ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and other Chemical Compounds

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Preparation
2.2. Extraction Procedure
2.3. Physicochemical Analyses
2.4. Identification and Quantification of Phenolic Compounds by the LC-PDA-QTOF-ESI-MS and UPLC-PDA Methods
2.5. Quantification of Polymeric Procyanidins by the UPLC-PDA-FLMethod
2.6. Determination of Antioxidant and In Vitro Biological Activities
2.7. Antioxidant On-Line Profiling by HPLC-PDA Coupled with Post-Column Derivatization with ABTS
2.8. Statistical Analysis
3. Results and Discussion
3.1. Physiochemical Analysis
3.2. Polyphenol Compounds
3.3. Antioxidant and In Vitro Biological Activities
3.4. Antioxidant On-Line Profiling by HPLC-PDA Coupled with Post-Column Derivatization with ABTS
3.5. Agglomerative Hierarchical Clustering (AHC)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nahorska, A.; Dzwoniarska, M.; Thiem, B. Owoce pigwowca japońskiego (Chaenomeles japonica (Thunb.) Lindl. ex Spach) źródłem substancji biologicznie aktywnych. Postępy Fitoterapii 2014, 4, 239–246. [Google Scholar]
- Du, H.; Wu, J.; Li, H.; Zhong, P.-X.; Xu, Y.-J.; Li, C.-H.; Ji, K.-X.; Wang, L. Polyphenols and triterpenes from Chaenomeles fruits: Chemical analysis and antioxidant activities assessment. Food Chem. 2013, 141, 4260–4268. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Han, L.Y.; Zhang, H.; Xin, H.L. Chaenomeles speciosa: A review of chemistry and pharmacology. Biomed. Rep. 2014, 2, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Gorlach, S.; Wagner, W.; Podsędek, A.; Szewczyk, K.; Koziołkiewicz, M.; Dastych, J. Procyanidins from Japanese quince (Chaenomeles japonica) fruit induce apoptosis in human colon cancer Caco-2 cells in a degree of polymerization-dependent manner. Nutr. Cancer 2011, 63, 1348–1360. [Google Scholar] [CrossRef] [PubMed]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Hernández, F. Antidiabetic, anticholinesterase and antioxidant activity vs. terpenoids and phenolic compounds in selected new cultivars and hybrids of artichoke Cynara scolymus L. Molecules 2019, 24, 1222. [Google Scholar] [CrossRef] [PubMed]
- Wojdyło, A.; Nowicka, P.; Oszmiański, J.; Golis, T. Phytochemical compounds and biological effects of Actinidia fruits. J. Funct. Foods 2017, 30, 194–202. [Google Scholar] [CrossRef]
- Pijanowski, E.; Mrożewski, S.; Horubała, A.; Jarczyk, A. Technologia Produktów Owocowych i Warzywnych; Państwowe Wydawnictwa Rolnicze iLeśne: Warsaw, Poland, 1973; Volume 1, p. 634. [Google Scholar]
- Wojdyło, A.; Oszmiański, J.; Bielicki, P. Comparison of polyphenolic composition, antioxidant activity and PPO activity of some cultivars of quince (Cydonia oblonga Miller) fruits. J. Agric. Food Chem. 2013, 61, 2762–2772. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Bąbelewski, P. Phenolic and carotenoid profile of new goji cultivars and their anti-hyperglycemic, anti-aging and antioxidant properties. J. Funct. Foods 2018, 48, 632–642. [Google Scholar] [CrossRef]
- Chung, L.Y.; Soo, W.K.; Chan, K.Y.; Mustafa, M.R.; Goh, S.H.; Imiyabir, Z. Lipoxygenase inhibiting activity of some Malaysian plants. Pharm. Biol. 2009, 47, 1142–1148. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Lech, K.; Tkacz, K.; Nowicka, P. Influence of different drying methods on the quality of Japanese quince fruit. LWT 2019, 114, 108416. [Google Scholar] [CrossRef]
- Thomas, M.; Guillemin, F.; Guillon, F.; Thibault, J.-F. Pectins in the fruits of Japanese quince (Chaenomeles japonica). Carbohydr. Polym. 2003, 53, 361–372. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Satora, P.; Sroka, P.; Pogoń, P.; Machalica, J. Chaenomeles japonica, Cornus mas, Morus nigra fruits characteristics and their processing potential. J. Food Sci. Technol. 2014, 51, 3934–3941. [Google Scholar] [CrossRef]
