Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences

 ,
,  ,
,  , , ,
, , , 
Abstract
1. Introduction
1.1. Background
1.2. Implication of Se in Mammalian Reproduction: An Overview
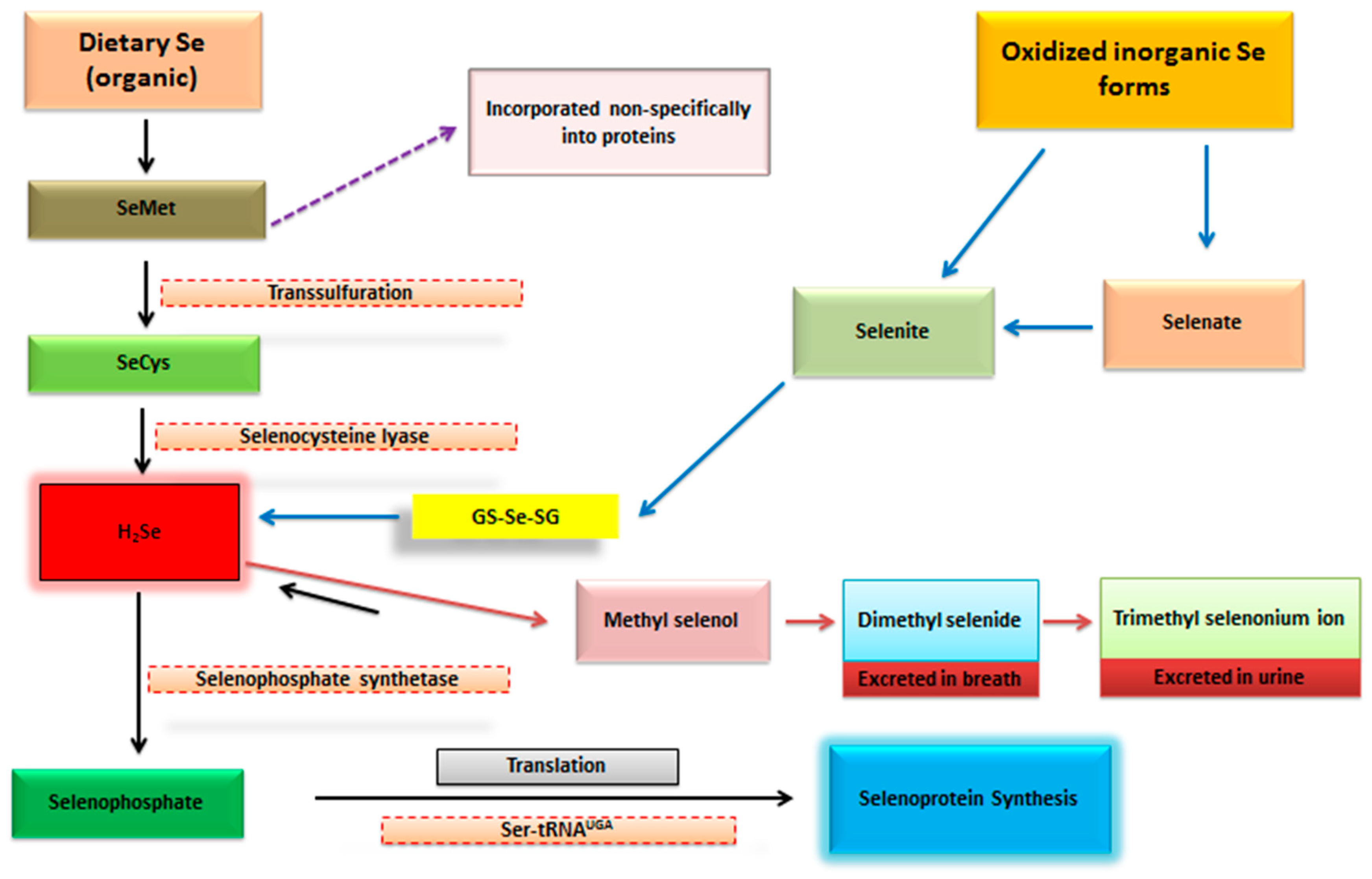
1.3. Selenium Biochemistry and Significance
1.4. Rationale
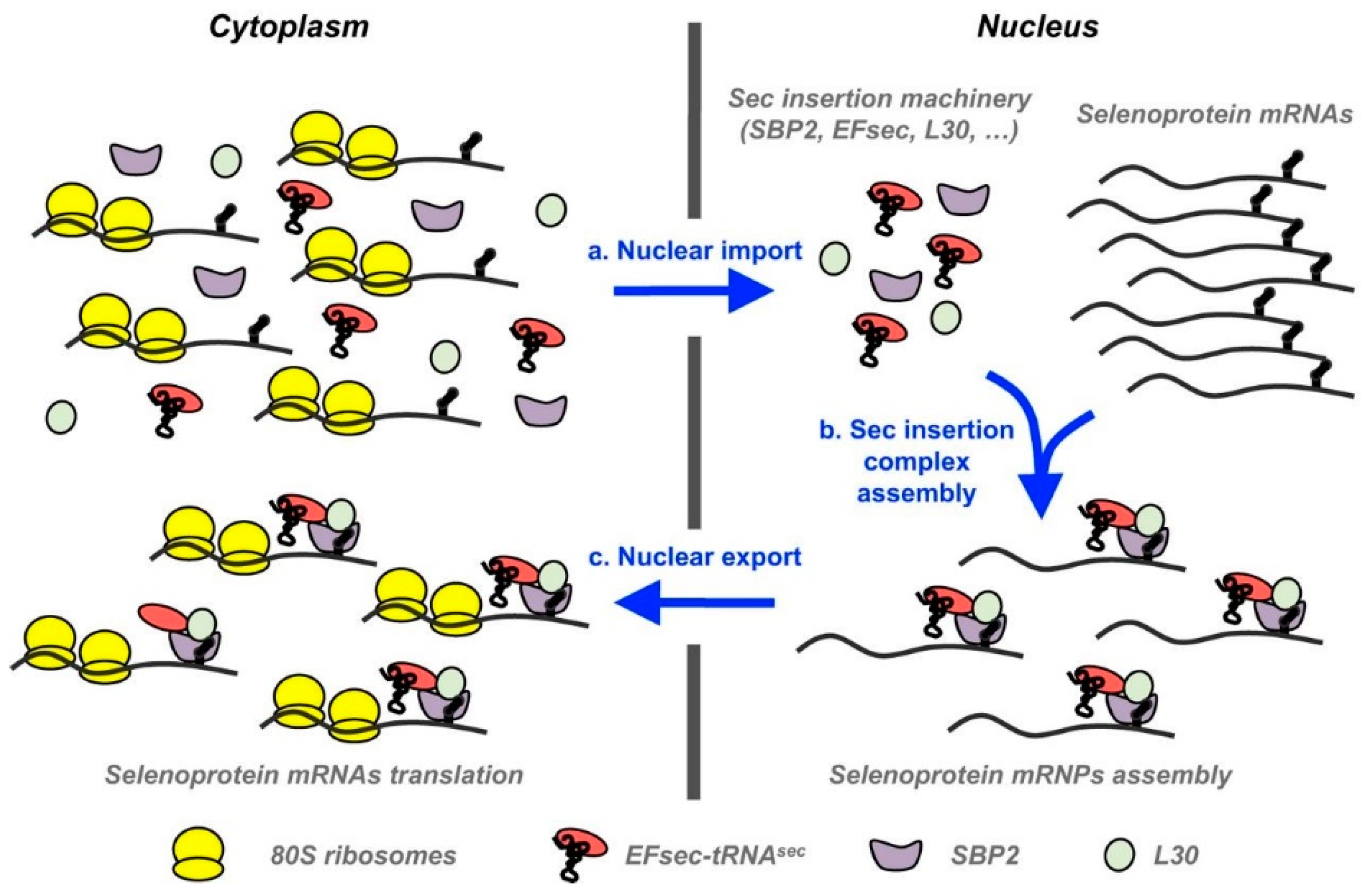
2. Important Selenoproteins Relevant to Male Reproduction
3. Role of Selenium in Male Reproduction
3.1. Role of Selenium in Steroidogenesis and Spermatogenesis
3.2. Implication of Se on Male Fertility-Related Parameters
Selenium in Seminal Plasma and its Implication in Male Fertility
3.3. Combinatorial Effects of Se (as a Part of Micronutrient Supplement) on Male Fertility Outcomes (Animal Studies)
3.4. Selenium and Sperm Cryopreservation
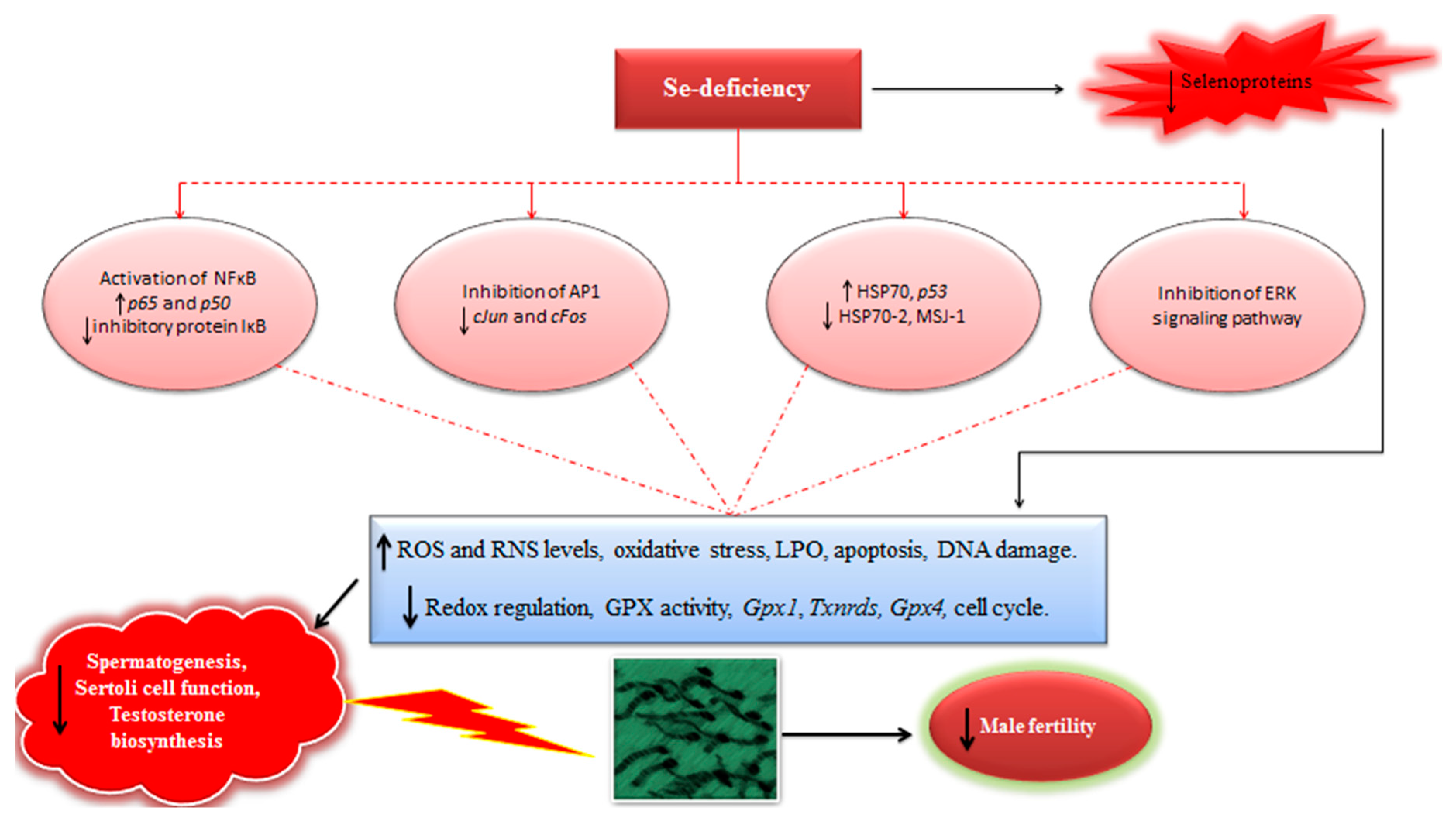
4. Effects of Se Supplementation on Different Experimentally-Induced Challenges in Male Reproductive System (Animal Model Studies)
5. Human Studies (Clinical Evidences)
6. Discussion and Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Oldfield, J.E. The two faces of selenium. J. Nutr. 1987, 117, 2002–2008. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- O’Dell, B.L.; Sunde, R.A. Handbook of Nutritionally Essential Mineral Elements; CRC Press: New York, NY, USA; Basel, Switzerland; Hong Kong, China, 1997. [Google Scholar]
- Ceko, M.J.; O’leary, S.; Harris, H.H.; Hummitzsch, K.; Rodgers, R.J. Trace elements in ovaries: Measurement and physiology. Biol. Reprod. 2016, 94, 86. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Lan, D.; Li, J.; Lin, Y.; Li, M. Selenium supplementation during in vitro maturation enhances meiosis and developmental capacity of yak oocytes. Anim. Sci. J. 2018, 89, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Hedaoo, M.; Khllare, K.; Meshram, M.; Sahatpure, S.; Patil, M. Study of some serum trace minerals in cyclic and non-cyclic surti buffaloes. Vet. World 2008, 1, 71. [Google Scholar]
- Mirone, M.; Giannetta, E.; Isidori, A. Selenium and reproductive function. A systematic review. J. Endocrinol. Investig. 2013, 36, 28–36. [Google Scholar]
- Pappas, A.; Zoidis, E.; Surai, P.; Zervas, G. Selenoproteins and maternal nutrition. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Flohé, L.; Garolla, A.; Roveri, A.; Ursini, F.; Maiorino, M. Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase. Biol. Reprod. 2002, 67, 967–971. [Google Scholar] [CrossRef]
- Kommisrud, E.; Østerås, O.; Vatn, T. Blood selenium associated with health and fertility in Norwegian dairy herds. Acta Vet. Scand. 2005, 46, 229. [Google Scholar] [CrossRef]
- Behne, D.; Höfer, T.; von Berswordt-Wallrabe, R.; Elger, W. Selenium in the testis of the rat: Studies on its regulation and its importance for the organism. J. Nutr. 1982, 112, 1682–1687. [Google Scholar] [CrossRef]
- Behne, D.; Weiler, H.; Kyriakopoulos, A. Effects of selenium deficiency on testicular morphology and function in rats. J. Reprod. Fertil. 1996, 106, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L. Selenium in mammalian spermiogenesis. Biol. Chem. 2007, 388, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, U.; Kamran, Z.; Raza, I.; Ahmad, S.; Babar, W.; Riaz, M.; Iqbal, Z. Role of selenium in male reproduction—A review. Anim. Reprod. Sci. 2014, 146, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Irvine, D.S. Glutathione as a treatment for male infertility. Rev. Reprod. 1996, 1, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Letavayová, L.; Vlčková, V.; Brozmanová, J. Selenium: From cancer prevention to DNA damage. Toxicology 2006, 227, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schrauzer, G.N. Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 2000, 130, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: Spotlight on speciation. Br. J. Nutr. 2008, 100, 238–253. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Regulation of selenium metabolism and transport. Annu. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I. Selenium in Pig Nutrition and reproduction: Boars and semen quality—A Review. Asian-Australas. J. Anim. Sci. 2015, 28, 730. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I. Selenium in livestock and other domestic animals. In Selenium; Hatfield, D., Schweizer, U., Tsuji, P., Gladyshev, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 595–606. [Google Scholar]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: How does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef]
- Dalto, D.; Matte, J.-J. Pyridoxine (vitamin B6) and the glutathione peroxidase system; a link between one-carbon metabolism and antioxidation. Nutrients 2017, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Dalto, D.B.; Audet, I.; Lapointe, J.; Matte, J.J. The importance of pyridoxine for the impact of the dietary selenium sources on redox balance, embryo development, and reproductive performance in gilts. J. Trace Elem. Med. Biol. 2016, 34, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Dalto, D.B.; Roy, M.; Audet, I.; Palin, M.-F.; Guay, F.; Lapointe, J.; Matte, J.J. Interaction between vitamin B6 and source of selenium on the response of the selenium-dependent glutathione peroxidase system to oxidative stress induced by oestrus in pubertal pig. J. Trace Elem. Med. Biol. 2015, 32, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484S–1491S. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Xia, Y.; Hill, K.E.; Byrne, D.W.; Xu, J.; Burk, R.F. Effectiveness of selenium supplements in a low-selenium area of China. Am. J. Clin. Nutr. 2005, 81, 829–834. [Google Scholar] [CrossRef]
- Swanson, C.; Patterson, B.; Levander, O.; Veillon, C.; Taylor, P.; Helzlsouer, K.; McAdam, P.; Zech, L. Human [74Se] selenomethionine metabolism: A kinetic model. Am. J. Clin. Nutr. 1991, 54, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Huerta, V.D.; Sánchez, M.L.F.; Sanz-Medel, A. Quantitative selenium speciation in cod muscle by isotope dilution ICP-MS with a reaction cell: Comparison of different reported extraction procedures. J. Anal. At. Spectrom. 2004, 19, 644–648. [Google Scholar] [CrossRef]
- Steen, A.; Strøm, T.; Bernhoft, A. Organic selenium supplementation increased selenium concentrations in ewe and newborn lamb blood and in slaughter lamb meat compared to inorganic selenium supplementation. Acta Vet. Scand. 2008, 50, 7. [Google Scholar] [CrossRef]
- Juniper, D.T.; Phipps, R.H.; Ramos-Morales, E.; Bertin, G. Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle. J. Anim. Sci. 2008, 86, 3100–3109. [Google Scholar] [CrossRef]
- Deagen, J.; Butler, J.; Beilstein, M.; Whanger, P. Effects of dietary selenite, selenocystine and selenomethionine on selenocysteine lyase and glutathione peroxidase activities and on selenium levels in rat tissues. J. Nutr. 1987, 117, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Behne, D.; Alber, D.; Kyriakopoulos, A. Long-term selenium supplementation of humans: Selenium status and relationships between selenium concentrations in skeletal muscle and indicator materials. J. Trace Elem. Med. Biol. 2010, 24, 99–105. [Google Scholar] [CrossRef]
- Suzuki, K.T.; Doi, C.; Suzuki, N. Metabolism of 76Se-methylselenocysteine compared with that of 77Se-selenomethionine and 82Se-selenite. Toxicol. Appl. Pharmacol. 2006, 217, 185–195. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Selenium-enriched yeast as source for selenium added for nutritional purposes in foods for particular nutritional uses and foods (including food supplements) for the general population-Scientific Opinion of the Panel on Food Additives. EFSA J. 2008, 766, 1–42. [Google Scholar] [CrossRef]
