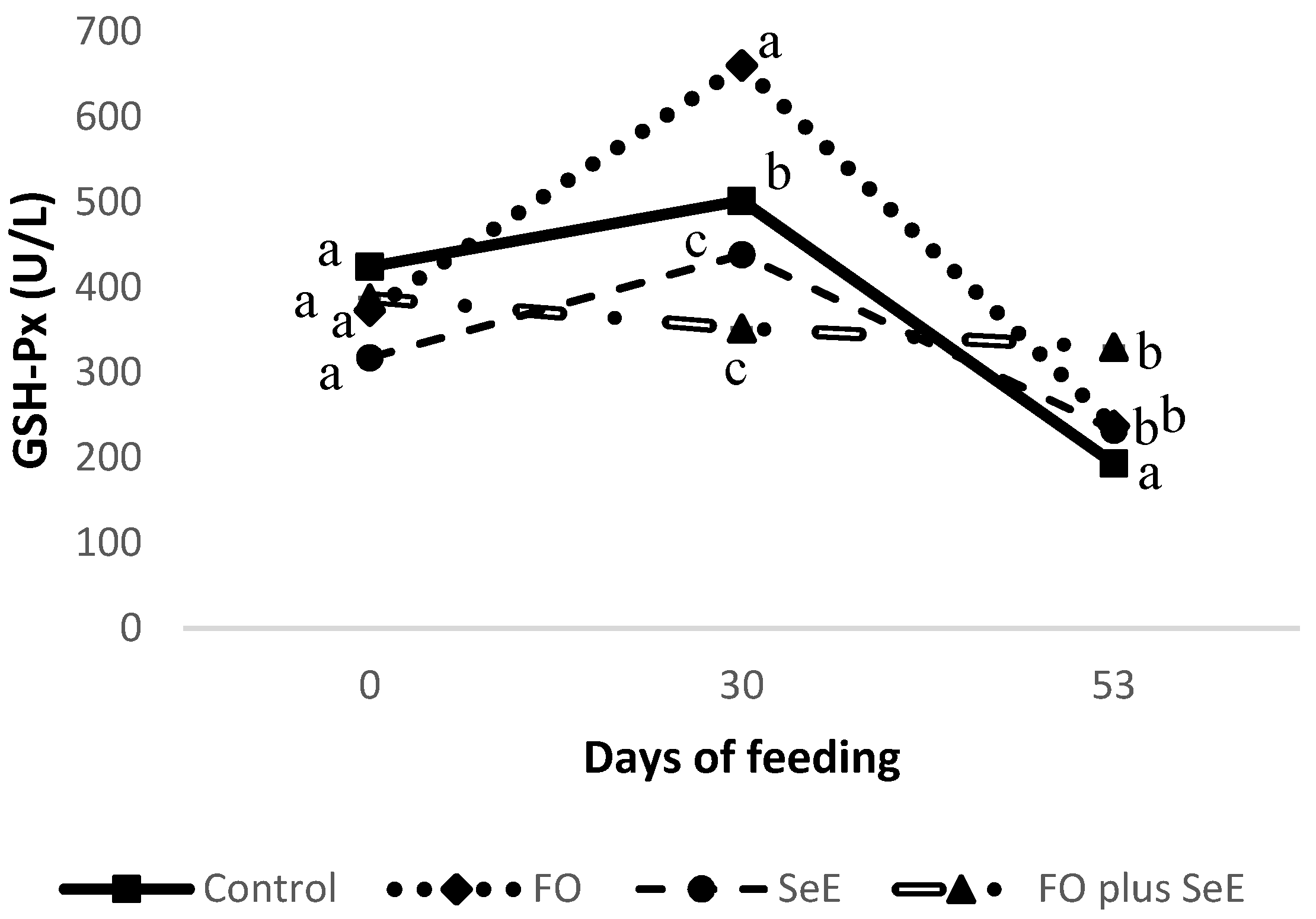
| GSH-Px | 373.12 ± 213 | 423.62 ± 235 | 356.57 ± 113 | 329.58 ± 150 | 375.56 ± 102 b | 488.14 ±149 a | 248.46 ± 132 c | 0.157 | <0.0001 | 0.006 |
| Catalase | 2.18 ± 2.26 | 2.34 ± 1.44 | 1.64 ± 2.79 | 2.46 ± 1.25 | 2.32 ±0.96 | 1.71 ±1.75 | 2.44 ± 1.49 | 0.531 | 0.353 | 0.852 |
| Peroxidases | 3.58 ± 1.37 b | 4.57 ± 1.27 ab | 8.11 ± 1.40 a | 2.65 ± 1.30 b | 5.56 ± 1.17 a | 2.23 ± 1.16 b | 6.40 ± 1.44 a | 0.035 | 0.031 | 0.539 |
| H2O2 | 932.56 ± 478 | 896.10± 181 | 891.83 ± 181 | 906.45 ± 247 | 988.38 ± 375 | 894.08 ± 261 | 837.74 ± 236 | 0.972 | 0.146 | 0.273 |
| ALT | 12.00 ± 4.81 a | 14.24 ± 6.54 a | 9.08 ± 5.74 b | 9.71 ± 3.93 b | 13.00 ± 4.93 | 10.46 ± 5.07 | 10.31 ± 5.16 | 0.006 | 0.095 | 0.935 |
