

Integration of Transcriptomic Analysis, Network Pharmacology, and Experimental Validation Demonstrates Enhanced Muscle-Protective Effects of Ethanol Extract of Jakyak-Gamcho-Tang
 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Herbal Materials
2.3. Preparation of JGT-W and JGT-E
2.4. C2C12 Cell Culture and Differentiation
2.5. Assessment of Cell Viability and Myotube Degradation in C2C12 Cells
2.6. Transcriptomic Analysis
2.7. Detection of Intracellular ROS Level
2.8. Measurement of Mitochondrial Mass
2.9. Free Radical Scavenging Assay
2.10. Quantitative Analysis of JGT Using UHPLC-MS/MS
2.11. Network Pharmacology Analysis
2.12. Statistical Analysis
3. Results
3.1. Transcriptomic Analysis Shows That JGT-E Regulated Muscle-Protective Pathways Under Oxidative Stress More Effectively than Jgt-W
3.2. JGT-E Demonstrated Greater Efficacy than JGT-W in Mitigating Cell Damage and ROS Production in C2C12 Myoblasts Under Oxidative Stress
3.3. JGT-E Was More Effective than JGE-W in Promoting Mitochondrial Biogenesis, Reducing ROS Levels, and Inhibiting the Degradation of C2C12 Myotubes Exposed to H2O2
3.4. Quantitative Analysis of JGT-W and JGT-E Using UHPLC-MS/MS
3.5. Prediction of Potential Compounds in JGT Involved in Muscle-Protective Mechanisms
3.6. Identification of Compounds That Contribute to Protection Against Oxidative Stress and Muscle Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| JGT | Jakyak-gamcho-tang |
| ABTS | 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt |
| BPC | Base peak ion chromatogram |
| DEG | Differentially expressed genes |
| DM | Differentiation medium |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl sulfoxide |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FDR | False discovery rate |
| GM | Growth medium |
| GO | Gene Ontology |
| GOBP | Gene Ontology Biological Process |
| GSEA | Gene Set Enrichment Analysis |
| H2O2 | Hydrogen peroxide |
| JGT | Jakyak-gamcho-tang |
| JGT-E | Ethanol extract of Jakyak-gamcho-tang |
| JGT-W | Water extract of Jakyak-gamcho-tang |
| MRM | Multiple reaction monitoring |
| NES | Normalized enrichment score |
