Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L.

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Obtention of Phenolic-Rich Extracts
2.3. Evaluation of Radical Scavenging and Inhibition of 5-Lipoxygenase Activities
2.3.1. Reducing Power, DPPH• Scavenging Activity
2.3.2. NO• Scavenging Test
2.3.3. Inhibition of 5-Lipoxygenase (5-LOX)
2.4. Evaluation of Biological Activity in Cellular Assays
2.4.1. Cell Culture and Treatments
2.4.2. MTT Assay
2.4.3. Intracellular ROS production
2.5. Analysis of the Phenolic Compounds
2.6. Statistical Analysis
3. Results and Discussion
3.1. Antioxidant and Anti-Inflammatory Properties (in Chemical Models)
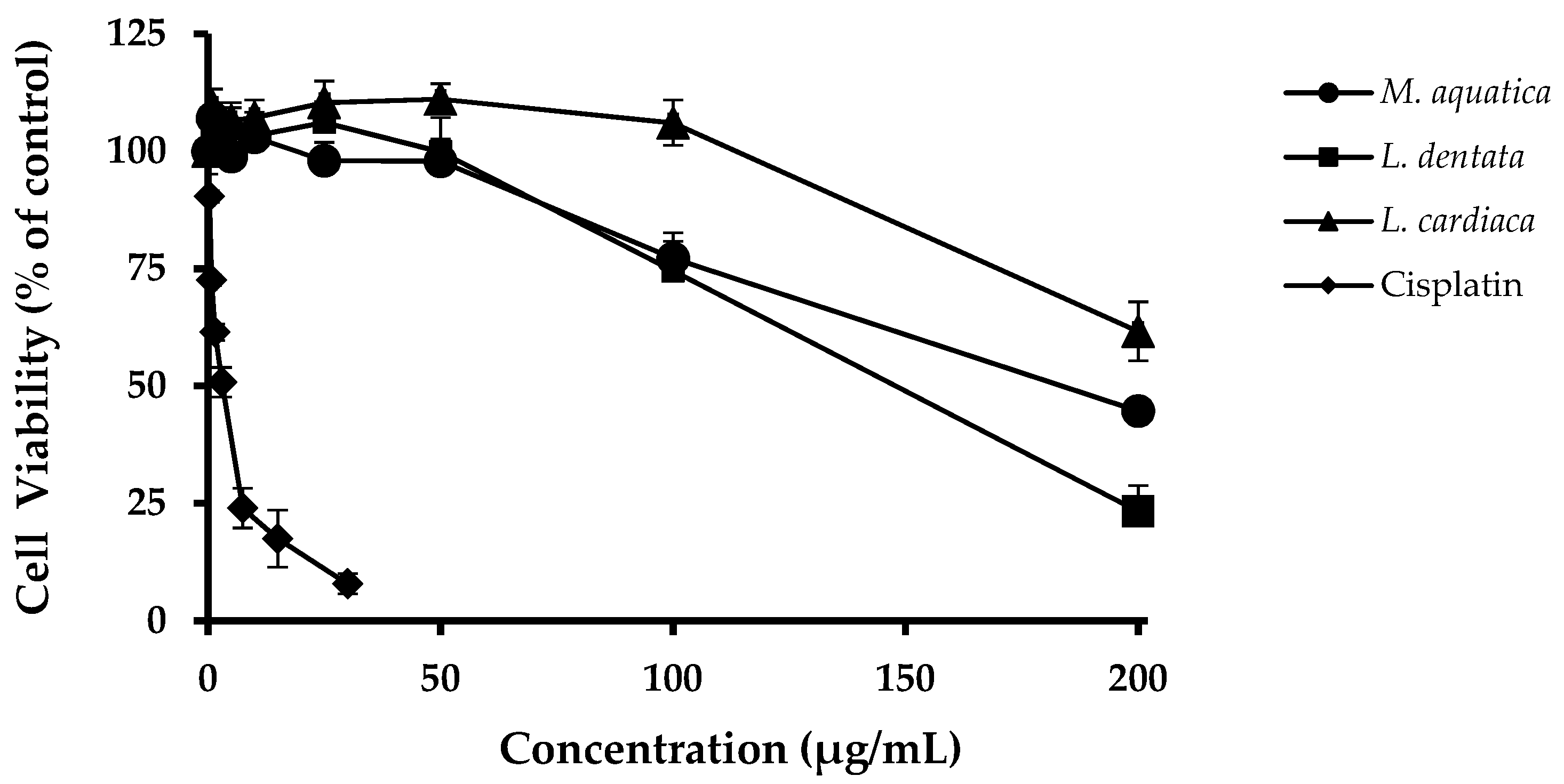
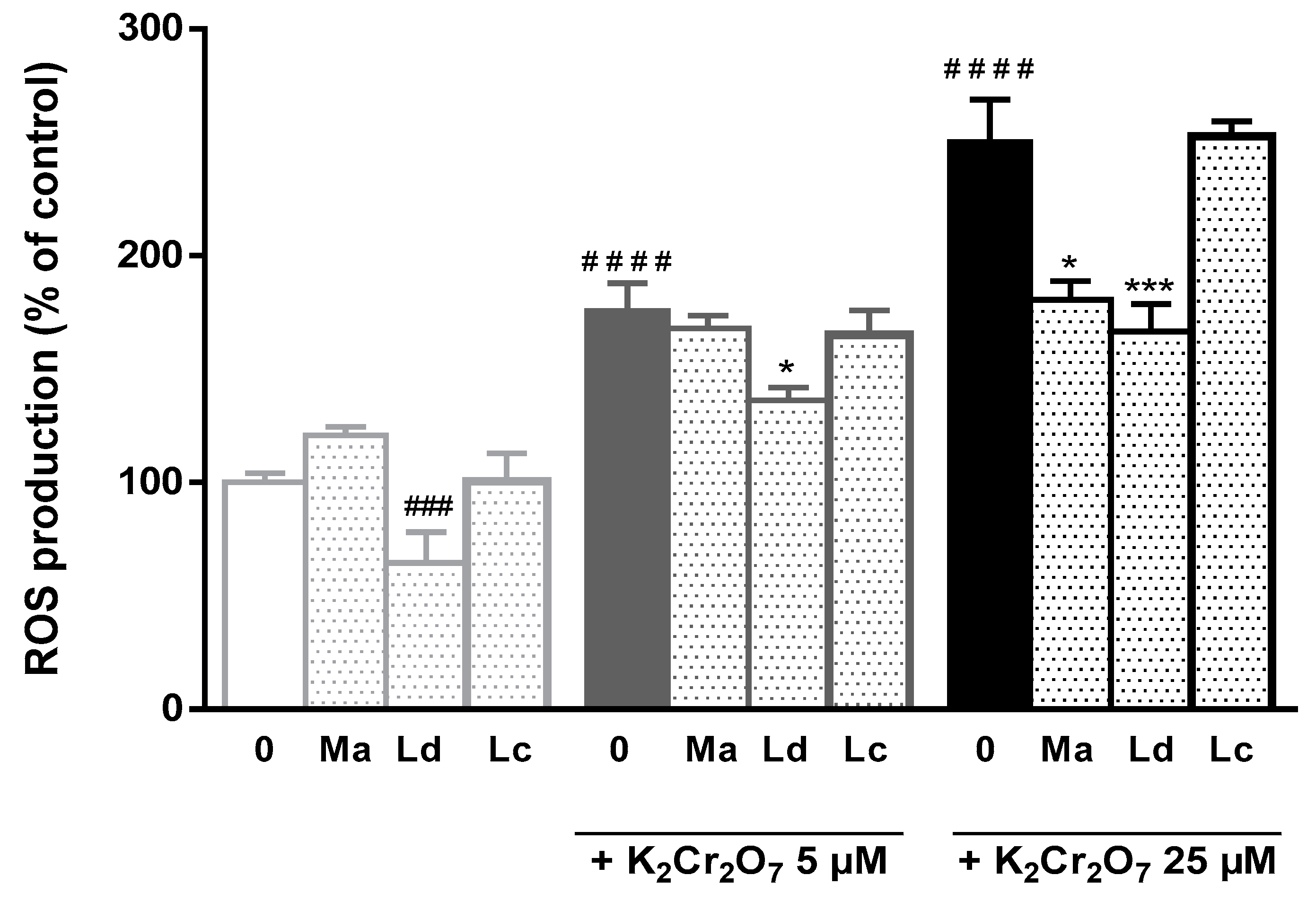
3.2. Hepatoprotective Activities in HepG2 Cells
3.3. Phenolic Characterization of M. aquatica, L. dentata and L. cardiaca Hydroethanolic Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Marin, J.J.G.; Briz, O.; Herraez, E.; Lozano, E.; Asensio, M.; Di Giacomo, S.; Romero, M.R.; Osorio-Padilla, L.M.; Santos-Llamas, A.I.; Serrano, M.A.; et al. Molecular bases of the poor response of liver cancer to chemotherapy. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Li, S.; Tan, H.Y.; Wang, N.; Tsao, S.; Feng, Y. Current Status of Herbal Medicines in Chronic Liver Disease Therapy: The Biological Effects, Molecular Targets and Future Prospects. Int. J. Mol. Sci. 2015, 16, 28705–28745. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Cheung, F.; Hong, M.; Feng, Y. The Potential and Action Mechanism of Polyphenols in the Treatment of Liver Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 8394818. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Alves-silva, J.M.; Pereira, O.R.; Cardoso, S.M. Antioxidant Capacities of Flavones and Benefits in Oxidative-Stress Related Diseases. Curr. Top. Med. Chem. 2015, 15, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Liu, C.; Gao, L.; Xu, X.; Zhu, N.; Cao, L. Hepatoprotective Effect of Aqueous Extract from the Seeds of Orychophragmus violaceus against Liver Injury in Mice and HepG2 Cells. Int. J. Mol. Sci. 2017, 18, 1197. [Google Scholar] [CrossRef]
- Stafford, G.I.; Pedersen, P.D.; Jäger, A.K.; Staden, J. Van Monoamine oxidase inhibition by southern African traditional medicinal plants. S. Afr. J. Bot. 2007, 73, 384–390. [Google Scholar] [CrossRef]
- Conforti, F.; Sosa, S.; Marrelli, M.; Menichini, F.; Statti, G.A.; Uzunov, D.; Tubaro, A.; Menichini, F.; Loggia, R.D. In vivo anti-inflammatory and in vitro antioxidant activities of Mediterranean dietary plants. J. Ethnopharmacol. 2008, 116, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Olsen, H.T.; Stafford, G.I.; Van Staden, J.; Christensen, S.B.; Anna, K.J. Isolation of the MAO-inhibitor naringenin from Mentha aquatica L. J. Ethnopharmacol. 2008, 117, 500–502. [Google Scholar] [CrossRef]
- Lis-Balchin, M. New research into Lavandula species, hybrids and cultivars. In Lavander The Genus Lavanda; Hardman, R., Ed.; Taylor & Francis: London, UK, 2002; pp. 206–213. [Google Scholar]
- Khalil, A.M.; Ashy, M.A.; Eltawil, B.A.H.; Tawfiq, N.I. Constituents of local plants. 5. Coumarin and triterpenoid constituents of Lavandula-dentata L plant. Pharmazie 1979, 34, 564–565. [Google Scholar]
- Yang, Z.; Pan, S.; Huo, K.; Wu, B.; Chao, Z. Molecular Analysis of Leonurus Species in China Based on ITS and mat K Sequences. Am. J. Chin. Med. 2011, 39, 411–422. [Google Scholar] [CrossRef]
