Strawberry Intake Ameliorates Oxidative Stress and Decreases GABA Levels Induced by High-Fat Diet in Frontal Cortex of Rats


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Production of Strawberry Extract
2.2. Animal Care and Strawberry Extract Treatment
2.3. Measurement of Lipid Peroxidation and Oxidized Proteins
2.4. GABA Level Determination
2.5. Statistical Analysis
3. Results
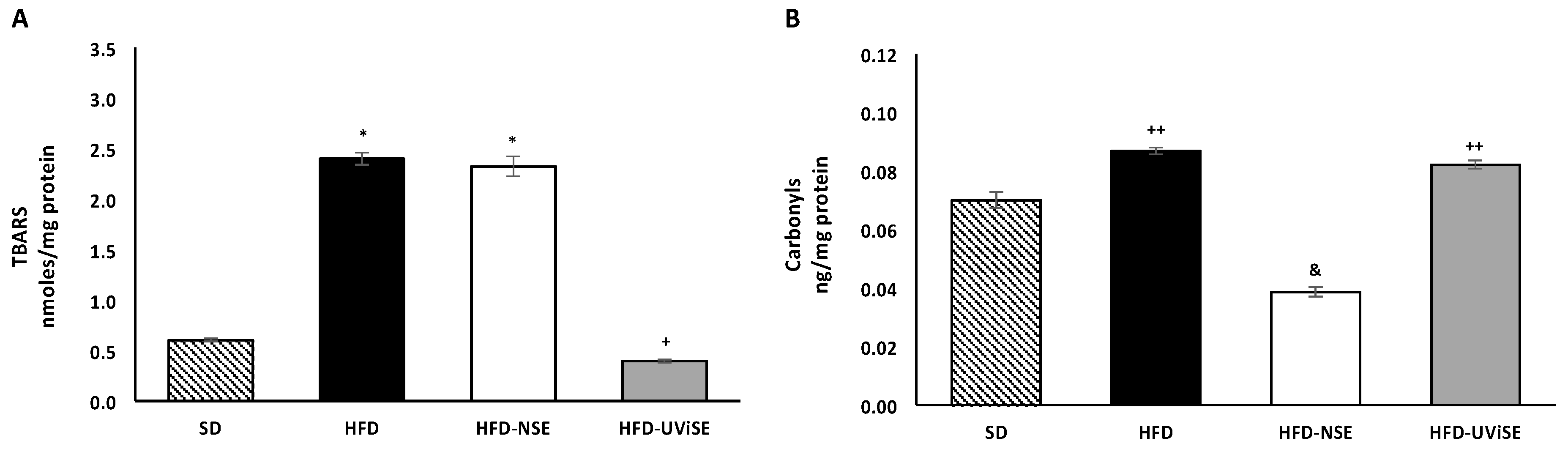
3.1. Effect of Strawberry Extracts on Oxidative Damage in the Brain
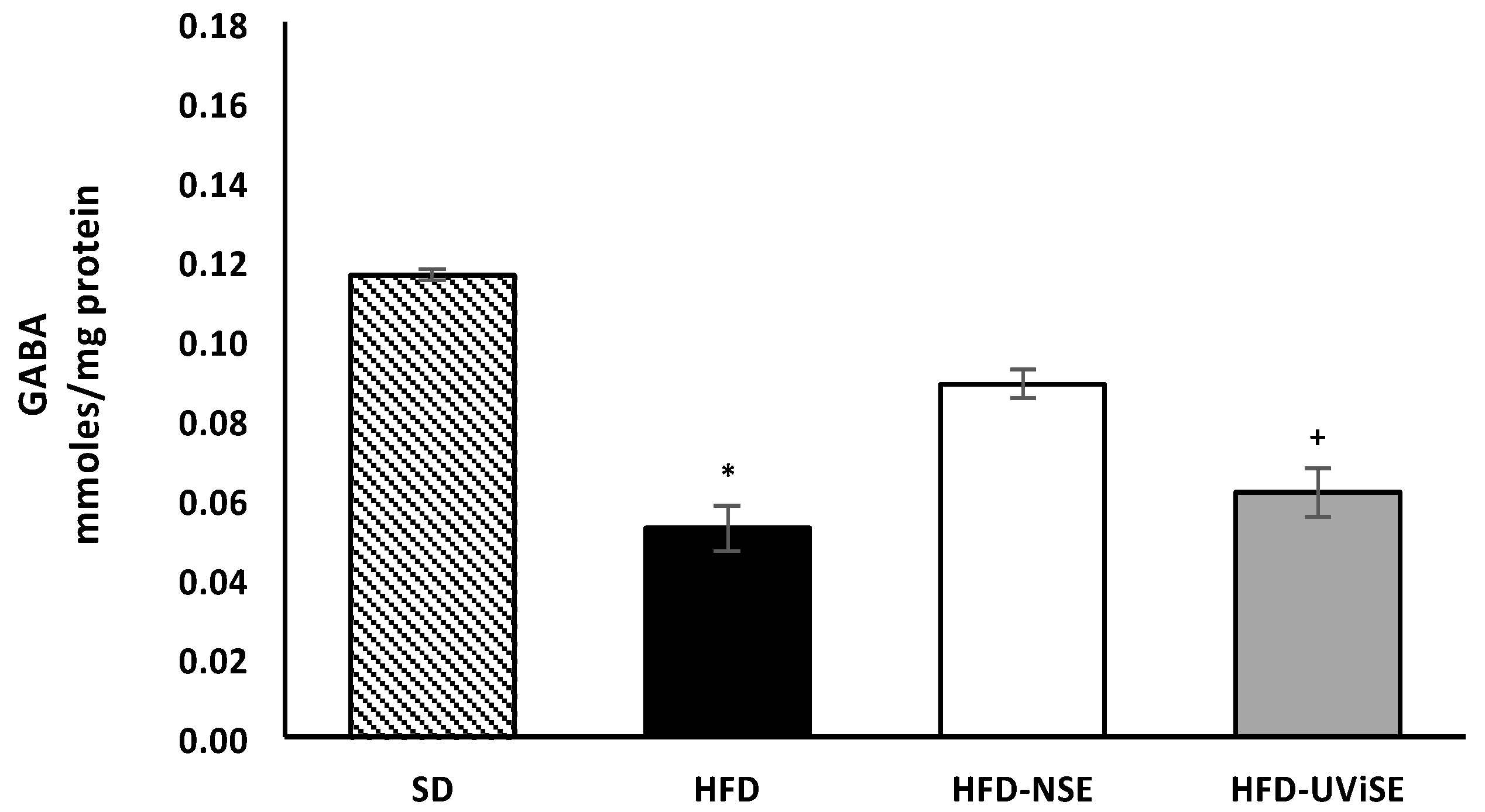
3.2. A High-Fat Diet Decreases GABA Levels in the Frontal Cortex and Hippocampus
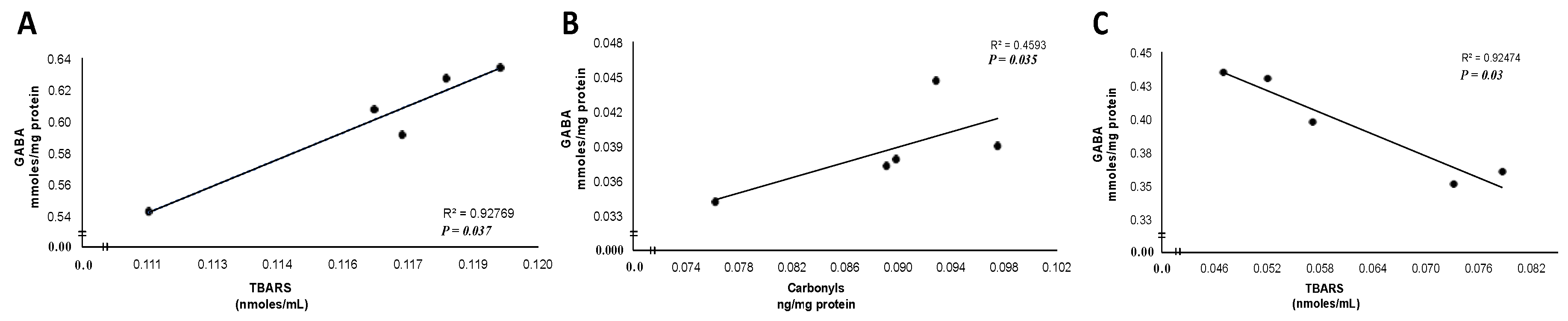
3.3. Correlation between Oxidative Damage and GABA Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Han, J.C.; Lawlor, D.A.; Kimm, S.Y. Childhood obesity. Lancet 2010, 375, 1737–1748. [Google Scholar] [CrossRef]
- Freeman, L.R.; Haley-Zitlin, V.; Rosenberger, D.S.; Granholm, A.C. Damaging effects of a high-fat diet to the brain and cognition: A review of proposed mechanisms. Nutr. Neurosci. 2014, 17, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Delgado, T.C. Glutamate and GABA in appetite regulation. Front. Endocrinol. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Salazar, C.; Ramírez-Emiliano, J.; Trejo-Bahena, T.; Oviedo-Solís, C.I.; Solís-Ortiz, M.S. A high-fat diet decreases GABA concentration in the frontal cortex and hippocampus of rats. Biol. Res. 2016, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; DeLorey, T.M. GABA and glycine. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999. [Google Scholar]
