

Antioxidant Intervention Against Microplastic Hazards

 ,
,
Abstract
1. Introduction
2. Microplastics: Sources and Impact on Health
2.1. Sources of Microplastics
2.2. Impact on Human Health
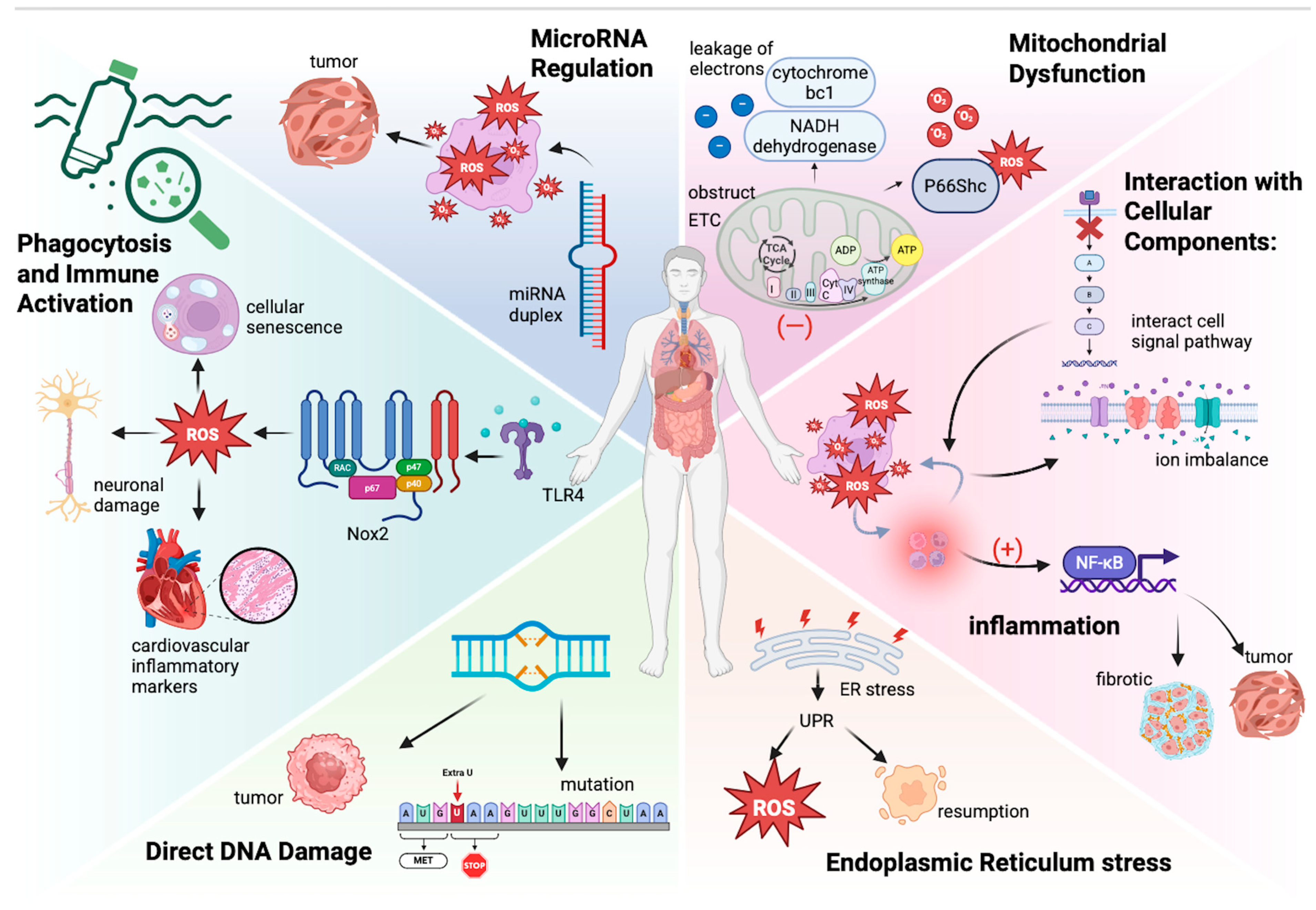
3. Oxidative Stress Mechanisms Induced by Microplastics
3.1. Understanding Oxidative Stress
3.2. Molecular Mechanisms of Microplastic-Induced Oxidative Stress
3.3. Evidence of Oxidative Stress Induction
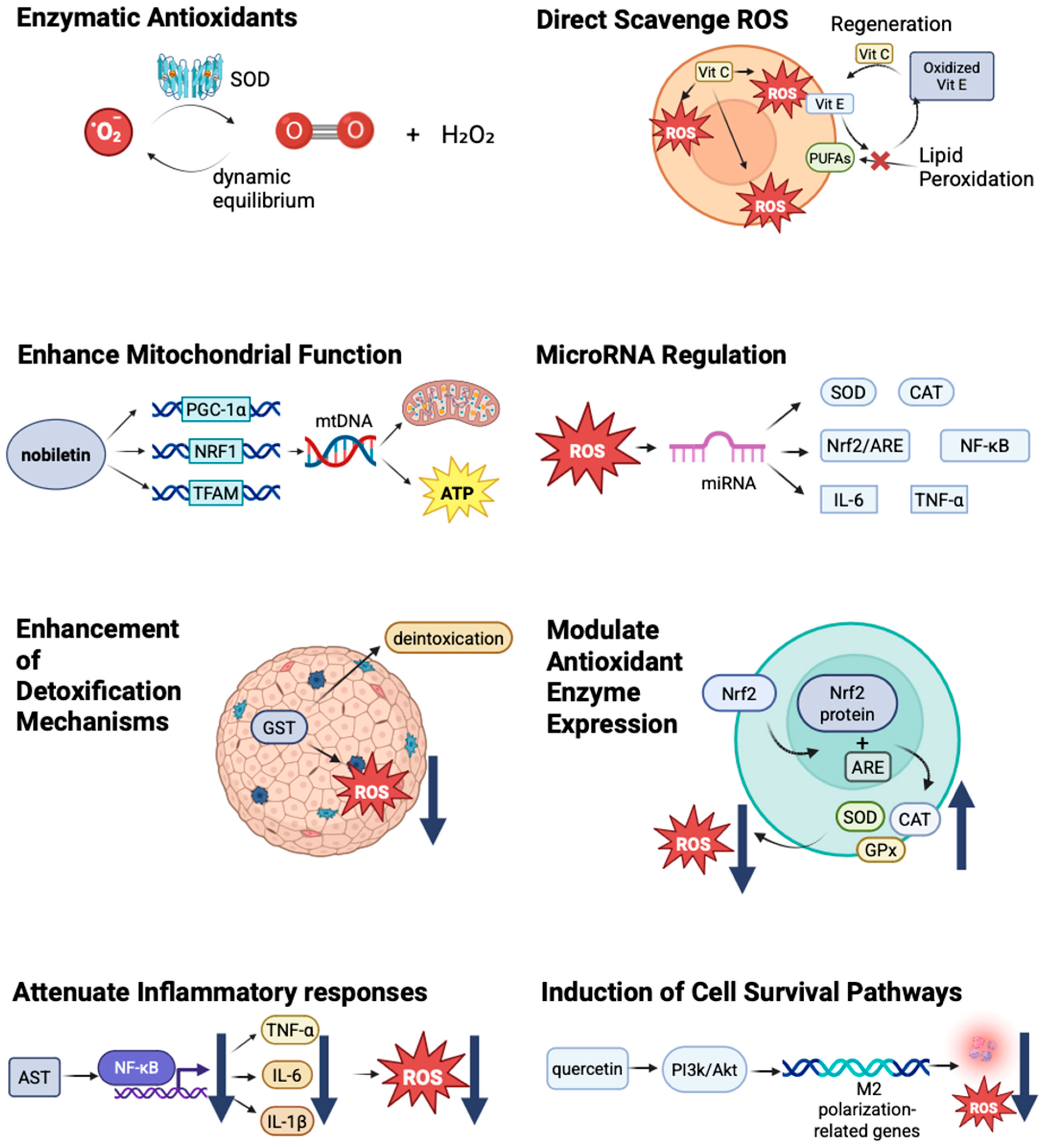
4. Antioxidant Interventions: Mechanisms and Efficacy
4.1. Types of Antioxidants
4.2. Mechanisms of Action
4.3. Evidence Supporting Antioxidant Efficacy
4.3.1. In Vitro Studies
4.3.2. In Vivo Studies
4.4. Mechanistic Insights
5. Advantages of Antioxidant Interventions
5.1. Multifaceted Protection
5.2. Nutritional Accessibility
5.3. Potential for Public Health Strategies
5.4. Low Toxicity and Side Effects
5.5. Cost Effectiveness
6. Challenges and Limitations of Antioxidant Interventions
6.1. Variability in Efficacy
6.2. Bioavailability Concerns
6.3. Lack of Standardization in Research Protocols
6.4. Long-Term Effects Require Further Investigation
6.5. Complexity of Mixtures
6.6. Lack of Vitro and Vivo Experiments
6.7. Antioxidant Toxicity
6.8. Antioxidant Contraindications
6.9. Lack of Human Exposure Research
6.10. Lack of Relevant Human Epidemiological Studies
7. Future Directions
7.1. Understanding Molecular Pathways
7.2. Development of Novel Antioxidant Formulations
7.3. Mechanistic Studies on Antioxidants
7.4. Interdisciplinary Approaches
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| GPx | Glutathione peroxidase |
| PVC | Polyvinyl chloride |
| PS | Polystyrene |
| PLA | Polylactide acid |
| Th2 | T-helper type 2 |
| MPs | Microplastics |
| NPs | Nanoplastics |
| ETC | Electron transport chain |
| mPTP | Mitochondrial permeability transition pore |
| BPA | Bisphenol A |
| MDA | Malondialdehyde |
| BER | Base excision repair |
| NER | Nucleotide excision repair |
| PS-MPs | Polystyrene microplastics |
| PE-NPs | Polyethylene nanoplastics |
| ER | Endoplasmic reticulum |
| UPR | Unfolded protein response |
| miRNAs | MicroRNAs |
| GSH | Glutathione |
| PUFAs | Polyunsaturated fatty acids |
| ARE | Antioxidant response element |
| SFN | Sulforaphane |
| MMP | Mitochondrial membrane potential |
| AST | Astaxanthin |
| ALT | Alanine aminotransferase |
| I/R | Ischemia/reperfusion |
| GST | Glutathione S-transferase |
| T2D | Type 2 diabetes |
| AAC | Abdominal aortic calcification |
| QSN | Quercetin |
| MT | Melatonin |
| PE-MPs | Polyethylene microplastics |
| NAC | N-acetylcysteine |
| Gly | Glutathione |
| SIR 2.1 | Sirtuin 2.1 |
| HLH-30 | Helix–loop–helix 30 |
| PP-MPs | Polypropylene microplastics |
| CoQ10 | Coenzyme Q10 |
References
- Schmid, C.; Cozzarini, L.; Zambello, E. Microplastic’s story. Mar. Pollut. Bull. 2021, 162, 111820. [Google Scholar] [CrossRef] [PubMed]
- Brahney, J.; Hallerud, M.; Heim, E.; Hahnenberger, M.; Sukumaran, S. Plastic rain in protected areas of the United States. Science 2020, 368, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, H.T.; MacDonald, C.; Santos, R.G.; Ali, R.; Bobat, A.; Cresswell, B.J.; Francini-Filho, R.; Freitas, R.; Galbraith, G.F.; Musembi, P.; et al. Plastic pollution on the world’s coral reefs. Nature 2023, 619, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Courtene-Jones, W.; Boucher, J.; Pahl, S.; Raubenheimer, K.; Koelmans, A.A. Twenty years of microplastic pollution research-what have we learned? Science 2024, 386, eadl2746. [Google Scholar] [CrossRef]
- Thompson, R.C.; Swan, S.H.; Moore, C.J.; vom Saal, F.S. Our plastic age. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 1973–1976. [Google Scholar] [CrossRef]
- Seo, J.H.; Shin, Y.; Song, I.-g.; Lim, J.; Ok, Y.S.; Weon, S. Atmospheric microplastics: Challenges in site- and target-specific measurements. TrAC Trends Anal. Chem. 2024, 178, 117859. [Google Scholar] [CrossRef]
- Debnath, R.; Prasad, G.S.; Amin, A.; Malik, M.M.; Ahmad, I.; Abubakr, A.; Borah, S.; Rather, M.A.; Impellitteri, F.; Tabassum, I.; et al. Understanding and addressing microplastic pollution: Impacts, mitigation, and future perspectives. J. Contam. Hydrol. 2024, 266, 104399. [Google Scholar] [CrossRef]
- Kurniawan, T.A.; Haider, A.; Ahmad, H.M.; Mohyuddin, A.; Umer Aslam, H.M.; Nadeem, S.; Javed, M.; Othman, M.H.D.; Goh, H.H.; Chew, K.W. Source, occurrence, distribution, fate, and implications of microplastic pollutants in freshwater on environment: A critical review and way forward. Chemosphere 2023, 325, 138367. [Google Scholar] [CrossRef]
- Kumar, K.; Umapathi, R.; Ghoreishian, S.M.; Tiwari, J.N.; Hwang, S.K.; Huh, Y.S.; Venkatesu, P.; Shetti, N.P.; Aminabhavi, T.M. Microplastics and biobased polymers to combat plastics waste. Chemosphere 2023, 341, 140000. [Google Scholar] [CrossRef]
- Bhatt, P.; Pathak, V.M.; Bagheri, A.R.; Bilal, M. Microplastic contaminants in the aqueous environment, fate, toxicity consequences, and remediation strategies. Environ. Res. 2021, 200, 111762. [Google Scholar] [CrossRef]
- Sarkar, B.; Dissanayake, P.D.; Bolan, N.S.; Dar, J.Y.; Kumar, M.; Haque, M.N.; Mukhopadhyay, R.; Ramanayaka, S.; Biswas, J.K.; Tsang, D.C.W.; et al. Challenges and opportunities in sustainable management of microplastics and nanoplastics in the environment. Environ. Res. 2022, 207, 112179. [Google Scholar] [CrossRef]
- Hatwar, N.; Qureshi, A. Comprehensive Review on Bio-Based Treatments for Polyvinyl Chloride Plastic. Appl. Biochem. Biotechnol. 2025, 197, 2769–2798. [Google Scholar] [CrossRef]
- Zhao, B.; Rehati, P.; Yang, Z.; Cai, Z.; Guo, C.; Li, Y. The potential toxicity of microplastics on human health. Sci. Total Environ. 2024, 912, 168946. [Google Scholar] [CrossRef]
- Ijaz, M.U.; Rafi, Z.; Hamza, A.; Sayed, A.A.; Albadrani, G.M.; Al-Ghadi, M.Q.; Abdel-Daim, M.M. Mitigative potential of kaempferide against polyethylene microplastics induced testicular damage by activating Nrf-2/Keap-1 pathway. Ecotoxicol. Environ. Saf. 2024, 269, 115746. [Google Scholar] [CrossRef]
