Exogenous Plant-Based Nutraceutical Supplementation and Peripheral Cell Mononuclear DNA Damage Following High Intensity Exercise



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Exercise Habituation
2.3. Supplementation Protocol and Nutraceutical Preparation
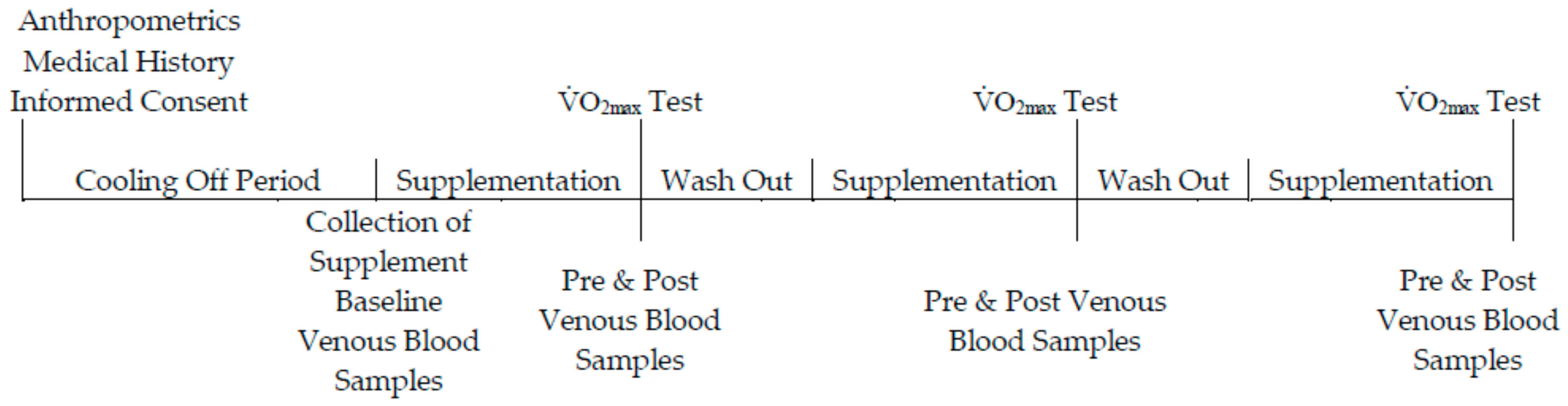
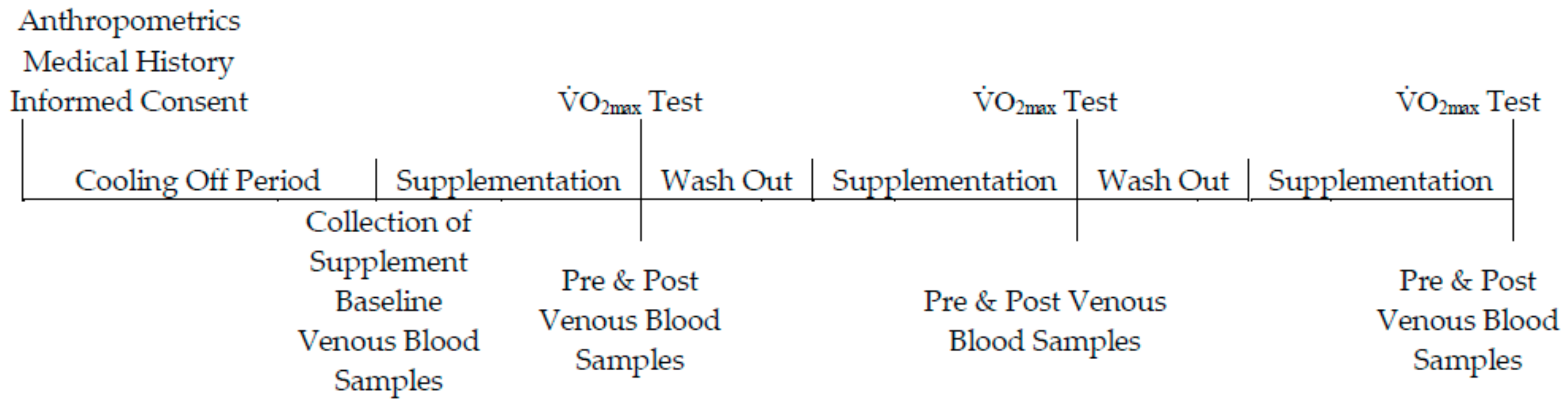
2.4. Experimental Testing
2.5. Biochemical Indices
2.6. Deoxyribonucleic Acid (DNA)