- Abreu, A.R.; De Abreu, A.R.; Santos, L.T.; de Souza, A.A.; da Silva, L.G., Jr.; Chianca, D.A., Jr.; de Menezes, R.C. Blunted GABA-mediated inhibition within the dorsomedial hypothalamus potentiates the cardiovascular response to emotional stress in rats fed a high-fat diet. Neuroscience 2014, 262, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Vong, L.; Ye, C.; Yang, Z.; Choi, B.; Chua, S., Jr.; Lowell, B.B. Leptin action on GABAergic neurons prevents obesity and reduces inhibitory tone to POMC Neurons. Neuron 2011, 71, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Ochner, C.N.; Green, D.; van Steenburgh, J.J.; Kounios, J.; Lowe, M.R. Asymemetric prefrontal cortex activation in relation to markers of overeating in obese humans. Appetite 2009, 53, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Lenard, N.R.; Berthoud, H.R. Central and peripheral regulation of food intake and physical activity: Pathways and genes. Obesity 2008, 16, S11–S22. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.W.; Zhang, J.; Townsend, D.M.; Tew, K.D. Oxidative stress, redox regulation and diseases of cellular differentiation. Biochim. Biophys. Acta 2015, 1850, 1607–1621. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Tulipani, S.; Gonzàles-Paramàs, A.M.; Santos-Buelga, C.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Photoprotective potential of strawberry (Fragaria × ananassa) extract against UV-A irradiation damage on human fibroblasts. J. Agric. Food Chem. 2012, 60, 2323–2327. [Google Scholar] [CrossRef]
- Kårlund, A.; Salminen, J.P.; Koskinen, P.; Ahern, J.R.; Karonen, M.; Tiilikkala, K.; Karjalainen, R.O. Polyphenols in strawberry (Fragaria × ananassa) leaves induced by plant activators. J. Agric. Food Chem. 2014, 62, 4592–4600. [Google Scholar] [CrossRef]