- Chen, W.; Zhu, R.; Ye, X.; Sun, Y.; Tang, Q.; Liu, Y.; Yan, F.; Yu, T.; Zheng, X.; Tu, P. Food-derived cyanidin-3-O-glucoside reverses microplastic toxicity via promoting discharge and modulating the gut microbiota in mice. Food Funct. 2022, 13, 1447–1458. [Google Scholar] [CrossRef]
- Yu, J.; Yoon, J.H.; Park, M.; Lee, H.J. Nobiletin-mediated autophagy mitigates nanoplastic-induced toxicity in human intestinal Caco-2 cells. FASEB J. 2025, 39, e70452. [Google Scholar] [CrossRef]
- Peng, B.Y.; Sun, Y.; Li, P.; Yu, S.; Xu, Y.; Chen, J.; Zhou, X.; Wu, W.M.; Zhang, Y. Biodegradation of polyvinyl chloride, polystyrene, and polylactic acid microplastics in Tenebrio molitor larvae: Physiological responses. J. Environ. Manag. 2023, 345, 118818. [Google Scholar] [CrossRef]
- Rizwan, A.; Ijaz, M.U.; Hamza, A.; Anwar, H. Attenuative effect of astilbin on polystyrene microplastics induced testicular damage: Biochemical, spermatological and histopathological-based evidences. Toxicol. Appl. Pharmacol. 2023, 471, 116559. [Google Scholar] [CrossRef]
- Qu, M.; Chen, H.; Lai, H.; Liu, X.; Wang, D.; Zhang, X. Exposure to nanopolystyrene and its 4 chemically modified derivatives at predicted environmental concentrations causes differently regulatory mechanisms in nematode Caenorhabditis elegans. Chemosphere 2022, 305, 135498. [Google Scholar] [CrossRef]
- Khan, K.Y.; Tang, Y.; Cheng, P.F.; Song, Y.L.; Li, X.Y.; Lou, J.B.; Iqbal, B.; Zhao, X.; Hameed, R.; Li, G.L.; et al. Effects of degradable and non-degradable microplastics and oxytetracycline co-exposure on soil N2O and CO2 emissions. Appl. Soil Ecol. 2024, 197, 105331. [Google Scholar] [CrossRef]
- Zhang, T.T.; Luo, X.S.; Xu, J.B.; Yao, X.W.; Fan, J.Y.; Mao, Y.D.; Song, Y.J.; Yang, J.S.; Pan, J.Y.; Khattak, W.A. Dry-wet cycle changes the influence of microplastics (MPs) on the antioxidant activity of lettuce and the rhizospheric bacterial community. J. Soils Sediments 2023, 23, 2189–2201. [Google Scholar] [CrossRef]
- Choudhury, A.; Simnani, F.Z.; Singh, D.; Patel, P.; Sinha, A.; Nandi, A.; Ghosh, A.; Saha, U.; Kumari, K.; Jaganathan, S.K.; et al. Atmospheric microplastic and nanoplastic: The toxicological paradigm on the cellular system. Ecotoxicol. Environ. Saf. 2023, 259, 115018. [Google Scholar] [CrossRef] [PubMed]
- Schwabl, P.; Köppel, S.; Königshofer, P.; Bucsics, T.; Trauner, M.; Reiberger, T.; Liebmann, B. Detection of Various Microplastics in Human Stool: A Prospective Case Series. Ann. Intern. Med. 2019, 171, 453–457. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef]
- Wright, S.L.; Kelly, F.J. Plastic and Human Health: A Micro Issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef]
- Zuri, G.; Karanasiou, A.; Lacorte, S. Human biomonitoring of microplastics and health implications: A review. Environ. Res. 2023, 237, 116966. [Google Scholar] [CrossRef]
- Shaoyong, W.; Jin, H.; Jiang, X.; Xu, B.; Liu, Y.; Wang, Y.; Jin, M. Benzo [a] pyrene-loaded aged polystyrene microplastics promote colonic barrier injury via oxidative stress-mediated notch signalling. J. Hazard. Mater. 2023, 457, 131820. [Google Scholar] [CrossRef]
- Cheng, W.; Li, X.; Zhou, Y.; Yu, H.; Xie, Y.; Guo, H.; Wang, H.; Li, Y.; Feng, Y.; Wang, Y. Polystyrene microplastics induce hepatotoxicity and disrupt lipid metabolism in the liver organoids. Sci. Total Environ. 2022, 806, 150328. [Google Scholar] [CrossRef]
- Sangkham, S.; Faikhaw, O.; Munkong, N.; Sakunkoo, P.; Arunlertaree, C.; Chavali, M.; Mousazadeh, M.; Tiwari, A. A review on microplastics and nanoplastics in the environment: Their occurrence, exposure routes, toxic studies, and potential effects on human health. Mar. Pollut. Bull. 2022, 181, 113832. [Google Scholar] [CrossRef]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef]
- Iqbal, B.; Zhao, T.T.; Yin, W.Q.; Zhao, X.; Xie, Q.J.; Khan, K.Y.; Zhao, X.X.; Nazar, M.; Li, G.L.; Du, D.L. Impacts of soil microplastics on crops: A review. Appl. Soil Ecol. 2023, 181, 104680. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Huang, Z.Y.; Li, Y.F.; Lu, X.L.; Li, G.R.; Qi, S.S.; Khan, I.U.; Li, G.L.; Dai, Z.C.; Du, D.L. The Degradability of Microplastics May Not Necessarily Equate to Environmental Friendliness: A Case Study of Cucumber Seedlings with Disturbed Photosynthesis. Agriculture 2024, 14, 53. [Google Scholar] [CrossRef]
- La Porta, E.; Exacoustos, O.; Lugani, F.; Angeletti, A.; Chiarenza, D.S.; Bigatti, C.; Spinelli, S.; Kajana, X.; Garbarino, A.; Bruschi, M.; et al. Microplastics and Kidneys: An Update on the Evidence for Deposition of Plastic Microparticles in Human Organs, Tissues and Fluids and Renal Toxicity Concern. Int. J. Mol. Sci. 2023, 24, 4391. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, S.A.M.; Elashal, M.; Kieliszek, M.; Ghazala, N.E.; Farag, M.A.; Saeed, A.; Xiao, J.B.; Zou, X.B.; Khatib, A.; Göransson, U.; et al. Recent insights into chemical and pharmacological studies of bee bread. Trends Food Sci. Technol. 2020, 97, 300–316. [Google Scholar] [CrossRef]
- Dong, C.D.; Chen, C.W.; Chen, Y.C.; Chen, H.H.; Lee, J.S.; Lin, C.H. Polystyrene microplastic particles: In vitro pulmonary toxicity assessment. J. Hazard. Mater. 2020, 385, 121575. [Google Scholar] [CrossRef]
- Campen, M.; Nihart, A.; Garcia, M.; Liu, R.; Olewine, M.; Castillo, E.; Bleske, B.; Scott, J.; Howard, T.; Gonzalez-Estrella, J.; et al. Bioaccumulation of Microplastics in Decedent Human Brains Assessed by Pyrolysis Gas Chromatography-Mass Spectrometry. Res. Sq. 2024, 6, rs.3.rs-4345687. [Google Scholar] [CrossRef]
- Tang, K.H.D.; Li, R. Aged Microplastics and Antibiotic Resistance Genes: A Review of Aging Effects on Their Interactions. Antibiotics 2024, 13, 941. [Google Scholar] [CrossRef]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.J. Microplastics in aquatic environments: Occurrence, accumulation, and biological effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef]
- Trushina, E.N.; Riger, N.A.; Mustafina, O.K.; Timonin, A.N. [Effect of microand nanoplastics as food contaminants on the immune system]. Vopr. Pitan. 2023, 92, 6–15. [Google Scholar] [CrossRef]