- Yamashita, Y.; Yamashita, M. Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J. Biol. Chem. 2010, 285, 18134–18138. [Google Scholar] [CrossRef] [PubMed]
- Kotrebai, M.; Birringer, M.; Tyson, J.F.; Block, E.; Uden, P.C. Selenium speciation in enriched and natural samples by HPLC-ICP-MS and HPLC-ESI-MS with perfluorinated carboxylic acid ion-pairing agents. Analyst 2000, 125, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Birringer, M.; Block, E.; Kotrebai, M.; Tyson, J.F.; Uden, P.C.; Lisk, D.J. Chemical speciation influences comparative activity of selenium-enriched garlic and yeast in mammary cancer prevention. J. Agric. Food Chem. 2000, 48, 2062–2070. [Google Scholar] [CrossRef] [PubMed]
- Cappon, C.J.; Smith, J.C. Chemical form and distribution of mercury and selenium in edible seafood. J. Anal. Toxicol. 1982, 6, 10–21. [Google Scholar] [CrossRef] [PubMed]
- JohnáLewis, D. Investigation of selenium speciation in in vitro gastrointestinal extracts of cooked cod by high-performance liquid chromatography–inductively coupled plasma mass spectrometry and electrospray mass spectrometry. J. Anal. At. Spectrom. 1996, 11, 1177–1182. [Google Scholar]
- Bergdahl, I.A. Fractionation of soluble selenium compounds from fish using size-exclusion chromatography with on-line detection by inductively coupled plasma mass spectrometry. Analyst 1999, 124, 1435–1438. [Google Scholar] [CrossRef]
- Qazi, I.H.; Angel, C.; Yang, H.; Pan, B.; Zoidis, E.; Zeng, C.J.; Han, H.; Zhou, G.B. Selenium, Selenoproteins, and Female Reproduction: A Review. Molecules 2018, 23, 3053. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Yim, S.H.; Clish, C.B.; Gladyshev, V.N. Selenium Deficiency Is Associated with Pro-longevity Mechanisms. Cell Rep. 2019, 27, 2785–2797. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, V.N.; Arnér, E.S.; Berry, M.J.; Brigelius-Flohé, R.; Bruford, E.A.; Burk, R.F.; Carlson, B.A.; Castellano, S.; Chavatte, L.; Conrad, M. Selenoprotein gene nomenclature. J. Biol. Chem. 2016, 291, 24036–24040. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Touat-Hamici, Z.; Legrain, Y.; Bulteau, A.-L.; Chavatte, L. Selective up-regulation of human selenoproteins in response to oxidative stress. J. Biol. Chem. 2014, 289, 14750–14761. [Google Scholar] [CrossRef]
- Reeves, M.; Hoffmann, P. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef]
- Sunde, R.A.; Raines, A.M.; Barnes, K.M.; Evenson, J.K. Selenium status highly regulates selenoprotein mRNA levels for only a subset of the selenoproteins in the selenoproteome. Biosci. Rep. 2009, 29, 329–338. [Google Scholar] [CrossRef]
- Hammad, G.; Legrain, Y.; Touat-Hamici, Z.; Duhieu, S.; Cornu, D.; Bulteau, A.-L.; Chavatte, L. Interplay between selenium levels and replicative senescence in WI-38 human fibroblasts: A proteomic approach. Antioxidants 2018, 7, 19. [Google Scholar] [CrossRef]
- Carlson, B.A.; Novoselov, S.V.; Kumaraswamy, E.; Lee, B.J.; Anver, M.R.; Gladyshev, V.N.; Hatfield, D.L. Specific excision of the selenocysteine tRNA [Ser] Sec (Trsp) gene in mouse liver demonstrates an essential role of selenoproteins in liver function. J. Biol. Chem. 2004, 279, 8011–8017. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Schweizer, U.; Holtmann, B.; Flohé, L.; Sendtner, M.; Köhrle, J. Gene disruption discloses role of selenoprotein P in selenium delivery to target tissues. Biochem. J. 2003, 370, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Dikiy, A.; Novoselov, S.V.; Fomenko, D.E.; Sengupta, A.; Carlson, B.A.; Cerny, R.L.; Ginalski, K.; Grishin, N.V.; Hatfield, D.L.; Gladyshev, V.N. SelT, SelW, SelH, and Rdx12: Genomics and molecular insights into the functions of selenoproteins of a novel thioredoxin-like family. Biochemistry 2007, 46, 6871–6882. [Google Scholar] [CrossRef] [PubMed]
- Zoidis, E.; Pappas, A. The health effects of selenoproteins. In Selenium: Sources, Functions and Health Effects; Aomori, C., Hokkaido, M., Eds.; Nova Science Publishers: New York, NY, USA, 2012. [Google Scholar]
- Conrad, M.; Schweizer, U. Mouse Models that Target Individual Selenoproteins. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 567–578. [Google Scholar]
- Riese, C.; Michaelis, M.; Mentrup, B.; Gotz, F.; Kohrle, J.; Schweizer, U.; Schomburg, L. Selenium-dependent pre-and posttranscriptional mechanisms are responsible for sexual dimorphic expression of selenoproteins in murine tissues. Endocrinology 2006, 147, 5883–5892. [Google Scholar] [CrossRef] [PubMed]
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Knapen, M.F.; Zusterzeel, P.L.; Peters, W.H.; Steegers, E.A. Glutathione and glutathione-related enzymes in reproduction: A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 82, 171–184. [Google Scholar] [CrossRef]
- Ursini, F.; Heim, S.; Kiess, M.; Maiorino, M.; Roveri, A.; Wissing, J.; Flohé, L. Dual function of the selenoprotein PHGPx during sperm maturation. Science 1999, 285, 1393–1396. [Google Scholar] [CrossRef] [PubMed]
- Safarinejad, M.R.; Safarinejad, S. Efficacy of selenium and/or N-acetyl-cysteine for improving semen parameters in infertile men: A double-blind, placebo controlled, randomized study. J. Urol. 2009, 181, 741–751. [Google Scholar] [CrossRef]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Conrad, M.; Moreno, S.; Sinowatz, F.; Ursini, F.; Kölle, S.; Roveri, A.; Brielmeier, M.; Wurst, W.; Maiorino, M.; Bornkamm, G. The nuclear form of phospholipid hydroperoxide glutathione peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol. Cell. Biol. 2005, 25, 7637–7644. [Google Scholar] [CrossRef]
- Chabory, E.; Damon, C.; Lenoir, A.; Henry-Berger, J.; Vernet, P.; Cadet, R.; Saez, F.; Drevet, J. Mammalian glutathione peroxidases control acquisition and maintenance of spermatozoa integrity 1. J. Anim. Sci. 2010, 88, 1321–1331. [Google Scholar] [CrossRef]
- Schneider, M.; Förster, H.; Boersma, A.; Seiler, A.; Wehnes, H.; Sinowatz, F.; Neumüller, C.; Deutsch, M.J.; Walch, A.; de Angelis, M.H. Mitochondrial glutathione peroxidase 4 disruption causes male infertility. FASEB J. 2009, 23, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Suzuki, K.; Ishizaka, K.; Ichinose, S.; Oshima, H.; Okayasu, I.; Emoto, K.; Umeda, M.; Nakagawa, Y. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males. Biol. Reprod. 2001, 64, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Hakkaku, N.; Iwamoto, R.; Suzuki, J.; Suzuki, T.; Tajima, Y.; Konishi, K.; Minami, S.; Ichinose, S.; Ishizaka, K. Depletion of selenoprotein GPx4 in spermatocytes causes male infertility in mice. J. Biol. Chem. 2009, 284, 32522–32532. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; Maria, J.; Simón, C.; Pellicer, A.; Remohí, J.; Garrido, N. Effect of sperm glutathione peroxidases 1 and 4 on embryo asymmetry and blastocyst quality in oocyte donation cycles. Fertil. Steril. 2006, 86, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Parillo, F.; Sylla, L.; Palombi, C.; Monaci, M.; Stradaioli, G. Immunocytochemical Localisation of Phospholipid Hydroperoxide Glutathione Peroxidase in Bull’s Spermatogenic Cells. Ital. J. Anim. Sci. 2014, 13, 3483. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Apolipoprotein E receptor-2 (ApoER2) mediates selenium uptake from selenoprotein P by the mouse testis. J. Biol. Chem. 2007, 282, 12290–12297. [Google Scholar] [CrossRef]
- Hill, K.E.; Zhou, J.; McMahan, W.J.; Motley, A.K.; Atkins, J.F.; Gesteland, R.F.; Burk, R.F. Deletion of selenoprotein P alters distribution of selenium in the mouse. J. Biol. Chem. 2003, 278, 13640–13646. [Google Scholar] [CrossRef]