| OD | Optical density |
| PBS | Phosphate-buffered saline |
| P/S | Penicillin and streptomycin |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| SDS | Sodium dodecyl sulfate |
| UHPLC | Ultrahigh-performance liquid chromatography |
References
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug. Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, A.B.; Neri, S.G.R.; Oliveira, R.J.; Bottaro, M.; David, A.C.; Vainshelboim, B.; Lima, R.M. Severity of sarcopenia is associated with postural balance and risk of falls in community-dwelling older women. Exp. Aging Res. 2018, 44, 258–269. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, S.S.; Singh, S.; Dabur, R. Natural products: Potential therapeutic agents to prevent skeletal muscle atrophy. Eur. J. Pharmacol. 2022, 925, 174995. [Google Scholar] [CrossRef]
- Jung, H.W.; Kang, A.N.; Kang, S.Y.; Park, Y.K.; Song, M.Y. The root extract of Pueraria lobata and its main compound, puerarin, prevent obesity by increasing the energy metabolism in skeletal muscle. Nutrients 2017, 9, 33. [Google Scholar] [CrossRef]
- Ota, K.; Fukui, K.; Nakamura, E.; Oka, M.; Ota, K.; Sakaue, M.; Sano, Y.; Takasu, A. Effect of Shakuyaku-kanzo-to in patients with muscle cramps: A systematic literature review. J. Gen. Fam. Med. 2020, 21, 56–62. [Google Scholar] [CrossRef]
- Kim, A.; Kim, Y.R.; Park, S.M.; Lee, H.; Park, M.; Yi, J.M.; Cha, S.; Kim, N.S. Jakyak-gamcho-tang, a decoction of Paeoniae Radix and Glycyrrhizae Radix et Rhizoma, ameliorates dexamethasone-induced muscle atrophy and muscle dysfunction. Phytomedicine 2024, 123, 155057. [Google Scholar] [CrossRef]
- Abubakar, A.R.; Haque, M. Preparation of medicinal plants: Basic extraction and fractionation procedures for experimental purposes. J. Pharm. Bioallied Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Plaskova, A.; Mlcek, J. New insights of the application of water or ethanol-water plant extract rich in active compounds in food. Front. Nutr. 2023, 10, 1118761. [Google Scholar] [CrossRef]