- Gautam, R.; Jo, J.; Acharya, M.; Maharjan, A.; Lee, D.; K, C.P.; Kim, C.; Kim, K.; Kim, H.; Heo, Y. Evaluation of potential toxicity of polyethylene microplastics on human derived cell lines. Sci. Total Environ. 2022, 838, 156089. [Google Scholar] [CrossRef]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, Z.; Wang, Z.; Liu, K.; Huang, S.; Liang, J.; Dai, Z.; Guo, W.; Mao, C.; Chen, S.; et al. Polyethylene microplastics promote nucleus pulposus cell senescence by inducing oxidative stress via TLR4/NOX2 axis. Ecotoxicol. Environ. Saf. 2025, 292, 117950. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yang, Y.; Lu, C.; Cheng, Y.; Jiang, X.; Yang, B.; Li, Y.; Chen, Q.; Ao, L.; Cao, J.; et al. Polystyrene nanoplastics exposure triggers spermatogenic cell senescence via the Sirt1/ROS axis. Ecotoxicol. Environ. Saf. 2024, 279, 116461. [Google Scholar] [CrossRef]
- Kang, Y.; Yin, S.; Zhou, X.; Liu, J.; Tan, X.; Zhang, C.; Lai, S.; Shao, L. Intranasal Zinc Oxide Nanoparticles Induce Neuronal PANoptosis via Microglial Pathway. Small 2025, 21, e2408139. [Google Scholar] [CrossRef]
- Kuntic, M.; Kuntic, I.; Cleppien, D.; Pozzer, A.; Nußbaum, D.; Oelze, M.; Junglas, T.; Strohm, L.; Ubbens, H.; Daub, S.; et al. Differential inflammation, oxidative stress and cardiovascular damage markers of nano- and micro-particle exposure in mice: Implications for human disease burden. Redox Biol. 2025, 83, 103644. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Y.; Liu, T.; Chen, M.; Qian, W.P.; Jiang, B.; Barwick, B.G.; Zhang, L.; Styblo, T.M.; Li, X.; et al. Inhibition of NADPH Oxidase-ROS Signal using Hyaluronic Acid Nanoparticles for Overcoming Radioresistance in Cancer Therapy. ACS Nano 2022, 16, 18708–18728. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, T.; Chen, M.; Gao, S.; Staley, C.A.; Yang, L.; Zhu, L. Dual inhibition of oxidative phosphorylation and glycolysis using a hyaluronic acid nanoparticle NOX inhibitor enhanced response to radiotherapy in colorectal cancer. Biomaterials 2025, 323, 123437. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, H.; Wang, C.; Su, X.L.; Song, Y.; Wu, P.; Yang, Z.; Wong, M.H.; Cai, Z.; Zheng, C. Metabolomics Reveal Nanoplastic-Induced Mitochondrial Damage in Human Liver and Lung Cells. Environ. Sci. Technol. 2022, 56, 12483–12493. [Google Scholar] [CrossRef]
- Das, A. The emerging role of microplastics in systemic toxicity: Involvement of reactive oxygen species (ROS). Sci. Total Environ. 2023, 895, 165076. [Google Scholar] [CrossRef]
- Zhang, D.; Ji, L.; Yang, Y.; Weng, J.; Ma, Y.; Liu, L.; Ma, W. Ceria Nanoparticle Systems Alleviate Degenerative Changes in Mouse Postovulatory Aging Oocytes by Reducing Oxidative Stress and Improving Mitochondrial Functions. ACS Nano 2024, 18, 13618–13634. [Google Scholar] [CrossRef]
- Gu, Y.; Xu, D.; Liu, J.; Chen, Y.; Wang, J.; Song, Y.; Sun, B.; Xia, B. Bioaccumulation of functionalized polystyrene nanoplastics in sea cucumber Apostichopus japonicus (Selenka, 1867) and their toxic effects on oxidative stress, energy metabolism and mitochondrial pathway. Environ. Pollut. 2023, 319, 121015. [Google Scholar] [CrossRef]
- Rong, J.; Yuan, C.; Yin, X.; Wu, X.; He, F.; Wang, Y.; Leung, K.S.; Lin, S. Co-exposure of polystyrene nanoplastics and copper induces development toxicity and intestinal mitochondrial dysfunction in vivo and in vitro. Sci. Total Environ. 2024, 930, 172681. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.H.; Seo, H.J.; Lee, J.Y.; Lee, I.; Jeon, K.; Kim, B.; Lee, K. Polypropylene nanoplastic exposure leads to lung inflammation through p38-mediated NF-κB pathway due to mitochondrial damage. Part. Fibre Toxicol. 2023, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Fang, Z.; Wang, P.; Qian, Z.; Yang, Y.; Ran, L.; Zheng, J.; Tang, Y.; Cui, X.; Li, Y.Y.; et al. Polylactic Acid Micro/Nanoplastic Exposure Induces Male Reproductive Toxicity by Disrupting Spermatogenesis and Mitochondrial Dysfunction in Mice. ACS Nano 2025, 19, 5589–5603. [Google Scholar] [CrossRef]
- Halimu, G.; Zhang, Q.; Liu, L.; Zhang, Z.; Wang, X.; Gu, W.; Zhang, B.; Dai, Y.; Zhang, H.; Zhang, C.; et al. Toxic effects of nanoplastics with different sizes and surface charges on epithelial-to-mesenchymal transition in A549 cells and the potential toxicological mechanism. J. Hazard. Mater. 2022, 430, 128485. [Google Scholar] [CrossRef]
- Lerchner, T.; Jedlička, J.; Kripnerová, M.; Dejmek, J.; Kuncová, J. Influence of micro- and nanoplastics on mitochondrial function in the cardiovascular system: A review of the current literature. Physiol. Res. 2024, 73, S685–S695. [Google Scholar] [CrossRef]
- Deng, Y.; Yan, Z.; Shen, R.; Wang, M.; Huang, Y.; Ren, H.; Zhang, Y.; Lemos, B. Microplastics release phthalate esters and cause aggravated adverse effects in the mouse gut. Environ. Int. 2020, 143, 105916. [Google Scholar] [CrossRef]
- Huang, Y.; Liang, B.; Li, Z.; Zhong, Y.; Wang, B.; Zhang, B.; Du, J.; Ye, R.; Xian, H.; Min, W.; et al. Polystyrene nanoplastic exposure induces excessive mitophagy by activating AMPK/ULK1 pathway in differentiated SH-SY5Y cells and dopaminergic neurons in vivo. Part. Fibre Toxicol. 2023, 20, 44. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, K.; Wang, D.; Wang, Y.; Lu, H.; Zhao, H.; Xing, M. Polystyrene microplastics-induced cardiotoxicity in chickens via the ROS-driven NF-κB-NLRP3-GSDMD and AMPK-PGC-1α axes. Sci. Total Environ. 2022, 840, 156727. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Lee, Y.H.; Hsu, Y.H.; Chiu, I.J.; Huang, C.C.; Huang, C.C.; Chia, Z.C.; Lee, C.P.; Lin, Y.F.; Chiu, H.W. The Kidney-Related Effects of Polystyrene Microplastics on Human Kidney Proximal Tubular Epithelial Cells HK-2 and Male C57BL/6 Mice. Environ. Health Perspect. 2021, 129, 57003. [Google Scholar] [CrossRef] [PubMed]
- Järvenpää, J.; Perkkiö, M.; Laitinen, R.; Lahtela-Kakkonen, M. PE and PET oligomers’ interplay with membrane bilayers. Sci. Rep. 2022, 12, 2234. [Google Scholar] [CrossRef]
