First Evidence of a Protective Effect of Plant Bioactive Compounds against H2O2-Induced Aconitase Damage in Durum Wheat Mitochondria


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Plant Materials
2.2. In Vitro Antioxidant Capacity (AC) Determination by Means of Oxygen Radical Absorbance Capacity (ORAC) and Trolox Equivalent Antioxidant Capacity (TEAC) Methods
2.2.1. ORAC Method
2.2.2. TEAC Method
2.3. Durum Wheat Mitochondria (DWM) Isolation
2.4. Aconitase Activity Assay
2.5. Statistical Analysis
3. Results and Discussion
3.1. In Vitro Antioxidant Capacity (AC) of Pure Phytochemicals
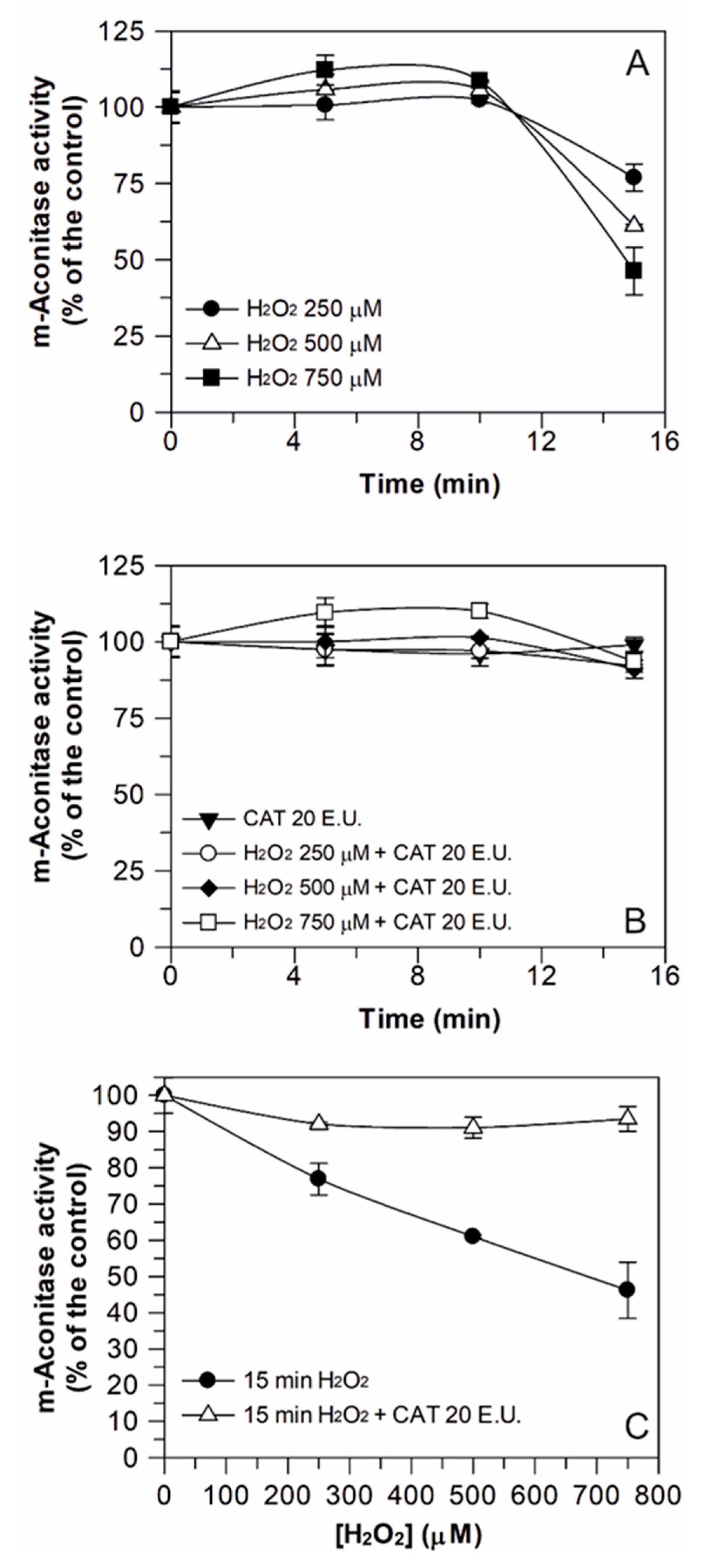
3.2. Induction of H2O2 Damage to Aconitase Activity
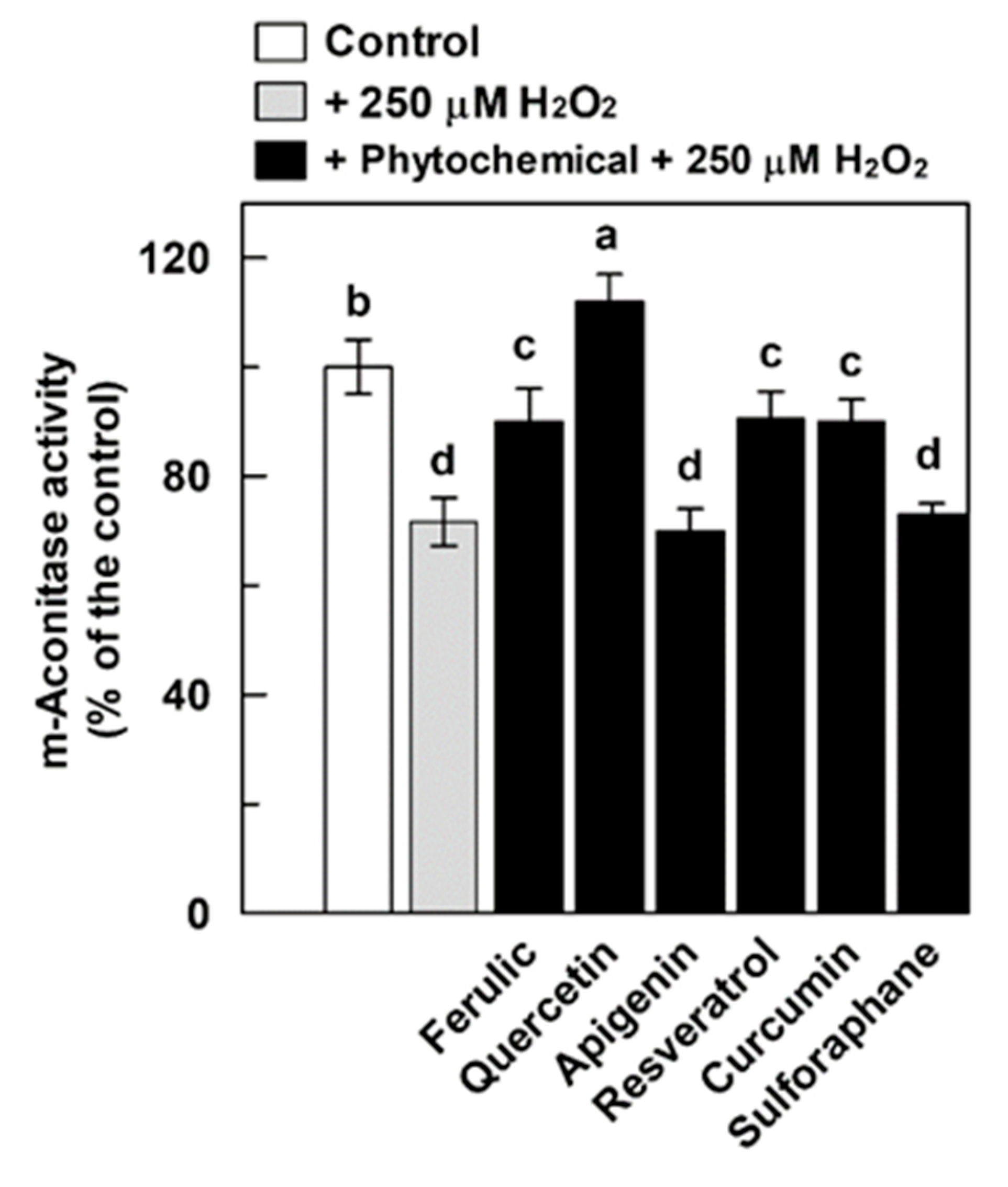
3.3. Effect of Phytochemicals on the H2O2-Induced Damage of Aconitase
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leonov, A.; Arlia-Ciommo, A.; Piano, A.; Svistkova, V.; Lutchman, V.; Medkour, Y.; Titorenko, V.I. Longevity extension by phytochemicals. Molecules 2015, 20, 6544–6572. [Google Scholar] [CrossRef] [PubMed]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Zhong, Y. Measurement of antioxidant activity. J. Funct. Foods 2015, 18, 757–781. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxycol. 2020, 94, 657–715. [Google Scholar] [CrossRef] [PubMed]
- Soccio, M.; Laus, M.N.; Flagella, Z.; Pastore, D. Assessment of antioxidant capacity and putative healthy effects of natural plant products using soybean lipoxygenase-based methods. An overview. Molecules 2018, 23, 3244. [Google Scholar] [CrossRef]
- Huang, D.; Boxin, O.U.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Fraga, C.G.; Oteiza, P.I.; Galleano, M. In vitro measurements and interpretation of total antioxidant capacity. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 931–934. [Google Scholar] [CrossRef]
- Pompella, A.; Sies, H.; Wacker, R.; Brouns, F.; Grune, T.; Biesalski, H.K.; Frank, J. The use of total antioxidant capacity as surrogate marker for food quality and its effect on health is to be discouraged. Nutrition 2014, 30, 791–793. [Google Scholar] [CrossRef]
- Gibellini, L.; Bianchini, E.; De Biasi, S.; Nasi, M.; Cossarizza, A.; Pinti, M. Natural Compounds Modulating Mitochondrial Functions. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef]
- Grabacka, M.M.; Gawin, M.; Pierzchalska, M. Phytochemical modulators of mitochondria: The search for chemopreventive agents and supportive therapeutics. Pharmaceuticals 2014, 7, 913–942. [Google Scholar] [CrossRef]