- Martínez-Morúa, A.; Soto-Urquieta, M.G.; Franco-Robles, E.; Zúñiga-Trujillo, I.; Campos-Cervantes, A.; Pérez-Vázquez, V.; Ramírez-Emiliano, J. Curcumin decreases oxidative stress in mitochondria isolated from liver and kidneys of high-fat diet-induced obese mice. J. Asian Nat. Prod. Res. 2013, 15, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Flores, L.M.; López-Briones, S.; Macías-Cervantes, M.H.; Ramírez-Emiliano, J.; Pérez-Vázquez, V. A PPARγ, NF-κB and AMPK-dependent mechanism may be involved in the beneficial effects of curcumin in the diabetic db/db mice liver. Molecules 2004, 19, 8289–8302. [Google Scholar] [CrossRef]
- Franco-Robles, E.; Campos-Cervantes, A.; Murillo-Ortiz, B.O.; Segovia, J.; López-Briones, S.; Vergara, P.; Pérez-Vázquez, V.; Solís-Ortiz, M.S.; Ramírez-Emiliano, J. Effects of curcumin on brain-derived neurotrophic factor levels and oxidative damage in obesity and diabetes. Appl. Physiol. Nutr. Metab. 2014, 39, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Lau, F.C.; Carey, A.N.; Galli, R.L.; Spangler, E.L.; Ingram, D.K.; Joseph, J.A. Blueberry polyphenols attenuate kainic acid- induced decrements in cognition and alter inflammatory gene expression in rat hippocampus. Nutr. Neurosci. 2008, 11, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Gu, L.; Hager, T.J.; Hager, A.; Howard, L.R. Whole berries versus berry anthocyanins: Interactions with dietary fat levels in the C57BL/6J mouse model of obesity. J. Agric. Food Chem. 2008, 56, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Bielinski, D.F.; Carrihill-Knoll, K.L.; Rabin, B.M.; Shukitt-Hale, B. Protective effects