- Tagorti, G.; Kaya, B. Genotoxic effect of microplastics and COVID-19: The hidden threat. Chemosphere 2022, 286, 131898. [Google Scholar] [CrossRef]
- Demir, E. Adverse biological effects of ingested polystyrene microplastics using Drosophila melanogaster as a model in vivo organism. J. Toxicol. Environ. Health A 2021, 84, 649–660. [Google Scholar] [CrossRef]
- Baj, J.; Dring, J.C.; Czeczelewski, M.; Kozyra, P.; Forma, A.; Flieger, J.; Kowalska, B.; Buszewicz, G.; Teresiński, G. Derivatives of Plastics as Potential Carcinogenic Factors: The Current State of Knowledge. Cancers 2022, 14, 4637. [Google Scholar] [CrossRef]
- Wang, K.; Du, Y.; Li, P.; Guan, C.; Zhou, M.; Wu, L.; Liu, Z.; Huang, Z. Nanoplastics causes heart aging/myocardial cell senescence through the Ca2+/mtDNA/cGAS-STING signaling cascade. J. Nanobiotechnol. 2024, 22, 96. [Google Scholar] [CrossRef]
- Li, X.; Huang, Y.; Zu, D.; Liu, H.; He, H.; Bao, Q.; He, Y.; Liang, C.; Luo, G.; Teng, Y.; et al. PMMA nanoplastics induce gastric epithelial cellular senescence and cGAS-STING-mediated inflammation via ROS overproduction and NHEJ suppression. Ecotoxicol. Environ. Saf. 2024, 287, 117284. [Google Scholar] [CrossRef]
- Kaur, M.; Xu, M.; Wang, L. Cyto-Genotoxic Effect Causing Potential of Polystyrene Micro-Plastics in Terrestrial Plants. Nanomaterials 2022, 12, 24. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Li, Y.; Yao, C.; Qu, J.; Tang, J.; Chen, G.; Han, Y. Acute exposure to polystyrene nanoplastics induces unfolded protein response and global protein ubiquitination in lungs of mice. Ecotoxicol. Environ. Saf. 2024, 280, 116580. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, C.; Liu, D.; Wang, Y.; Qi, H.; Liu, X.; Zhang, Y.; Chen, H.; Zeng, Y.; Li, J. Polystyrene nanoplastics-induced lung apoptosis and ferroptosis via ROS-dependent endoplasmic reticulum stress. Sci. Total Environ. 2024, 912, 169260. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, Y.; Zhang, H.; Wang, Y.; Wu, C.; Huang, W. Malvidin induces hepatic stellate cell apoptosis via the endoplasmic reticulum stress pathway and mitochondrial pathway. Food Sci. Nutr. 2020, 8, 5095–5106. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Gu, T.; Zhong, T.; Xiao, Y.; Sun, Q.C. Endoplasmic reticulum stress in metabolic disorders: Opposite roles of phytochemicals and food contaminants. Curr. Opin. Food Sci. 2022, 48, 100913. [Google Scholar] [CrossRef]
- Pontecorvi, P.; Ceccarelli, S.; Cece, F.; Camero, S.; Lotti, L.V.; Niccolai, E.; Nannini, G.; Gerini, G.; Anastasiadou, E.; Scialis, E.S.; et al. Assessing the Impact of Polyethylene Nano/Microplastic Exposure on Human Vaginal Keratinocytes. Int. J. Mol. Sci. 2023, 24, 11379. [Google Scholar] [CrossRef]
- Banikazemi, Z.; Farshadi, M.; Rajabi, A.; Homayoonfal, M.; Sharifi, N.; Sharafati Chaleshtori, R. Nanoplastics: Focus on the role of microRNAs and long non-coding RNAs. Chemosphere 2022, 308, 136299. [Google Scholar] [CrossRef]
- Huang, Z.; Weng, Y.; Shen, Q.; Zhao, Y.; Luo, T.; Xiao, Y.; Yang, G.; Jin, Y. Nano- and micro-polystyrene plastics interfered the gut barrier function mediated by exosomal miRNAs in rats. Environ. Pollut. 2023, 335, 122275. [Google Scholar] [CrossRef]
- Yuan, L.; Lin, J.; Peng, Y.; Gao, R.; Sun, Q. Chlorantraniliprole induces adipogenesis in 3T3-L1 adipocytes via the AMPKα pathway but not the ER stress pathway. Food Chem. 2020, 311, 125953. [Google Scholar] [CrossRef]
- Xu, Z.; Shen, J.; Lin, L.; Chen, J.; Wang, L.; Deng, X.; Wu, X.; Lin, Z.; Zhang, Y.; Yu, R.; et al. Exposure to irregular microplastic shed from baby bottles activates the ROS/NLRP3/Caspase-1 signaling pathway, causing intestinal inflammation. Environ. Int. 2023, 181, 108296. [Google Scholar] [CrossRef]
- Tang, Y.; Han, Y.; Zhang, W.; Yu, Y.; Huang, L.; Zhou, W.; Shi, W.; Tian, D.; Liu, G. Bisphenol A and microplastics weaken the antimicrobial ability of blood clams by disrupting humoral immune responses and suppressing hemocyte chemotactic activity. Environ. Pollut. 2022, 307, 119497. [Google Scholar] [CrossRef]
- Yang, S.; Cheng, Y.; Chen, Z.; Liu, T.; Yin, L.; Pu, Y.; Liang, G. In vitro evaluation of nanoplastics using human lung epithelial cells, microarray analysis and co-culture model. Ecotoxicol. Environ. Saf. 2021, 226, 112837. [Google Scholar] [CrossRef]
- Ma, J.Y.; Islam, F.; Ayyaz, A.; Fang, R.Y.; Hannan, F.; Farooq, M.A.; Ali, B.; Huang, Q.; Sun, R.; Zhou, W.J. Wood vinegar induces salinity tolerance by alleviating oxidative damages and protecting photosystem II in rapeseed cultivars. Ind. Crops Prod. 2022, 189, 115763. [Google Scholar] [CrossRef]
- Wang, S.; Han, Q.; Wei, Z.; Wang, Y.; Xie, J.; Chen, M. Polystyrene microplastics affect learning and memory in mice by inducing oxidative stress and decreasing the level of acetylcholine. Food Chem. Toxicol. 2022, 162, 112904. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Li, D.; Yuan, X.; Wang, Z.; Shao, Z.; Feng, X.; Yang, C.; Liu, H.; Zhang, G.; Wang, Y.; et al. Potential Effects of Orally Ingesting Polyethylene Terephthalate Microplastics on the Mouse Heart. Cardiovasc. Toxicol. 2024, 24, 291–301. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuan, Q.; Zhang, L.; Meng, L.; Fu, X.; Hou, Y. Polystyrene microplastics induced female reproductive toxicity in mice. J. Hazard. Mater. 2022, 424, 127629. [Google Scholar] [CrossRef]
- Luo, D.; Chu, X.; Wu, Y.; Wang, Z.; Liao, Z.; Ji, X.; Ju, J.; Yang, B.; Chen, Z.; Dahlgren, R.; et al. Micro- and nano-plastics in the atmosphere: A review of occurrence, properties and human health risks. J. Hazard. Mater. 2024, 465, 133412. [Google Scholar] [CrossRef]
- Santos, A.L.; Rodrigues, L.C.; Rodrigues, C.C.; Cirqueira, F.; Malafaia, G.; Rocha, T.L. Polystyrene nanoplastics induce developmental impairments and vasotoxicity in zebrafish (Danio rerio). J. Hazard. Mater. 2024, 464, 132880. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Yu, H.; Ye, L.; Li, T.; Zhang, X.; Wang, C.; Li, P.; Ji, H.; Gao, Q.; et al. Nanoplastics promote arsenic-induced ROS accumulation, mitochondrial damage and disturbances in neurotransmitter metabolism of zebrafish (Danio rerio). Sci. Total Environ. 2023, 863, 161005. [Google Scholar] [CrossRef]