- Ferhat, M.; Erol, E.; Beladjila, K.A.; Çetintaş, Y.; Duru, M.E.; Öztürk, M.; Kabouche, A.; Kabouche, Z. Antioxidant, anticholinesterase and antibacterial activities of Stachys guyoniana and Mentha aquatica. Pharm. Biol. 2017, 55, 324–329. [Google Scholar] [CrossRef] [PubMed]
- López, V.; Akerreta, S.; Casanova, E.; García-mina, J.M.; Cavero, R.Y. In Vitro Antioxidant and Anti-rhizopus Activities of Lamiaceae Herbal Extracts. Plants Food Hum. Nutr. 2007, 62, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Benabdallah, A.; Rahmoune, C.; Boumendjel, M.; Aissi, O.; Messaoud, C. Total phenolic content and antioxidant activity of six wild Mentha species (Lamiaceae) from northeast of Algeria. Asian Pac. J. Trop. Biomed. 2016, 6, 760–766. [Google Scholar] [CrossRef]
- Kosar, M.; Dorman, H.; Baser, K.; Filtunen, M. Screening of Free Radical Scavenging Compounds in Water Extracts of Mentha Samples Using a Postcolumn Derivatization. J. Agric. Food Chem. 2004, 52, 5004–5010. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, T.; Melzig, M.F. Medicinal Plants Traditionally Used for Treatment of Obesity and Diabetes Mellitus—Screening for Pancreatic Lipase and α-Amylase Inhibition. Phyther. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Sariri, R.; Seifzadeh, S.; Sajedi, R.H. Anti-tyrosinase and antioxidant activity of Lavandula sp. extracts. Pharmacol. Online 2009, 3, 319–326. [Google Scholar]
- Algieri, F.; Rodriguez-Nogales, A.; Vezza, T.; Garrido-Mesa, J.; Garrido-Mesa, N.; Utrilla, M.P.; González-Tejero, M.R.; Casares-Porcel, M.; Molero-Mesa, J.; del Mar Contreras, M.; et al. Anti-inflammatory activity of hydroalcoholic extracts of Lavandula dentata L. and Lavandula stoechas L. J. Ethnopharmacol. 