of blueberry- and strawberry diets on neuronal stress following exposure to 56Fe particles. Brain Res. 2014, 1593, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Jiao, H. Scavenging capacity of berry crops on superoxide radicals, hydrogen peroxide, hydroxyl radicals, and singlet oxygen. J. Agric. Food Chem. 2000, 48, 5677–5684. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Yu, Y.M.; Chiang, S.Y.; Tseng, C.Y. Ellagic acid suppresses oxidised low-density lipoprotein-induced aortic smooth muscle cell proliferation: Studies on the activation of extracellular signal-regulated kinase 1/2 and proliferating cell nuclear antigen expression. Br. J. Nutr. 2008, 99, 709–714. [Google Scholar] [CrossRef]
- Basu, A.; Wilkinson, M.; Penugonda, K.; Simmons, B.; Betts, N.M.; Lyons, T.J. Freeze-dried strawberry powder improves lipid profile and lipid peroxidation in women with metabolic syndrome: Baseline and post intervention effects. Nutr. J. 2009, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Alothman, R.; Bhat, R.; Karim, A.A. UV radiation-induced changes of antioxidant capacity of fresh-cut tropical fruits. Innov. Food Sci. Emerg. Technol. 2009, 10, 512–516. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Wang, C.Y.; Buta, J.G.; Krizek, D.T. Use of UV-C irradiation to prevent decay and maintain postharvest quality of ripe “Tommy Atkins” mangoes. Int. J. Food Sci. Technol. 2008, 36, 767–773. [Google Scholar] [CrossRef]
- Ayala-Gil, M.E.; Lozoya-Gloria, E. Methods for Increasing the Nutraceutical Content of Perishable Fruits. U.S. Patent US2016/0235080A1. Available online: https://www.google.com/patents/US20160235080 (accessed on 18 August 2016).