- Leslie, H.A.; van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and quantification of plastic particle pollution in human blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, F.; Jiang, Z.; Du, Z.; Liu, S.; Zhang, M.; Jin, Y.; Qin, Y.; Yang, X.; Wang, C.; et al. Microplastics are associated with elevated atherosclerotic risk and increased vascular complexity in acute coronary syndrome patients. Part. Fibre Toxicol. 2024, 21, 34. [Google Scholar] [CrossRef]
- Lv, W.; Gu, H.; He, D.; Liu, Z.; Yao, C.; Huang, W.; Yuan, Q.; Zhou, W. Polystyrene nanospheres-induced hepatotoxicity in swamp eel (Monopterus albus): From biochemical, pathological and transcriptomic perspectives. Sci. Total Environ. 2023, 893, 164844. [Google Scholar] [CrossRef]
- Cao, H.; Ding, P.; Li, X.; Huang, C.; Li, X.; Chen, X.; Zhang, L.; Qi, J. Environmentally persistent free radicals on photoaged microplastics from disposable plastic cups induce the oxidative stress-associated toxicity. J. Hazard. Mater. 2024, 464, 132990. [Google Scholar] [CrossRef] [PubMed]
- Jeyavani, J.; Vaseeharan, B. Combined toxic effects of environmental predominant microplastics and ZnO nanoparticles in freshwater snail Pomaceae paludosa. Environ. Pollut. 2023, 325, 121427. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zhang, J.; Feng, Y.; Duan, Y.; Ma, H.; Zhang, H. Purification and identification of novel antioxidant peptides from watermelon seed protein hydrolysates and their cytoprotective effects on H2O2-induced oxidative stress. Food Chem. 2020, 327, 127059. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, J.; Dong, X.; Tang, J.; Xie, Y.; Yang, J.; Zou, L.; Wu, L.; Fan, J. Bifidobacterium longumBL-19 inhibits oxidative stress and inflammatory damage in the liver of mice with NAFLD by regulating the production of butyrate in the intestine. Food Sci. Nutr. 2024, 12, 6442–6460. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, C.; Ma, Z.; Zeng, L.; Wang, H.; Cheng, X.; Zhang, C.; Xue, Y.; Yuan, Y.; Li, J.; et al. Co-exposure to polystyrene nanoplastics and triclosan induces synergistic cytotoxicity in human KGN granulosa cells by promoting reactive oxygen species accumulation. Ecotoxicol. Environ. Saf. 2024, 273, 116121. [Google Scholar] [CrossRef]
- Dhanasekaran, S.; Yang, Q.; Godana, E.A.; Liu, J.; Li, J.; Zhang, H. Trehalose supplementation enhanced the biocontrol efficiency of Sporidiobolus pararoseus Y16 through increased oxidative stress tolerance and altered transcriptome. Pest. Manag. Sci. 2021, 77, 4425–4436. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Wu, H.; Chai, Z.; Hutabarat, R.P.; Zeng, Q.; Niu, L.; Li, D.; Yu, H.; Huang, W. Blueberry leaves from 73 different cultivars in southeastern China as nutraceutical supplements rich in antioxidants. Food Res. Int. 2019, 122, 548–560. [Google Scholar] [CrossRef]
- Chen, X. A review on coffee leaves: Phytochemicals, bioactivities and applications. Crit. Rev. Food Sci. Nutr. 2019, 59, 1008–1025. [Google Scholar] [CrossRef]
- Dai, Y.Q.; Wu, H.; Liu, X.L.; Liu, H.; Yin, L.Q.; Wang, Z.; Xia, X.D.; Zhou, J.Z. Antioxidant activities and inhibitory effects of blueberry pomace and wine pomace crude extracts on oxidation of oil in water emulsion and fish mince. J. Food Process. Preserv. 2021, 45, 15509. [Google Scholar] [CrossRef]
- Krinsky, N.I. Carotenoids as antioxidants. Nutrition 2001, 17, 815–817. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Puig, S.; Fenollosa, E.; Casadesús, A.; Fernández, E. Vitamin E protects from lipid peroxidation during winter stress in the seagrass Cymodocea nodosa. Planta 2022, 255, 41. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Role of vitamin E as a lipid-soluble peroxyl radical scavenger: In vitro and in vivo evidence. Free Radic. Biol. Med. 2014, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ishteyaque, S.; Yadav, K.S.; Verma, S.; Washimkar, K.R.; Mugale, M.N. CYP2E1 triggered GRP78/ATF6/CHOP signaling axis inhibit apoptosis and promotes progression of hepatocellular carcinoma. Arch. Biochem. Biophys. 2023, 745, 109701. [Google Scholar] [CrossRef]
- Chan, A.C. Partners in defense, vitamin E and vitamin C. Can. J. Physiol. Pharmacol. 1993, 71, 725–731. [Google Scholar] [CrossRef]
- Lee, G.Y.; Han, S.N. The Role of Vitamin E in Immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef]
- Schroeder, P.; Krutmann, J. In vivo relevance of infrared a radiation-induced skin damage: Reply to Piazena and Kelleher, Letter to the Editor, Free Radical Biology and Medicine, 2008. Free Radic. Biol. Med. 2008, 44, 1870–1871. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, J.; Zheng, C.; Zhang, P.; Zhou, W.; Cui, W.; Mo, X.; Li, L.; Xu, L.; Gao, J. Nrf2 antioxidant pathway suppresses Numb-mediated epithelial-mesenchymal transition during pulmonary fibrosis. Cell Death Dis. 2018, 9, 83. [Google Scholar] [CrossRef]
- Wei, K.; Wei, Y.; Xu, W.; Lu, F.; Ma, H. Corn peptides improved obesity-induced non-alcoholic fatty liver disease through relieving lipid metabolism, insulin resistance and oxidative stress. Food Funct. 2022, 13, 5782–5793. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Lin, X.; Bai, D.; Wei, Z.; Zhang, Y.; Huang, Y.; Deng, H.; Huang, X. Curcumin attenuates oxidative stress in RAW264.7 cells by increasing the activity of antioxidant enzymes and activating the Nrf2-Keap1 pathway. PLoS ONE 2019, 14, e0216711. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Amador-Martínez, I.; Medina-Campos, O.N.; Garcia-Garcia, M.; Bernabe-Yepes, B.; León-Contreras, J.C.; Hernández-Pando, R.; Aparicio-Trejo, O.E.; Sánchez-Lozada, L.G.; et al. Sulforaphane protects from kidney damage during the release of unilateral ureteral obstruction (RUUO) by activating nuclear factor erythroid 2-related factor 2 (Nrf2): Role of antioxidant, anti-inflammatory, and antiapoptotic mechanisms. Free Radic. Biol. Med. 2024, 212, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Kubben, N.; Zhang, W.; Wang, L.; Voss, T.C.; Yang, J.; Qu, J.; Liu, G.H.; Misteli, T. Repression of the Antioxidant NRF2 Pathway in Premature Aging. Cell 2016, 165, 1361–1374. [Google Scholar] [CrossRef]