2.7. Lipid Hydroperoxides (LOOH)
2.8. Lipid Soluble Antioxidants (LSA)
2.9. Electron Paramagnetic Resonance (EPR) Spectroscopy
2.10. Superoxide Dismutase (SOD)
2.11. Statistical Analysis
3. Results
3.1. Compliance
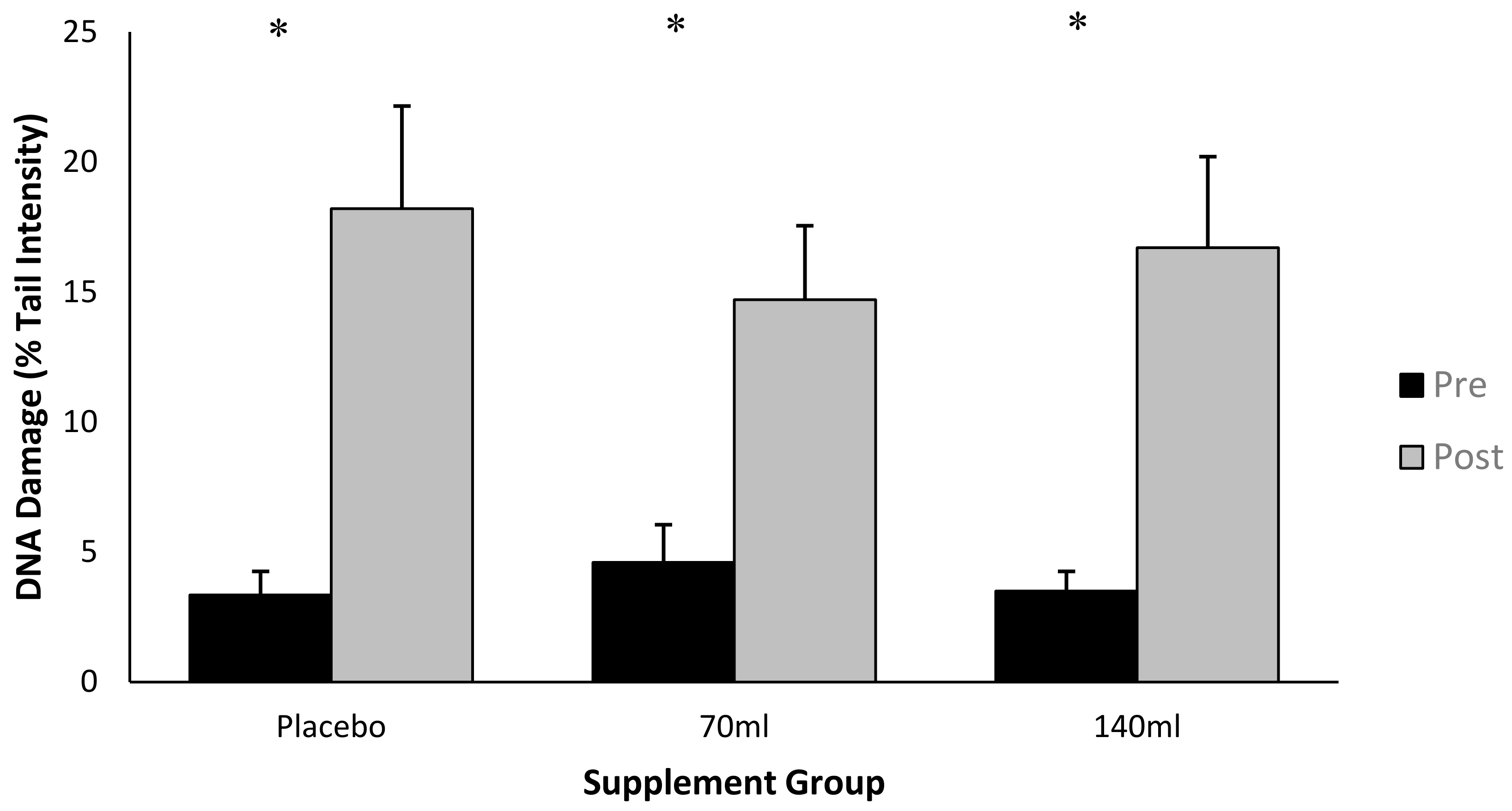
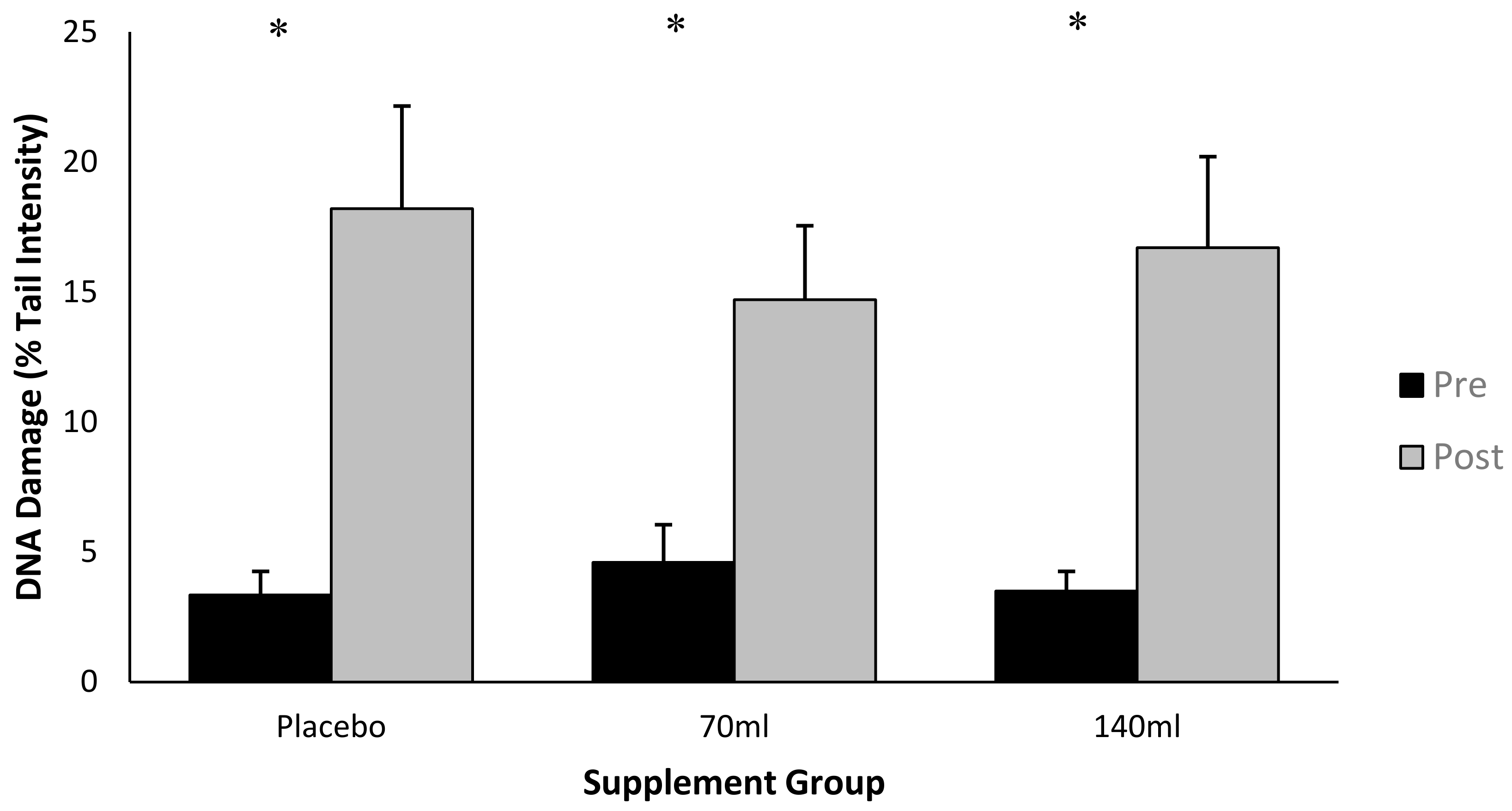
3.2. DNA Damage
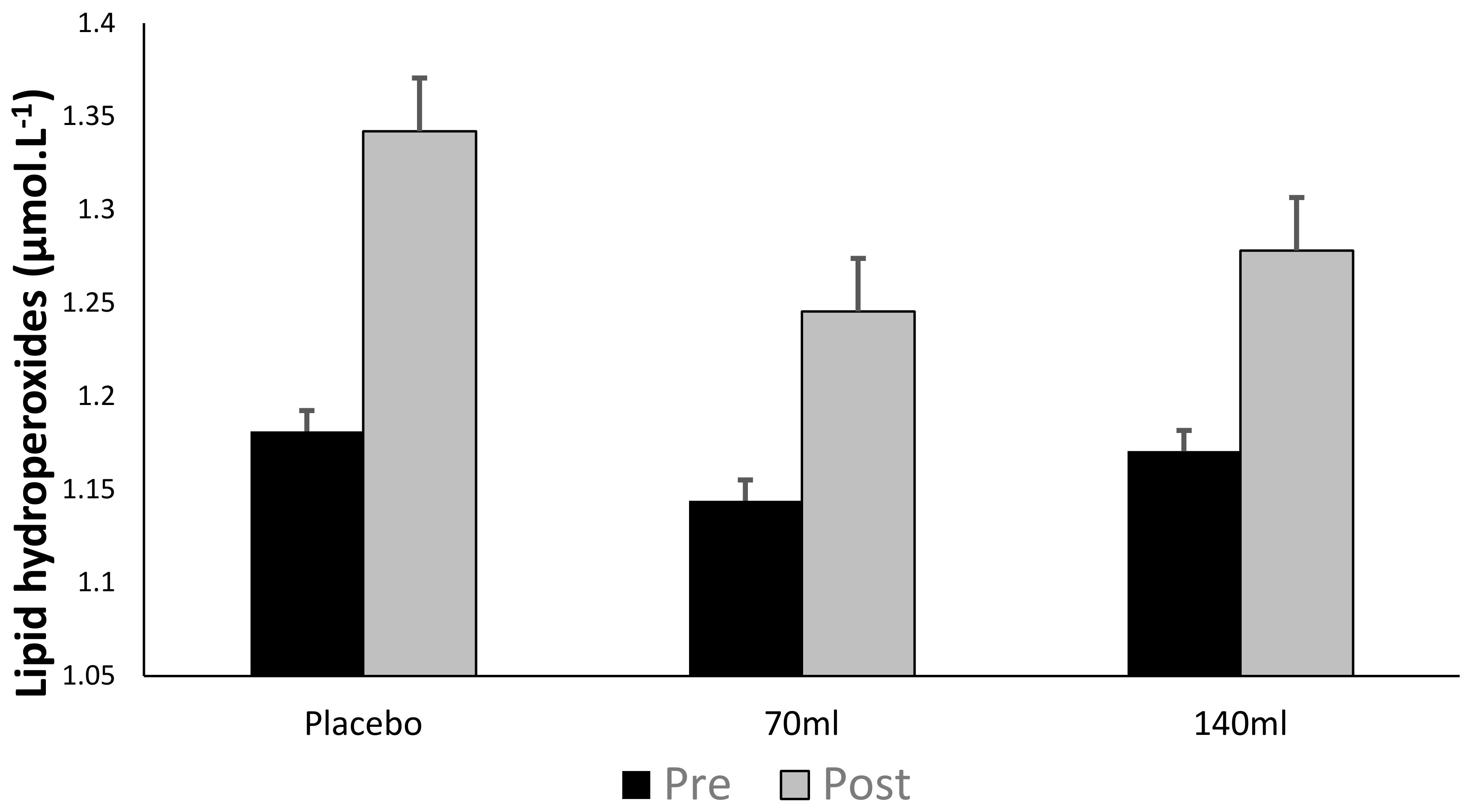
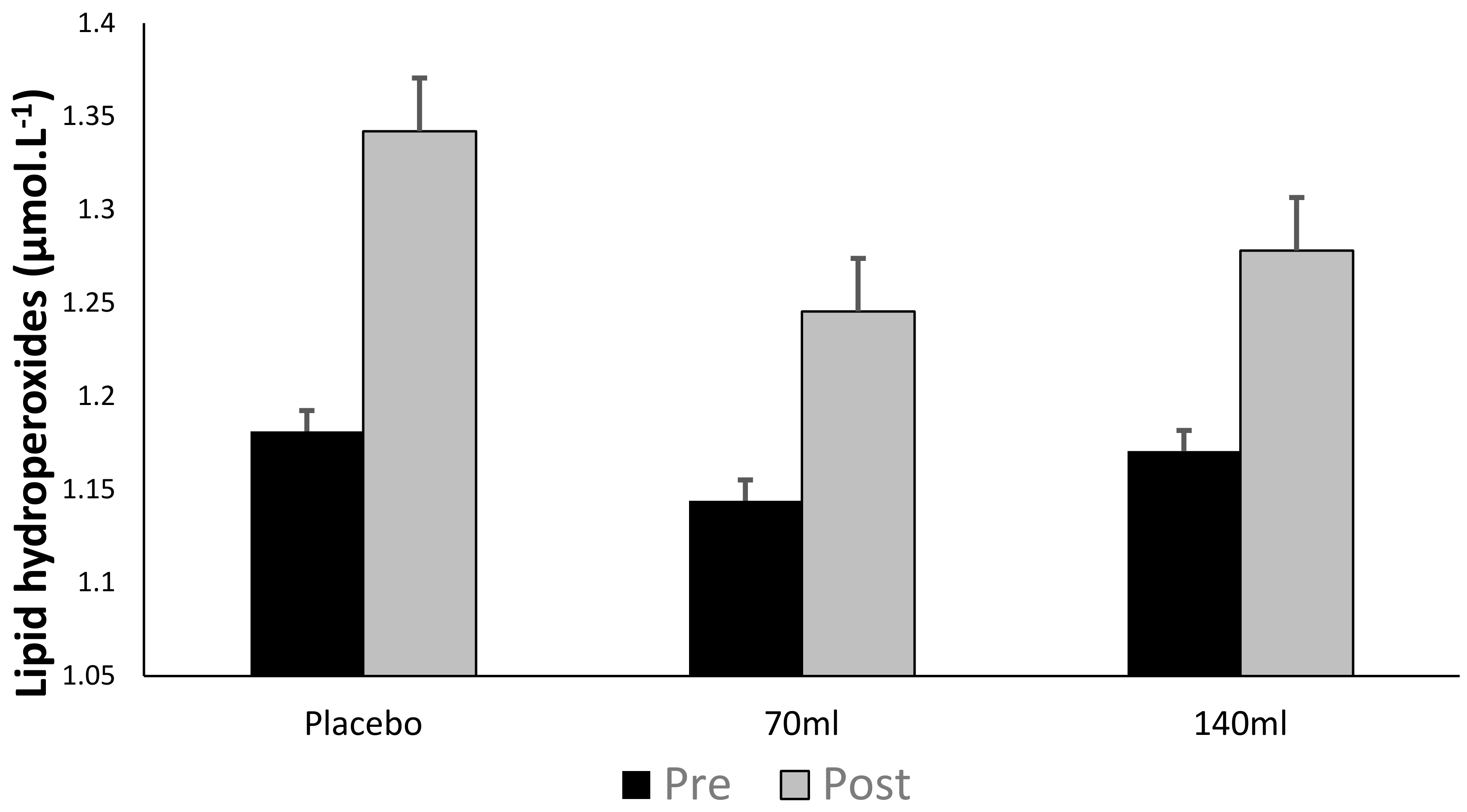
3.3. Lipid Hydroperoxide