2016, 190, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.; Moradi, A.; Salaritabar, A.; Hadjiakhoondi, A.; Khanavi, M. Determination of total phenolic and flavonoid contents of Leonurus cardiaca. Res. J. Biol. Sci. 2010, 5, 484–487. [Google Scholar]
- Sadowska, B.; Micota, B.; Rozalski, M.; Redzynia, M.; Rozalski, M. The immunomodulatory potential of Leonurus cardiaca extract in relation to endothelial cells and platelets. Innate Immun. 2017, 23, 285–295. [Google Scholar] [CrossRef]
- López, V.; Martín, S.; Gómez-serranillos, M.P.; Carretero, M.E.; Jäger, A.K.; Calvo, M.I. Neuroprotective and Neurochemical Properties of Mint Extracts. Phytother. Res. 2010, 874, 869–874. [Google Scholar]
- Conforti, F.; Ioele, G.; Statti, G.A.; Marrelli, M.; Ragno, G.; Menichini, F. Antiproliferative activity against human tumor cell lines and toxicity test on Mediterranean dietary plants. Food Chem. Toxicol. 2008, 46, 3325–3332. [Google Scholar] [CrossRef]
- Flemmig, J.; Noetzel, I.; Arnhold, J.; Rauwald, H. Leonurus cardiaca L. herb extracts and their constituents promote lactoperoxidase activity. J. Funct. Foods 2015, 17, 328–339. [Google Scholar] [CrossRef]
- Willer, H.; Schaack, D.; Lernoud, J. Organic Farming and Market Development in Europe and the European Union. In The World of Organic Agriculture—Statistics and Emerging Trends 2017; Organics International: Bonn, Germany, 2017. [Google Scholar]
- American Herbal Products Association and Organic Trade Association. Guidance on Formulation and Marketing of Organic Dietary Supplements under the National Organic Program; AHPA: Silver Spring, MD, USA; QAI: San Diego, CA, USA; OTA: Washington, DC, USA, 2017.