- Pastore, D.; Trono, D.; Laus, M.N.; Di Fonzo, N.; Flagella, Z. Possible plant mitochondria involvement in cell adaptation to drought stress - A case study: Durum wheat mitochondria. J. Exp. Bot. 2007, 58, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Raza, H.; John, A.; Brown, E.M.; Benedict, S.; Kambal, A. Alterations in mitochondrial respiratory functions, redox metabolism and apoptosis by oxidant 4-hydroxynonenal and antioxidants curcumin and melatonin in PC12 cells. Toxicol. Appl. Pharmacol. 2008, 226, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ravanel, P.; Tissut, M.; Douce, R. Effects of flavone on the oxidative properties of intact plant mitochondria. Phytochemistry 1981, 20, 2101–2103. [Google Scholar] [CrossRef]
- Verniquet, F.; Gaillard, J.; Neuberger, M.; Douce, R. Rapid inactivation of plant aconitase by hydrogen peroxide. Biochem. J. 1991, 276, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.V.; Piroddi, M.; Galli, F.; Lushchak, V.I. Aconitase post-translational modification as a key in linkage between Krebs cycle, iron homeostasis, redox signaling, and metabolism of reactive oxygen species. Redox Rep. 2014, 19, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A.U. Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot. 2012, 63, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Gardeström, P. Regulation of NAD- and NADP-dependent isocitrate dehydrogenases by reduction levels of pyridine nucleotides in mitochondria and cytosol of pea leaves. Biochim. Biophys. Acta Bioenerg. 2003, 1606, 117–125. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Nikitina, M.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of aconitase in maize scutellum. J. Plant Physiol. 2015, 181, 14–19. [Google Scholar] [CrossRef]
- Gardner, P.R. Aconitase: Sensitive target and measure of superoxide. Methods Enzymol. 2002, 349, 9–23. [Google Scholar] [CrossRef]
- Palmieri, E.M.; Gonzalez-Cotto, M.; Baseler, W.A.; Davies, L.C.; Ghesquière, B.; Maio, N.; Rice, C.M.; Rouault, T.A.; Cassel, T.; Higashi, R.M.; et al. Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Ratcliffe, R.G.; Gupta, K.J. Plant mitochondria: Source and target for nitric oxide. Mitochondrion 2014, 19, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Negrette-Guzmán, M.; García-Niño, W.R.; Tapia, E.; Zazueta, C.; Huerta-Yepez, S.; León-Contreras, J.C.; Hernández-Pando, R.; Aparicio-Trejo, O.E.; Madero, M.; Pedraza-Chaverri, J. Curcumin Attenuates Gentamicin-Induced Kidney Mitochondrial Alterations: Possible Role of a Mitochondrial Biogenesis Mechanism. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef]