- Lesińska, E. Charakterystyka skladu chemicznego owocow pigwowca i ocena ich technologicznej przydatnosci dla przetworstwa owocowo-warzywnego. In Zeszyty Naukowe Akademi Rolniczej im. H. Kołłątaja w Krakowie. Rozprawa Habilitacyjna; Wydawnictwo Akademii Rolniczej im. H. Kołłątaja: Kraków, Poland, 1986; p. 118. [Google Scholar]
- Rubinskienė, M.; Viškelis, P.; Viškelis, J.; Bobinaitė, R.; Shalkevich, M.; Pigul, M.; Urbonavičienė, D. Biochemical composition and antioxidant activity of Japanese quince (Chaenomeles japonica) fruit, their syrup and candied fruit slices. Sodininkystė ir Daržininkystė 2014, 33, 45–52. [Google Scholar]
- Ros, J.; Laencina, J.; Hellın, P.; Jordan, M.; Vila, R.; Rumpunen, K. Characterization of juice in fruits of different Chaenomeles species. LWT Food Sci. Technol. 2004, 37, 301–307. [Google Scholar] [CrossRef]
- Bieniasz, M.; Dziedzic, E.; Kaczmarczyk, E. The effect of storage and processing on vitamin C content in Japanese quince fruit. Folia Hortic. 2017, 29, 83–93. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Hellín, P.; Vila, R.; Jordán, M.J.; Laencina Sánchez, J.; Rumpunen, K.; Ros-García, J.M. Characteristics and composition of Chaenomeles fruit juice. In Japanese Quince—Potential Fruit Crop for Northern Europe; Rumpunen, K., Ed.; Swedish University of Agricultural Sciences: Alnarp, Sweden, 2003; pp. 127–140. [Google Scholar]
- Lesińska, E. Characteristics of sugars and acids in the fruits of East Asian quince. Die Nahr. 1987, 31, 763–765. [Google Scholar] [CrossRef]
- Lesińska, E.; Przybylski, R.; Eskin, M. Some Volatile and nonvolatile flavor components of the dwarf quince (Chaenomeles japonica). J. Food Sci. 1988, 53, 854–856. [Google Scholar] [CrossRef]
- Owczarek, K.; Hrabec, E.; Fichna, J.; Sosnowska, D.; Koziołkiewicz, M.; Szymański, J.; Lewandowska, U. Flavanols from Japanese quince (Chaenomeles japonica) fruit suppress expression of cyclooxygenase-2, metalloproteinase-9, and nuclear factor-kappaB in human colon cancer cells. Acta Biochim. Pol. 2017, 64, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, R.; Zhou, S.; Liu, W.; Liang, Y.; Zhao, Z.; Li, S.; Wang, X.; Wong, T.; Zhao, H. Chemical characterization and evaluation of the antioxidants in Chaenomeles fruits by an improved HPLC-TOF/MS coupled to an on-line DPPH-HPLC method. J. Environ. Sci. Health Part C 2018, 36, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Teleszko, M.; Wojdyło, A. Comparison of phenolic compounds and antioxidant potential between selected edible fruits and their leaves. J. Funct. Foods 2015, 14, 736–746. [Google Scholar] [CrossRef]
- Sun, J.; Liang, F.; Bin, Y.; Li, P.; Duan, C. Screening non-colored phenolics in red wines using liquid chromatography/ultraviolet and mass spectrometry/mass spectrometry libraries. Molecules 2007, 12, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Bravo, M.; Silva, S.; Coelho, A.; Boas, L.V.; Bronze, M. Analysis of phenolic compounds in Muscatel wines produced in Portugal. Anal. Chim. Acta 2006, 563, 84–92. [Google Scholar] [CrossRef]
- Stöggl, W.; Huck, C.; Bonn, G.K. Structural elucidation of catechin and epicatechin in sorrel leaf extracts using liquid-chromatography coupled to diode array-, fluorescence-, and mass spectrometric detection. J.Sep. Sci. 2004, 27, 524–528. [Google Scholar] [CrossRef]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsędek, A.; Koziołkiewicz, M.; Hrabec, E. Flavanols from Japanese quince (Chaenomeles japonica) fruit inhibit human prostate and breast cancer cell line invasiveness and cause favorable changes in Bax/Bcl-2 mRNA ratio. Nutr. Cancer 2013, 65, 273–285. [Google Scholar] [CrossRef]
- Wojdyło, A.; Figiel, A.; Lech, K.; Nowicka, P.; Oszmiański, J. Effect of convective and vacuum–microwave drying on the bioactive compounds, color, and antioxidant capacity of sour cherries. Food Bioprocess Technol. 2014, 7, 829–841. [Google Scholar] [CrossRef]