- Stanciauskaite, M.; Marksa, M.; Babickaite, L.; Majiene, D.; Ramanauskiene, K. Comparison of ethanolic and aqueous Populus balsamifera L. Bud Extracts by different extraction methods: Chemical composition, antioxidant and antibacterial activities. Pharmaceuticals 2021, 14, 1018. [Google Scholar] [CrossRef]
- Ju, J.B.; Kim, J.S.; Choi, C.W.; Lee, H.K.; Oh, T.K.; Kim, S.C. Comparison between ethanolic and aqueous extracts from Chinese juniper berries for hypoglycaemic and hypolipidemic effects in alloxan-induced diabetic rats. J. Ethnopharmacol. 2008, 115, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Jose, H.; Homem, V.; Simoes, M. Comparison of techniques and solvents on the antimicrobial and antioxidant potential of extracts from Acacia dealbata and Olea europaea. Antibiotics 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.Y.; Lee, Y.S.; Han, S.Y.; Jeong, E.J.; Lee, M.K.; Kong, J.Y.; Lee, D.H.; Cho, K.J.; Lee, H.S.; Ahn, M.J. A Comparison between water and ethanol extracts of Rumex acetosa for protective effects on gastric ulcers in mice. Biomol. Ther. 2012, 20, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Kim, A.; Lee, H.; Baek, S.J.; Kim, N.S.; Park, M.; Yi, J.M.; Cha, S. Systematic transcriptome analysis reveals molecular mechanisms and indications of bupleuri radix. Front. Pharmacol. 2022, 13, 1010520. [Google Scholar] [CrossRef]
- Park, M.; Park, S.M.; Lee, H.; Kim, A.; Kim, N.S.; Kim, Y.R.; Yi, J.M.; Cha, S. KORE-Map 1.0: Korean Medicine Omics Resource Extension Map on transcriptome data of tonifying herbal medicine. Sci. Data 2024, 11, 974. [Google Scholar] [CrossRef]
- Kim, Y.S.; Yuk, H.J.; Kim, D.S. Effect of Jakyakgamcho-Tang extracts on H2O2-induced C2C12 myoblasts. Molecules 2021, 26, 215. [Google Scholar] [CrossRef]
- Zhang, G.B.; Li, Q.Y.; Chen, Q.L.; Su, S.B. Network pharmacology: A new approach for chinese herbal medicine research. Evid. Based Complement. Alternat. Med. 2013, 2013, 621423. [Google Scholar] [CrossRef]
- Noor, F.; Tahir Ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.S.; Aljasir, M.A. Network pharmacology approach for medicinal plants: Review and assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar] [CrossRef]