- Guerrero-Beltrán, C.E.; Calderón-Oliver, M.; Martínez-Abundis, E.; Tapia, E.; Zarco-Márquez, G.; Zazueta, C.; Pedraza-Chaverri, J. Protective effect of sulforaphane against cisplatin-induced mitochondrial alterations and impairment in the activity of NAD(P)H: Quinone oxidoreductase 1 and γ glutamyl cysteine ligase: Studies in mitochondria isolated from rat kidney and in LLC-PK1 cells. Toxicol. Lett. 2010, 199, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Granados-Castro, L.F.; Rodríguez-Rangel, D.S.; Fernández-Rojas, B.; Leõn-Contreras, J.C.; Hernández-Pando, R.; Medina-Campos, O.N.; Eugenio-Pérez, D.; Pinzõn, E.; Pedraza-Chaverri, J. Curcumin prevents paracetamol-induced liver mitochondrial alterations. J. Pharm. Pharmacol. 2016, 68, 245–256. [Google Scholar] [CrossRef]
- Correa, F.; Buelna-Chontal, M.; Hernández-Reséndiz, S.; García-Niño, W.R.; Roldán, F.J.; Soto, V.; Silva-Palacios, A.; Amador, A.; Pedraza-Chaverrí, J.; Tapia, E.; et al. Curcumin maintains cardiac and mitochondrial function in chronic kidney disease. Free Radic. Biol. Med. 2013, 61, 119–129. [Google Scholar] [CrossRef]
- García-Niño, W.R.; Zazueta, C.; Tapia, E.; Pedraza-Chaverri, J. Curcumin attenuates cr(VI)-induced ascites and Changes in the activity of aconitase and F1F0 ATPase and the ATP content in rat liver mitochondria. J. Biochem. Mol. Toxicol. 2014, 28, 522–527. [Google Scholar] [CrossRef]
- Tapia, E.; Sánchez-Lozada, L.G.; García-Niño, W.R.; García, E.; Cerecedo, A.; García-Arroyo, F.E.; Osorio, H.; Arellano, A.; Cristóbal-García, M.; Loredo, M.L.; et al. Curcumin prevents maleate-induced nephrotoxicity: Relation to hemodynamic alterations, oxidative stress, mitochondrial oxygen consumption and activity of respiratory complex I. Free Radic. Res. 2014, 48, 1342–1354. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; de Bittencourt Brasil, F.; Fürstenau, C.R. Sulforaphane Promotes Mitochondrial Protection in SH-SY5Y Cells Exposed to Hydrogen Peroxide by an Nrf2-Dependent Mechanism. Mol. Neurobiol. 2018, 55, 4777–4787. [Google Scholar] [CrossRef]
- Mouzaoui, S.; Banerjee, S.; Djerdjouri, B. Low-dose curcumin reduced TNBS-associated mucin depleted foci in mice by scavenging superoxide anion and lipid peroxides, rebalancing matrix NO synthase and aconitase activities, and recoupling mitochondria. Inflammopharmacology 2020, 28, 949–965. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A. Sulforaphane: Its “Coming of Age” as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease. Oxidative Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The therapeutic potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic acid: Therapeutic potential through its antioxidant property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef]