- Barański, M.; Średnicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, C.; et al. Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analyses. Br. J. Nutr. 2014, 112, 794–811. [Google Scholar]
- Afonso, A.F.; Pereira, O.R.; Válega, M.; Silva, A.M.S.; Cardoso, S.M. Metabolites and biological activities of Thymus zygis, Thymus pulegioides, and Thymus fragrantissimus grown under organic cultivation. Molecules 2018, 23, 1514. [Google Scholar] [CrossRef]
- Pereira, O.R.; Domingues, M.R.M.; Silva, A.M.S.; Cardoso, S.M. Phenolic constituents of Lamium album: Focus on isoscutellarein derivatives. Food Res. Int. 2012, 48, 330–335. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef]
- Pereira, O.R.; Macias, R.I.R.; Perez, M.J.; Marin, J.J.G.; Cardoso, S.M. Protective effects of phenolic constituents from Cytisus multiflorus, Lamium album L. and Thymus citriodorus on liver cells. J. Funct. Foods 2013, 5, 1170–1179. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Neto, R.T.; Silva, A.M.S.; Cardoso, S.M. Health-promoting effects of Thymus herba-barona, Thymus pseudolanuginosus, and Thymus caespititius decoctions. Int. J. Mol. Sci. 2017, 18, 1879. [Google Scholar] [CrossRef]
- Briz, O.; Serrano, M.A.; Macias, R.I.R.; Marin, J.J.G. Overcoming cisplatin resistance in vitro by a free and liposome-encapsulated bile acid derivative: Bamet-R2. Int. J. Cancer. 2000, 88, 287–292. [Google Scholar] [CrossRef]
- Ebrahimzadeh, M.A.; Nabavi, S.F.; Nabavi, S.M.; Eslami, B.; Asgarirad, H. In vitro antioxidant and free radical scavenging activity of Leonurus cardiaca subsp. Persicus, Grammosciadium platycarpum and Onosma demawendicum. Afr. J. Biotechnol. 2010, 9, 8865–8871. [Google Scholar]
- Matkowski, A.; Tasarz, P.; Szypuła, E. Antioxidant activity of herb extracts from five medicinal plants from Lamiaceae, subfamily Lamioideae. J. Med. Plant. Res. 2008, 2, 321–330. [Google Scholar]
- Sanchez-Vicente, L.; Herraez, E.; Briz, O.; Nogales, R.; Molina-Alcaide, E.; Marin, J.J.G. Biodetection of potential genotoxic pollutants entering the human food chain through ashes used in livestock diet. Food Chem. 2016, 205, 81–88. [Google Scholar] [CrossRef]
- González, L.T.; Minsky, N.W.; Elsa, L.; Espinosa, M.; Aranda, R.S.; Meseguer, J.P.; Pérez, P.C. In vitro assessment of hepatoprotective agents against damage induced by acetaminophen and CCl4. BMC Complement. Altern. Med. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Lee, J.; Yang, J.; Jeon, J.; Jeong, H.S.; Lee, J.; Sung, J. Hepatoprotective e ff ect of esculetin on ethanol-induced liver injury in human HepG2 cells and C57BL/6J mice. J. Funct. Foods 2018, 40, 536–543. [Google Scholar] [CrossRef]
- Avila-Rojas, S.H.; Tapia, E.; Briones-Herrera, A.; Aparicio-Trejo, O.E.; León-Contreras, J.C.; Hernández-Pando, R.; Pedraza-Chaverri, J. Curcumin prevents potassium dichromate (K2Cr2O7)-induced renal hypoxia. Food Chem. Toxicol. 2018, 121, 472–482. [Google Scholar] [CrossRef]