3.4. Lipid Soluble Antioxidants
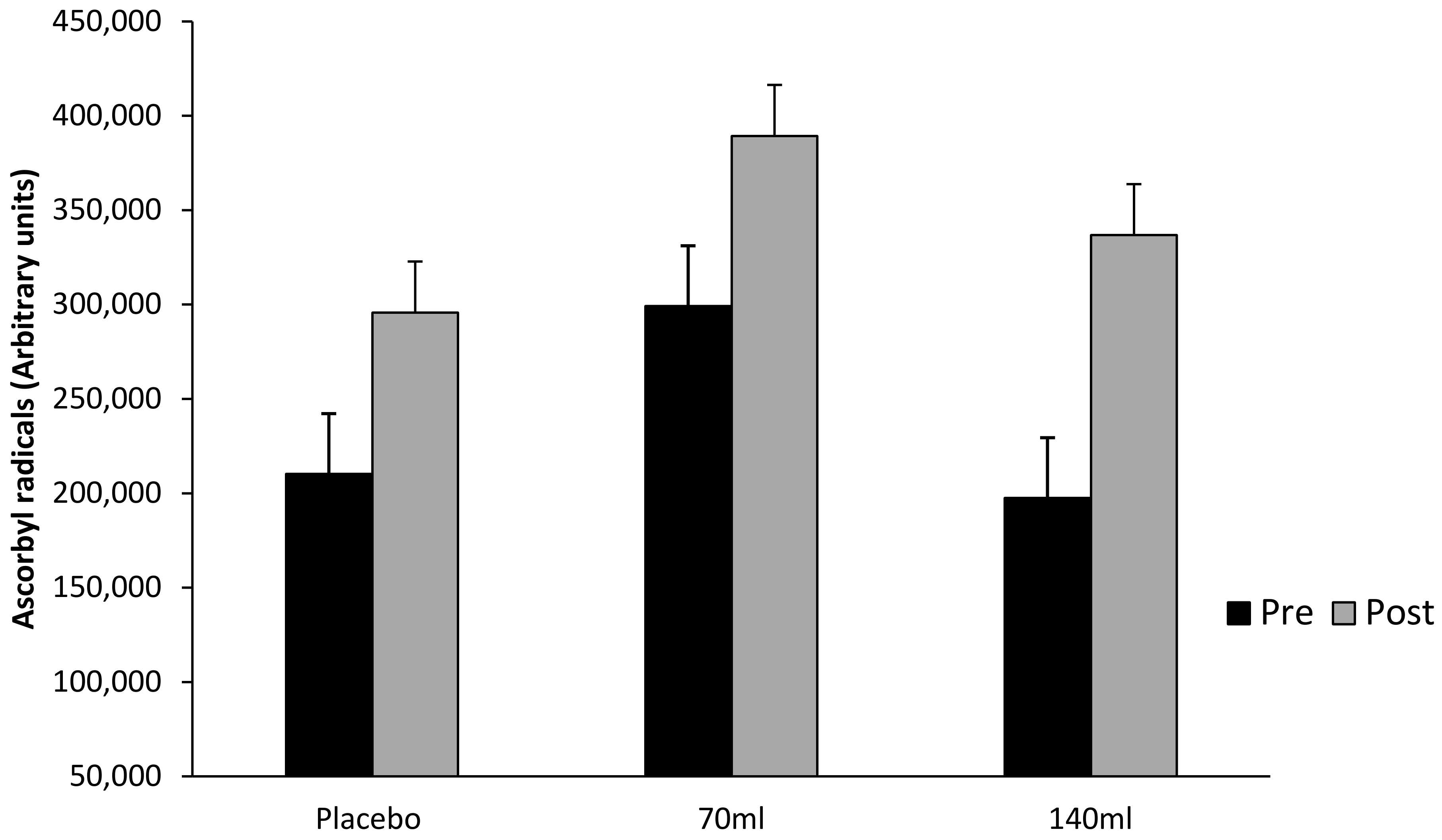
3.5. Ascorbyl Free Radicals
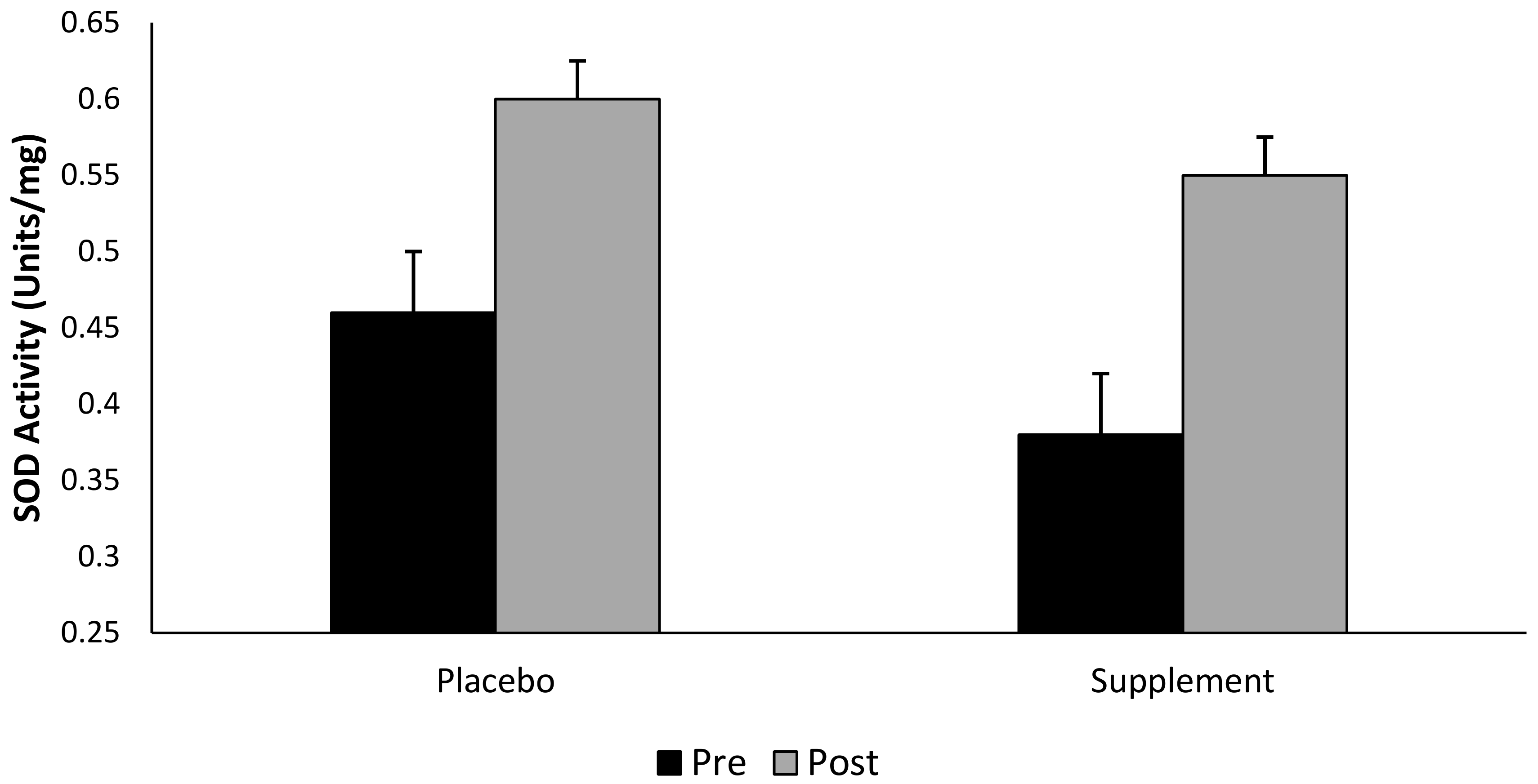
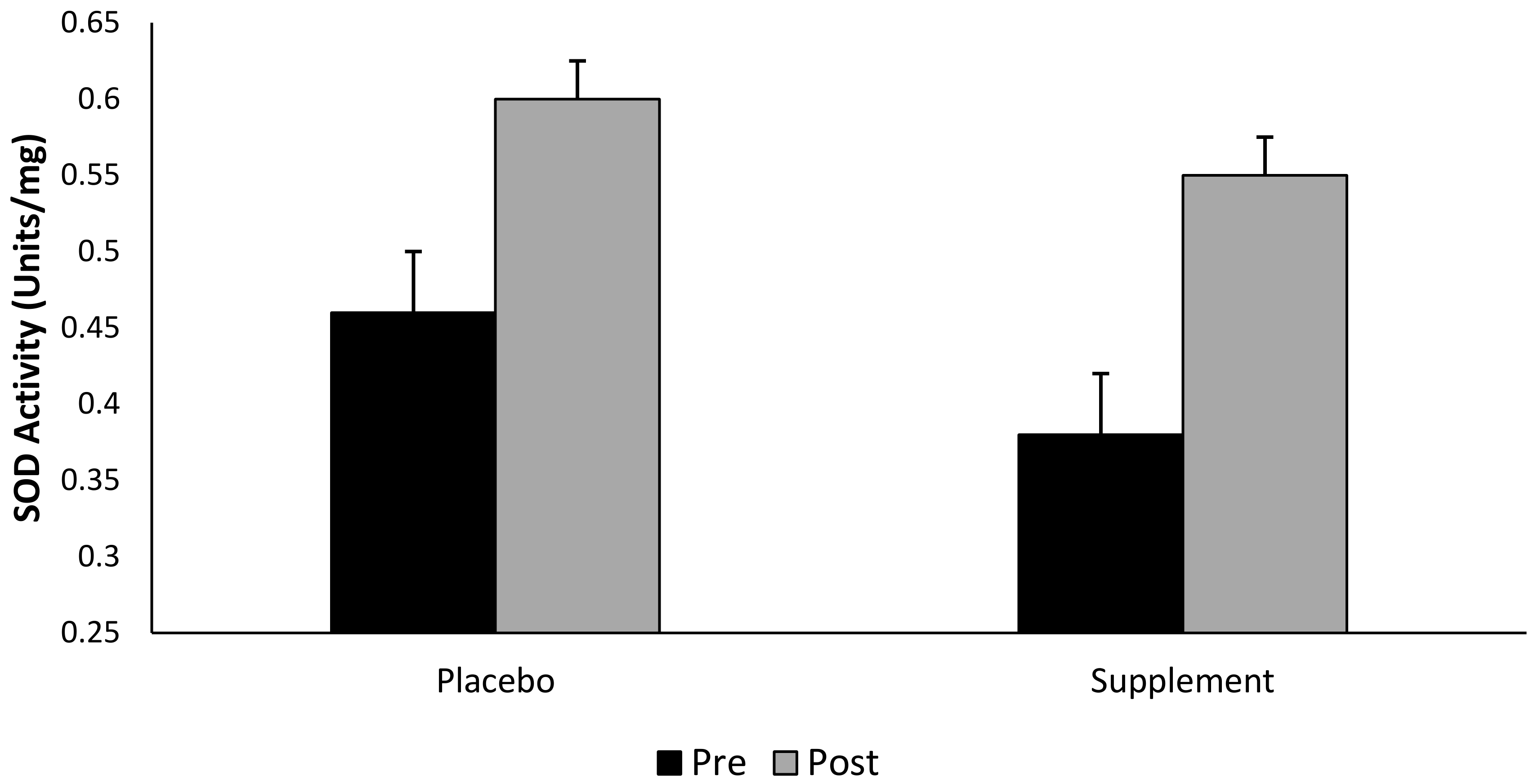
3.6. Superoxide Dismutase
4. Discussion
4.1. DNA Damage
4.2. Lipid Peroxidation
4.3. Prophylactic Effect of Plant-Derived Nutraceuticals
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Elokda, A.S.; Nielsen, D.H. Effects of exercise training on the glutathione antioxidant system. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Marton, O.; Nagy, E.; Koltai, E.; Goto, S. The complex role of physical exercise and reactive oxygen species on brain. J. Sport Health Sci. 2013, 2, 87–93. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Sweazea, K.L.; Johnston, C.S.; Knurick, J.; Bliss, C.D. Plant-based nutraceutical increases plasma catalase activity in healthy participants: A small double-blind, randomised, placebo-controlled, proof of concept trial. J. Diet. Suppl. 2017, 14, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Exercise, oxidative stress, and antioxidants. Am. J. Sports Med. 1996, 24, S20–S24. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J. Handbook of Oxidants and Antioxidants in Exercise. Hanninen, O., Packer, L., Sen, C.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 57–68.