- Kehr, S.; Malinouski, M.; Finney, L.; Vogt, S.; Labunskyy, V.M.; Kasaikina, M.V.; Carlson, B.A.; Zhou, Y.; Hatfield, D.L.; Gladyshev, V.N. X-ray fluorescence microscopy reveals the role of selenium in spermatogenesis. J. Mol. Biol. 2009, 389, 808–818. [Google Scholar] [CrossRef]
- Noblanc, A.; Kocer, A.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Conrad, M.; Drevet, J.R. Glutathione peroxidases at work on epididymal spermatozoa: An example of the dual effect of reactive oxygen species on mammalian male fertilizing ability. J. Androl. 2011, 32, 641–650. [Google Scholar] [CrossRef]
- Boitani, C.; Puglisi, R. Selenium, a key element in spermatogenesis and male fertility. In Molecular Mechanisms in Spermatogenesis; Springer: Berlin/Heidelberg, Germany, 2009; pp. 65–73. [Google Scholar]
- Su, D.; Novoselov, S.V.; Sun, Q.-A.; Moustafa, M.E.; Zhou, Y.; Oko, R.; Hatfield, D.L.; Gladyshev, V.N. Mammalian selenoprotein thioredoxin-glutathione reductase roles in disulfide bond formation and sperm maturation. J. Biol. Chem. 2005, 280, 26491–26498. [Google Scholar] [CrossRef]
- Michaelis, M.; Gralla, O.; Behrends, T.; Scharpf, M.; Endermann, T.; Rijntjes, E.; Pietschmann, N.; Hollenbach, B.; Schomburg, L. Selenoprotein P in seminal fluid is a novel biomarker of sperm quality. Biochem. Biophys. Res. Commun. 2014, 443, 905–910. [Google Scholar] [CrossRef]
- Renko, K.; Werner, M.; Renner-Muller, I.; Cooper, T.G.; Yeung, C.H.; Hollenbach, B.; Scharpf, M.; Kohrle, J.; Schomburg, L.; Schweizer, U. Hepatic selenoprotein P (SePP) expression restores selenium transport and prevents infertility and motor-incoordination in Sepp-knockout mice. Biochem. J. 2008, 409, 741–749. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Selenoprotein P is required for mouse sperm development. Biol. Reprod. 2005, 73, 201–211. [Google Scholar] [CrossRef]
- Turanov, A.A.; Malinouski, M.; Gladyshev, V.N. Selenium and male reproduction. In Selenium: Its Molecular Biology and Role in Human Health; Hatfield, D.L., Berry, M.J., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2012; pp. 409–417. [Google Scholar]
- Behne, D.; Duk, M.; Elger, W. Selenium content and glutathione peroxidase activity in the testis of the maturing rat. J. Nutr. 1986, 116, 1442–1447. [Google Scholar] [CrossRef]
- Nishimura, K.; Matsumiya, K.; Tsujimura, A.; Koga, M.; Kitamura, M.; Okuyama, A. Association of selenoprotein P with testosterone production in cultured Leydig cells. Arch. Androl. 2001, 47, 67–76. [Google Scholar] [CrossRef]
- Koga, M.; Tanaka, H.; Yomogida, K.; Tsuchida, J.; Uchida, K.; Kitamura, M.; Sakoda, S.; Matsumiya, K.; Okuyama, A.; Nishimune, Y. Expression of selenoprotein-P messenger ribonucleic acid in the rat testis. Biol. Reprod. 1998, 58, 261–265. [Google Scholar] [CrossRef][Green Version]
- Oluboyo, A.; Adijeh, R.; Onyenekwe, C.; Oluboyo, B.; Mbaeri, T.; Odiegwu, C.; Chukwuma, G.; Onwuasoanya, U. Relationship between serum levels of testosterone, zinc and selenium in infertile males attending fertility clinic in Nnewi, south east Nigeria. Afr. J. Med. Med. Sci. 2012, 41, 51–54. [Google Scholar]
- Villaverde, A.I.S.; Fioratti, E.G.; Ramos, R.S.; Neves, R.C.; Ferreira, J.C.P.; Cardoso, G.S.; Padilha, P.M.; Lopes, M.D. Blood and seminal plasma concentrations of selenium, zinc and testosterone and their relationship to sperm quality and testicular biometry in domestic cats. Anim. Reprod. Sci. 2014, 150, 50–55. [Google Scholar] [CrossRef]
- Richards, J.; Hallford, D.; Duff, G. Serum luteinizing hormone, testosterone, and thyroxine and growth responses of ram lambs fed locoweed (Oxvtropis sericea) and treated with vitamin e/selenium. Theriogenology 1999, 52, 1055–1066. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, C.; Yue, W.; Shi, L.; Zhu, X.; Lei, F. Short-term effect of dietary selenium-enriched yeast on semen parameters, antioxidant status and Se concentration in goat seminal plasma. Anim. Feed Sci. Technol. 2010, 157, 104–108. [Google Scholar] [CrossRef]
- Ren, X.-M.; Wang, G.-G.; Xu, D.-Q.; Luo, K.; Liu, Y.-X.; Zhong, Y.-H.; Cai, Y.-Q. The protection of selenium on cadmium-induced inhibition of spermatogenesis via activating testosterone synthesis in mice. Food Chem. Toxicol. 2012, 50, 3521–3529. [Google Scholar] [CrossRef]
- Shi, L.; Song, R.; Yao, X.; Ren, Y. Effects of selenium on the proliferation, apoptosis and testosterone production of sheep Leydig cells in vitro. Theriogenology 2017, 93, 24–32. [Google Scholar] [CrossRef]
- Liu, H.; Li, X.; Qin, F.; Huang, K. Selenium suppresses oxidative-stress-enhanced vascular smooth muscle cell calcification by inhibiting the activation of the PI3K/AKT and ERK signaling pathways and endoplasmic reticulum stress. JBIC J. Biol. Inorg. Chem. 2014, 19, 375–388. [Google Scholar] [CrossRef]
- Shi, L.; Song, R.; Yao, X.; Duan, Y.; Ren, Y.; Zhang, C.; Yue, W.; Lei, F. Effects of maternal dietary selenium (Se-enriched yeast) on testis development, testosterone level and testicular steroidogenesis-related gene expression of their male kids in Taihang Black Goats. Theriogenology 2018, 114, 95–102. [Google Scholar] [CrossRef]
- Cerny, K.; Garbacik, S.; Skees, C.; Burris, W.; Matthews, J.; Bridges, P. Gestational form of selenium in free-choice mineral mixes affects transcriptome profiles of the neonatal calf testis, including those of steroidogenic and spermatogenic pathways. Biol. Trace Elem. Res. 2016, 169, 56–68. [Google Scholar] [CrossRef]
- Brennan, K.M.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Selenium content in blood fractions and liver of beef heifers is greater with a mix of inorganic/organic or organic versus inorganic supplemental selenium but the time required for maximal assimilation is tissue-specific. Biol. Trace Elem. Res. 2011, 144, 504–516. [Google Scholar] [CrossRef]
- Liao, S.F.; Brown, K.R.; Stromberg, A.J.; Burris, W.R.; Boling, J.A.; Matthews, J.C. Dietary supplementation of selenium in inorganic and organic forms differentially and commonly alters blood and liver selenium concentrations and liver gene expression profiles of growing beef heifers. Biol. Trace Elem. Res. 2011, 140, 151–169. [Google Scholar] [CrossRef]
- Watanabe, T.; Endo, A. Effects of selenium deficiency on sperm morphology and spermatocyte chromosomes in mice. Mutat. Res. Lett. 1991, 262, 93–99. [Google Scholar] [CrossRef]
- Kaur, P.; Bansal, M.P. Effect of selenium-induced oxidative stress on the cell kinetics in testis and reproductive ability of male mice. Nutrition 2005, 21, 351–357. [Google Scholar] [CrossRef]
- Adegoke, E.; Wang, X.; Wang, H.; Wang, C.; Zhang, H.; Zhang, G. Selenium (Na 2 SeO 3) Upregulates Expression of Immune Genes and Blood–Testis Barrier Constituent Proteins of Bovine Sertoli Cell In Vitro. Biol. Trace Elem. Res. 2018, 185, 332–343. [Google Scholar] [CrossRef]
- Adegoke, E.; Xue, W.; Machebe, N.; Adeniran, S.; Hao, W.; Chen, W.; Han, Z.; Guixue, Z.; Peng, Z. Sodium Selenite inhibits mitophagy, downregulation and mislocalization of blood–testis barrier proteins of bovine Sertoli cell exposed to microcystin-leucine arginine (MC-LR) via TLR4/NF-kB and mitochondrial signaling pathways blockage. Ecotoxicol. Environ. Saf. 2018, 166, 165–175. [Google Scholar] [CrossRef]
- Kaushal, N.; Bansal, M. Inhibition of CDC2/Cyclin B1 in response to selenium-induced oxidative stress during spermatogenesis: Potential role of Cdc25c and p21. Mol. Cell. Biochem. 2007, 298, 139–150. [Google Scholar] [CrossRef]