- Vaiserman, A.; Koliada, A.; Zayachkivska, A.; Lushchak, O. Curcumin: A therapeutic potential in ageing-related disorders. PharmaNutrition 2020, 14, 100226. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Fragasso, M.; Nigro, F.; Platani, C.; Papa, R.; Beleggia, R.; Trono, D. Analysis of metabolic and mineral changes in response to salt stress in durum wheat (Triticum turgidum ssp. durum) genotypes, which differ in salinity tolerance. Plant Physiol. Biochem. 2018, 133, 57–70. [Google Scholar] [CrossRef]
- Brini, F.; Hanin, M.; Lumbreras, V.; Irar, S.; Pagès, M.; Masmoudi, K. Functional characterization of DHN-5, a dehydrin showing a differential phosphorylation pattern in two Tunisian durum wheat (Triticum durum Desf.) varieties with marked differences in salt and drought tolerance. Plant Sci. 2007, 172, 20–28. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Sairam, R.K.; Srivastava, G.C. Physiological and biochemical responses of hexaploid and tetraploid wheat to drought stress. J. Agron. Crop Sci. 2000, 185, 219–227. [Google Scholar] [CrossRef]
- Pastore, D.; Laus, M.N.; Di Fonzo, N.; Passarella, S. Reactive oxygen species inhibit the succinate oxidation-supported generation of membrane potential in wheat mitochondria. FEBS Lett. 2002, 516, 15–19. [Google Scholar] [CrossRef]
- Trono, D.; Laus, M.N.; Soccio, M.; Pastore, D. Transport pathways-proton motive force interrelationship in durum wheat mitochondria. Int. J. Mol. Sci. 2014, 15, 8186–8215. [Google Scholar] [CrossRef] [PubMed]
- Trono, D.; Laus, M.N.; Soccio, M.; Alfarano, M.; Pastore, D. Modulation of potassium channel activity in the balance of ROS and ATP production by durum wheat mitochondria—An amazing defense tool against hyperosmotic stress. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Pastore, D.; Trono, D.; Passarella, S. Substrate oxidation and ADP/ATP exchange in coupled durum wheat (Triticum durum Desf.) mitochondria. Plant Biosyst. 1999, 133, 219–228. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Laus, M.N.; Soccio, M.; Alfarano, M.; Pasqualone, A.; Lenucci, M.S.; Di Miceli, G.; Pastore, D. Different effectiveness of two pastas supplemented with either lipophilic or hydrophilic/phenolic antioxidants in affecting serum as evaluated by the novel Antioxidant/Oxidant Balance approach. Food Chem. 2017, 221. [Google Scholar] [CrossRef] [PubMed]