- Davison, G.W.; Hughes, C.M.; Bell, R.A. Exercise and Mononuclear Cell DNA Damage: The Effects of Antioxidant Supplementation. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.; Lima, L.C.F. The Association between Physical Exercise and Reactive Oxygen Species (ROS) Production. J. Sports Med. Doping Stud. 2014, 5, 152. [Google Scholar]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defence. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Bohn, T. Exogenous Antioxidants—Double-Edged Swords in Cellular Redox State. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Sasazuki, S.; Hayashi, T.; Nakachi, K.; Tsubono, Y.; Okubo, S.; Hayashi, M.; Tsugane, S. Protective Effect of Vitamin C on Oxidative Stress: A Randomized Controlled Trial. Int. J. Vitam. Nutr. Res. 2008, 78, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Role of Vitamin E as a Lipid-Soluble Peroxyl Radical Scavenger: In Vitro and In Vivo. Free Radic. Biol. Med. 2014, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Keong, C.C.; Singh, H.J.; Singh, R. Effects of Palm Vitamin E Supplementation on Exercise-Induced Oxidative Stress and Endurance Performance in the Heat. J. Sports Sci. Med. 2006, 5, 629–639. [Google Scholar] [PubMed]
- Taghiyar, M.; Darvishi, L.; Askari, G.; Feizi, A.; Hariri, M.; Mashhadi, N.S.; Ghiasvand, R. The Effects of Vitamin C and E Supplementation on Muscle Damage and Oxidative Stress in Female Athletes: A Clinical Trial. Int. J. Prev Med. 2013, 4, S16–S23. [Google Scholar] [PubMed]
- Morillas-Ruiz, J.M.; Villegas Garcia, J.A.; Lopez, F.J.; Vidal-Guevara, M.L.; Zafrilla, P. Effects of Polyphenolic Antioxidants on Exercise-Induced Oxidative Stress. Clin. Nutr. 2006, 25, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jew, S.S.; Chang, P.S.; Hong, I.J.; Hwang, E.S.; Kim, K.S.; Kim, K.T. Free radical scavenging effect and antioxidant activities of barley leaves. Food Sci. Biotechnol. 2003, 12, 268–273. [Google Scholar]
- Paulíčková, I.; Ehrenbergerová, J.; Fiedlerová, V.; Gabrovska, D.; Havlova, P.; Holasova, M.; Kopáček, J.; Ouhrabková, J.; Pinkrová, J.; Rysová, J.; et al. Evaluation of barley grass as a potential source of some nutritional substances. Czech J. Food Sci. 2007, 25, 65–72. [Google Scholar] [CrossRef]
- Ghavami, L.; Goliaei, B.; Taghizadeh, B.; Nikoofar, A. Effects of barley β-glucan on radiation damage in the human hepatoma cell line HepG2. Mutat. Res. 2014, 775–776, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kulkami, S.D.; Tilak, J.C.; Acharya, R.; Rajurkar, N.S.; Devasagayam, T.P.; Reddy, A.V. Evaluation of the antioxidant activity of wheatgrass (Triticum aestivum, L.) as a function of growth under different conditions. Phytother. Res. 2006, 20, 218–227. [Google Scholar]
- Sethi, J.; Yadav, M.; Dahiya, K.; Sood, S.; Singh, V.; Bhattacharya, S.B. Antioxidant effect of Triticum aestivium (wheat grass) in high-fat diet-induced oxidative stress in rabbits. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Simundic, A.; Cornes, M.; Grankvist, K.; Lippi, G.; Nybo, M. Standardization of collection requirements for fasting samples: For the Working Group on Preanalytical Phase (WG-PA) of the European Federation of Clinical Chemistry and Laboratory Medicine (EFLM). Clin. Chim. Acta. 2013, 432, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Boorsma, R.K.; Whitfeld, J.; Spriett, L.L. Beetroot juice supplementation does not improve performance of elite 1500-m runners. Med. Sci. Sport Exerc. 2014, 46, 2326–2334. [Google Scholar] [CrossRef] [PubMed]
- Dill, D.B.; Costill, D.L. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Dacie, J.V.; Lewis, S.M. Practical Haematology; Churchill: London, UK, 1968. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Wolff, S.P. Ferrous Ion Oxidation in Presence of Ferric Ion Indicator Xylenol Orange for Measurement of Hydroperoxides. Methods Enzymol. 1994, 223, 182–189. [Google Scholar]
- Thurnham, D.I.; Smith, E.; Flora, P.S. Concurrent Lipid-Chromatpgraphic Assay of Retinol, α-Tocopherol, β-Carotene, α-Carotene, Lycopene, and β-Cryptoxanthin in Plasma, with Tocopherol Acetate as an Internal Standard. Clin. Chem. 1988, 34, 377–381. [Google Scholar] [PubMed]
- Clliford, T.; Berntzen, B.; Davison, G.W.; West, D.J.; Howatson, G.; Steevenson, E.J. Effects of Beetroot Juice on Recovery of Muscle Function and Performance between Bouts of Repeated Sprint Exercise. Nutrients 2016, 8, E506. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, M.C.; Hughes, C.M.; Burke, G.; Brown, J.C.; Trinick, T.R.; Duly, E.; Bailey, D.M.; Davison, G.W. Exercise-induced lipid peroxidation: Implications for deoxyribonucleic acid damage and systemic free radical generation. Environ. Mol. Mutagen. 2011, 52, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Lambertucci, R.H.; Levada-Pires, A.C.; Rossoni, L.V.; Curi, R.; Pithon-Curi, T.C. Effects of aerobic exercise training on antioxidant enzyme activities and mRNA levels in soleus muscle from young and aged rats. Mech. Ageing Dev. 2007, 128, 267–75. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R.; Oscoz, A.A.; Brunborg, G.; Gaivão, I.; Giovannelli, L.; Kruszewski, M.; Smith, C.C.; Stetina, R. The comet assay: Topical issues. Mutagenesis 2008, 23, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.W. Exercise and Oxidative Damage in Nucleoid DNA Quantified Using Single Cell Gel Electrophoresis: Present and Future Application. Front. Physiol. 