- Kaushal, N.; Bansal, M. Selenium variation induced oxidative stress regulates p53 dependent germ cell apoptosis: Plausible involvement of HSP70-2. Eur. J. Nutr. 2009, 48, 221–227. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M. Alterations in selenium status influences reproductive potential of male mice by modulation of transcription factor NFκB. Biometals 2007, 20, 49–59. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M.P. Role of selenium in spermatogenesis: Differential expression of cjun and cfos in tubular cells of mice testis. Mol. Cell. Biochem. 2006, 292, 27–38. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M. Role of selenium in regulation of spermatogenesis: Involvement of activator protein 1. Biofactors 2005, 23, 151–162. [Google Scholar] [CrossRef]
- Kaushal, N.; Bansal, M. Diminished reproductive potential of male mice in response to selenium-induced oxidative stress: Involvement of HSP70, HSP70-2, and MSJ-1. J. Biochem. Mol. Toxicol. 2009, 23, 125–136. [Google Scholar] [CrossRef]
- Sattar, H.; Yang, J.; Zhao, X.; Cai, J.; Liu, Q.; Ishfaq, M.; Yang, Z.; Chen, M.; Zhang, Z.; Xu, S. Selenoprotein-U (SelU) knockdown triggers autophagy through PI3K–Akt–mTOR pathway inhibition in rooster Sertoli cells. Metallomics 2018, 10, 929–940. [Google Scholar] [CrossRef]
- Bano, I.; Malhi, M.; Soomro, S.; Kandhro, S.; Awais, M.; Baloch, S.; Perveen, S.; Sajjad, H. Effect of Dietary Selenium Supplementation on Morphology and Antioxidant Status in Testes of Goat. J. Basic Appl. Sci. 2018, 14, 53–61. [Google Scholar] [CrossRef]
- Stefanov, R.; Chervenkov, M.; Anev, G.; Maksimović, N.; Andreeva, M.; Ivanova, T.; Milovanović, A. Effect of supplementation with inorganic and organic selenium on sperm quality and quantity in north-east Bulgarian merino rams. Biotechnol. Anim. Husb. 2018, 34, 69–81. [Google Scholar] [CrossRef]
- Asri-Rezaei, S.; Nourian, A.; Shalizar-Jalali, A.; Najafi, G.; Nazarizadeh, A.; Koohestani, M.; Karimi, A. Selenium supplementation in the form of selenium nanoparticles and selenite sodium improves mature male mice reproductive performances. Iran. J. Basic Med Sci. 2018, 21, 577. [Google Scholar]
- Martins, S.M.M.K.; Afonso, E.R.; Parazzi, L.J.; Andrade, A.F.C.D.; Leal, D.F.; Gameiro, A.H.; Moretti, A.D.S.A.; Arruda, R.P.D. Organic selenium supplementation is cost-effective for increasing the number of seminal doses produced by sexually mature boars. Rev. Bras. Zootec. 2018, 47. [Google Scholar] [CrossRef]
- Liu, L.; He, Y.; Xiao, Z.; Tao, W.; Zhu, J.; Wang, B.; Liu, Z.; Wang, M. Effects of selenium nanoparticles on reproductive performance of male Sprague-Dawley rats at supranutritional and nonlethal levels. Biol. Trace Elem. Res. 2017, 180, 81–89. [Google Scholar] [CrossRef]
- Zhou, J.-C.; Zheng, S.; Mo, J.; Liang, X.; Xu, Y.; Zhang, H.; Gong, C.; Liu, X.-L.; Lei, X.G. Dietary selenium deficiency or excess reduces sperm quality and testicular mRNA abundance of nuclear glutathione peroxidase 4 in rats. J. Nutr. 2017, 147, 1947–1953. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Zhu, Y.; Wu, C.; Lu, C.; Ezekwe, M.O.; Liao, S.F.; Haung, K. Selenium-enriched probiotics improves murine male fertility compromised by high fat diet. Biol. Trace Elem. Res. 2012, 147, 251–260. [Google Scholar] [CrossRef]
- Shalini, S.; Bansal, M. Dietary selenium deficiency as well as excess supplementation induces multiple defects in mouse epididymal spermatozoa: Understanding the role of selenium in male fertility. Int. J. Androl. 2008, 31, 438–449. [Google Scholar] [CrossRef]
- Sánchez-Gutiérrez, M.; García-Montalvo, E.; Izquierdo-Vega, J.; Del Razo, L. Effect of dietary selenium deficiency on the in vitro fertilizing ability of mice spermatozoa. Cell Biol. Toxicol. 2008, 24, 321–329. [Google Scholar] [CrossRef]
- Mohammadi, S.; Movahedin, M.; Mowla, S.J. Up-regulation of CatSper genes family by selenium. Reprod. Biol. Endocrinol. 2009, 7, 126. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.; Ahmed, A.; Hassan, H.; El-Sadek, M.A.; Ghazalah, A.; Lohakare, J. Nutritional impact of nano-selenium, garlic oil, and their combination on growth and reproductive performance of male Californian rabbits. Anim. Feed Sci. Technol. 2019, 249, 37–45. [Google Scholar] [CrossRef]
- Baiomy, A.; Mohamed, A.; Mottelib, A. Effect of dietary selenium and vitamin E supplementation on productive and reproductive performance in rams. In Proceedings of the 14th International Congress of the International Society for Animal Hygiene (ISAH), Vechta, Germany, 9–23 July 2009; pp. 43–46. [Google Scholar]
- Jacyno, E.; Kawecka, M.; Kamyczek, M. Influence of inorganic Se+ vitamin E and organic Se+ vitamin E on reproductive performance of young boars. Agric. Food Sci. 2002, 11, 175–184. [Google Scholar] [CrossRef]
- Hajalshaikh, A.; Al-Hassan, M.; Mohamed, H. The influence of injectable sodium selenite on semen characteristics and testosterone concentration in Aardi goats. Indian J. Anim. Res. 2015, 49, 793–797. [Google Scholar] [CrossRef][Green Version]
- Martins, S.M.M.K.; De Andrade, A.; Zaffalon, F.G.; Parazzi, L.J.; Bressan, F.F.; Pugine, S.M.P.; Melo, M.P.d.; Chiaratti, M.; Marino, C.T.; Afonso, E.R. Organic selenium increases PHGPx, but does not affect quality sperm in raw boar semen. Livest. Sci. 2014, 164, 175–178. [Google Scholar] [CrossRef]
- Petrujkić, B.; Šefer, D.; Jovanović, I.; Jovičin, M.; Janković, S.; Jakovljević, G.; Beier, R.; Anderson, R. Effects of commercial selenium products on glutathione peroxidase activity and semen quality in stud boars. Anim. Feed Sci. Technol. 2014, 197, 194–205. [Google Scholar] [CrossRef]
- El-Sharawy, M.; Eid, E.; Darwish, S.; Abdel-Razek, I.; Islam, M.R.; Kubota, K.; Yamauchi, N.; El-Shamaa, I. Effect of organic and inorganic selenium supplementation on semen quality and blood enzymes in buffalo bulls. Anim. Sci. J. 2017, 88, 999–1005. [Google Scholar] [CrossRef]
- Lukusa, K.; Lehloenya, K. Selenium supplementation improves testicular characteristics and semen quality of Saanen bucks. Small Rumin. Res. 2017, 151, 52–58. [Google Scholar] [CrossRef]
- Anchordoquy, J.P.; Anchordoquy, J.M.; Lizarraga, R.M.; Nikoloff, N.; Pascua, A.M.; Furnus, C.C. The importance of trace minerals copper, manganese, selenium and zinc in bovine sperm–zona pellucida binding. Zygote 2019, 1–8. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Sharma, R.; Sikka, S.; Agarwal, A. Diagnostic application of total antioxidant capacity in seminal plasma to assess oxidative stress in male factor infertility. J. Assist. Reprod. Genet. 2016, 33, 627–635. [Google Scholar] [CrossRef]
- Gharagozloo, P.; Gutiérrez-Adán, A.; Champroux, A.; Noblanc, A.; Kocer, A.; Calle, A.; Pérez-Cerezales, S.; Pericuesta, E.; Polhemus, A.; Moazamian, A. A novel antioxidant formulation designed to treat male infertility associated with oxidative stress: Promising preclinical evidence from animal models. Hum. Reprod. 2016, 31, 252–262. [Google Scholar] [CrossRef]
- Anel-López, L.; Ortega-Ferrusola, C.; Martínez-Rodríguez, C.; Álvarez, M.; Borragán, S.; Chamorro, C.; Peña, F.; Anel, L.; de Paz, P. Analysis of seminal plasma from brown bear (Ursus arctos) during the breeding season: Its relationship with testosterone levels. PLoS ONE 2017, 12, e0181776. [Google Scholar] [CrossRef]
- Bertelsmann, H.; Keppler, S.; Höltershinken, M.; Bollwein, H.; Behne, D.; Alber, D.; Bukalis, G.; Kyriakopoulos, A.; Sieme, H. Selenium in blood, semen, seminal plasma and spermatozoa of stallions and its relationship to sperm quality. Reprod. Fertil. Dev. 2010, 22, 886–891. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Turek, P.J. Effects of dietary selenium on sperm motility in healthy men. J. Androl. 2001, 22, 764–772. [Google Scholar]