- Pereira, O.R.; Cardoso, S.M. Overview on Mentha and Thymus Polyphenols. Curr. Anal. Chem. 2013, 9, 382–396. [Google Scholar] [CrossRef]
- Zhogova, A.A.; Perova, I.B.; Samylina, I.A.; Eller, K.I.; Ramenskaya, G.V. Identification and quantitative determination of the main biologically active substances in motherwort herb by HPLC—Mass Spectrometry. Pharm. Chem. J. 2014, 48, 461–466. [Google Scholar] [CrossRef]
- Renzulli, C.; Galvano, F.; Pierdomenico, L.; Speroni, E.; Guerra, M.C. Effects of Rosmarinic Acid against Aflatoxin B 1 and Ochratoxin-A-induced Cell Damage in a Human Hepatoma Cell Line (Hep G2). J. Appl. Toxicol. 2004, 24, 289–296. [Google Scholar] [CrossRef]
- Madushan, P.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Hewage, S.R.; Chae, S.W.; Hyun, J.W. Rosmarinic Acid Attenuates Cell Damage against UVB Radiation-Induced Oxidative Stress via Enhancing Antioxidant Effects in Human HaCaT Cells. Biomol. Ther. 2016, 24, 75–84. [Google Scholar]
- Liu, J.; Li, X.; Lin, J.; Li, Y.; Wang, T.; Jiang, Q.; Chen, D. Sarcandra glabra (Caoshanhu) protects mesenchymal stem cells from oxidative stress: A bioevaluation and mechanistic chemistry. BMC Complement. Altern. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Lin, P.; Tian, X.; Yi, Y.S.; Jiang, W. Luteolin-induced protection of H2O2-Iinduced apoptosis in PC12 cells and the associated pathway. Mol. Med. Rep. 2015, 12, 7699–7704. [Google Scholar] [CrossRef]
- Nogata, Y.; Ohta, H.; Ishii, T.; Sekiya, K. Isolation of eriocitrin (eriodictyol 7-O-rutinoside) as an arachidonate lipoxygenase inhibitor from Lumie fruit (Citrus lumia) and its distribution in Citrus species. J. Sci. Food Agric. 2007, 87, 82–89. [Google Scholar] [CrossRef]
- Lee, J.K. Anti-inflammatory Effects of Eriodictyol in Lipopolysaccharide-stimulated Raw 264.7 Murine Macrophages. Arch. Pharm. Res. 2011, 34, 671–679. [Google Scholar] [CrossRef]
- Miguel, F.; Rego, A.C.; Oliveira, C.R.; Maria, R. Antioxidant effect of flavonoids after ascorbate/Fe2+-induced oxidative stress in cultured retinal cells. Biochem. Pharmacol. 2001, 62, 111–118. [Google Scholar]
- Lou, H.; Jing, X.; Ren, D.; Wei, X.; Zhang, X. Neurochemistry International Eriodictyol protects against H2O2-induced neuron-like PC12 cell death through activation of Nrf2/ARE signaling pathway. Neurochem. Int. 2012, 61, 251–257. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | Yield (%) | DPPH Scavenging (μg/mL) | Reducing Power (μg/mL) | NO• Scavenging (μg/mL) | 5-LOX Inhibition (μg/mL) |
|---|---|---|---|---|---|
| M. aquatica | 11.3 | 8.1 ± 1.3 a | 51.9 ± 12.6 a | 217.0 ± 19.0 a | 174.5 ± 30.5 a |
| L. dentata | 3.2 | 11.6 ± 1.1 b | 78.9 ± 2.6 b | 879.3 ± 192.8 b | 237.9 ± 15.2 b |
| L. cardiaca | 3.7 | 18.3 ± 1.5 c | 94.7 ± 12.1 b | >1000 | >1000 |
| AA | - | 2.5 ± 0.4 d | - | 92.0 ± 7.3 c | 7.8 ± 1.0 c |
| BHA | - | - | 27.1 ± 1.2 c | - | - |
| - | Plant Extract | ||||||
|---|---|---|---|---|---|---|---|