- Choi, W.G.; Choi, N.R.; Park, E.J.; Kim, B.J. A study of the therapeutic mechanism of Jakyakgamcho-Tang about functional dyspepsia through network pharmacology research. Int. J. Med. Sci. 2022, 19, 1824–1834. [Google Scholar] [CrossRef]
- Hu, Y.H.; Wang, X.Y.; Zhang, X.W.; Chen, J.; Li, F. Investigation of the mechanisms and experimental verification of Shao yao gan cao decoction against Sphincter of Oddi Dysfunction via systems pharmacology. Math. Biosci. Eng. 2022, 19, 13374–13398. [Google Scholar] [CrossRef]
- Alamdari, N.; Aversa, Z.; Castillero, E.; Gurav, A.; Petkova, V.; Tizio, S.; Hasselgren, P.O. Resveratrol prevents dexamethasone-induced expression of the muscle atrophy-related ubiquitin ligases atrogin-1 and MuRF1 in cultured myotubes through a SIRT1-dependent mechanism. Biochem. Biophys. Res. Commun. 2012, 417, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Bagherniya, M.; Mahdavi, A.; Shokri-Mashhadi, N.; Banach, M.; Von Haehling, S.; Johnston, T.P.; Sahebkar, A. The beneficial therapeutic effects of plant-derived natural products for the treatment of sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 2772–2790. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Shanavas, S.; Chaudhury, D.; Bose, B.; Das, U.N.; Shenoy, P.S. Mitigation of chronic glucotoxicity-mediated skeletal muscle atrophy by arachidonic acid. Life Sci. 2023, 333, 122141. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ji, Y.; Liu, R.; Zhu, X.; Wang, K.; Yang, X.; Liu, B.; Gao, Z.; Huang, Y.; Shen, Y.; et al. Mitochondrial dysfunction: Roles in skeletal muscle atrophy. J. Transl. Med. 2023, 21, 503. [Google Scholar] [CrossRef]
- Popov, L.D. Mitochondrial biogenesis: An update. J. Cell. Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef]
- Sandri, M.; Lin, J.; Handschin, C.; Yang, W.; Arany, Z.P.; Lecker, S.H.; Goldberg, A.L.; Spiegelman, B.M. PGC-1alpha protects skeletal muscle from atrophy by suppressing FoxO3 action and atrophy-specific gene transcription. Proc. Natl. Acad. Sci. USA 2006, 103, 16260–16265. [Google Scholar] [CrossRef]
- Handschin, C.; Spiegelman, B.M. The role of exercise and PGC-1α in inflammation and chronic disease. Nature 2008, 454, 463–469. [Google Scholar] [CrossRef]