- Domosławska, A.; Zdunczyk, S.; Franczyk, M.; Kankofer, M.; Janowski, T. Selenium and vitamin E supplementation enhances the antioxidant status of spermatozoa and improves semen quality in male dogs with lowered fertility. Andrologia 2018, e13023. [Google Scholar] [CrossRef]
- Domosławska, A.; Zduńczyk, S.; Niżański, W.; Jurczak, A.; Janowski, T. Effect of selenium and vitamin E supplementation on semen quality in dogs with lowered fertility. Bull. Vet. Inst. Pulawy 2015, 59, 85–90. [Google Scholar] [CrossRef]
- Butt, M.A.; Shahid, M.Q.; Bhatti, J.A.; Khalique, A. Effect of Dietary Vitamin E and Selenium Supplementation on Physiological Responses and Reproductive Performance in Holstein Friesian Bulls during Humid Hot Summer. Pak. Vet. J. 2019, 1–5. [Google Scholar] [CrossRef]
- Ghorbani, A.; Moeini, M.M.; Souri, M.; Hajarian, H. Influences of dietary selenium, zinc and their combination on semen characteristics and testosterone concentration in mature rams during breeding season. J. Appl. Anim. Res. 2018, 46, 813–819. [Google Scholar] [CrossRef]
- Kirchhoff, K.; Failing, K.; Goericke-Pesch, S. Effect of dietary vitamin E and selenium supplementation on semen quality in Cairn Terriers with normospermia. Reprod. Domest. Anim. 2017, 52, 945–952. [Google Scholar] [CrossRef]
- Alonge, S.; Melandri, M.; Leoci, R.; Lacalandra, G.M.; Caira, M.; Aiudi, G.G. The Effect of Dietary Supplementation of Vitamin E, Selenium, Zinc, Folic Acid, and N-3 Polyunsaturated Fatty Acids on Sperm Motility and Membrane Properties in Dogs. Animals 2019, 9, 34. [Google Scholar] [CrossRef]
- Domosławska, A.; Zduńczyk, S.; Janowski, T. Improvement of sperm motility within one month under selenium and vitamin E supplementation in four infertile dogs with low selenium status. J. Vet. Res. 2019. [Google Scholar] [CrossRef]
- Ran, M.-X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.-N.; Zhang, M.; Zhou, G.-B.; Zhou, Y.-M.; Wu, K.; Wang, C.-D. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. Int. J. Mol. Sci. 2018, 19, 3066. [Google Scholar] [CrossRef]
- Dai, D.-H.; Qazi, I.H.; Ran, M.-X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.-B.; Angel, C.; Zeng, C.-J. Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef]
- Jamali, N.U.; Kaka, A.; Khatri, P.; Malhi, M.; Naeem, M.; Memon, A.A.; Kaleri, R.R.; Janyaro, H.; Kalhoro, D.H. Effect of in vitro Selenium Addition to the Semen Extender on the Spermatozoa Characteristics before and after Freezing in Kundhi Buffalo Bull and in vivo Fertility Rate. Pak. J. Zool. 2019, 51, 317–323. [Google Scholar] [CrossRef]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.; Hassan, M.A. Impact of selenium nano-particles in semen extender on bull sperm quality after cryopreservation. Theriogenology 2019, 126, 121–127. [Google Scholar] [CrossRef]
- Graupner, A.; Instanes, C.; Andersen, J.M.; Brandt-Kjelsen, A.; Dertinger, S.D.; Salbu, B.; Brunborg, G.; Olsen, A.-K. Genotoxic effects of two-generational selenium deficiency in mouse somatic and testicular cells. Mutagenesis 2015, 30, 217–225. [Google Scholar] [CrossRef]
- Taghizadeh, L.; Eidi, A.; Mortazavi, P.; Rohani, A.H. Effect of selenium on testicular damage induced by varicocele in adult male Wistar rats. J. Trace Elem. Med. Biol. 2017, 44, 177–185. [Google Scholar] [CrossRef]
- Gan, X.; Zhang, X.; Qiannan, E.; Zhang, Q.; Ye, Y.; Cai, Y.; Han, A.; Tian, M.; Wang, C.; Su, Z. Nano-selenium attenuates nickel-induced testosterone synthesis disturbance through inhibition of MAPK pathways in Sprague-Dawley rats. Environ. Toxicol. 2019. [Google Scholar] [CrossRef]
- Kaur, S.; Saluja, M.; Bansal, M. Bisphenol A induced oxidative stress and apoptosis in mice testes: Modulation by selenium. Andrologia 2018, 50, e12834. [Google Scholar] [CrossRef]
- Long, M.; Yang, S.; Wang, Y.; Li, P.; Zhang, Y.; Dong, S.; Chen, X.; Guo, J.; He, J.; Gao, Z. The protective effect of selenium on chronic zearalenone-induced reproductive system damage in male mice. Molecules 2016, 21, 1687. [Google Scholar] [CrossRef]
- Rashad, M.M.; Galal, M.K.; Abou-El-Sherbini, K.S.; El-Behairy, A.M.; Gouda, E.M.; Moussa, S.Z. Nano-sized selenium attenuates the developmental testicular toxicity induced by di-n-butyl phthalate in pre-pubertal male rats. Biomed. Pharmacother. 2018, 107, 1754–1762. [Google Scholar] [CrossRef]
- EL-Maraghy, S.A.; Nassar, N.N. Modulatory effects of lipoic acid and selenium against cadmium-induced biochemical alterations in testicular steroidogenesis. J. Biochem. Mol. Toxicol. 2011, 25, 15–25. [Google Scholar] [CrossRef]
- Cao, Z.; Shao, B.; Xu, F.; Liu, Y.; Li, Y.; Zhu, Y. Protective effect of selenium on aflatoxin B1-induced testicular toxicity in mice. Biol. Trace Elem. Res. 2017, 180, 233–238. [Google Scholar] [CrossRef]
- Oda, S.S.; El-Maddawy, Z.K. Protective effect of vitamin E and selenium combination on deltamethrin-induced reproductive toxicity in male rats. Exp. Toxicol. Pathol. 2012, 64, 813–819. [Google Scholar] [CrossRef]
- Dkhil, M.; Zrieq, R.; Al-Quraishy, S.; Abdel Moneim, A. Selenium nanoparticles attenuate oxidative stress and testicular damage in streptozotocin-induced diabetic rats. Molecules 2016, 21, 1517. [Google Scholar] [CrossRef]
- Rungsung, S.; Khan, A.M.; Sood, N.K.; Rampal, S.; Saini, S.P.S. Evaluation of ameliorative potential of supranutritional selenium on enrofloxacin-induced testicular toxicity. Chem.-Biol. Interact. 2016, 252, 87–92. [Google Scholar] [CrossRef]
- Zhang, X.; Gan, X.E.Q.; Zhang, Q.; Ye, Y.; Cai, Y.; Han, A.; Tian, M.; Wang, Y.; Wang, C. Ameliorative effects of nano-selenium against NiSO4-induced apoptosis in rat testes. Toxicol. Mech. Methods 2019, 1–36. [Google Scholar] [CrossRef]
- Aydin, S.; Erkekoglu, P.; Başaran, N.; Kocer-Gumusel, B. Impact of selenium status on Aroclor 1254-induced DNA damage in sperm and different tissues of rats. Toxicol. Mech. Methods 2018, 28, 252–261. [Google Scholar] [CrossRef]
- Kheradmand, N.; Kamkar, R.; Moshajjari, M.; Baazm, M. Effect of selenium and pentoxifylline on expression of CATSPER1 and 2 genes and FSH/LH levels in treated mice by dexamethasone. Andrologia 2019, e13279. [Google Scholar] [CrossRef]
- Isidori, A.M.; Pozza, C.; Gianfrilli, D.; Isidori, A. Medical treatment to improve sperm quality. Reprod. Biomed. Online 2006, 12, 704–714. [Google Scholar] [CrossRef]
- Smits, R.M.; Mackenzie-Proctor, R.; Yazdani, A.; Stankiewicz, M.T.; Jordan, V.; Showell, M.G. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2019, 14. [Google Scholar] [CrossRef]
- Steiner, A.; Hansen, K.; Diamond, M.; Coutifaris, C.; Cedars, M.; Legro, R.; Usadi, R.; Baker, V.; Coward, R.; Santoro, N. Antioxidants in the treatment of male factor infertility: Results from the double blind, multi-center, randomized controlled Males, Antioxidants, and Infertility (MOXI) trial. Hum. Reprod. 2018, 33, i30. [Google Scholar]
- Aktan, G.; Doğru-Abbasoğlu, S.; Küçükgergin, C.; Kadıoğlu, A.; Özdemirler-Erata, G.; Koçak-Toker, N. Mystery of idiopathic male infertility: Is oxidative stress an actual risk? Fertil. Steril. 2013, 99, 1211–1215. [Google Scholar] [CrossRef]
- Rezaeian, Z.; Yazdekhasti, H.; Nasri, S.; Rajabi, Z.; Fallahi, P.; Amidi, F. Effect of selenium on human sperm parameters after freezing and thawing procedures. Asian Pac. J. Reprod. 2016, 5, 462–466. [Google Scholar] [CrossRef]
- Ghafarizadeh, A.; Vaezi, G.; Shariatzadeh, M.; Malekirad, A. Effect of in vitro selenium supplementation on sperm quality in asthenoteratozoospermic men. Andrologia 2018, 50, e12869. [Google Scholar] [CrossRef]