- Oviedo-Solís, C.I.; Sandoval-Salazar, C.; Lozoya-Gloria, E.; Maldonado-Aguilera, G.A.; Aguilar-Zavala, H.; Beltrán-Campos, V.; Pérez-Vázquez, V.; Ramírez-Emiliano, J. Ultraviolet light-C increases antioxidant capacity of the strawberry (Fragaria × ananassa) in vitro and in high-fat diet-induced obese rats. Food Sci. Nutr. 2017, 5, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Institute of Laboratory Animal Resources. Guide for the Care and Use of Laboratory Animals; National Institutes of Health Publication No. 85–23 (revised 1996); National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- Norma Oficial Mexicana (NOM-062-ZOO-1999). In Especificaciones Técnicas Para la Producción, Cuidado y uso de los Animales de Laboratorio; Ochoa, M.L.I., Ed.; Diario Oficial de la Federación: México city, México, 2001. [Google Scholar]
- Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Strawberry as a health promoter: An evidence based review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and human health: Effects beyond antioxidant activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Solís, C.I.; Cornejo-Manzo, S.; Murillo-Ortiz, B.O.; Guzmán-Barrón, M.M.; Ramírez-Emiliano, J. Strawberry polyphenols decrease oxidative stress in chronic diseases. Gaceta Medica de Mexico 2018, 154, 60–65. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.; Mazzoni, L.; Forbes-Hernandez, T.; Gasparrini, M.; Gonzàlez-Paramàs, A.; Santos-Buelga, C.; Quiles, J.; Bompadre, S.; Mezzetti, B.; et al. Polyphenol-rich strawberry extract protects human dermal fibroblasts against hydrogen peroxide oxidative damage and improves mitochondrial functionality. Molecules 2004, 19, 7798–7816. [Google Scholar] [CrossRef]
- Forbes-Hernández, T.Y.; Afrin, S.; Cianciosi, D.; Manna, P.P.; Zhang, J.; Gasparrini, M.; Reboredo-Rodriguez, P. Strawberry extract attenuates oxidative stress in 3T3-L1 cells. J. Berry Res. 2018, 8, 193–203. [Google Scholar] [CrossRef]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of malondialdehyde (MDA) by high-performance liquid chromatography in serum and liver as a biomarker for oxidative stress. Application to a rat model for hypercholesterolemia and evaluation of the effect of diets rich in phenolic antioxidant. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 827, 76–82. [Google Scholar] [CrossRef]
- Forbes-Hernández, T.; Giampieri, F.; Gasparrini, M.; Afrin, S.; Mazzoni, L.; Cordero, M.; Mezzetti, B.; Quiles, J.; Battino, M. Lipid Accumulation in HepG2 Cells Is Attenuated by Strawberry Extract through AMPK Activation. Nutrients 2017, 9, 621. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Dekanski, D.; Ristić, S. Strawberry polyphenols attenuate ethanol-induced gastric lesions in rats by activation of antioxidant enzymes and attenuation of MDA increase. PLoS ONE 2011, 6, e25878. [Google Scholar] [CrossRef]
- Morrison, C.D.; Pistell, P.J.; Ingram, D.K.; Johnson, W.D.; Liu, Y.; Fernandez-Kim, S.O.; White, C.L.; Purpera, M.N.; Uranga, R.M.; Bruce-Keller, A.J.; et al. High fat diet increases hippocampal oxidative stress and cognitive impairment in aged mice: Implications for decreased Nrf2 signaling. J. Neurochem. 2010, 114, 1581–1589. [Google Scholar] [CrossRef]
- Cobb, C.A.; Cole, M.P. Oxidative and nitrative stress in neurodegeneration. Neurobiol. Dis. 2015, 84, 4–21. [Google Scholar] [CrossRef]
- Valladolid-Acebes, I.; Merino, B.; Principato, A.; Fole, A.; Barbas, C.; Lorenzo, M.P.; García, A.; Del Olmo, N.; Ruiz-Gayo, M.; Cano, V. High-fat diets induce changes in hippocampal glutamate metabolism and neurotransmission. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E396–E402. [Google Scholar] [CrossRef] [PubMed]