- Gatica, D.; Klionsky, D.J. New insights into MTORC1 amino acid sensing and activation. Biotarget 2017, 1, 2. [Google Scholar] [CrossRef]
- Rudrappa, S.S.; Wilkinson, D.J.; Greenhaff, P.L.; Smith, K.; Idris, I.; Atherton, P.J. Human skeletal muscle disuse atrophy: Effects on muscle protein synthesis, breakdown, and insulin resistance-A qualitative review. Front. Physiol. 2016, 7, 361. [Google Scholar] [CrossRef]
- Mendias, C.L.; Gumucio, J.P.; Davis, M.E.; Bromley, C.W.; Davis, C.S.; Brooks, S.V. Transforming growth factor-beta induces skeletal muscle atrophy and fibrosis through the induction of atrogin-1 and scleraxis. Muscle Nerve 2012, 45, 55–59. [Google Scholar] [CrossRef]
- Gurd, B.J. Deacetylation of PGC-1alpha by SIRT1: Importance for skeletal muscle function and exercise-induced mitochondrial biogenesis. Appl. Physiol. Nutr. Metab. 2011, 36, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.N.; Emter, R.; Hock, M.B.; Knutti, D.; Cardenas, J.; Podvinec, M.; Oakeley, E.J.; Kralli, A. The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 6472–6477. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Yang, J.; Jiang, Y.; Wen, L.; Wang, Z.; Yang, B. The antioxidant activity and neuroprotective mechanism of isoliquiritigenin. Free Radic. Biol. Med. 2020, 152, 207–215. [Google Scholar] [CrossRef]
- Wang, T.T.; Chen, Z.Z.; Xie, P.; Zhang, W.J.; Du, M.Y.; Liu, Y.T.; Zhu, H.Y.; Guo, Y.S. Isoliquiritigenin suppresses the proliferation and induced apoptosis via miR-32/LATS2/Wnt in nasopharyngeal carcinoma. Eur. J. Pharmacol. 2019, 856, 172352. [Google Scholar] [CrossRef]
- Al-Qahtani, W.H.; Alshammari, G.M.; Ajarem, J.S.; Al-Zahrani, A.Y.; Alzuwaydi, A.; Eid, R.; Yahya, M.A. Isoliquiritigenin prevents doxorubicin-induced hepatic damage in rats by upregulating and activating SIRT1. Biomed. Pharmacother. 2022, 146, 112594. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, X.; Zhong, B.; Liao, Q.; Wang, X.; Xie, Y.; He, X. Review on the diverse biological effects of glabridin. Drug Des. Devel. Ther. 2023, 17, 15–37. [Google Scholar] [CrossRef]