- Scott, R.; MacPherson, A.; Yates, R. Selenium supplementation in sub-fertile human males. Br. J. Urol. 1998, 82, 76–80. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Alkan, Z.; Wong, K. Selenium supplementation does not affect testicular selenium status or semen quality in North American men. J. Androl. 2009, 30, 525–533. [Google Scholar] [CrossRef]
- Zadeh, A.A.; Arab, D.; Kia, N.S.; Heshmati, S.; Amirkhalili, S.N. The role of Vitamin E–Selenium-Folic acid supplementation in improving the sperm parameters after varicocelectomy: A randomized clinical trial. Urol. J. 2019. [Google Scholar] [CrossRef]
- Mossa, M.; Azzawi, M.; Dekhel, H. Effect of Selenium in Treatment of Male Infertility. Exp. Tech. Urol. Nephrol. 2018, 1, ETUN.000521. [Google Scholar]
- Busetto, G.M.; Koverech, A.; Messano, M.; Antonini, G.; De Berardinis, E.; Gentile, V. Prospective open-label study on the efficacy and tolerability of a combination of nutritional supplements in primary infertile patients with idiopathic astenoteratozoospermia. Arch. Ital. Urol. 2012, 84, 137–140. [Google Scholar]
- Lombardo, F.; Fiducia, M.; Lunghi, R.; Marchetti, L.; Palumbo, A.; Rizzo, F.; Koverech, A.; Lenzi, A.; Gandini, L. Effects of a dietary supplement on chronic pelvic pain syndrome (Category IIIA), leucocytospermia and semen parameters. Andrologia 2012, 44, 672–678. [Google Scholar] [CrossRef]
- Moslemi, M.K.; Tavanbakhsh, S. Selenium-vitamin E supplementation in infertile men: Effects on semen parameters and pregnancy rate. Int. J. Gen. Med. 2011, 4, 99–104. [Google Scholar] [CrossRef]
- Rayman, M.P.; Winther, K.H.; Pastor-Barriuso, R.; Cold, F.; Thvilum, M.; Stranges, S.; Guallar, E.; Cold, S. Effect of long-term selenium supplementation on mortality: Results from a multiple-dose, randomised controlled trial. Free Radic. Biol. Med. 2018, 127, 46–54. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | General Description [27] | Relevant Literature |
|---|---|---|
| Selenomethionine | It is a Se analogue of amino acid, methionine; this form is typically obtained from plant sources (in particular the cereal grains), Se yeast, and other Se supplements. Also reported in animal foodstuffs. It is inserted non-specifically into body proteins in position of methionine. The supplements containing selenomethionine, therefore, are considered to have more bioavailable Se. | [18,26,27,28,29,30,31,32,33,34,35,36] |
| Selenocysteine | This form is found in animal foods (from their selenoproteins) and is Se analogue of the cysteine (amino acid). | [18,26,27,30,32,33,36] |
| Selenoneine (2-selenyl-Nα,Nα,Nα-trimethyl-L-histidine) | This is a newly disclosed major Se species in fish i.e., tuna and mackerel; however, at lower levels it is found in squid, tilapia, pig, and chickens. This form possesses strong radical-scavenging activity. | [27,37] |
| Se-methyl-selenocysteine and γ-glutamyl-Se-methyl-selenocysteine | These are also obtained from plant sources viz. Se-enriched yeast, garlic, onions, and broccoli. Generally considered as detoxification products, particularly formed in Se-accumulators and plants of the Brassica and Allium families. It is metabolized to methyl selenol, which is believed to possess anti-tumor effects. No bioavailability data exist for these [18]. | [18,26,27,35,38,39] |
| Sodium selenite and selenate | The components of dietary supplements; selenate at times appears in water supplies. Some selenate is found in plant sources (cabbage) and fish. | [18,26,27,32,33,35,36,40,41,42] |
| Species | RDA |
|---|---|
| Adult men | M: 55 µg/d *; F: 55 µg/d * |
| Sheep and Goat | 100–200 µg/kg dry matter of feed/d |
| Pig | 150–300 µg/kg dry matter of feed/d |
| Horse | 100 µg/kg dry matter of feed/d |
| Donkey | 150 µg/100 kg BW |
| Dairy cow | 100 µg/kg dry matter of feed/d |
| Beef cow | 300 µg/kg dry matter of feed/d |
| Bovine calf | 100 µg/kg dry matter of feed/d |
| Camel | 400–800 µg/d |
| Selenoprotein Gene * | Symbol [12] | General Description/Function [8,27,43,44,47,56] | mRNA * | Protein * | Relevance to Male Reproductive Function |
|---|---|---|---|---|---|
| Glutathione peroxidase 4 | Gpx4 | Detoxification of lipid hydroperoxides, Antioxidant in membranes, functions as structural protein in sperm, also implicated in apoptosis | ++++ | ++++ | Structural protein of sperm midpiece mitochondrial sheath and involved in sperm chromatin condensation [75]. Implication in male fertility [68]. |
| Thioredoxin-glutathione reductase | Txnrd3 (TGR) | Part of the thioredoxin system, Antioxidant function, redox regulation, cell signaling | + | + | Implicated in formation of disulfide bond and sperm maturation process [76]. Expressed in post-pubertal testis, particularly abundant in elongated spermatids at the site of mitochondrial sheath formation [75]. |
| Selenoprotein P | Selenop | Primarily responsible for Se transport and also performs antioxidative role. Considered as a major contributor to plasma Se and a reliable biomarker of Se status. Its deficiency causes infertility characterized by abnormal sperm in mice | + | + | Implicated in male fertility [77,78]. Implicated in transport of Se to spermatogenic cells [71]. Essential for sperm development in mice [79]. |
| Selenoprotein V | Selenov | Largely unidentified, potential role in redox regulation | + | n.d. | Specifically expressed in rodent testes [80]. In situ hybridization trials have demonstrated the expression of Selenov mRNA in seminiferous tubules in mouse, however, its precise function in spermatogenesis is largely unexplored [48,80]. |
| Selenoprotein W | Selenow | Antioxidant protection | + | + | n.d. * |
| Selenoprotein K | Selenok | Possible antioxidant protection in cardiomyocytes, Endoplasmic reticulum transmembrane protein | ++ | n.d. | n.d. * |
| Selenoprotein F | Selenof | Role in cell apoptosis and mediation of chemo-preventive effects of Se | + | n.d. | n.d. * |
| Selenoprotein S | Selenos | Cellular redox balance, Possible influence in inflammatory response | + | n.d. | n.d. * |
| Selenophosphate synthetase 2 | Sephs2 | Required for biosynthesis of selenophosphate, a precursor of selenocysteine, and thus for selenoprotein synthesis | + | n.d. | n.d. |
| Model | Treatment | Key Observations Reported | Ref. |
|---|---|---|---|
| Sprague-Dawley rats | Se nanoparticles at supranutritional levels (0.2, 0.4, or 0.8 mg Se per kg body weight) | Sperm parameters such as, sperm concentration, motility, and morphological features were all improved at supranutritional levels. However, these parameters were significantly affected when rats were supplemented with higher levels (nonlethal level) of Se nanoparticles i.e., at 2.0, 4.0, or 8.0 mg Se per kg body weight. | [110] |
| Sprague-Dawley rats | Treated with inorganic Se [0.01(deficient); 0.25 (adequate); 3 (excess); or 5 (excess) mg per kg] for four weeks | The U-shaped response of dietary Se was observed on DNA damage and sperm quality. Se deficiency showed a lower expression of sensitive antioxidant selenoproteins (Gpx1 and Txnrds). However, excessive doses of Se impaired sperm quality and this was linked with reduced mRNA expression of nGpx4. | [111] |
| Mouse | Se-supplement (inorganic Se (0.3 μg/g Se) or organic Se-enriched probiotics (containing 0.3 μg/g Se) given for 75 days | Organic Se co-supplemented with probiotics significantly improved male fertility in mice. The ameliorated fertility index included the parameters such as, reduced testicular tissue injury, increased levels of serum testosterone, and improved sperm indices in Se-supplemented group. As such, these improved fertility-related parameters were ascribed to be the result of the antioxidant function of Se. | [112] |