| AST | 112.62 ± 32.54 a | 108.24 ± 35.83 a | 92.75 ± 14.58 b | 98.00 ± 25.78 b | 84.33 ± 10.34 c | 113.13 ± 20.14 ab | 111.25 ± 21.63 b | 0.033 | <0.0001 | 0.641 |
| ALP | 593.24 ± 208 | 608.79 ± 250 | 609.75 ± 165 | 600.29 ± 216 | 651.91 ± 187 | 605.49 ± 209 | 551.66 ± 209 | 0.990 | 0.193 | 1.000 |
| GGT | 87.03 ± 30.11 | 78.85 ± 30.29 | 75.54 ± 18.22 | 82.25 ± 35.39 | 59.79 ± 13.70 cc | 90.36 ± 19.96 ab | 92.59 ± 22.20 a | 0.447 | <0.0001 | 0.857 |
| Cholesterol | 62.75 ± 13.25 | 58.53 ± 15.05 | 53.90 ± 11.54 | 60.29 ± 14.47 | 49.87 ± 9.12 | 64.35 ± 10.19 ab | 62.38 ± 11.56 ab | 0.124 | <0.0001 | 0.114 |
| Measurements | Treatment | Days on Display | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | FO | FO Plus SeE | SeE | 0 | 3 | 5 | Treatment | Time | Treatment * Time | |
| Color L* | 42.18 ± 3.74 c | 44.02 ± 3.74 ab | 44.79 ± 2.60 ab | 44.84 ± 2.60 ab | 42.56 ± 3.81 bc | 45.40 ± 4.17 a | 43.91 ± 2.04 bc | 0.026 | 0.005 | 0.630 |