- Denzer, I.; Munch, G.; Pischetsrieder, M.; Friedland, K. S-allyl-L-cysteine and isoliquiritigenin improve mitochondrial function in cellular models of oxidative and nitrosative stress. Food Chem. 2016, 194, 843–848. [Google Scholar] [CrossRef]
- Yang, E.J.; Min, J.S.; Ku, H.Y.; Choi, H.S.; Park, M.K.; Kim, M.K.; Song, K.S.; Lee, D.S. Isoliquiritigenin isolated from Glycyrrhiza uralensis protects neuronal cells against glutamate-induced mitochondrial dysfunction. Biochem. Biophys. Res. Commun. 2012, 421, 658–664. [Google Scholar] [CrossRef]
- Jiao, X.; Zhang, Y.; Wang, Z.; Chang, H.; Li, Y.; Wang, B.; Gan, Y.; Gu, D. Isoliquiritigenin, an extract from Licorice, attenuates dexamethasone-induced muscle atrophy via Akt/mTOR pathway. Mol. Nutr. Food Res. 2025, 69, e70000. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, A.; Xiong, W.; Lin, H.; Xiao, W.; Huang, J.; Zhang, S.; Liu, Z. Catechins enhance skeletal muscle performance. Crit. Rev. Food Sci. Nutr. 2020, 60, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, K.M.; Byun, M.R.; Hwang, J.H.; Park, J.I.; Oh, H.T.; Kim, H.K.; Jeong, M.G.; Hwang, E.S.; Hong, J.H. Catechins activate muscle stem cells by Myf5 induction and stimulate muscle regeneration. Biochem. Biophys. Res. Commun. 2017, 489, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, A.; Liu, C.; Qu, Z.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Role and mechanism of catechin in skeletal muscle cell differentiation. J. Nutr. Biochem. 2019, 74, 108225. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Kubota, Y.; Samukawa, Y.; Yamashita, Y.; Ashida, H. Glabridin inhibits dexamethasone-induced muscle atrophy. Arch. Biochem. Biophys. 2019, 664, 157–166. [Google Scholar] [CrossRef]
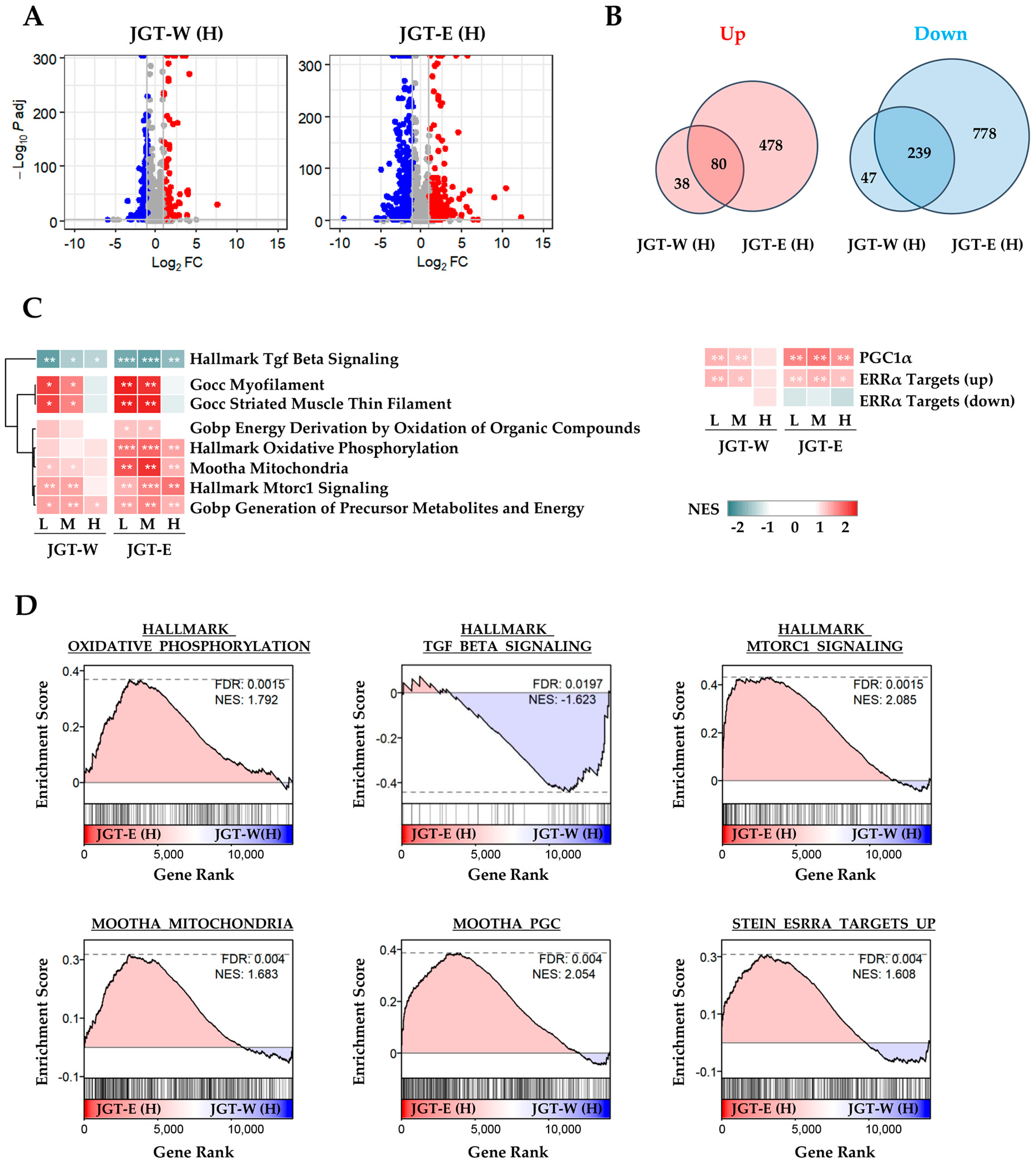


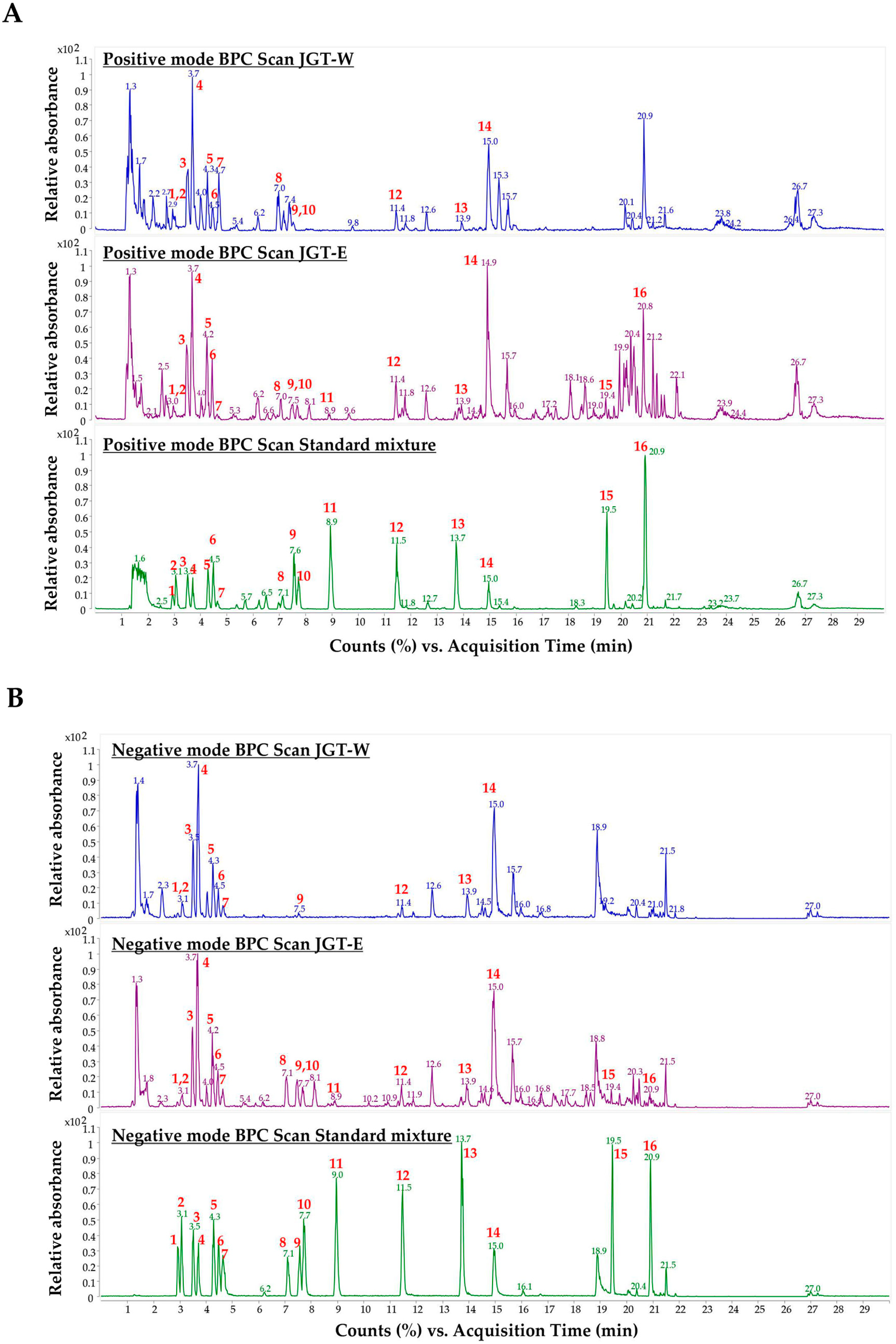
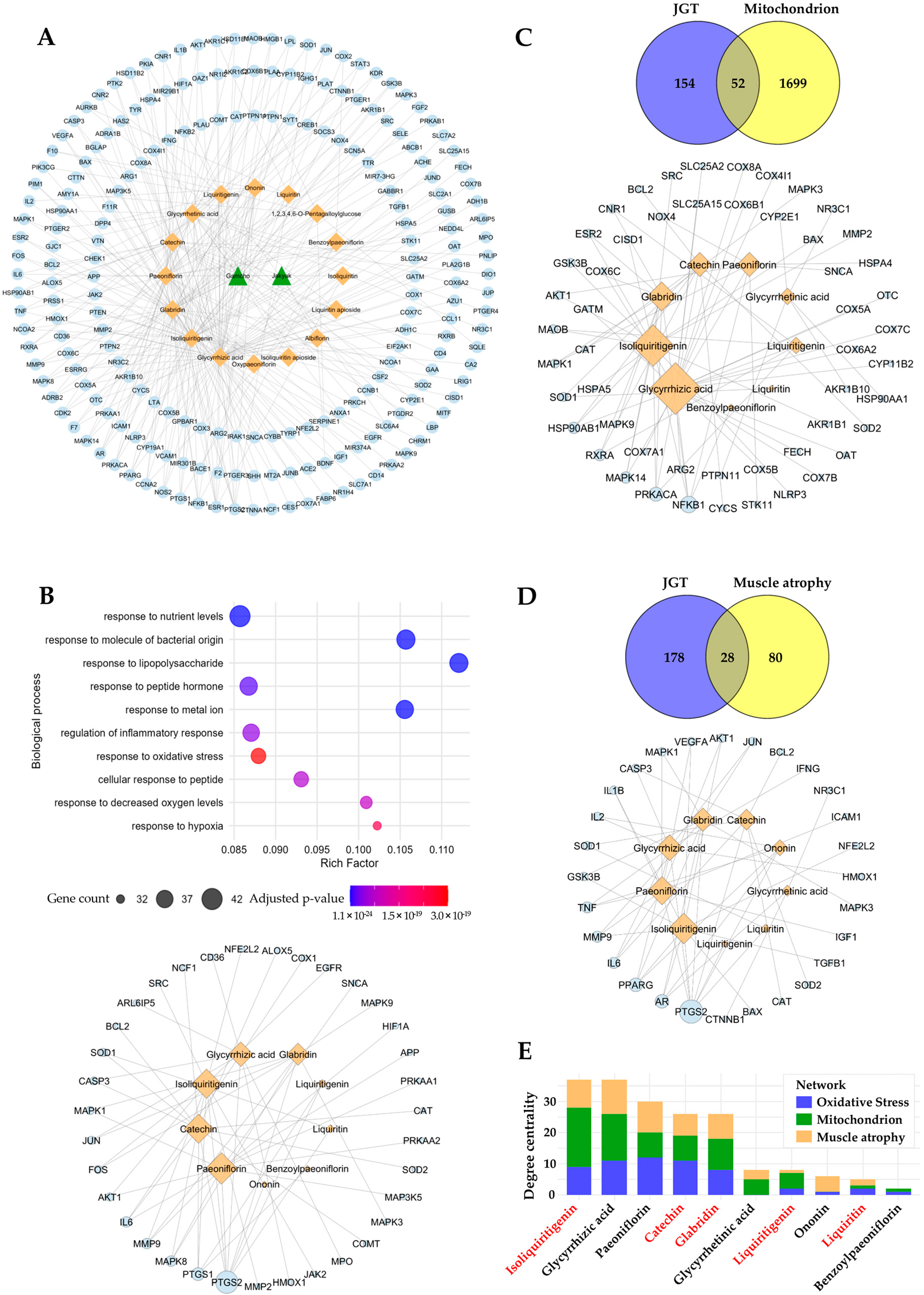

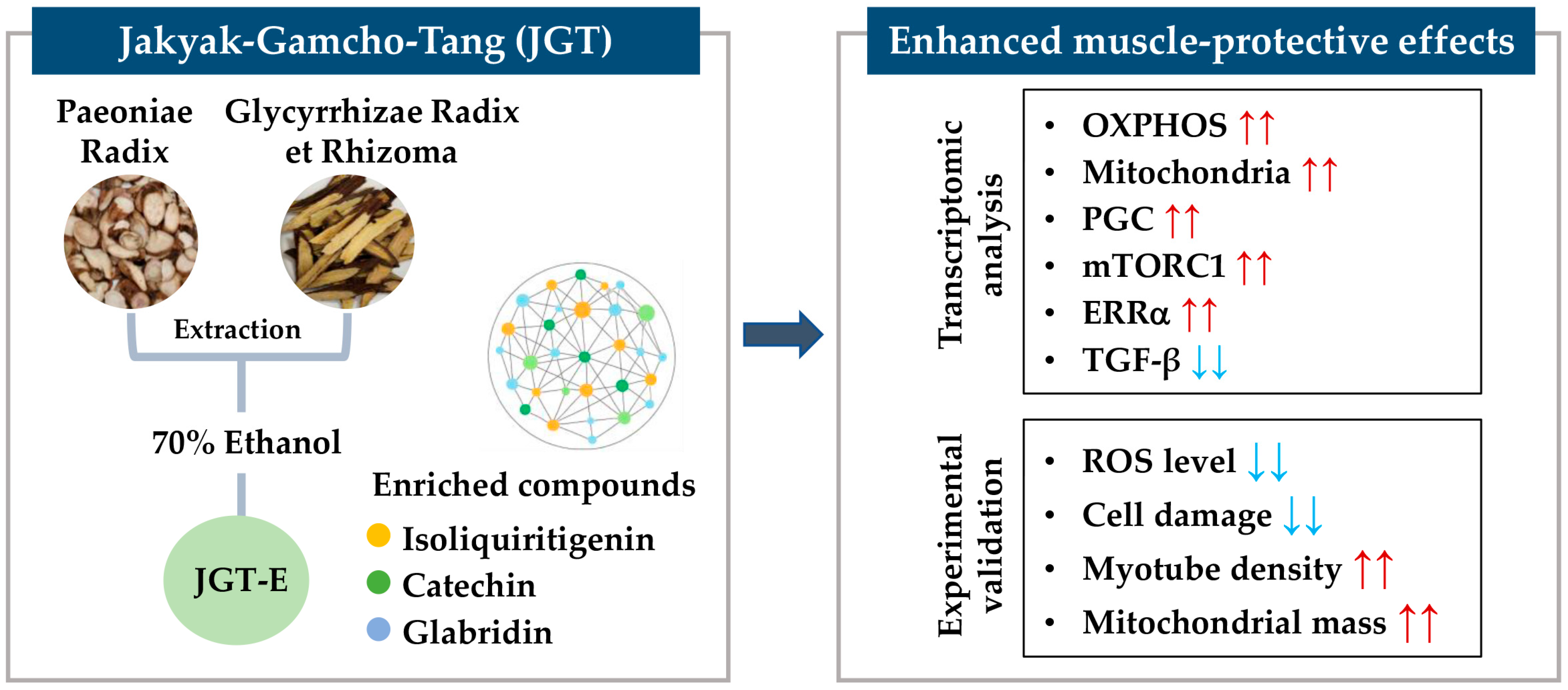

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Components | Rt (min) | Precursor Ion (m/z) | Product Ion (m/z) | Collision Energy (V) | Ion Polarity |
|---|---|---|---|---|---|---|