2016, 7, 249. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J.; Vasilaki, A.; McArdle, A. Cellular mechanisms underlying oxidative stress in human exercise. Free Radic. Biol. Med. 2016, 98, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Urso, M.L.; Clarkson, P.M. Oxidative stress, exercise, and antioxidant supplementation. Toxicology 2003, 189, 41–54. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Kushnareva, Y.E.; Murphy, A.N.; Starkov, A.A. Mitochondrial ROS metabolism: 10 years later. Biochemistry 2015, 80, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002, 227, 44784–44790. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, R.L.S.; Quinlan, C.L.; Perevoshchikova, I.V.; Hey-Mogensen, M.; Brand, M.D. Sites of superoxide and hydrogen peroxide production by muscle mitochondria assessed ex vivo under conditions mimicking rest and exercise. J. Biol. Chem. 2015, 290, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.; Klimcakova, E.; St-Pierre, J. Impact of PGC-1alpha on the topology and rate of superoxide production by the mitochondrial electron transport chain. Free Radic. Biol. Med. 2011, 51, 2243–2248. [Google Scholar] [CrossRef] [PubMed]
- Saint-Pierre, J.; Drori, S.; Uldry, M.; Silvaggi, J.M.; Rhee, J.; Jäger, S.; Handschin, C.; Zheng, K.; Lin, J.; Yang, W.; et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional cofactors. Cell 2006, 127, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.; Hsu, T.G.; Hsu, K.M.; Cheng, H.; Liu, T.Y.; Hsu, C.F.; Kong, C.W. Oxidative DNA damage in human peripheral leukocytes induced by massive aerobic exercise. Free Radic. Biol. Med. 2001, 31, 1465–1472. [Google Scholar] [CrossRef]
- Guetens, G.; De Boeck, G.; Highley, M. Oxidative DNA damage: Biological significance and methods of analysis. Crit. Rev. Clin. Lab. Sci. 2002, 39, 331–457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Izumi, I.; Kagamimori, S.; Sokejima, S.; Yamagami, T.; Liu, Z.; Qi, B. Role of taurine supplementation to prevent exercise-induced oxidative stress in healthy young men. Amino Acids 2004, 26, 203–207. [Google Scholar] [PubMed]
- Bo, H.; Jiany, N.; Ji, L.L.; Zhang, Y. Mitochondrial redox metabolism in aging: Effects of exercise interventions. J. Sport Health Sci. 2013, 2, 67–74. [Google Scholar] [CrossRef]
- Langmaier, J.; Samec, Z.; Samcova, E.; Hobza, P.; Reha, D. Origin of difference between one-electron redox potentials of guanosine and guanine: Electrochemical and quantum chemical study. J. Phys. Chem. 2004, 108, 15896–15899. [Google Scholar] [CrossRef]
- Roszkowski, K.; Jozwicki, W.; Blaszcyk, P.; Mucha-Malecka, A.; Siomek, A. Oxidative damage DNA: 8-oxoGua and 8-oxodG as molecular markers of cancer. Med. Sci. Monit. 2011, 17, CR329–CR333. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Barnes, D.E. Repair of endogenous DNA damage. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhu, B.; Hao, W.; Zeng, X.; Vlahopoulos, S.A.; Hazra, T.K.; Hegde, M.L.; Radak, Z.; Bacsi, A.; Brasier, A.R.; et al. Oxidized guanine base lesions function in 8-oxoguanine DNA gylcosylase1-mediated epigenetic regulation of nuclear factor kappaB-driven gene expression. J. Biol. Chem. 2016, 291, 25553–25566. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Hagiwara, Y.; Hagiwara, H.; Shibamoto, T. Inhibitory effect of 2′′-O-glycosyl isovitexin and α-tocopherol on genotoxic glyoxal formation in a lipid peroxidation system. Food Chem. Toxicol. 1994, 32, 1047–1051. [Google Scholar] [CrossRef]
- McClean, C.; Harris, R.A.; Brown, M.; Brown, J.C.; Davison, G.W. Effects of Exercise Intensity on Post-Exercise Endothelial Function and Oxidative Stress. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Marnett, L.J. Oxy radicals, lipid peroxidation and DNA damage. Toxicology 2002, 181–182, 219–222. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Free Radicals in Biology and Medicine; Oxford Press: Oxford, UK, 2007. [Google Scholar]
- Davison, G.W.; Ashton, T.; George, L.; Young, I.S.; McEneny, J.; Davies, B.; Jackson, S.K.; Peters, J.R.; Bailey, D.M. Molecular detection of exercise-induced free radicals following ascorbate prophylaxis in type 1 diabetes mellitus: A randomised controlled trial. Diabetologia 2008, 51, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, C.W. Superoxide dismutase in barley. J. Inst. Brew. 1983, 89, 420–423. [Google Scholar] [CrossRef]
- Markham, K.R.; Mitchell, K.A. The mis-identification of the major antioxidant flavonoids in young barley (Hordeum vulgare) leaves. J. Biosci. 2003, 58, 53–56. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Rios-Gonzalez, K.; Veisz, O.; Paldi, E. Comparative study of frost tolerance and antioxidant activity in cereals. Plant Sci. 2003, 164, 301–306. [Google Scholar] [CrossRef]
- Yu, Y.M.; Chang, W.C.; Chang, C.T.; Hsieh, C.L.; Tsai, C.E. Effects of young barley leaf extract and antioxidant vitamins on LDL oxidation and free radical scavenging activities in type 2 diabetes. Diabetes Metab. 2002, 28, 107–114. [Google Scholar] [PubMed]
- Yi, B.; Kasai, H.; Lee, H.S.; Kang, Y.; Park, J.Y.; Yang, M. Inhibition by wheat sprout (Triticum aestivum) Juice of bisphenol A-Induced oxidative stress in young women. Mutat. Res. 2011, 724, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ayre, E.; Goldin, E.; Wengrower, D.; Stamper, A.; Kohn, R.; Berry, E. Wheat Grass Juice in the Treatment of Active Distal Ulcerative Colitis: A Randomised Double-Blind Placebo-Controlled Trial. Scand. J. Gastroenterol. 2002, 37, 444–449. [Google Scholar] [CrossRef]