| Color a* | 14.87 ± 4.49 | 14.29 ± 3.76 | 13.84 ± 3.64 | 14.09 ± 3.78 | 15.43 ± 2.70 | 13.76 ± 4.50 | 13.63 ± 1.79 | 0.830 | 0.140 | 0.999 |
| Color b* | 11.87 ± 2.08 | 12.99 ± 2.30 | 12.62 ± 1.09 | 13.11 ± 1.74 | 11.14 ± 3.14 c | 13.41 ± 3.07 ab | 13.38 ± 1.58 ab | 0.121 | <0.0001 | 0.462 |
| TBARS 1 | 0.142 ± 0.09 a | 0.197 ± 0.015 a | 0.072 ± 0.04 b | 0.054 ± 0.03 b | 0.104 ± 0.05 | 0.079 ± 0.05 | 0.166 ± 0.13 | 0.001 | 0.020 | 0.401 |
| Mesophilic | 6.31 ±0.39 | 5.71 ± 0.33 | 5.58 ± 0.32 | 5.50 ± 0.20 | 2.07 ± 0.74 | 5.06 ± 0.38 | 6.37 ± 0.35 | 0.443 | 0.081 | 0.573 |
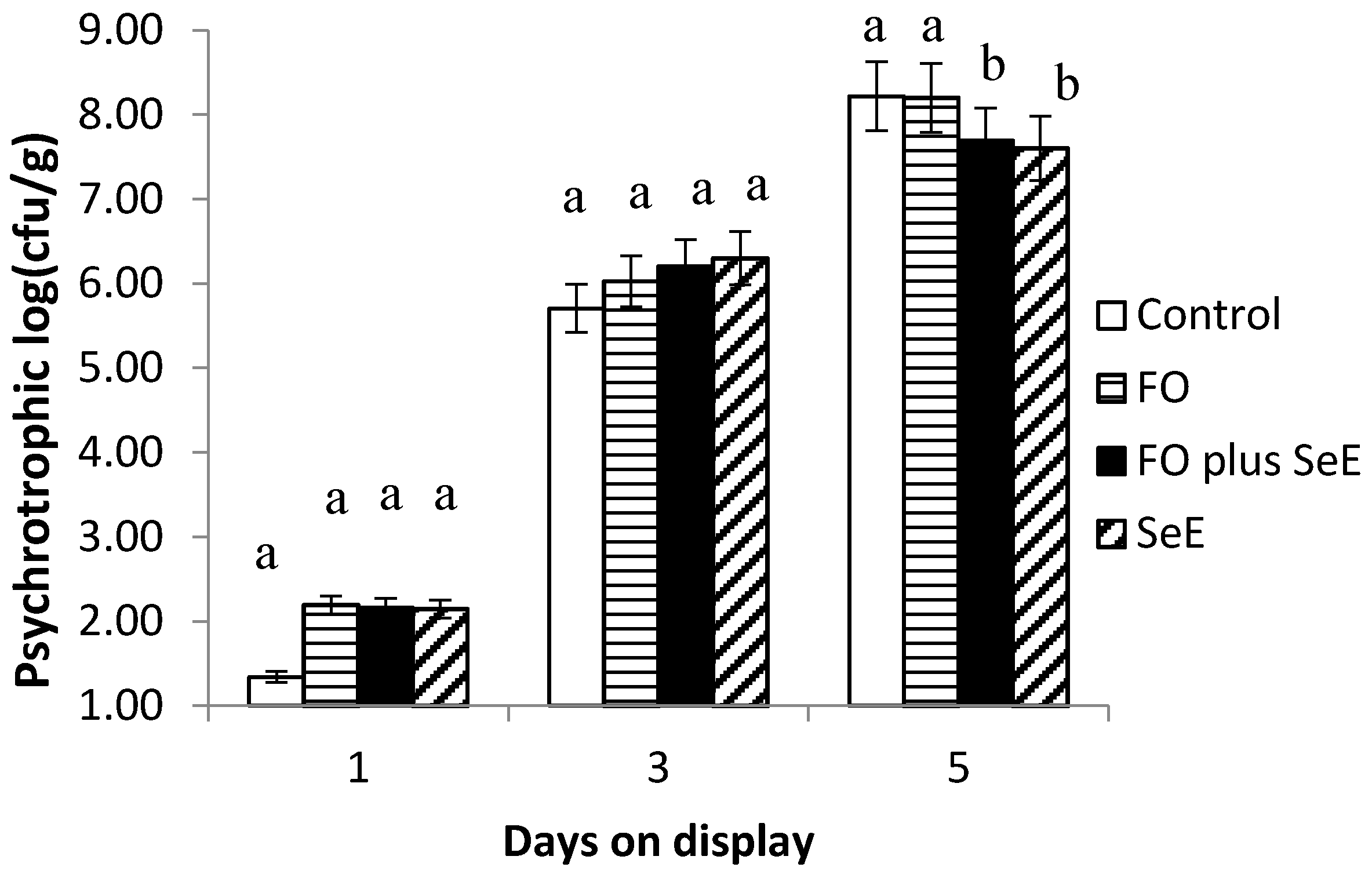
| Psychrotrophic | 7.74 ± 0.30 | 7.72 ± 0.46 | 7.23 ± 0.19 | 7.14 ± 0.18 | 2.06 ± 0.20 | 6.11 ± 0.17 | 8.01 ± 0.84 | 0.056 | <0.0001 | 0.021 |