- Lee, J.M.; Li, J.; Johnson, D.A.; Stein, T.D.; Kraft, A.D.; Calkins, M.J.; Jakel, R.J.; Johnson, J.A. Nrf2, a multi-organ protector? FASEB J. 2005, 19, 1061–1066. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, Y.; Xu, C.; An, P.; Luo, Y.; Jiao, L.; Luo, J.; Li, Y. Mitochondrial Dysfunction and Therapeutic Perspectives in Cardiovascular Diseases. Int. J. Mol. Sci. 2022, 23, 53. [Google Scholar] [CrossRef]
- Wang, H.H.; Sun, Y.N.; Qu, T.Q.; Sang, X.Q.; Zhou, L.M.; Li, Y.X.; Ren, F.Z. Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function. Int. J. Mol. Sci. 2022, 23, 11963. [Google Scholar] [CrossRef]
- Lone, J.; Parray, H.A.; Yun, J.W. Nobiletin induces brown adipocyte-like phenotype and ameliorates stress in 3T3-L1 adipocytes. Biochimie 2018, 146, 97–104. [Google Scholar] [CrossRef]
- Li, H.; Weng, Q.; Gong, S.; Zhang, W.; Wang, J.; Huang, Y.; Li, Y.; Guo, J.; Lan, T. Kaempferol prevents acetaminophen-induced liver injury by suppressing hepatocyte ferroptosis via Nrf2 pathway activation. Food Funct. 2023, 14, 1884–1896. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, W.; Zheng, Z.; Wang, W.; Yuan, Y.; Hong, Q.; Lin, J.; Li, X.; Meng, Y. Cigarette smoke-inactivated SIRT1 promotes autophagy-dependent senescence of alveolar epithelial type 2 cells to induce pulmonary fibrosis. Free Radic. Biol. Med. 2021, 166, 116–127. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol Treats UVB-Induced Photoaging by Anti-MMP Expression, through Anti-Inflammatory, Antioxidant, and Antiapoptotic Properties, and Treats Photoaging by Upregulating VEGF-B Expression. Oxid. Med. Cell Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-inflammatory action of astaxanthin and its use in the treatment of various diseases. Biomed. Pharmacother. 2022, 145, 112179. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Li, F.; Zhu, X.; Xu, J.; Muhammad, A.; Chen, Y.; Li, D.; Liu, B.; Wang, C.; Wang, Z.; et al. Alfalfa saponins inhibit oxidative stress-induced cell apoptosis through the MAPK signaling pathway. Redox Rep. 2022, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J. Surg. Oncol. 2018, 16, 108. [Google Scholar] [CrossRef]
- Gao, R.; Zhu, L.; Zhang, W.; Jin, W.; Bai, F.; Xu, P.; Wang, J.; Sun, Q.; Guo, Z.; Yuan, L. Novel Peptides from Sturgeon Ovarian Protein Hydrolysates Prevent Oxidative Stress-Induced Dysfunction in Osteoblast Cells: Purification, Identification, and Characterization. J. Agric. Food Chem. 2024, 72, 10076–10088. [Google Scholar] [CrossRef]
- Guo, L.T.; Wang, S.Q.; Su, J.; Xu, L.X.; Ji, Z.Y.; Zhang, R.Y.; Zhao, Q.W.; Ma, Z.Q.; Deng, X.Y.; Ma, S.P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflam. 2019, 16, 95. [Google Scholar] [CrossRef]
- Li, L.; Jiang, W.; Yu, B.; Liang, H.; Mao, S.; Hu, X.; Feng, Y.; Xu, J.; Chu, L. Quercetin improves cerebral ischemia/reperfusion injury by promoting microglia/macrophages M2 polarization via regulating PI3K/Akt/NF-κB signaling pathway. Biomed. Pharmacother. 2023, 168, 115653. [Google Scholar] [CrossRef]
- Switzer, C.H.; Kasamatsu, S.; Ihara, H.; Eaton, P. SOD1 is an essential H(2)S detoxifying enzyme. Proc. Natl. Acad. Sci. USA 2023, 120, e2205044120. [Google Scholar] [CrossRef]
- Panda, C.; Komarnytsky, S.; Fleming, M.N.; Marsh, C.; Barron, K.; Le Brun-Blashka, S.; Metzger, B. Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants. Nutrients 2023, 15, 2209. [Google Scholar] [CrossRef]
- Hayes, J.D.; Pulford, D.J. The glutathione S-transferase supergene family: Regulation of GST and the contribution of the isoenzymes to cancer chemoprotection and drug resistance. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–600. [Google Scholar] [CrossRef]
- Ahmed, F.; Ijaz, B.; Ahmad, Z.; Farooq, N.; Sarwar, M.B.; Husnain, T. Modification of miRNA Expression through plant extracts and compounds against breast cancer: Mechanism and translational significance. Phytomedicine 2020, 68, 153168. [Google Scholar] [CrossRef]
- Ahmad, N.; Naeem, M.; Ali, H.; Alabbosh, K.F.; Hussain, H.; Khan, I.; Siddiqui, S.A.; Khan, A.A.; Iqbal, B. From challenges to solutions: The impact of melatonin on abiotic stress synergies in horticultural plants via redox regulation and epigenetic signaling. Sci. Hortic. 2023, 321, 112369. [Google Scholar] [CrossRef]
- Vezza, T.; de Marañón, A.M.; Canet, F.; Díaz-Pozo, P.; Marti, M.; D’Ocon, P.; Apostolova, N.; Rocha, M.; Víctor, V.M. MicroRNAs and Oxidative Stress: An Intriguing Crosstalk to Be Exploited in the Management of Type 2 Diabetes. Antioxidants 2021, 10, 802. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Cao, Y.; Chai, X.; Zhao, Q.; Geng, Y.; Liu, D.; Tian, S. Potential health risks of the interaction of microplastics and lung surfactant. J. Hazard. Mater. 2022, 429, 128109. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yoon, S.J.; Jung, N.Y.; Lee, W.S.; Jeong, J.; Park, Y.J.; Kim, W.; Oh, D.B.; Seo, J. Antioxidants prevent particulate matter-induced senescence of lung fibroblasts. Heliyon 2023, 9, e14179. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, J.; He, Q.; Wang, M.; Liu, Q.; Wang, T.; Chen, X.; Wang, W.; Xu, H. Association between dietary vitamin C and abdominal aortic calcification among the US adults. Nutr. J. 2023, 22, 58. [Google Scholar] [CrossRef]
- Ji, D.; Wang, Q.; Lu, T.; Ma, H.; Chen, X. The effects of ultrasonication on the phytochemicals, antioxidant, and polyphenol oxidase and peroxidase activities in coffee leaves. Food Chem. 2022, 373, 131480. [Google Scholar] [CrossRef]
- Sun, W.; Chen, Y.; Wang, L.; Wang, Z.; Liu, S.; Zhang, M.; Liu, Y.; Li, Q.; Zhang, H. Gram-scale preparation of quercetin supramolecular nanoribbons for intestinal inflammatory diseases by oral administration. Biomaterials 2023, 295, 122039. [Google Scholar] [CrossRef]