- Tkacz, K.; Wojdyło, A.; Nowicka, P.; Turkiewicz, I.; Golis, T. Characterization in vitro potency of biological active fractions of seeds, skins and flesh from selected Vitis vinifera L. cultivars and interspecific hybrids. J. Funct. Foods 2019, 56, 353–363. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Report on Diabetes. 2016. Available online: https://www.who.int/diabetes/global-report/en/ (accessed on 10 October 2019).
- Miao, J.; Li, X.; Zhao, C.; Gao, X.; Wang, Y.; Cheng, K.; Gao, W. Solvents effect on active chemicals and activities of antioxidant, anti-α-glucosidase and inhibit effect on smooth muscle contraction of isolated rat jejunum of Chaenomeles speciosa. J. Funct. Foods 2018, 40, 146–155. [Google Scholar] [CrossRef]
- Miao, J.; Zhao, C.; Li, X.; Chen, X.; Mao, X.; Huang, H.; Wang, T.; Gao, W. Chemical composition and bioactivities of two common Chaenomeles fruits in China: Chaenomeles speciosa and Chaenomeles sinensis. J. Food Sci. 2016, 81, H2049–H2058. [Google Scholar] [CrossRef]
- Nowicka, P.; Wojdyło, A.; Laskowski, P. Inhibitory potential against digestive enzymes linked to obesity and type 2 diabetes and content of bioactive compounds in 20 cultivars of the peach fruit grown in Poland. Plant Foods Hum. Nutr. 2018, 73, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Honarvar, N.M.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Sangsari, G.T.; Rad, B.H.; Muench, G. Molecular anti-inflammatory mechanisms of retinoids and carotenoids in Alzheimer’s disease: A review of current evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Sancheti, S.; Sancheti, S.; Seo, S.-Y. Antidiabetic and antiacetylcholinesterase effects of ethyl acetate fraction of Chaenomeles sinensis (Thouin) Koehne fruits in streptozotocin-induced diabetic rats. Exp. Toxicol. Pathol. 2013, 65, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Orafaie, A.; Matin, M.M.; Sadeghian, H. The importance of 15-lipoxygenase inhibitors in cancer treatment. Cancer Metastasis Rev. 2018, 37, 397–408. [Google Scholar] [CrossRef]
- Burnaz, N.A.; Küçük, M.; Akar, Z. An on-line HPLC system for detection of antioxidant compounds in some plant extracts by comparing three different methods. J. Chromatogr. B 2017, 1052, 66–72. [Google Scholar] [CrossRef]
- Wood, J.E.; Senthilmohan, S.T.; Peskin, A.V. Antioxidant activity of procyanidin-containing plant extracts at different pHs. Food Chem. 2002, 77, 155–161. [Google Scholar] [CrossRef]
- Raudone, L.; Raudonis, R.; Liaudanskas, M.; Viskelis, J.; Pukalskas, A.; Janulis, V. Phenolic profiles and contribution of individual compounds to antioxidant activity of apple powders. J. Food Sci. 2016, 81, C1055–C1061. [Google Scholar] [CrossRef]
- Çelik, S.E.; Özyürek, M.; Güçlü, K.; Apak, R. Solvent effects on the antioxidant capacity of lipophilic and hydrophilic antioxidants measured by CUPRAC, ABTS/persulphate and FRAP methods. Talanta 2010, 81, 1300–1309. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Species | Chaenomeles × Superba | ||||||||||
| Cultivar | Crimson and Gold | Texas Scarlet | Nicoline | Andenken an Karl Ramcke | Pink Lady | Colour Trail | Flocon Rose | Hollandia | Jet Trail | Wild | Cameo |
| dry matter (%) | 13.46 ± 0.20h | 16.21 ± 0.21d | 17.51 ± 0.30bc | 17.13 ± 0.13c | 14.76 ± 0.20f | 13.82 ± 0.20gh | 11.95 ± 0.25ij | 15.58 ± 0.18e | 13.92 ± 0.22gh | 20.40 ± 0.20a | 12.31 ± 0.21i |
| ash content (%) | 0.32 ± 0.12j | 0.43 ± 0.33fghi | 0.48 ± 0.15defg | 0.53 ± 0.13bcdef | 0.64 ± 0.24a | 0.41 ± 0.11ghij | 0.46 ± 0.15fg | 0.41 ± 0.10ghij | 0.51 ± 0.11cdefg | 0.57 ± 0.14abcde | 0.34 ± 0.44ij |
| SSC (°Brix) | 6.8 ± 0.1h | 11.6 ± 0.0b | 10.4 ± 0.1c | 7.9 ± 0.1f | 9.4 ± 0.0d | 9.5 ± 0.1d | 7.3 ± 0.0g | 10.3 ± 0.1c | 5.9 ± 0.0j | 12.1 ± 0.1a | 7.2 ± 0.1g |