| Mouse | 0.2 ppm sodium selenite; 1.0 ppm sodium selenite | Mice in both groups showed an increased occurrence of mitochondria- and plasma membrane-related defects, and DNA damage in sperm. However, these damages were more pronounced in mice exposed to Se-deficient feed. | [113] |
| Mouse | Se-deficient diet (0.02 ppm) Se-sufficient (0.2 ppm); organic Se | Sperm from Se-deficient mice demonstrated vitiated chromatin condensation, declined in vitro fertilization ability and increased lipid peroxidation (LPO) in both testes and sperm compared to the Se-sufficient mice. | [114] |
| Mouse | Se-deficient (0.02 ppm) Se-excess (0.2 ppm); yeast-based Se. Mice were fed for 4 months | Se concentration and GPX activity (in testis) were significantly reduced. The fertility percentage and size of litter were both reduced in Se-deficient group. | [96] |
| Aged mice | Inorganic Se 0.2 mg/kg body weight | Improved sperm parameters and increased expression of CatSper genes were observed in Se-treated group. | [115] |
| Rabbit | Treated with Se nanoparticles (400 μg/kg) for 60 days | Improved serum testosterone levels were recorded in Se-treated group compared to the control. Besides, improved ejaculate volume and sperm quality parameters such as, sperm morphology, viability were observed. | [116] |
| Ram | 0.5 ppm organic Se; 0.2 ppm organic Se | A significantly higher concentration of Se and improved ejaculate and sperm quality were observed in seminal plasma of rams exposed to a feed containing 0.5 ppm organic Se compared to those who received 0.2 ppm organic Se. | [117] |
| Boar | Organic Se (0.2 mg per kg); Inorganic Se (0.2 mg per kg) | Ejaculate quality and sperm parameters were significantly improved in boars following dietary supplementation of organic Se (0.2 mg per kg) compared to those treated with sodium selenite at the same dose. | [118] |
| Aardi buck | Sodium selenite 0.1 mg/kg, Sodium selenite 0.05 mg/kg | Improved sperm count and motility was observed in both Se-treated groups. However, relatively better outcomes were observed in 0.1 mg/kg group. | [119] |
| Boar | 0.5 ppm organic Se | Following 11 weeks of feeding trail, organic Se supplementation increased glutathione peroxidase 4 (GPX4) activity (raw semen) and number of seminal doses in boars. | [120] |
| Boar | 0.3 ppm organic Se; 0.3 ppm inorganic Se | Following 12 weeks of Se supplementation, Se content and GPX activity were increased in semen of boars treated with organic and inorganic Se. Besides, semen quality parameters namely semen concentration and progressive motility of sperm were improved compared to the control group without Se. Improved resistance of liquid stored semen to hypo-osmotic shock and thermal tests, and improved fertility rates were observed in semen of boars treated with Se. All mentioned indices were slightly higher in the organic Se group compared to the inorganic group. | [121] |
| Buffalo bulls | 10 mg organic Se/animal twice a week; and 10 mg inorganic Se/animal twice a week | Three months long Se supplementation significantly improved the sperm quality parameters (ejaculate volume, sperm motility, concentration, and morphology) in buffalo bulls. Besides, testosterone concentrations were also increased in Se-treated groups. | [122] |
| Saanen bucks | Inorganic Se 0.34 mg/kg body weight supplemented at ten-day intervals for three months | Se supplementation improved the testicular biometry and sperm parameters. GPX activity, plasma testosterone and LH levels significantly increased in Se-treated group from days 40 to 80 compared to the control group. These indices reached peak reached peak at day 80 of the trial. | [123] |
| Bovine bull | In vitro fertilization (IVF) medium supplemented with Se (100 ng/mL) | A significant increase in sperm mitochondrial activity was observed after 1 h of incubation in Se-supplemented IVF medium. Moreover, Se supplementation after 2 h of incubation showed an increase in HOST-positive (hypo-osmotic swelling test) sperm and sperm acrosome integrity. Increased number of sperm bound to zona pellucida (ZP) was observed in Se-treated group compared to the control. | [124] |
| Animal Model and Number | Treatment Regime and Duration | Key Findings | Ref. |
|---|---|---|---|
| Male CD-1 mice (n = 12 per experimental group) | Fertilix® (CellOxess, Princeton, NJ, USA) was supplemented for two months. (Se 55 μg, zinc 7.5–11 mg, Full spectrum natural vitamin E 104–290 mg, Lycopene 7.5–15 mg Carnitine blend 200–800 mg Folic acid 400–500 mg Vitamin C 30–90 mg). | Eight weeks long pretreatment with the antioxidant formulation completely protected oxidative stress-induced DNA damage in Gpx5 KO mice sperm. In mouse models of scrotal heat stress, only 35% (19/54) of female mice became pregnant resulting in 169 fetuses with 18% fetal resorption (30/169). Conversely, in antioxidant pretreated group 74% (42/57) of female mice became pregnant, resulting in 427 fetuses with 9% fetal resorption (38/427). | [126] |
| Four infertile male dogs with low blood Se levels (86.0–165.0 μg/L) | Organic Se 0.6 mg/kg and vitamin E (5 mg/kg) orally supplemented for 60 days. | Treated dogs showed improved sperm parameters. Increase in blood Se concentration (401 μg/L) was observed at the end of trial. When these dogs were used for matting purpose, bitches successfully conceived and gave birth to 4–6 pups. | [136] |
| Sixteen healthy normospermic dogs (two patients were excluded after adaptation period) | A supplement comprising of Se 0.27 mg/kg vitamin E 250 mg/kg, vitamin B9 1.5 mg/kg, zinc 180 mg/kg, and n-3 PUFA 0.5%, given for 90 days. | In treated group, sperm quality parameters i.e., total sperm count, concentration, sperm vitality and membrane integrity were significantly improved compared to the control group. | [135] |
| Model | Experimental Condition/Treatment Regime | Relevant Results | Ref. |
|---|---|---|---|
| Rats | Se nanoparticles (0.2 and 0.5 mg/kg/d) supplementation ameliorated developmental testicular toxicity induced by maternal exposure of di-n-butyl phthalate (DBP) in Pre-pubertal male rat offspring. Note: Pregnant female rats treated from gestation day 12 to postnatal day 14 day with two doses of Se-nanoparticles (0.2 and 0.5 mg/kg/d) against developmental testicular toxicity induced by DBP (500 mg/kg/d). | Maternal Se treatment significantly increased mRNA expression of Gpx and Sod, Insl3, and Mr in pre-pubertal male rat offspring. Malondialdehyde (MDA) and GSH levels were also significantly reduced and increased, respectively in testicular tissue. Besides, histological assessment revealed that damage in testicular parenchyma was also ameliorated. | [146] |
| Wistar rats | Cadmium-exposed rats treated with Se (0.35 mg per kg body weight) for 28 days. | The activities of testosterone biosynthesis-related and antioxidant enzymes, levels of steroid hormones, and testicular Se levels were adequately ameliorated compared to the Cd-exposed rats. In addition, Se treatment alleviated, at least partly, Cd-induced damage to architecture of testis in rats. Se-treatment also modulated the key testicular injury-related marker enzymes including LDH, SDH, G6PD, G6Pase, ACP, ALP, and AST. | [147] |