- Wen, S.Y.; Wei, B.Y.; Ma, J.Q.; Wang, L.; Chen, Y.Y. Phytochemicals, Biological Activities, Molecular Mechanisms, and Future Prospects of Plantago asiatica L. J. Agric. Food Chem. 2023, 71, 143–173. [Google Scholar] [CrossRef]
- Gu, T.Y.; Wang, N.M.; Wu, T.; Ge, Q.; Chen, L. Antioxidative Stress Mechanisms behind Resveratrol: A Multidimensional Analysis. J. Food Qual. 2021, 2021, 5571733. [Google Scholar] [CrossRef]
- Bhattarai, G.; Poudel, S.B.; Kook, S.H.; Lee, J.C. Resveratrol prevents alveolar bone loss in an experimental rat model of periodontitis. Acta Biomater. 2016, 29, 398–408. [Google Scholar] [CrossRef]
- Shahani, A.A.A.; Abbas, A.; Hameed, R.; Iqbal, A.; Chen, S.B.; Liu, Q.; Liu, Y.X.; Zhang, D.; Zhu, R.G.; Fayyaz, A.; et al. Melatonin in plants: A pleiotropic molecule for abiotic stresses and pathogens infection. Sci. Hortic. 2023, 322, 112387. [Google Scholar] [CrossRef]
- He, R.; Cui, M.; Lin, H.; Zhao, L.; Wang, J.; Chen, S.; Shao, Z. Melatonin resists oxidative stress-induced apoptosis in nucleus pulposus cells. Life Sci. 2018, 199, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Luo, S.F.; Huali, H.; Zhou, H.S.; Zhang, Y.T.; An, R.H.; Ling, J.; Li, P.X. Melatonin promotes the normal cellular mitochondrial function of lotus seeds through stimulating nitric oxide production. Postharvest Biol. Technol. 2022, 185, 111814. [Google Scholar] [CrossRef]
- Farag, A.A.; Bayoumi, H.; Radwaan, S.E.; El Gazzar, W.B.; Youssef, H.S.; Nasr, H.E.; Badr, A.M.; Mansour, H.M.; Elalfy, A.; Sayed, A.E.H.; et al. Melatonin counteracts polyethylene microplastics induced adreno-cortical damage in male albino rats. Ecotoxicol. Environ. Saf. 2024, 279, 116499. [Google Scholar] [CrossRef]
- Song, S.; van Dijk, F.; Vasse, G.F.; Liu, Q.; Gosselink, I.F.; Weltjens, E.; Remels, A.H.V.; de Jager, M.H.; Bos, S.; Li, C.; et al. Inhalable Textile Microplastic Fibers Impair Airway Epithelial Differentiation. Am. J. Respir. Crit. Care Med. 2024, 209, 427–443. [Google Scholar] [CrossRef]
- Wen, J.; Sun, H.; Yang, B.; Song, E.; Song, Y.; Jiang, G. Environmentally Relevant Concentrations of Microplastic Exposure Cause Cholestasis and Bile Acid Metabolism Dysregulation through a Gut-Liver Loop in Mice. Environ. Sci. Technol. 2024, 58, 1832–1841. [Google Scholar] [CrossRef]
- Bashir, D.W.; Ahmed, Y.H.; El-Sakhawy, M.A. Ameliorative effect of vitamin E and selenium against bisphenol A-induced toxicity in spinal cord and submandibular salivary glands of adult male albino rats. Int. J. Environ. Health Res. 2023, 33, 993–1009. [Google Scholar] [CrossRef]
- Sun, R.; Lin, Z.; Wang, X.; Liu, L.; Huo, M.; Zhang, R.; Lin, J.; Xiao, C.; Li, Y.; Zhu, W.; et al. AADAC protects colorectal cancer liver colonization from ferroptosis through SLC7A11-dependent inhibition of lipid peroxidation. J. Exp. Clin. Cancer Res. 2022, 41, 284. [Google Scholar] [CrossRef]
- Kumar, P.; Osahon, O.W.; Sekhar, R.V. GlyNAC (Glycine and N-Acetylcysteine) Supplementation in Mice Increases Length of Life by Correcting Glutathione Deficiency, Oxidative Stress, Mitochondrial Dysfunction, Abnormalities in Mitophagy and Nutrient Sensing, and Genomic Damage. Nutrients 2022, 14, 1114. [Google Scholar] [CrossRef]
- Alvarenga, L.; Saldanha, J.F.; Stockler-Pinto, M.B.; Fouque, D.; Soulage, C.O.; Mafra, D. Effects of resveratrol on inflammation and oxidative stress induced by the uremic toxin indoxyl sulfate in Murine macrophage-like RAW 264.7. Biochimie 2023, 213, 22–29. [Google Scholar] [CrossRef]
- Cong, L.; Lei, M.Y.; Liu, Z.Q.; Liu, Z.F.; Ma, Z.; Liu, K.; Li, J.; Deng, Y.; Liu, W.; Xu, B. Resveratrol attenuates manganese-induced oxidative stress and neuroinflammation through SIRT1 signaling in mice. Food Chem. Toxicol. 2021, 153, 112283. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Z.; Peng, P.; Gong, Z.; Huang, J.; Peng, H. Astaxanthin attenuates osteoarthritis progression via inhibiting ferroptosis and regulating mitochondrial function in chondrocytes. Chem. Biol. Interact. 2022, 366, 110148. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.N.; Wen, B.; Li, X.X.; Xu, L.; Gao, J.Z.; Chen, Z.Z. Astaxanthin mitigates oxidative stress caused by microplastics at the expense of reduced skin pigmentation in discus fish. Sci. Total Environ. 2023, 874, 162494. [Google Scholar] [CrossRef]
- Geng, Q.; Wang, S.; Heng, K.; Zhai, J.; Song, X.; Xia, L.; Wang, L.; Lin, Q.; Li, H.; Guo, Y. Astaxanthin attenuates irradiation-induced osteoporosis in mice by inhibiting oxidative stress, osteocyte senescence, and SASP. Food Funct. 2022, 13, 11770–11779. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Qu, H.; Cai, C.; Liu, H.; Chu, J. β-Carotene extracted from Blakeslea trispora attenuates oxidative stress, inflammatory, hepatic injury and immune damage induced by copper sulfate in zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 258, 109366. [Google Scholar] [CrossRef]
- Rocha, F.; Yumi Sugahara, L.; Leimann, F.V.; de Oliveira, S.M.; da Silva Brum, E.; Calhelha, R.C.; Barreiro, M.F.; Ferreira, I.; Porto Ineu, R.; Gonçalves, O.H. Nanodispersions of beta-carotene: Effects on antioxidant enzymes and cytotoxic properties. Food Funct. 2018, 9, 3698–3706. [Google Scholar] [CrossRef]
- Kashif, M.; Yao, H.; Schmidt, S.; Chen, X.; Truong, M.; Tüksammel, E.; Liu, Y.; Bergo, M.O. ROS-lowering doses of vitamins C and A accelerate malignant melanoma metastasis. Redox Biol. 2023, 60, 102619. [Google Scholar] [CrossRef]
- Izumiya, M.; Haniu, M.; Ueda, K.; Ishida, H.; Ma, C.; Ideta, H.; Sobajima, A.; Ueshiba, K.; Uemura, T.; Saito, N.; et al. Evaluation of MC3T3-E1 Cell Osteogenesis in Different Cell Culture Media. Int. J. Mol. Sci. 2021, 22, 7752. [Google Scholar] [CrossRef]
- Nicoliche, T.; Bartolomeo, C.S.; Lemes, R.M.R.; Pereira, G.C.; Nunes, T.A.; Oliveira, R.B.; Nicastro, A.L.M.; Soares, É.N.; da Cunha Lima, B.F.; Rodrigues, B.M.; et al. Antiviral, anti-inflammatory and antioxidant effects of curcumin and curcuminoids in SH-SY5Y cells infected by SARS-CoV-2. Sci. Rep. 2024, 14, 10696. [Google Scholar] [CrossRef]