| M. aquatica | L. dentata | L. cardiaca | |||||
| Peak | RT (min) | λmax (nm) | Compound | ESI—MSn Fragmentation (m/z) | Phenolic Content (μg/mg of extract) | ||
| Caffeic acid derivatives | |||||||
| 1 | 8.2 | 290,329 | Caffeic acid glc | 341→179→135 | - | - | 3.7 ± 0.8 |
| 16 | 21.1 | 290,328 | Rosmarinic acid | 359→179→135, 161 | 64.2 ± 8.8 | 67.8 ± 6.7 | - |
| Phenylethanoid glycosides | |||||||
| 5 | 16.7 | 290,329 | Lavandulifolioside | 755→593→461→315 | - | - | 253.6 ± 35.8 |
| 7 | 17.5 | 290,329 | Verbascoside | 623→461→315→135 | - | - | 137.4 ± 19.9 |
| 14 | 19.4 | ND | Leucoseptoside A | 637→461→315→135 | - | - | 31.5 ± 4.6 |
| 15 | 20.0 | ND | Leonoside B | 783→607→475→329 | - | - | 25.1±4.7 |
| Flavones | |||||||
| 6 | 16.8 | 254,267,345 | Luteolin-7-O-rut | 593→285→241 | 43.3 ± 10.0 | - | - |
| 10 | 18.1 | 253,267,345 | Luteolin-7-O-glcA | 461→285→241 | D | 26.2 ± 4.0 | - |
| 12 | 18.9 | ND | Luteolin-7-O-rut | 593→285→257 | - | - | D |
| 17 | 21.1 | 266,329 | Apigenin-7-O-gl | 431→269→225 | - | D | - |
| 18 | 26.3 | 266,330 | Apigenin-7-O-(6″ acetyl)glc | 473→269, 413 | - | D | - |
| Flavonols | |||||||
| 2 | 8.2 | ND | Rutin-O-glc | 771→609→301 | - | D | - |
| 3 | 13.7 | ND | Quercetin-3-O-soph | 625→301→179 | - | - | 5.7 ± 1.1 |
| 9 | 18.0 | 256,267,355 | Rutin | 609→301→179 | - | - | 15.8 ± 2.1 |
| 11 | 18.9 | 256,267,357 | Quercetin-3-O-glc | 463→301→179 | - | - | 24.9 ± 3.8 |
| Flavanones | |||||||
| 4 | 15.0 | 283,325sh | Eriodictyol-O-rut | 595→287→151→107 | 144.6 ± 22.4 | - | - |
| 8 | 17.9 | 282,333sh | Naringenin-7-O-rut | 579→271→151 | 24.4 ± 3.7 | - | - |
| 13 | 19.5 | 283,325sh | Hesperetin-7-O-rut | 609→301→286→241 | 25.9 ± 3.6 | - | - |
| Assay | Flavan | Flav | Flavo | PEG | RAc |
|---|---|---|---|---|---|
| DPPH | 0.889 | 0.986 | −0.841 | −0.841 | 0.815 |
| RP | 0.973 | 0.917 | −0.688 | −0.688 | 0.653 |
| NO | 0.990 | 0.878 | −0.620 | −0.620 | 0.582 |
| ROS | 0.373 | 0.856 | −0.990 | −0.990 | 0.996 |
| CytP | 0.803 | 0.284 | 0.115 | 0.115 | −0.161 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, O.R.; Macias, R.I.R.; Domingues, M.R.M.; Marin, J.J.G.; Cardoso, S.M. Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L. Antioxidants 2019, 8, 267. https://doi.org/10.3390/antiox8080267
Pereira OR, Macias RIR, Domingues MRM, Marin JJG, Cardoso SM. Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L. Antioxidants. 2019; 8(8):267. https://doi.org/10.3390/antiox8080267
Chicago/Turabian StylePereira, Olívia R., Rocio I. R. Macias, Maria R. M. Domingues, Jose J. G. Marin, and Susana M. Cardoso. 2019. "Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L." Antioxidants 8, no. 8: 267. https://doi.org/10.3390/antiox8080267
APA StylePereira, O. R., Macias, R. I. R., Domingues, M. R. M., Marin, J. J. G., & Cardoso, S. M. (2019). Hepatoprotection of Mentha aquatica L., Lavandula dentata L. and Leonurus cardiaca L. Antioxidants, 8(8), 267. https://doi.org/10.3390/antiox8080267