| pectin (%) | 1.23 ± 0.10def | 1.57 ± 0.10abc | 1.62 ± 0.12ab | 1.69 ± 0.09ab | 1.10 ± 0.10efg | 0.99 ± 0.11efgh | 0.68 ± 0.18ij | 0.98 ± 0.08fghi | 1.41 ± 0.10bcd | 1.72 ± 0.20a | 0.71 ± 0.10hij |
| TA (g of malic acid/100 g of fw) | 4.27 ± 0.13ef | 4.60 ± 0.10cde | 4.66 ± 0.49cde | 5.30 ± 0.15b | 4.64 ± 0.49cde | 5.20 ± 0.12bc | 4.20 ± 0.10ef | 4.25 ± 0.15ef | 3.45 ± 0.10gh | 6.16 ± 0.16a | 4.66 ± 0.10cde |
| pH | 2.927 ± 0.01d | 2.801 ± 0.00h | 2.782 ± 0.01hi | 2.738 ± 0.01j | 2.772 ± 0.00i | 2.897 ± 0.01e | 2.855 ± 0.02g | 2.842 ± 0.00g | 2.975 ± 0.01ab | 2.713 ± 0.01j | 2.892 ± 0.01ef |
| L-ascorbic acid (mg/100 g of fw) | 40.83 ± 0.55jk | 175.32 ± 0.68ab | 134.38 ± 0.23cd | 144.17 ± 0.50c | 111.81 ± 0.88def | 47.86 ± 0.74ijk | 110.99 ± 0.29ef | 195.05 ± 0.30a | 70.96 ± 0.55gh | 143.09 ± 1.00c | 70.29 ± 0.67ghi |
| Sugars (g/100g fw) | |||||||||||
| xylose | 0.03 ± 0.00b | nd | nd | 0.05 ± 0.00a | nd | nd | nd | nd | nd | nd | nd |
| fructose | 0.58 ± 0.02g | 1.80 ± 0.35a | 0.60 ± 0.02g | 0.51 ± 0.03gh | 1.34 ± 0.13bcd | 1.10 ± 0.07def | 0.94 ± 0.02f | 1.62 ± 0.03ab | 0.12 ± 0.02i | 1.31 ± 0.07cde | 1.18 ± 0.04def |
| sorbitol | 0.30 ± 0.00jk | 0.95 ± 0.16b | 0.56 ± 0.00fgh | 0.75 ± 0.03cde | 0.72 ± 0.06def | 0.72 ± 0.04def | 0.47 ± 0.01hij | 0.79 ± 0.01bcde | 0.22 ± 0.01k | 1.40 ± 0.05a | 0.37 ± 0.00ijk |
| glucose | 0.29 ± 0.00ghi | 1.23 ± 0.26a | 0.44 ± 0.02efg | 0.29 ± 0.01ghi | 0.91 ± 0.07bc | 0.88 ± 0.06bc | 0.51 ± 0.01def | 0.91 ± 0.02bc | 0.10 ± 0.01i | 0.85 ± 0.04c | 0.93 ± 0.03bc |
| total | 1.20 ± 0.02ij | 3.98 ± 0.37a | 1.60 ± 0.04hi | 1.60 ± 0.27hi | 2.97 ± 0.26cde | 2.70 ± 0.17def | 1.92 ± 0.04gh | 3.32 ± 0.06bcd | 0.44 ± 0.04k | 3.56 ± 0.17abc | 2.48 ± 0.08fg |
| fructose:glucose ratio | 2.0 | 1.5 | 1.4 | 1.8 | 1.5 | 1.2 | 1.8 | 1.8 | 1.2 | 1.5 | 1.3 |
| Organic Acids (g/kg fw) | |||||||||||
| oxalic | 0.06 ± 0.00e | 0.70 ± 0.05b | 0.24 ± 0.06bcde | 0.27 ± 0.00bcde | 0.18 ± 0.02bcde | 0.23 ± 0.02bcde | 0.20 ± 0.03bcde | 0.59 ± 0.09a | 0.23 ± 0.03bcde | 0.28 ± 0.04bcd | 0.17 ± 0.02bcde |
| maleic | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | nd | 0.01 ± 0.00a | 0.01 ± 0.00a |
| citric | 0.78 ± 0.07de | 0.90 ± 0.12de | 0.41 ± 0.02fg | 0.75 ± 0.05de | 0.66 ± 0.09ef | 1.20 ± 0.02ab | 1.29 ± 0.10ab | 1.35 ± 0.09ab | 0.31 ± 0.02g | 1.42 ± 0.07a | 0.91 ± 0.02cde |
| malic | 62.52 ± 1.33efg | 56.12 ± 1.00gh | 48.61 ± 1.97hi | 64.34 ± 3.54ef | 64.87 ± 3.43ef | 81.44 ± 2.12abc | 62.14 ± 1.70efg | 57.64 ± 0.39fg | 38.83 ± 1.47j | 88.75 ± 1.32a | 65.94 ± 2.52e |
| quinic | 10.51 ± 0.16fg | 7.52 ± 0.12ij | 14.50 ± 0.32bc | 12.37 ± 0.15de | 9.91 ± 0.04fgh | 10.70 ± 0.48efg | 14.52 ± 0.18bc | 17.28 ± 1.39a | 11.50 ± 0.79def | 17.15 ± 0.29a | 8.70 ± 0.04hi |
| shikimic | 0.11 ± 0.01fg | 0.14 ± 0.01fg | 0.73 ± 0.05de | 0.91 ± 0.07cd | 0.12 ± 0.01fg | 0.07 ± 0.00g | 0.22 ± 0.01efg | 0.17 ± 0.00efg | 0.65 ± 0.00def | 2.70 ± 0.06a | 0.10 ± 0.00fg |
| total | 73.99 ± 1.57de | 65.07 ± 1.29ef | 64.50 ± 2.37ef | 78.65 ± 3.82d | 75.75 ± 3.55d | 93.65 ± 2.64c | 78.39 ± 1.96d | 77.04 ± 1.96d | 51.52 ± 2.26g | 110.31 ± 1.12a | 75.82 ± 2.60d |
| sugars:acids ratio | 0.2 | 0.6 | 0.2 | 0.2 | 0.4 | 0.3 | 0.2 | 0.4 | 0.1 | 0.3 | 0.3 |
| Species | Chaenomeles Japonica | Chaenomeles Speciosa | |||||||||
| Cultivar | Cido | Red Joy | Wild #1 | Wild #2 | n1 (New) | Nivalis | Rubra | Simonii | |||
| dry matter (%) | 13.45 ± 0.15h | 10.95 ± 0.15k | 17.76 ± 0.14b | 14.35 ± 0.15fg | 10.09 ± 0.11l | 17.04 ± 0.24c | 11.95 ± 0.15ij | 11.71 ± 0.20j | |||
| ash content (%) | 0.47 ± 0.23efg | 0.35 ± 0.15hij | 0.61 ± 0.31abc | 0.58 ± 0.22abcd | 0.41 ± 0.21ghij | 0.43 ± 0.33fghi | 0.45 ± 0.15fgh | 0.63 ± 0.23ab | |||
| SSC (°Brix) | 8.4 ± 0.0e | 6.3 ± 0.1i | 10.3 ± 0.0c | 8.6 ± 0.0e | 6.3 ± 0.1i | 10.5 ± 0.0c | 6.6 ± 0.0h | 5.8 ± 0.1j | |||