- Soccio, M.; Laus, M.N.; Alfarano, M.; Dalfino, G.; Panunzio, M.F.; Pastore, D. Antioxidant/Oxidant Balance as a novel approach to evaluate the effect on serum of long-term intake of plant antioxidant-rich foods. J. Funct. Foods 2018, 40, 778–784. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Laus, M.N.; Di Benedetto, N.A.; Caporizzi, R.; Tozzi, D.; Soccio, M.; Giuzio, L.; De Vita, P.; Flagella, Z.; Pastore, D. Evaluation of Phenolic Antioxidant Capacity in Grains of Modern and Old Durum Wheat Genotypes by the Novel QUENCHERABTS Approach. Plant Foods Hum. Nutr. 2015, 70, 207–214. [Google Scholar] [CrossRef]
- Moore, A.L.; Proudlove, M.O. Purification of plant mitochondria on silica sol gradients. Methods Enzymol. 1987, 148, 415–420. [Google Scholar] [CrossRef]
- Laus, M.N.; Soccio, M.; Trono, D.; Cattivelli, L.; Pastore, D. Plant Inner Membrane Anion Channel (PIMAC) function in plant mitochondria. Plant Cell Physiol. 2008, 49, 1039–1055. [Google Scholar] [CrossRef]
- Laus, M.N.; Soccio, M.; Trono, D.; Liberatore, M.T.; Pastore, D. Activation of the plant mitochondrial potassium channel by free fatty acids and acyl-CoA esters: A possible defence mechanism in the response to hyperosmotic stress. J. Exp. Bot. 2011, 62, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Soccio, M.; Laus, M.N.; Trono, D.; Pastore, D. A new simple fluorimetric method to assay cytosolic ATP content: Application to durum wheat seedlings to assess modulation of mitochondrial potassium channel and uncoupling protein activity under hyperosmotic stress. Biologia 2013, 68, 421–432. [Google Scholar] [CrossRef]
- Harris, D. Spectophotometric assay. In Spectophotometry and Spectrofluorimetry: A Practical Approach; Bashford, C.L., Harris, D., Eds.; IRL Press: Oxford, UK, 1987; pp. 59–61. [Google Scholar]
- Baumgart, M.; Bott, M. Biochemical characterisation of aconitase from Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC-Fluorescein) Assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Lucas-Abellán, C.; Mercader-Ros, M.T.; Zafrilla, M.P.; Gabaldón, J.A.; Núñez-Delicado, E. Comparative study of different methods to measure antioxidant activity of resveratrol in the presence of cyclodextrins. Food Chem. Toxicol. 2011, 49, 1255–1260. [Google Scholar] [CrossRef]