| 1 | Oxypaeoniflorin | 2.985 | 495.1 | 137.1 | 26.0 | Negative |
| 2 | (+)-Catechin | 3.115 | 291.1 | 139.0 | 14.0 | Positive |
| 3 | Albiflorin | 3.566 | 481.2 | 105.0 | 22.0 | Positive |
| 4 | Paeoniflorin | 3.759 | 525.2 | 449.1 | 10.0 | Negative |
| 5 | Liquiritin apioside | 4.341 | 549.2 | 255.1 | 34.0 | Negative |
| 6 | Liquiritin | 4.535 | 417.2 | 255.0 | 20.0 | Negative |
| 7 | 1,2,3,4,6-Pentagalloyl glucose | 4.680 | 958.1 | 153.0 | 40.0 | Positive |
| 8 | Isoliquiritin apioside | 7.169 | 549.1 | 255.1 | 30.0 | Negative |
| 9 | Ononin | 7.634 | 431.1 | 269.1 | 18.0 | Positive |
| 10 | Isoliquiritin | 7.789 | 417.0 | 255.1 | 18.0 | Negative |
| 11 | Liquiritigenin | 9.025 | 257.0 | 137.0 | 26.0 | Positive |
| 12 | Benzoylpaeoniflorin | 11.531 | 629.1 | 553.2 | 10.0 | Negative |
| 13 | Isoliquiritigenin | 13.811 | 257.0 | 137.0 | 22.0 | Positive |
| 14 | Glycyrrhizin | 15.030 | 821.4 | 351.0 | 40.0 | Negative |
| 15 | Glabridin | 19.529 | 325.1 | 189.1 | 14.0 | Positive |
| 16 | Glycyrrhetinic acid | 21.004 | 471.2 | 189.1 | 38.0 | Positive |
| No. | Components | Regression Equation | R2 | Range (μg/mL) | Concentration (mg/g, n = 6) | |
|---|---|---|---|---|---|---|
| JGT-W | JGT-E | |||||
| 1 | Oxypaeoniflorin | y = 86,522x + 6367 | 0.9990 | 0.16–2.50 | 0.959 (±0.024) | 0.875 (±0.012) |
| 2 | (+)-Catechin | y = 484,425x + 23,507 | 0.9990 | 0.08–2.5 | 0.089 (±0.003) | 1.101 (±0.016) |
| 3 | Albiflorin | y = 590,324x + 8051 | 0.9992 | 0.04–5.00 | 15.141 (±1.636) | 14.881 (±0.675) |
| 4 | Paeoniflorin | y = 205,795x + 93,092 | 0.9990 | 0.63–10.00 | 66.180 (±6.085) | 70.331 (±3.824) |
| 5 | Liquiritin apioside | y = 379,464x + 22,860 | 0.9991 | 0.04–2.50 | 9.013 (±0.931) | 10.271 (±0.748) |
| 6 | Liquiritin | y = 459,755x + 37,366 | 0.9990 | 0.08–1.25 | 5.158 (±0.461) | 6.956 (±0.584) |
| 7 | 1,2,3,4,6-Pentagalloyl glucose | y = 220,570x + 3267 | 0.9991 | 0.04–5.00 | 0.295 (±0.022) | 2.240 (±0.047) |
| 8 | Isoliquiritin apioside | y = 162,498x + 80,392 | 0.9991 | 0.63–10.00 | 0.165 (±0.013) | 6.092 (±0.066) |
| 9 | Ononin | y = 2,566,720x + 52,6143 | 0.9991 | 0.31–5.00 | 3.896 (±0.039) | 4.234 (±0.069) |