| pectin (%) | 0.76 ± 0.16hij | 0.95 ± 0.15fghij | 1.08 ± 0.18efg | 0.90 ± 0.10ghij | 0.65 ± 0.15j | 1.29 ± 0.09cde | 0.85 ± 0.15ghij | 0.88 ± 0.18ghij | |||
| TA (g of malic acid/100 g of fw) | 3.11 ± 0.11h | 4.90 ± 0.10bcd | 5.50 ± 0.15b | 5.32 ± 0.12b | 3.97 ± 0.10fg | 5.44 ± 0.14b | 4.58 ± 0.12de | 3.45 ± 0.10gh | |||
| pH | 2.867 ± 0.01fg | 2.941 ± 0.01cd | 2.965 ± 0.01bc | 2.966 ± 0.01bc | 2.843 ± 0.00g | 2.862 ± 0.00g | 2.992 ± 0.01ab | 2.994 ± 0.00a | |||
| L-ascorbic acid (mg/100 g of fw) | 132.33 ± 0.35cde | 57.82 ± 0.23hij | 101.72 ± 0.21f | 114.13 ± 0.21def | 91.19 ± 0.57fg | 154.97 ± 0.33bc | 30.26 ± 0.20k | 53.6 ± 0.09hij | |||
| Sugars (g/100 g fw) | |||||||||||
| xylose | nd | nd | nd | nd | nd | nd | nd | nd | |||
| fructose | 1.57 ± 0.01abc | 0.18 ± 0.02i | 1.11 ± 0.04def | 1.02 ± 0.01ef | 0.62 ± 0.03g | 1.70 ± 0.02a | 0.24 ± 0.01hi | 0.25 ± 0.02hi | |||
| sorbitol | 0.66 ± 0.01efg | 0.25 ± 0.02k | 1.38 ± 0.13a | 0.92 ± 0.12bc | 0.33 ± 0.02jk | 0.88 ± 0.00bcd | 0.53 ± 0.03ghi | 0.25 ± 0.02k | |||
| glucose | 1.07 ± 0.03ab | 0.17 ± 0.01i | 0.65 ± 0.02d | 0.62 ± 0.02de | 0.42 ± 0.01fgh | 1.21 ± 0.01a | 0.22 ± 0.00hi | 0.15 ± 0.01i | |||
| total | 3.30 ± 0.05bcd | 0.60 ± 0.04jk | 3.14 ± 0.20cde | 2.56 ± 0.10ef | 1.37 ± 0.07hi | 3.79 ± 0.03ab | 0.99 ± 0.04ijk | 0.65 ± 0.04jk | |||
| fructose:glucose ratio | 1.5 | 1.1 | 1.7 | 1.6 | 1.5 | 1.4 | 1.1 | 1.6 | |||
| Organic Acids (g/kg fw) | |||||||||||
| oxalic | 0.13 ± 0.02cde | 0.20 ± 0.00bcde | 0.17 ± 0.05bcde | 0.25 ± 0.01bcde | 0.65 ± 0.25a | 0.26 ± 0.00bcde | 0.09 ± 0.01de | 0.32 ± 0.08bc | |||
| maleic | nd | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | 0.01 ± 0.00a | nd | 0.01 ± 0.00a | 0.01 ± 0.00a | |||
| citric | 0.30 ± 0.06g | 1.23 ± 0.10ab | 1.35 ± 0.04ab | 1.33 ± 0.05ab | 0.94 ± 0.17cd | 1.16 ± 0.09bc | 0.81 ± 0.07de | 0.75 ± 0.11de | |||
| malic | 32.08 ± 6.19j | 79.51 ± 3.82bc | 86.14 ± 1.30ab | 86.04 ± 0.78ab | 56.20 ± 1.79gh | 74.30 ± 3.99cd | 67.02 ± 1.20de | 47.93 ± 1.74i | |||
| quinic | 9.05 ± 0.76ghi | 9.15 ± 0.36ghi | 14.22 ± 0.80bc | 15.10 ± 0.56b | 13.09 ± 0.20cd | 16.89 ± 1.06a | 6.21 ± 0.02j | 12.33 ± 0.43de | |||
| shikimic | 0.08 ± 0.01g | 1.12 ± 0.08bcd | 1.34 ± 0.56bc | 0.72 ± 0.53de | 0.12 ± 0.01fg | 1.48 ± 0.12b | 1.17 ± 0.05bcd | 0.10 ± 0.01fg | |||
| total | 41.64 ± 7.04h | 91.21 ± 4.36c | 103.23 ± 2.58ab | 103.47 ± 1.81ab | 71.01 ± 1.52de | 94.09 ± 5.26bc | 75.30 ± 1.30d | 61.45 ± 2.37f | |||
| sugars:acids ratio | 0.8 | 0.1 | 0.3 | 0.2 | 0.2 | 0.4 | 0.1 | 0.1 | |||
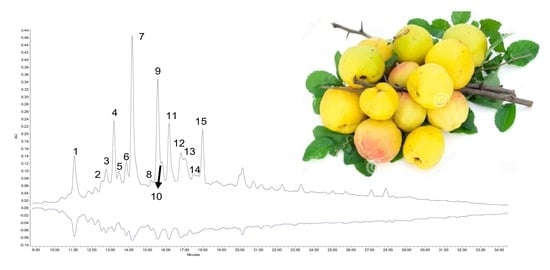
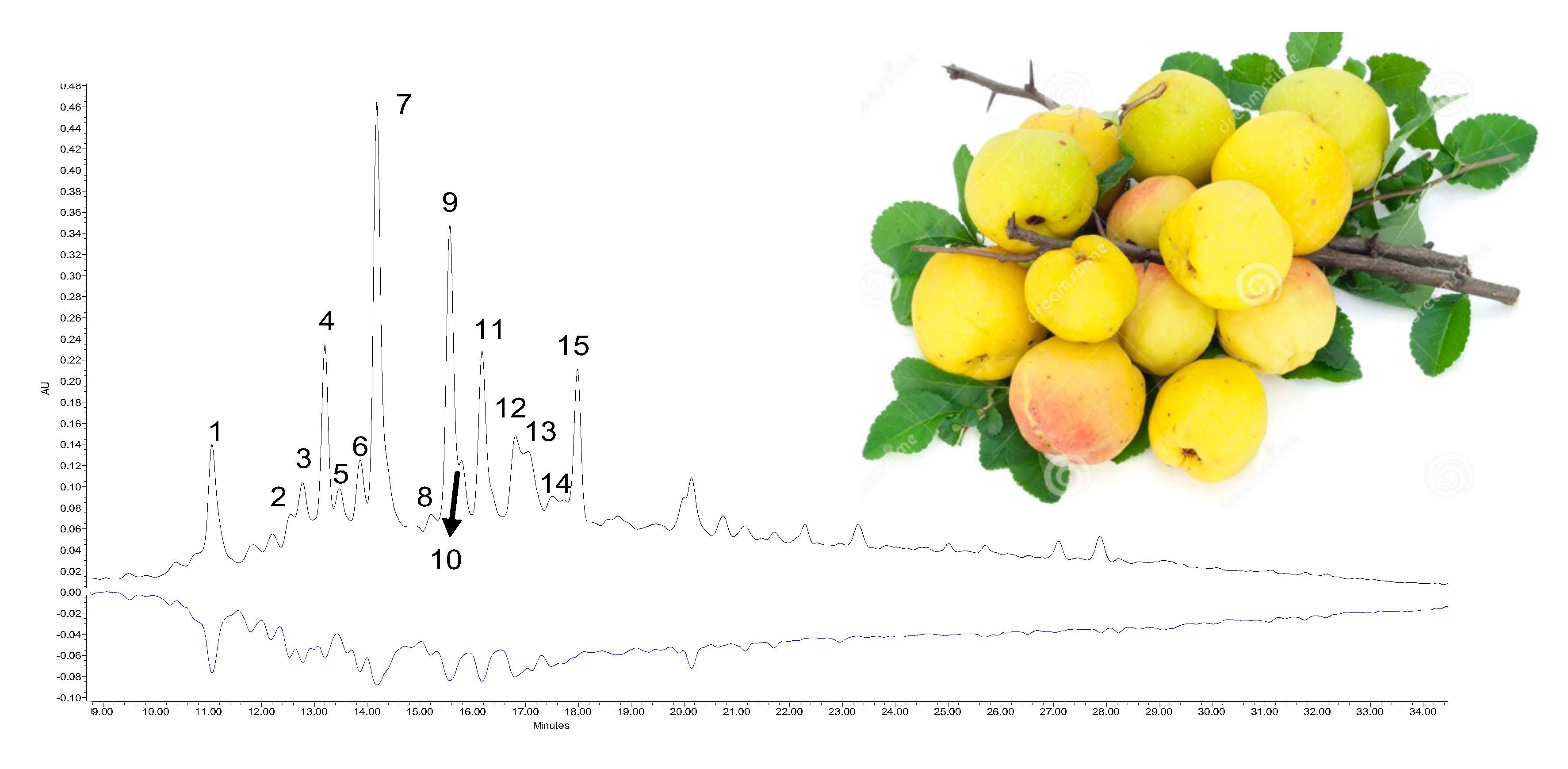
| Peak | Compound | Rt (min) | λmax (nm) | Molecular Formula | MS [M-H]− (m/z) | MS/MS (m/z) |
|---|---|---|---|---|---|---|
| 1 | Procyanidin B3 | 11.09 | 280 | C30H26O12 | 577.13 | 425.08/451.00/407.05/289.05 |
| 2 | (+)-Catechin | 12.53 | 240/280 | C15H13O6 | 289.06 | 245.06/205.03/125.01 |
| 3 | Procyanidin trimer | 12.79 | 280 | C45H37O18 | 865.21 | 577.13/425.08/289.06 |
| 4 | 5-O-Caffeoylquinic acid (chlorogenic) | 13.20 | 246/326 | C22H27O14 | 353.08 | 191.04 |
| 5 | 4-O-Caffeoylquinic acid (cryptochlorogenic) | 13.33 | 246/326 | C22H27O14 | 353.08 | 191.04 |
| 6 | Procyanidin trimer | 13.84 | 280 | C45H37O18 | 865.21 | 577.13/425.08/289.06 |
| 7 | Procyanidin B2 | 14.19 | 280 | C30H26O12 | 577.13 | 425.08/451.00/407.05/289.05 |
| 8 | Procyanidin dimer | 15.21 | 280 | C30H26O12 | 577.13 | 425.08/289.05 |
| 9 | (−)-Epicatechin | 15.58 | 240/280 | C15H13O6 | 289.06 | 245.06/205.03/125.01 |
| 10 | Procyanidin dimer | 15.80 | 280 | C30H26O12 | 577.13 | 425.08/289.05 |
| 11 | Procyanidin C1 | 16.18 | 280 | C45H37O18 | 865.21 | 577.13/289.06/245.06/125.01 |
| 12 | Procyanidin tetramer | 16.79 | 280 | C60H49O24 | 1153.3 | 865.21/576.12/289.05 |
| 13 | Procyanidin tetramer | 17.00 | 280 | C60H49O24 | 1153.3 | 865.21/576.12/289.05 |
| 14 | Procyanidin dimer | 17.20 | 280 | C30H26O12 | 577.13 | 425.08/289.05 |