- Fogarty, M.C.; Hughes, C.M.; Burke, G.; Brown, J.C.; Davison, G.W. Acute and chronic watercress supplementation attenuates exercise-induced peripheral mononuclear cell DNA damage and lipid peroxidation. Br. J. Nutr. 2012, 109, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, S.R.; McAnulty, L.S.; Nieman, D.C.; Morrow, J.D.; Shooter, L.A.; Holmes, S.; Heward, C.; Henson, D.A. Effect of alpha-tocopherol supplementation on plasma homocysteine and oxidative stress in highly trained athletes before and after exhaustive exercise. J. Nutr. Biochem. 2005, 16, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Rokitzki, L.; Logemann, E.; Huber, G.; Keck, E.; Keul, J. Alpha-tocopherol supplementation in racing cyclists during extreme endurance training. Int. J. Sport Nutr. Exerc. Metab. 1994, 4, 253–264. [Google Scholar] [CrossRef]
- Subudhi, A.W.; Mattson, J.P. Effects of Antioxidant Supplementation on Oxidative Stress in Trained Cyclists; USANA Health Sciences: Salt Lake City, UT, USA, 2000. [Google Scholar]
- Davison, G.W.; George, L.; Jackson, S.K.; Young, I.S.; Davies, B.; Bailey, D.M.; Peters, J.R.; Ashton, T. Exercise, Free Radicals, and Lipid Peroxidation in Type 1 Diabetes Mellitus. Free Radic. Biol. Med. 2002, 33, 1543–1551. [Google Scholar] [CrossRef]
- Long, W.; Wells, K.; Englert, V. Does prior acute exercise affect post exercise substrate oxidation in response to a high carbohydrate meal? Nutr. Metab. 2008, 5, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Haegele, A.D.; Gillette, C.; O’Neill, C. Plasma xanthophyll carotenoids correlate inversely with indices of oxidative DNA damage and lipid peroxidation. Cancer Epidemiol. Biomark. Prev. 2000, 9, 421–425. [Google Scholar]
- Eastep, J.; Chen, G. The Relationships of High-Fat and Metabolism of Lipophilic Vitamins. Integr. Food Nutr. Metab. 2015, 2, 174–179. [Google Scholar] [CrossRef]
- Mayne, S.T. Antioxidant Nutrients and Chronic Disease: Use of Biomarkers of Exposure and Oxidative Stress Status in Epidemiologic Research. J. Nutr. 2003, 133, 933S–940S. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, M.A.; Hammouda, O.; Matran, R.; Robin, S.; Fabre, C. Changes in Oxidative Stress Markers and Biological Markers of Muscle Injury with Aging at Rest and in Response to an Exhaustive Exercise. PLoS ONE 2014, 9, e90420. [Google Scholar] [CrossRef] [PubMed]
- Khassaf, M.; Child, R.B.; McArdle, A.; Brodie, D.A.; Esanu, C.; Jackson, M.J. Time course of responses of human skeletal muscle to oxidative stress induced by nondamaging exercise. J. Appl. Physiol. 2001, 90, 1031–1035. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant Characteristics | Baseline Data | ||
|---|---|---|---|
| Age (yrs) | 21.5 ± 2 | ||
| Height (cm) | 180.5 ± 6 | ||
| Weight (kg) | 86.6 ± 8 | ||
| Resting HR (bpm) | 69.5 ± 5 | ||
| Exercise Characteristics | Placebo | Low | High |
| Maximum HR (bpm) | 189 ± 8 | 187 ± 8 | 189 ± 8 |
| V̇O2max (mL kg−1 min−1) | 55.1 ± 6 | 57.2 ± 5 | 55.4 ± 7 |
| TTE (mins) | 9.3 ± 2 | 8.4 ± 2 | 8.5 ± 2 |
| Lipid Soluble Antioxidants | Baseline | Rest | Exercise | Δ% (B−R) | Δ% (R−E) |
|---|---|---|---|---|---|
| α-Tocopherol | |||||
| Placebo | 20.90 ± 1.7 | 21.17 ± 4.1 | 23.76 ± 4.3 | 1.3 | 12.2 # |
| Low | 20.90 ± 1.7 | 21.71 ± 3.5 | 22.97 ± 3.8 | 3.9 | 5.8 # |
| High | 20.90 ± 1.7 | 21.30 ± 3.8 | 22.53 ± 3.5 | 1.9 | 5.8 # |
| γ-Tocopherol | |||||
| Placebo | 1.19 ± 0.4 | 1.44 ± 0.1 | 1.83 ± 0.7 | 21.0 | 27.1 * |
| Low | 1.19 ± 0.4 | 1.25 ± 0.4 | 1.17 ± 0.3 | 5.0 | −6.4 |
| High | 1.19 ± 0.4 | 1.44 ± 0.5 | 1.41 ± 0.5 | 21.0 | −2.1 |
| α-Carotene | |||||
| Placebo | 0.05 ± 0.01 | 0.05 ± 0.01 | 0.05 ± 0.01 | 0 | 0 |
| Low | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 | −20.0 | 0 |
| High | 0.05 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0 | −20.0 |
| Retinol | |||||
| Placebo | 1.96 ± 0.4 | 1.94 ± 0.5 | 2.20 ± 0.5 | −1.0 | 13.4 |
| Low | 1.96 ± 0.4 | 1.94 ± 0.4 | 2.14 ± 0.6 | −1.0 | 10.3 |
| High | 1.96 ± 0.4 | 2.00 ± 0.5 | 2.32 ± 0.3 | 2.0 | 16.0 |
| Xanthophyll | |||||
| Placebo | 0.28 ± 0.1 | 0.30 ± 0.1 | 0.37 ± 0.1 | 7.1 | 23.3 # |
| Low | 0.28 ± 0.1 | 0.50 ± 0.3 | 0.54 ± 0.3 | 78.6 # | 8.0 # |
| High | 0.28 ± 0.1 | 0.45 ± 0.1 | 0.51 ± 0.2 | 60.7 # | 13.3 # |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williamson, J.; Hughes, C.M.; Davison, G.W. Exogenous Plant-Based Nutraceutical Supplementation and Peripheral Cell Mononuclear DNA Damage Following High Intensity Exercise. Antioxidants 2018, 7, 70. https://doi.org/10.3390/antiox7050070
Williamson J, Hughes CM, Davison GW. Exogenous Plant-Based Nutraceutical Supplementation and Peripheral Cell Mononuclear DNA Damage Following High Intensity Exercise. Antioxidants. 2018; 7(5):70. https://doi.org/10.3390/antiox7050070
Chicago/Turabian StyleWilliamson, Josh, Ciara M. Hughes, and Gareth W. Davison. 2018. "Exogenous Plant-Based Nutraceutical Supplementation and Peripheral Cell Mononuclear DNA Damage Following High Intensity Exercise" Antioxidants 7, no. 5: 70. https://doi.org/10.3390/antiox7050070
APA StyleWilliamson, J., Hughes, C. M., & Davison, G. W. (2018). Exogenous Plant-Based Nutraceutical Supplementation and Peripheral Cell Mononuclear DNA Damage Following High Intensity Exercise. Antioxidants, 7(5), 70. https://doi.org/10.3390/antiox7050070