| Kunming mice | Aflatoxin B1-Exposed mice treated with inorganic Se (0.2 and 0.4 mg/kg) for 45 days. | Se-treatment at both doses (0.2 and 0.4 mg/kg) significantly ameliorated the sperm quality parameters such as, morphology, concentration and motility compared to the aflatoxin B1-exposed group. Levels of reactive oxygen species (ROS), MDA were significantly decreased, and activity of Gpx was improved. The level of serum testosterone and protein expression of testosterone synthesis enzymes StAR, P450scc, and 17β-HSD were significantly improved in Se-treated groups. | [148] |
| Albino rats | Oral deltamethrin-exposed rats treated with combinatory supplementation of Se and vitamin E (1.2 mg/kg body weight Viteselen®, containing 1.67 mg sodium selenite + 150 mg vitamin E/mL). | Se treatment significantly ameliorated the sperm quality characteristics, improved the levels of testosterone and testicular GSH, and reduced MDA levels. Similarly, Se-treated group showed markedly improved spermatogenesis and histo-architecture of testis parenchyma compared to the deltamethrin-exposed group. | [149] |
| Rats | Streptozotocin-exposed diabetic rats treated with Se nanoparticles (0.1 mg per kg body weight). | Se-treated group showed improved antioxidant status and serum testosterone levels. Expression of apoptosis-related genes i.e., Bax and Bcl-2 was also significantly altered. Histological assessment revealed that Se-treatment significantly ameliorated the testicular damage caused by streptozotocin exposure, which was evident by an increased number of spermatogenic cells in the seminiferous parenchyma of rats. | [150] |
| Wistar rats | Enrofloxacin-exposed rats treated with supranutritional Se (dose not reported by authors) for 21 days. | Se co-administration moderately improved the activity of antioxidant enzymes in testicular tissue and reduced the levels of LPO. Sperm parameters such as, total count, viability were also partly improved. | [151] |
| SD rats | Nickel sulfate-exposed rats treated with Se-nanoparticles (0.5, 1, 2 mg Se/kg body weight) for 14 days. | Se-treatment adequately alleviated testicular damage in Ni-exposed rats. GPX activity was improved MDA levels were reduced in testes. Besides, the rate of apoptosis was significantly decreased in Se-treated group compared to the Ni-exposed rats. A significant decline was observed in caspase-3 positive cells. Se-treatment significantly decreased mRNA and protein expression of Bak, cytochrome c, and caspase-9 in the testis, and increased the expression of Bcl-2. These effects were more pronounced in rats treated with higher doses (2 mg) of Se. | [152] |
| SD rats | Aroclor 1254-exposed rats treated with Se (1 mg Se/kg) Both control and Se-deficient rats were used in this study. | DNA damage was more pronounced in Se-deficient rats exposed to Aroclor 1254. Se supplementation significantly ameliorated DNA damage in sperm in both normal Aroclor 1254-exposed and Se-deficient rats. | [153] |
| NMRI mice | Dexamethasone-treated mice treated with Se (0.3mg/kg) for 7 days. | Se-treatment increased the mRNA expression of Catsper1 and Catsper2 in testes. Improvements were also observed in serum levels of LH. It should be noted that Catsper1 and Catsper2 are implicated in important sperm functions. | [154] |
| Wistar rats | Experimentally varicocelized male rats supplemented with inorganic Se (0.05, 0.1, 0.2, and 0.4 mg per kg body weight). | Sperm quality parameters, antioxidative status were significantly ameliorated, and damage to histo-architecture of testes was significantly lower, and Johnsen’s score was also adequately improved compared to the varicocelized control rats. | [142] |
| Condition | Study Type and Location | No. of Subjects and Age | Type and Duration of Treatment | Key Results | Reference |
|---|---|---|---|---|---|
| Subjects diagnosed with varicocele and underwent sub-inguinal varicocelectomy | Randomized, single blind clinical trial (intervention vs. control) (Iran) | n = 60 infertile men Age: not reported | Oral supplementation of Se (200 ug), Folic acid (5 mg) and vitamin E (400IU) (6 months) | Sperm parameters were improved compared to the control group. | Zadeh et al. [163] |
| Men with male factor infertility | Multi-center, double blind, randomized, placebo-controlled trial conducted in eight American fertility centers (USA) | n = 174 couples Age of males: not reported | 500 mg vitamin C, 2000IU vitamin D3, 400IU vitamin E, 1 mg folic acid, 20 mg zinc, 200 μg Se, and 1000 mg L-carnitine (3 months) | No improvements were observed in semen quality parameters or DNA fragmentation. No improvements were observed in conception rate (in vivo). | Steiner et al. (2018) [157] |
| Infertile men | Longitudinal study (Iraq) | n = 12 Age: not reported | 50 μg Se (3 months) | Improved sperm count, motility, viability, sperm morphology, and ejaculate volume. | Mossa et al. (2018) [164] |
| Infertile patients with idiopathic astenoteratozoospermia | Prospective open-label study (Italy) | n = 114 (96 completed the study) Age: 21–46 years | Combination treatment including Se 50 mcg + L-carnitine 145 mg + acetyl-L-carnitine 64 mg + fructose 250 mg + citric acid 50 mg + coenzyme Q10 20 mg + zinc 10 mg + ascorbic acid 90 mg + cyanocobalamin 1.5 mcg + folic acid 200 mcg (4 months) | Improvements were observed in sperm parameters such as progressive motility and treatment was well tolerated. Whereas 16 patients achieved pregnancy during the study. | Busetto et al. (2012) [165] |
| Chronic prostatis | Prospective open-label study (Italy) | n = 60 Age: 30–55 years | Se 82.3 μg + lycopene (1.5 mg) + epigallocatechin gallate (250 mg) + ellagic acid (250 mg) + zinc (20 mg) (30 subjects) vs. No treatment (30 subjects) (6 months) | Improved sperm quality parameters (motility and morphology) were observed. Improvements were observed in leucocytospermia and Chronic Prostatitis Symptom Index. | Lombardo et al. (2012) [166] |
| Idiopatic asthenoteratospermia | Prospective single-arm study (Iran) | n = 690 Age: 20–45 years | 200 μg/d L-selenomethionine + 400 IU/d Vit E (3 months) | Improved sperm motility, morphology and pregnancy rate were observed. | Moslemi and Tavanbakhsh (2011) [167] |
| Healthy men | Double blind RCT (USA) | n = 42 Age: 18–45 years | 300 μg/d Se-yeast or placebo (11 months) | No effects on seminal parameters were observed. | Hawkes et al. (2009) [162] |
| Idiopatic asthenoteratospermia | Doubleblind RCT (Iran) | n = 468 Age: 25–48 years | 200 μg Se/d (116 subjects), Or 600 mg NAC/d (118 subjects), Or 200 μg Se+ 600 mg NAC/d (116 subjects) Or Placebo (118 subjects) (6 months) | Improved sperm count, motility and morphology were observed (both in Se + NAC and Se alone groups). | Safarinejad and Safarinejad (2009) [62] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qazi, I.H.; Angel, C.; Yang, H.; Zoidis, E.; Pan, B.; Wu, Z.; Ming, Z.; Zeng, C.-J.; Meng, Q.; Han, H.; et al. Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences. Antioxidants 2019, 8, 268. https://doi.org/10.3390/antiox8080268
Qazi IH, Angel C, Yang H, Zoidis E, Pan B, Wu Z, Ming Z, Zeng C-J, Meng Q, Han H, et al. Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences. Antioxidants. 2019; 8(8):268. https://doi.org/10.3390/antiox8080268
Chicago/Turabian StyleQazi, Izhar Hyder, Christiana Angel, Haoxuan Yang, Evangelos Zoidis, Bo Pan, Zhenzheng Wu, Zhang Ming, Chang-Jun Zeng, Qingyong Meng, Hongbing Han, and et al. 2019. "Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences" Antioxidants 8, no. 8: 268. https://doi.org/10.3390/antiox8080268
APA StyleQazi, I. H., Angel, C., Yang, H., Zoidis, E., Pan, B., Wu, Z., Ming, Z., Zeng, C.-J., Meng, Q., Han, H., & Zhou, G. (2019). Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences. Antioxidants, 8(8), 268. https://doi.org/10.3390/antiox8080268