| 15 | Procyanidin dimer | 17.92 | 280 | C30H26O12 | 577.13 | 425.08/289.05 |
| Chaenomeles × Superba | ||||||||||||||||||
| Peak no | Crimson and Gold | Texas Scarlet | Nicoline | Andenken an Karl Ramcke | Pink Lady | Colour Trail | Flocon Rose | Hollandia | Jet Trail | Wild | Cameo | |||||||
| Phenolic acids | ||||||||||||||||||
| 4 | 0.43 ± 0.15ef | 1.68 ± 0.20bc | 1.34 ± 0.21cd | 1.35 ± 0.10cd | 0.79 ± 0.13de | 0.23 ± 0.08ef | 1.65 ± 0.27bc | 1.08 ± 0.11cd | 1.12 ± 0.19cd | 3.04 ± 0.21a | 3.05 ± 0.15a | |||||||
| 5 | 0.46 ± 0.12bc | 0.35 ± 0.10bc | 0.04 ± 0.00c | nd | nd | 0.36 ± 0.08bc | 0.41 ± 0.22bc | 0.45 ± 0.11bc | 0.06 ± 0.00c | nd | 0.25 ± 0.14bc | |||||||
| Sum | 0.89 ± 0.15ef | 2.03 ± 0.20bc | 1.38 ± 0.00de | 1.35 ± 0.31de | 0.79 ± 0.20ef | 0.59 ± 0.10fg | 2.06 ± 0.33bc | 1.53 ± 0.18cd | 1.18 ± 0.27def | 3.04 ± 0.11a | 3.30 ± 0.19a | |||||||
| Flavan-3-ols | ||||||||||||||||||
| 1 | 0.28 ± 0.08i | 2.86 ± 0.22a | 1.85 ± 0.21cde | 1.47 ± 0.18efg | 0.65 ± 0.09hi | 1.00 ± 0.12gh | 2.02 ± 0.44bcde | 1.66 ± 0.36def | 1.76 ± 0.15cde | 1.75 ± 0.27cde | 2.75 ± 0.22a | |||||||
| 2 | 0.27 ± 0.10de | 0.29 ± 0.11cde | 0.42 ± 0.08bcde | 0.37 ± 0.10cde | 0.42 ± 0.15bcde | 0.75 ± 0.15abcde | 0.62 ± 0.22abcde | 0.69 ± 0.19abcde | 1.07 ± 0.08a | 0.86 ± 0.31abcd | 0.86 ± 0.28abcd | |||||||
| 3 | nd | 2.86 ± 0.44a | 2.64 ± 0.39a | 2.40 ± 0.30ab | nd | 0.40 ± 0.18f | 1.07 ± 0.19de | 0.88 ± 0.24def | 0.94 ± 0.09def | 1.16 ± 0.12de | 1.97 ± 0.33bc | |||||||
| 6 | 1.59 ± 0.18c | 2.33 ± 0.28b | 3.00 ± 0.30a | 2.28 ± .21b | 1.03 ± 0.18cde | 0.87 ± 0.22de | 0.91 ± 0.10de | 1.48 ± 0.19cd | 2.73 ± 0.28ab | 1.46 ± 0.17cd | 1.39 ± 0.19cde | |||||||
| 7 | 10.92 ± 1.21d | 13.40 ± 1.30c | 18.16 ± 1.02a | 14.19 ± 1.00b | 5.53 ± 0.50i | 7.90 ± 0.67f | 5.74 ± 0.78hi | 9.29 ± 0.62e | 14.72 ± 1.05b | 9.60 ± 0.77e | 8.24 ± 0.46f | |||||||
| 8 | 1.27 ± 0.62cde | 2.59 ± 0.80a | 2.55 ± 0.55a | 1.84 ± 0.42bc | 1.34 ± 0.42cde | 1.28 ± 0.33cde | 1.20 ± 0.27de | 1.34 ± 0.19cde | 2.16 ± 0.55ab | 1.17 ± 0.31def | 1.42 ± 0.41cd | |||||||
| 9 | 6.8 ± 0.99bc | 5.03 ± 0.55d | 6.99 ± 0.89b | 7.68 ± 0.77a | 2.35 ± 0.60gh | 5.02 ± 0.63d | 2.79 ± 0.49g | 4.35 ± 0.55f | 6.30 ± 0.68c | 4.59 ± 0.70def | 4.93 ± 0.66def | |||||||
| 10 | nd | 0.91 ± 0.10a | 0.48 ± 0.25abc | nd | 0.23 ± 0.11c | nd | 0.51 ± 0.21abc | 0.65 ± 0.10abc | 0.65 ± 0.10abc | nd | 0.79 ± 0.13abc | |||||||
| 11 | 3.90 ± 0.54de | 5.88 ± 0.52c | 7.58 ± 1.12a | 6.71 ± 0.66b | 1.87 ± 0.28ij | 3.05 ± 0.65fgh | 3.16 ± 0.33fg | 4.12 ± 0.33de | 5.84 ± 0.74c | 4.31 ± 0.56gh | 4.20 ± 0.60de | |||||||
| 12 | 2.39 ± 0.10fgh | 3.78 ± 0.25bc | 4.36 ± 0.85ab | 3.61 ± 0.33cd | 1.29 ± 0.00jk | 1.81 ± 0.31hij | 2.26 ± 0.55gh | 2.41 ± 0.33fgh | 3.07 ± 0.59de | 2.05 ± 0.27hi | 2.05 ± 0.55ghi | |||||||
| 13 | 4.28 ± 0.87d | 6.03 ± 0.54bc | 7.12 ± 0.42a | 5.81 ± 0.77c | 2.68 ± 0.28gh | 3.70 ± 0.44de | 2.44 ± 0.36ghi | 3.49 ± 0.50e | 6.60 ± 0.45ab | 3.38 ± 0.22ef | 2.87 ± 0.85fg | |||||||
| 14 | 0.61 ± 0.55fgh | 1.58 ± 0.30c | 2.33 ± 0.54b | 1.40 ± 0.26cde | 0.62 ± 0.40fgh | nd | 0.93 ± 0.58defgh | 0.90 ± 0.63efgh | 1.14 ± 0.28cdef | 1.12 ± 0.24cdefg | 0.67 ± 0.00fgh | |||||||
| 15 | 2.80 ± 0.60c | 4.67 ± 0.47a | 1.85 ± 0.32efg | 1.49 ± 0.40fgh | 0.93 ± 0.35hi | 0.48 ± 0.09ij | 2.20 ± 0.45cde | 3.55 ± 0.27b | 2.08 ± 0.65def | 1.17 ± 0.33h | 1.39 ± 0.14gh | |||||||
| Sum | 35.11 ± 1.18d | 52.21 ± 1.33b | 59.33 ± 2.15a | 49.25 ± 1.22c | 18.94 ± 1.66j | 26.26 ± 0.99hi | 25.85 ± 1.15hi | 34.81 ± 1.66de | 49.04 ± 1.44c | 32.62 ± 2.03efg | 33.53 ± 1.88def | |||||||
| Polymeric procyanidins | 51.73 ± 0.99g | 100.47 ± 1.33b | 109.67 ± 1.11a | 88.37 ± 1.15c | 74.33 ± 2.33d | 34.60 ± 1.21k | 48.46 ± 1.65hi | 63.69 ± 1.35f | 73.53 ± 0.87d | 68.15 ± 0.98e | 54.04 ± 2.22g | |||||||
| DP | 2.91 | 3.77 | 3.98 | 3.44 | 4.25 | 2.43 | 3.41 | 3.45 | 3.24 | 3.80 | 3.35 | |||||||