- Tumer, T.B.; Rojas-Silva, P.; Poulev, A.; Raskin, I.; Waterman, C. Direct and indirect antioxidant activity of polyphenol- and isothiocyanate-enriched fractions from moringa oleifera. J. Agric. Food Chem. 2015, 63, 1505–1513. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Mizgier, M.L.; Speisky, H.; Gotteland, M. Differential protective effects of quercetin, resveratrol, rutin and epigallocatechin gallate against mitochondrial dysfunction induced by indomethacin in Caco-2 cells. Chem. Biol. Interact. 2012, 195, 199–205. [Google Scholar] [CrossRef]
- Gao, C.; Chen, X.; Li, J.; Li, Y.; Tang, Y.; Liu, L.; Chen, S.; Yu, H.; Yao, P. Myocardial mitochondrial oxidative stress and dysfunction in intense exercise: Regulatory effects of quercetin. Eur. J. Appl. Physiol. 2014, 114, 695–705. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Hyland, R.A.; Rogers, P.J.; Higgins, V.J.; Myers, S.; Coorssen, J.R. Measuring hydrogen peroxide reduction using a robust, inexpensive, and sensitive method. J. Chem. Biol. 2012, 5, 143–150. [Google Scholar] [CrossRef]
- Kennedy, M.C.; Emptage, M.H.; Dreyer, J.L.; Beinert, H. The role of iron in the activation-inactivation of aconitase. J. Biol. Chem. 1983, 258, 11098–11105. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical | AC (μmol Trolox eq./μmol) | |
|---|---|---|
| TEAC | ORAC | |
| Ferulic acid | 1.52 ± 0.02 E | 5.18 ± 0.74 c |
| Apigenin | 2.74 ± 0.12 C | 8.46 ± 0.73 b |
| Quercetin | 4.99 ± 0.30 A | 8.71 ± 0.50 b |
| Curcumin | 2.30 ± 0.08 D | 8.79 ± 0.73 b |
| Resveratrol | 4.38 ± 0.14 B | 10.45 ± 1.16 a |
| Sulforaphane | N.D. | N.D. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laus, M.N.; Soccio, M. First Evidence of a Protective Effect of Plant Bioactive Compounds against H2O2-Induced Aconitase Damage in Durum Wheat Mitochondria. Antioxidants 2020, 9, 1256. https://doi.org/10.3390/antiox9121256
Laus MN, Soccio M. First Evidence of a Protective Effect of Plant Bioactive Compounds against H2O2-Induced Aconitase Damage in Durum Wheat Mitochondria. Antioxidants. 2020; 9(12):1256. https://doi.org/10.3390/antiox9121256
Chicago/Turabian StyleLaus, Maura N., and Mario Soccio. 2020. "First Evidence of a Protective Effect of Plant Bioactive Compounds against H2O2-Induced Aconitase Damage in Durum Wheat Mitochondria" Antioxidants 9, no. 12: 1256. https://doi.org/10.3390/antiox9121256
APA StyleLaus, M. N., & Soccio, M. (2020). First Evidence of a Protective Effect of Plant Bioactive Compounds against H2O2-Induced Aconitase Damage in Durum Wheat Mitochondria. Antioxidants, 9(12), 1256. https://doi.org/10.3390/antiox9121256