| 10 | Isoliquiritin | y = 406,963x + 15,5124 | 0.9991 | 0.31–5.00 | tr * | 1.849 (±0.029) |
| 11 | Liquiritigenin | y = 1,525,781x + 42,803 | 0.9992 | 0.04–1.25 | tr | 1.038 (±0.013) |
| 12 | Benzoylpaeoniflorin | y = 306,585x + 50,791 | 0.9990 | 0.16–5.00 | 0.643 (±0.015) | 0.975 (±0.013) |
| 13 | Isoliquiritigenin | y = 1,244,869x + 29,884 | 0.9992 | 0.04–2.50 | 0.006 (±0.002) | 1.184 (±0.017) |
| 14 | Glycyrrhizin | y = 80,641x + 18,808 | 0.9990 | 0.16–10.00 | 50.324 (±4.754) | 75.053 (±4.004) |
| 15 | Glabridin | y = 1,288,421x + 10,894 | 0.9996 | 0.04–0.63 | 0.022 (±0.001) | 0.838 (±0.077) |
| 16 | Glycyrrhetinic acid | y = 558,961x + 28,138 | 0.9990 | 0.08–1.25 | tr | 0.241 (±0.004) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, A.; Tran, M.N.; Lee, A.Y.; Yeo, H.; Baek, S.-J.; Kim, N.S.; Cha, S.; Park, S.-M. Integration of Transcriptomic Analysis, Network Pharmacology, and Experimental Validation Demonstrates Enhanced Muscle-Protective Effects of Ethanol Extract of Jakyak-Gamcho-Tang. Antioxidants 2025, 14, 795. https://doi.org/10.3390/antiox14070795
Kim A, Tran MN, Lee AY, Yeo H, Baek S-J, Kim NS, Cha S, Park S-M. Integration of Transcriptomic Analysis, Network Pharmacology, and Experimental Validation Demonstrates Enhanced Muscle-Protective Effects of Ethanol Extract of Jakyak-Gamcho-Tang. Antioxidants. 2025; 14(7):795. https://doi.org/10.3390/antiox14070795
Chicago/Turabian StyleKim, Aeyung, Minh Nhat Tran, A Yeong Lee, Heerim Yeo, Su-Jin Baek, No Soo Kim, Seongwon Cha, and Sang-Min Park. 2025. "Integration of Transcriptomic Analysis, Network Pharmacology, and Experimental Validation Demonstrates Enhanced Muscle-Protective Effects of Ethanol Extract of Jakyak-Gamcho-Tang" Antioxidants 14, no. 7: 795. https://doi.org/10.3390/antiox14070795
APA StyleKim, A., Tran, M. N., Lee, A. Y., Yeo, H., Baek, S.-J., Kim, N. S., Cha, S., & Park, S.-M. (2025). Integration of Transcriptomic Analysis, Network Pharmacology, and Experimental Validation Demonstrates Enhanced Muscle-Protective Effects of Ethanol Extract of Jakyak-Gamcho-Tang. Antioxidants, 14(7), 795. https://doi.org/10.3390/antiox14070795