| Total | 87.73 ± 2.22j | 154.71 ± 3.00b | 170.38 ± 1.25a | 138.97 ± 1.98c | 94.06 ± 1.98h | 61.45 ± 1.24m | 76.37 ± 1.56k | 100.03 ± 1.54f | 123.75 ± 1.88d | 103.81 ± 2.22e | 90.87 ± 1.78i | |||||||
| Peak no | Chaenomeles Japonica | Chaenomeles Speciosa | ||||||||||||||||
| Cido | Red Joy | Wild #1 | Wild #2 | n1 (New) | Nivalis | Rubra | Simonii | |||||||||||
| Phenolic acids | ||||||||||||||||||
| 4 | 0.09 ± 0.02f | 1.07 ± 0.17cd | 0.31 ± 0.15ef | 1.15 ± 0.10cd | 2.12 ± 0.20b | 2.05 ± 0.26b | 1.29 ± 0.15cd | 0.25 ± 0.05ef | ||||||||||
| 5 | 0.06 ± 0.01c | 0.04 ± 0.01c | 0.70 ± 0.22b | nd | 0.28 ± 0.16bc | 0.19 ± 0.10c | nd | 2.80 ± 0.33a | ||||||||||
| Sum | 0.15 ± 0.05g | 1.11 ± 0.21def | 1.01 ± 0.06def | 1.15 ± 0.15def | 2.40 ± 0.17b | 2.24 ± 0.25b | 1.29 ± 0.10de | 3.05 ± 0.44a | ||||||||||
| Flavan-3-ols | ||||||||||||||||||
| 1 | 0.38 ± 0.10i | 1.88 ± 0.19cde | 0.48 ± 0.11hi | 1.09 ± 0.17fgh | 2.29 ± 0.38abc | 2.08 ± 0.30bcd | 2.51 ± 0.50ab | 0.37 ± 0.14i | ||||||||||
| 2 | 0.39 ± 0.13cde | 0.71 ± 0.19abcde | 0.22 ± 0.13e | 0.56 ± 0.14abcde | 0.86 ± 0.21abc | 0.69 ± 0.38abcde | 0.99 ± 0.39ab | 0.81 ± 0.25abcd | ||||||||||
| 3 | nd | 1.04 ± 0.24de | nd | 0.75 ± 0.28ef | 1.20 ± 0.19de | 1.37 ± 0.26d | 1.40 ± 0.30cd | nd | ||||||||||
| 6 | 2.23 ± 0.15b | 0.84 ± 0.20e | 1.17 ± 0.26cde | 0.91 ± 0.15de | 1.34 ± 0.08cde | 1.09 ± 0.33cde | 1.13 ± 0.16cde | 2.32 ± 0.47b | ||||||||||
| 7 | 11.40 ± 0.99d | 3.39 ± 0.96j | 6.19 ± 0.54h | 5.36 ± 0.66i | 8.40 ± 0.72f | 7.15 ± 0.60g | 3.80 ± 0.58j | 8.49 ± 0.50f | ||||||||||
| 8 | 1.57 ± 0.50bcd | 0.48 ± 0.33g | 1.09 ± 0.44defg | 0.77 ± 0.20efg | 1.57 ± 0.00bcd | 1.11 ± 0.24def | 0.56 ± 0.10fg | nd | ||||||||||
| 9 | 4.37 ± 0.22f | 1.77 ± 0.11h | 4.40 ± 0.43ef | 2.31 ± 0.39gh | 2.92 ± 0.18g | 2.88 ± 0.52g | 1.97 ± 0.12h | 4.99 ± 0.12de | ||||||||||
| 10 | 0.36 ± 0.09abc | 0.41 ± 0.20abc | 0.32 ± 0.15bc | nd | 0.53 ± 0.22abc | 0.42 ± 0.29abc | 0.43 ± 0.08abc | 0.83 ± 0.00ab | ||||||||||
| 11 | 4.05 ± 0.61de | 1.54 ± 0.88j | 2.61 ± 0.71gh | 2.43 ± 0.70hi | 3.94 ± 0.59de | 3.63 ± 0.20ef | 1.75 ± 0.55j | 2.79 ± 0.80gh | ||||||||||
| 12 | 0.49 ± 0.14l | 0.39 ± 0.00l | 3.21 ± 0.27cde | 1.17 ± 0.40k | 2.67 ± 0.00efg | 4.68 ± 0.33a | 1.58 ± 0.22ijk | 2.96 ± 0.47ef | ||||||||||
| 13 | 2.41 ± 0.35ghi | 0.89 ± 0.61j | 2.16 ± 0.33hi | 1.91 ± 0.54i | 3.45 ± 0.96ef | 2.86 ± 0.46fg | 0.82 ± 0.28j | 2.55 ± 0.27gh | ||||||||||
| 14 | 4.74 ± 0.19a | 0.87 ± 0.18efgh | 0.56 ± 0.11fghi | 0.54 ± 0.27ghi | 0.72 ± 0.33fgh | 1.50 ± 0.21cd | 0.38 ± 0.20hi | 0.39 ± 0.15hi | ||||||||||
| 15 | 1.94 ± 0.30efg | 0.27 ± 0.08j | 2.64 ± 0.33cd | 0.45 ± 0.24ij | 1.15 ± 0.51h | 2.42 ± 0.36cde | 0.31 ± 0.36j | 1.41 ± 0.22gh | ||||||||||
| Sum | 34.33 ± 1.54de | 14.48 ± 1.11k | 25.05 ± 2.29i | 18.25 ± 1.55j | 31.04 ± 1.43g | 31.88 ± 2.00fg | 17.63 ± 1.55j | 27.10 ± 1.48h | ||||||||||
| Polymeric procyanidins | 67.48 ± 1.99e | 40.08 ± 1.64j | 37.37 ± 1.00jk | 51.35 ± 1.70gh | 62.31 ± 1.46f | 90.83 ± 2.15c | 38.92 ± 1.02j | 45.83 ± 0.99i | ||||||||||
| DP | 4.18 | 2.72 | 3.16 | 3.87 | 3.53 | 3.95 | 2.70 | 2.74 | ||||||||||
| Total | 101.96 ± 1.14ef | 55.67 ± 2.15n | 63.43 ± 1.11m | 70.75 ± 1.77l | 95.75 ± 1.65g | 124.95 ± 1.14d | 57.84 ± 2.05n | 76.79 ± 1.19k | ||||||||||
| Spcecies | Cultivar | Antioxidant Capacity | In Vitro Inhibition Activities | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ABTS | FRAP | ORAC | α-amylase | α-glucosidase | Pancreatic Lipase | AChE | BuChE | 15-LOX | ||
| Chaenomeles × superba | Crimson and Gold | 16.03 ± 1.04cdef | 16.00 ± 0.99de | 54.93 ± 1.11bc | 17.49 ± 0.88a | 7.03 ± 0.20def | 0.29 ± 0.01ab | 11.84 ± 0.24efg | 10.13 ± 0.97efgh | 99.81 ± 0.15a |
| Texas Scarlet | 19.63 ± 0.99ab | 17.90 ± 1.22bcde | 53.89 ± 1.37cd | 14.34 ± 0.48b | 5.08 ± 0.22g | 0.09 ± 0.00def | 17.43 ± 0.56ab | 8.90 ± 0.70fghi | 43.23 ± 0.73f | |