- Sickmann, H.M.; Waagepetersen, H.S.; Schousboe, A.; Benie, A.J.; Bouman, S.D. Obesity and type 2 diabetes in rats are associated with altered brain glycogen and amino-acid homeostasis. J. Cereb. Blood Flow Metab. 2010, 30, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Dey, A.; Yu, X.; Stranahan, A.M. Dietary obesity reversibly induces synaptic stripping by microglia and impairs hippocampal plasticity. Brain Behav. Immun. 2016, 51, 230–239. [Google Scholar] [CrossRef]
- Baldo, B.A.; Spencer, R.C.; Sadeghian, K.; Mena, J.D. GABA-mediated inactivation of medial prefrontal and agranular insular cortex in the rat: Contrasting effects on hunger- and palatability-driven feeding. Neuropsychopharmacology 2016, 41, 960–970. [Google Scholar] [CrossRef]
- Treviño, S.; Aguilar-Alonso, P.; Flores Hernandez, J.A.; Brambila, E.; Guevara, J.; Flores, G.; Lopez-Lopez, G.; Muñoz-Arenas, G.; Morales-Medina, J.C.; Toxqui, V.; et al. A high calorie diet causes memory loss, metabolic syndrome and oxidative stress into hippocampus and temporal cortex of rats. Synapse 2015, 69, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Koopman, K.E.; Booij, J.; Fliers, E.; Serlie, M.J.; la Fleur, S.E. Diet-induced changes in the lean brain: Hypercaloric high-fat-high-sugar snacking decreases serotonin transporters in the human hypothalamic region. Mol. Metab. 2013, 2, 417–422. [Google Scholar] [CrossRef]
- Abram, V.; Berlec, B.; Ota, A.; Šentjurc, M.; Blatnik, P.; Ulrih, N.P. Effect of flavonoid structure on the fluidity of model lipid membranes. Food Chem. 2013, 139, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Goyarzu, P.; Malin, D.H.; Lau, F.C.; Taglialatela, G.; Moon, W.D.; Jennings, R.; Moy, E.; Moy, D.; Lippold, S.; Shukitt-Hale, B.; et al. Blueberry supplemented diet: Effects on object recognition memory and nuclear factor-kappa B levels in aged rats. Nutr. Neurosci. 2004, 7, 75–83. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandoval-Salazar, C.; Oviedo-Solís, C.I.; Lozoya-Gloria, E.; Aguilar-Zavala, H.; Solís-Ortiz, M.S.; Pérez-Vázquez, V.; Balcón-Pacheco, C.D.; Ramírez-Emiliano, J. Strawberry Intake Ameliorates Oxidative Stress and Decreases GABA Levels Induced by High-Fat Diet in Frontal Cortex of Rats. Antioxidants 2019, 8, 70. https://doi.org/10.3390/antiox8030070
Sandoval-Salazar C, Oviedo-Solís CI, Lozoya-Gloria E, Aguilar-Zavala H, Solís-Ortiz MS, Pérez-Vázquez V, Balcón-Pacheco CD, Ramírez-Emiliano J. Strawberry Intake Ameliorates Oxidative Stress and Decreases GABA Levels Induced by High-Fat Diet in Frontal Cortex of Rats. Antioxidants. 2019; 8(3):70. https://doi.org/10.3390/antiox8030070
Chicago/Turabian StyleSandoval-Salazar, Cuauhtémoc, Cecilia I. Oviedo-Solís, Edmundo Lozoya-Gloria, Herlinda Aguilar-Zavala, Martha S. Solís-Ortiz, Victoriano Pérez-Vázquez, Cristina D. Balcón-Pacheco, and Joel Ramírez-Emiliano. 2019. "Strawberry Intake Ameliorates Oxidative Stress and Decreases GABA Levels Induced by High-Fat Diet in Frontal Cortex of Rats" Antioxidants 8, no. 3: 70. https://doi.org/10.3390/antiox8030070
APA StyleSandoval-Salazar, C., Oviedo-Solís, C. I., Lozoya-Gloria, E., Aguilar-Zavala, H., Solís-Ortiz, M. S., Pérez-Vázquez, V., Balcón-Pacheco, C. D., & Ramírez-Emiliano, J. (2019). Strawberry Intake Ameliorates Oxidative Stress and Decreases GABA Levels Induced by High-Fat Diet in Frontal Cortex of Rats. Antioxidants, 8(3), 70. https://doi.org/10.3390/antiox8030070