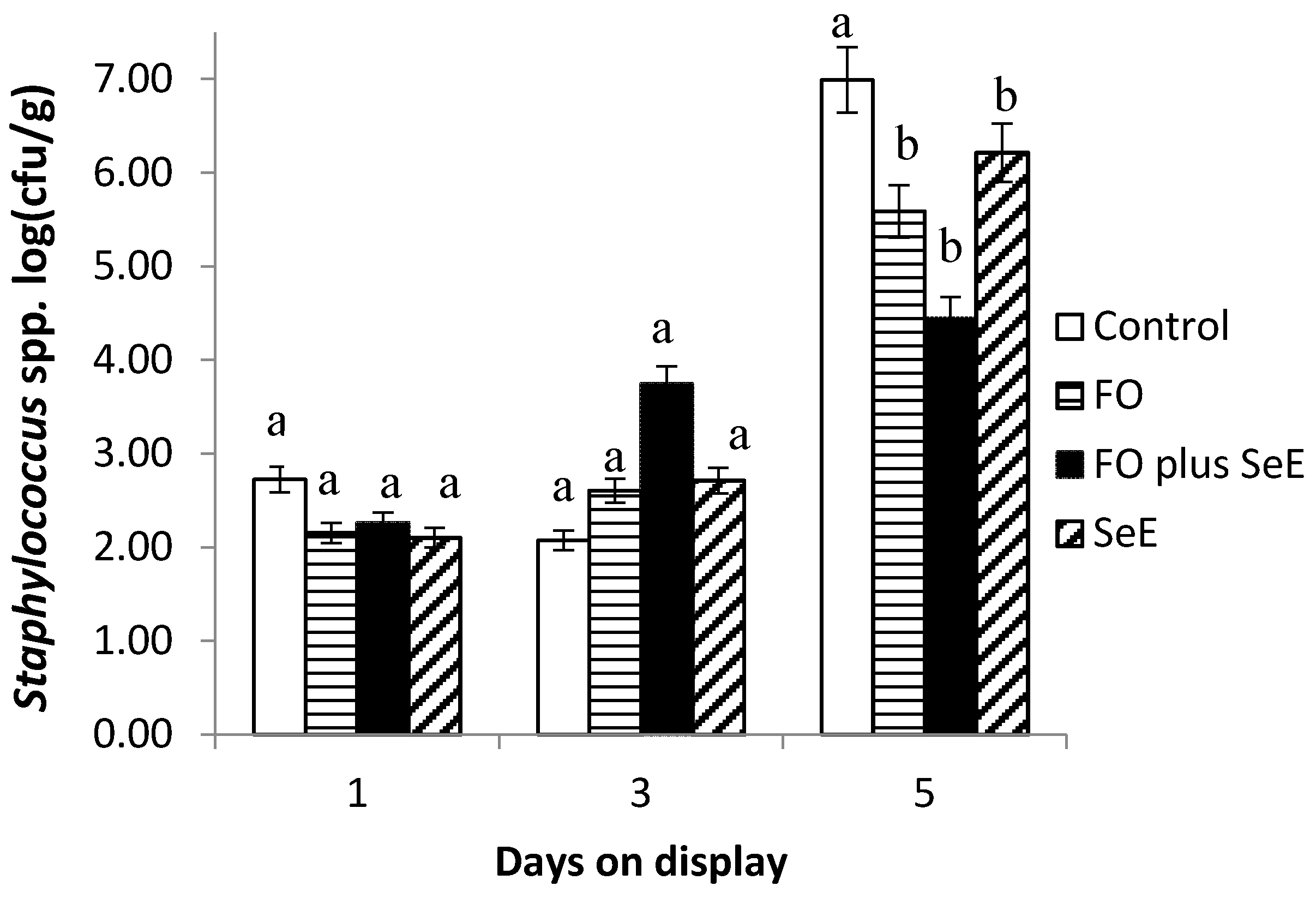
| Staphylococcus spp. | 6.51 ± 0.37 | 5.11 ± 0.24 | 4.05 ± 0.15 | 5.74 ± 0.11 | 2.39 ± 0.27 | 3.22 ± 0.20 | 6.47 ± 0.39 | 0.061 | 0.010 | 0.007 |
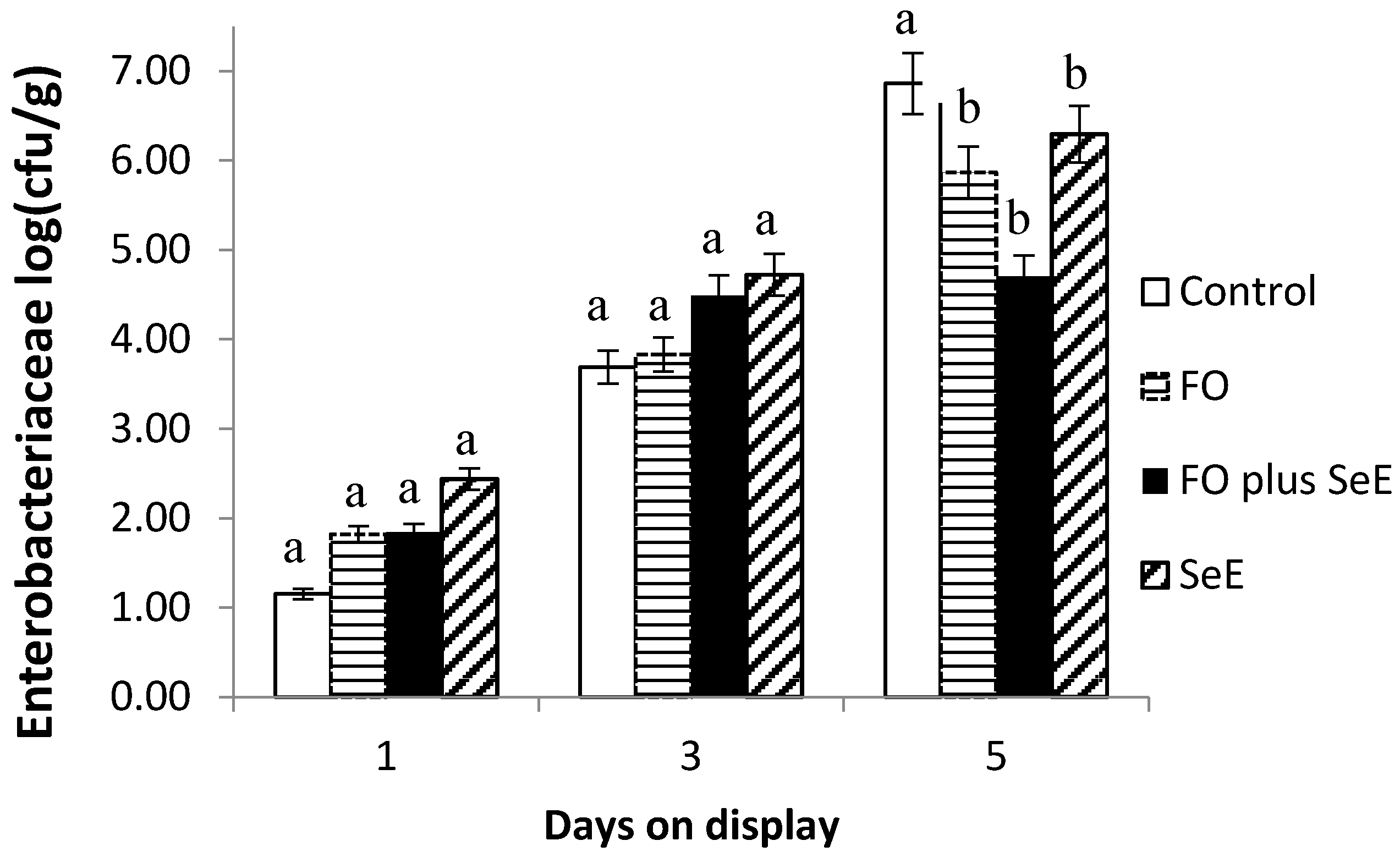
| Enterobacteriaceae | 6.38 ± 0.33 | 5.39 ± 0.42 | 4.43 ± 0.40 | 5.83 ± 0.12 | 2.03 ± 0.18 | 4.38 ± 0.36 | 6.40 ± 0.37 | 0.309 | 0.016 | 0.050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra, H.V.A.; Buarque, V.L.M.; Silva, L.S.B.; Leme, P.R.P.; Vidal, A.M.C.; Vaz, A.C.N.; Gallo, S.B.; Silva, S.L.; Leme, P.R. Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet. Antioxidants 2020, 9, 1298. https://doi.org/10.3390/antiox9121298
Bezerra HVA, Buarque VLM, Silva LSB, Leme PRP, Vidal AMC, Vaz ACN, Gallo SB, Silva SL, Leme PR. Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet. Antioxidants. 2020; 9(12):1298. https://doi.org/10.3390/antiox9121298
Chicago/Turabian StyleBezerra, Helena Viel Alves, Vicente Luiz Macedo Buarque, Lucas Santos Bermudes Silva, Paulo Roberto Pedroso Leme, Ana Maria Centola Vidal, Andréia Cristina Nakashima Vaz, Sarita Bonagurio Gallo, Saulo Luz Silva, and Paulo Roberto Leme. 2020. "Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet" Antioxidants 9, no. 12: 1298. https://doi.org/10.3390/antiox9121298
APA StyleBezerra, H. V. A., Buarque, V. L. M., Silva, L. S. B., Leme, P. R. P., Vidal, A. M. C., Vaz, A. C. N., Gallo, S. B., Silva, S. L., & Leme, P. R. (2020). Effect of Castor and Cashew Nut Shell Oils, Selenium and Vitamin E as Antioxidants on the Health and Meat Stability of Lambs Fed a High-Concentrate Diet. Antioxidants, 9(12), 1298. https://doi.org/10.3390/antiox9121298