| Nicoline | 20.61 ± 1.13a | 21.32 ± 0.85a | 51.86 ± 0.90cde | 13.88 ± 0.98b | 2.67 ± 0.17h | 0.07 ± 0.02ef | 16.02 ± 0.25bcd | 16.14 ± 0.79d | 71.24 ± 0.18d | |
| Andenken an Karl Ramcke | 18.80 ± 0.88abc | 15.51 ± 1.14def | 40.38 ± 0.83h | 16.28 ± 0.77ab | 6.71 ± 0.72defg | 0.04 ± 0.02f | 13.03 ± 0.81defg | 9.96 ± 0.17efgh | 74.81 ± 0.18c | |
| Pink Lady | 17.65 ± 1.50abcde | 17.29 ± 0.63bcde | 57.86 ± 1.00b | 18.01 ± 0.89a | 5.90 ± 0.85fg | 0.20 ± 0.10bcd | 15.94 ± 0.13bcd | 12.14 ± 0.87e | >100.00 | |
| Colour Trail | 11.02 ± 0.72h | 10.56 ± 0.55h | 66.59 ± 0.55a | 17.56 ± 0.99a | 7.95 ± 0.16cde | <0.01 | 17.53 ± 0.41ab | 7.85 ± 0.77hi | 75.94 ± 0.29c | |
| Flocon Rose | 15.38 ± 1.80defg | 15.32 ± 0.88efg | 45.25 ± 0.99g | 15.59 ± 0.66ab | 7.18 ± 0.11def | <0.01 | 11.73 ± 0.47efg | 22.70 ± 0.63bc | 98.15 ± 1.00a | |
| Hollandia | 18.36 ± 0.63abcd | 19.44 ± 1.11abc | 40.72 ± 0.78h | 16.49 ± 0.32ab | 6.10 ± 1.23fg | <0.01 | 10.73 ± 0.74gh | 15.89 ± 0.44d | >100.00 | |
| Jet Trail | 18.91 ± 0.91abc | 20.04 ± 1.37ab | 54.80 ± 0.46bc | 17.97 ± 1.00a | 6.97 ± 0.77def | 0.29 ± 0.01ab | 12.22 ± 0.65efg | 8.09 ± 0.99ghi | >100.00 | |
| wild | 17.46 ± 0.81bcde | 18.45 ± 0.45abcd | 50.18 ± 0.89ef | 18.25 ± 0.39a | 7.17 ± 0.57def | <0.01 | 11.05 ± 0.57g | 31.59 ± 0.95a | >100.00 | |
| Cameo | 15.24 ± 0.24efg | 11.93 ± 0.33h | 51.63 ± 1.62de | 16.75 ± 0.57ab | 8.54 ± 0.34cd | 0.12 ± 0.02def | 11.56 ± 0.84fg | 12.37 ± 1.22e | 36.84 ± 0.44g | |
| Chaenomeles japonica | Cido | 18.06 ± 1.52abcde | 18.00 ± 0.65bcde | 48.48 ± 1.55f | 16.47 ± 0.56ab | 6.49 ± 0.49efg | 0.17 ± 0.00cde | 14.75 ± 0.75bcde | 16.42 ± 0.31d | 42.11 ± 0.56f |
| Red Joy | 13.50 ± 0.50fgh | 12.76 ± 1.12fgh | 53.43 ± 0.87cd | 17.45 ± 0.54a | 15.19 ± 0.14a | 0.06 ± 0.01ef | 7.74 ± 0.34hi | 6.06 ± 0.41i | 73.31 ± 0.74cd | |
| wild #1 | 12.41 ± 0.41gh | 11.53 ± 0.55h | 40.28 ± 0.66h | 16.66 ± 0.87ab | 6.11 ± 0.19efg | 0.35 ± 0.05a | 12.14 ± 0.20efg | 32.11 ± 1.13a | 90.60 ± 0.69b | |
| wild #2 | 13.72 ± 0.72fgh | 12.30 ± 0.62gh | 33.99 ± 1.74i | 16.11 ± 1.13ab | 9.57 ± 0.55c | <0.01 | 10.13 ± 0.30gh | 20.68 ± 0.56c | 70.37 ± 0.55d | |
| n1 (new) | 16.96 ± 0.96bcde | 16.90 ± 0.22cde | 51.07 ± 0.77def | 16.89 ± 0.98ab | 6.09 ± 1.22fg | 0.25 ± 0.05abc | 14.18 ± 0.49cdef | 11.17 ± 0.66efg | 66.05 ± 0.99e | |
| Chaenomeles speciosa | Nivalis | 17.54 ± 0.54abcde | 16.39 ± 0.47cde | 44.30 ± 1.50g | 18.48 ± 0.43a | 6.56 ± 0.46efg | 0.20 ± 0.02bcd | 16.52 ± 35bc | 11.97 ± 0.20ef | 74.81 ± 0.45c |
| Rubra | 10.91 ± 0.91h | 10.24 ± 1.20h | 33.82 ± 0.49i | 18.38 ± 0.77a | 5.74 ± 0.84fg | 0.04 ± 0.00f | 6.65 ± 0.73i | 11.07 ± 0.81efg | 14.29 ± 0.99i | |
| Simonii | 17.37 ± 1.37bcde | 17.02 ± 0.98bcde | 45.18 ± 1.15g | 16.88 ± 1.00ab | 12.48 ± 0.68b | 0.20 ± 0.05bcd | 20.42 ± 0.99a | 25.79 ± 0.11b | 27.37 ± 0.30h | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Golis, T.; Bąbelewski, P. ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and other Chemical Compounds. Antioxidants 2020, 9, 60. https://doi.org/10.3390/antiox9010060
Turkiewicz IP, Wojdyło A, Tkacz K, Nowicka P, Golis T, Bąbelewski P. ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and other Chemical Compounds. Antioxidants. 2020; 9(1):60. https://doi.org/10.3390/antiox9010060
Chicago/Turabian StyleTurkiewicz, Igor Piotr, Aneta Wojdyło, Karolina Tkacz, Paulina Nowicka, Tomasz Golis, and Przemysław Bąbelewski. 2020. "ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and other Chemical Compounds" Antioxidants 9, no. 1: 60. https://doi.org/10.3390/antiox9010060
APA StyleTurkiewicz, I. P., Wojdyło, A., Tkacz, K., Nowicka, P., Golis, T., & Bąbelewski, P. (2020). ABTS On-Line Antioxidant, α-Amylase, α-Glucosidase, Pancreatic Lipase, Acetyl- and Butyrylcholinesterase Inhibition Activity of Chaenomeles Fruits Determined by Polyphenols and other Chemical Compounds. Antioxidants, 9(1), 60. https://doi.org/10.3390/antiox9010060