- Ren, B.C.; Zhang, Y.F.; Liu, S.S.; Cheng, X.J.; Yang, X.; Cui, X.G.; Zhao, X.R.; Zhao, H.; Hao, M.F.; Li, M.D.; et al. Curcumin alleviates oxidative stress and inhibits apoptosis in diabetic cardiomyopathy via Sirt1-Foxo1 and PI3K-Akt signalling pathways. J. Cell Mol. Med. 2020, 24, 12355–12367. [Google Scholar] [CrossRef]
- Sharma, S.; Kulkarni, S.K.; Chopra, K. Curcumin, the active principle of turmeric (Curcuma longa), ameliorates diabetic nephropathy in rats. Clin. Exp. Pharmacol. Physiol. 2006, 33, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Li, R.; Li, Y.; Wang, M.; Wang, L.; Wang, S.; Cheng, H.; Zhang, Q.; Fu, C.; Qian, Z.; et al. Inflammation-Targeted Nanomedicines Alleviate Oxidative Stress and Reprogram Macrophages Polarization for Myocardial Infarction Treatment. Adv. Sci. 2024, 11, e2308910. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, X.; Tang, Y.; Huang, C. Melatonin alleviates doxorubicin-induced cardiotoxicity via inhibiting oxidative stress, pyroptosis and apoptosis by activating Sirt1/Nrf2 pathway. Biomed. Pharmacother. 2023, 162, 114591. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.P.; Sri Hari, A.; Day, B.J.; Patel, M. Pharmacological elevation of glutathione inhibits status epilepticus-induced neuroinflammation and oxidative injury. Redox Biol. 2024, 73, 103168. [Google Scholar] [CrossRef]
- Luo, L.; Huang, F.; Zhong, S.; Ding, R.; Su, J.; Li, X. Astaxanthin attenuates ferroptosis via Keap1-Nrf2/HO-1 signaling pathways in LPS-induced acute lung injury. Life Sci. 2022, 311, 121091. [Google Scholar] [CrossRef]
- Cai, X.; Hua, S.; Deng, J.; Du, Z.; Zhang, D.; Liu, Z.; Khan, N.U.; Zhou, M.; Chen, Z. Astaxanthin Activated the Nrf2/HO-1 Pathway to Enhance Autophagy and Inhibit Ferroptosis, Ameliorating Acetaminophen-Induced Liver Injury. ACS Appl. Mater. Interfaces 2022, 14, 42887–42903. [Google Scholar] [CrossRef]
- Wu, L.; Mo, W.; Feng, J.; Li, J.; Yu, Q.; Li, S.; Zhang, J.; Chen, K.; Ji, J.; Dai, W.; et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br. J. Pharmacol. 2020, 177, 3760–3777. [Google Scholar] [CrossRef]
- Meng, M.; Jiang, Y.; Wang, Y.; Huo, R.; Ma, N.; Shen, X.; Chang, G. β-carotene targets IP3R/GRP75/VDAC1-MCU axis to renovate LPS-induced mitochondrial oxidative damage by regulating STIM1. Free Radic. Biol. Med. 2023, 205, 25–46. [Google Scholar] [CrossRef]
- Latief, U.; Ahmad, R. β-Carotene inhibits NF-κB and restrains diethylnitrosamine-induced hepatic inflammation in Wistar rats. Int. J. Vitam. Nutr. Res. 2022, 92, 301–310. [Google Scholar] [CrossRef]
- Han, X.; Luo, R.; Qi, S.; Wang, Y.; Dai, L.; Nie, W.; Lin, M.; He, H.; Ye, N.; Fu, C.; et al. “Dual sensitive supramolecular curcumin nanoparticles” in “advanced yeast particles” mediate macrophage reprogramming, ROS scavenging and inflammation resolution for ulcerative colitis treatment. J. Nanobiotechnol. 2023, 21, 321. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, P.; Li, H.; Wang, Y.; Huang, Y.; Fan, L.; Ma, X.; Qian, X.; Xi, J. Cerium-Luteolin Nanocomplexes in Managing Inflammation-Related Diseases by Antioxidant and Immunoregulation. ACS Nano 2024, 18, 6229–6242. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.Y.; Kim, M.Y.; Cho, J.Y. Antioxidant, Anti-Inflammatory, Anti-Menopausal, and Anti-Cancer Effects of Lignans and Their Metabolites. Int. J. Mol. Sci. 2022, 23, 15482. [Google Scholar] [CrossRef]
- Chen, S.M.; Zhou, J.; Igbokwe, C.J.; Duan, Y.Q.; Cai, M.H.; He, Y.Q.; Zhang, H.H. Oligopeptide of RDPEER from watermelon seeds prevents heat stress-induced liver injury by suppressing oxidative stress and inflammation responses. J. Funct. Foods 2023, 105, 105563. [Google Scholar] [CrossRef]
- Meng, M.; Huo, R.; Wang, Y.; Ma, N.; Shi, X.; Shen, X.; Chang, G. Lentinan inhibits oxidative stress and alleviates LPS-induced inflammation and apoptosis of BMECs by activating the Nrf2 signaling pathway. Int. J. Biol. Macromol. 2022, 222, 2375–2391. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xiao, M.; Eweys, A.S.; Bai, J.; Darwesh, O.M.; Xiao, X. Cinnamaldehyde Alleviates the Oxidative Stress of Caenorhabditis elegans in the Presence of Lactic Acid. Plant Foods Hum. Nutr. 2023, 78, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.; Chen, X.; Jia, G.; Chen, G.; Gong, L.; Cheng, Y.; Li, Z.; Wang, H.; Chen, A.; Zhang, G.; et al. WGX50 mitigates doxorubicin-induced cardiotoxicity through inhibition of mitochondrial ROS and ferroptosis. J. Transl. Med. 2023, 21, 823. [Google Scholar] [CrossRef]
- Zhang, J.; He, L.; Wang, A.; Wu, B.; Zhang, P.; Zhu, Y.; Jiang, Y.; Bai, J.; Xiao, X. Responses of bitter melon saponins to oxidative stress and aging via the IIS pathway linked with sir-2.1 and hlh-30. J. Food Biochem. 2022, 46, e14456. [Google Scholar] [CrossRef]
- Chan, H.S.H.; Dingle, C.; Not, C. Evidence for non-selective ingestion of microplastic in demersal fish. Mar. Pollut. Bull. 2019, 149, 110523. [Google Scholar] [CrossRef]
- Prata, J.C. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef]
- Shahsavaripour, M.; Abbasi, S.; Mirzaee, M.; Amiri, H. Human occupational exposure to microplastics: A cross-sectional study in a plastic products manufacturing plant. Sci. Total Environ. 2023, 882, 163576. [Google Scholar] [CrossRef]
- Prado, C.M.; Batsis, J.A.; Donini, L.M.; Gonzalez, M.C.; Siervo, M. Sarcopenic obesity in older adults: A clinical overview. Nat. Rev. Endocrinol. 2024, 20, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Cuffaro, F.; Lamminpää, I.; Niccolai, E.; Amedei, A. Nutritional and Microbiota-Based Approaches in Amyotrophic Lateral Sclerosis: From Prevention to Treatment. Nutrients 2024, 17, 102. [Google Scholar] [CrossRef]
- Heidari, H.; Bagherniya, M.; Majeed, M.; Sathyapalan, T.; Jamialahmadi, T.; Sahebkar, A. Curcumin-piperine co-supplementation and human health: A comprehensive review of preclinical and clinical studies. Phytother. Res. 2023, 37, 1462–1487. [Google Scholar] [CrossRef] [PubMed]
- Łysakowska, P.; Sobota, A.; Wirkijowska, A. Medicinal Mushrooms: Their Bioactive Components, Nutritional Value and Application in Functional Food Production—A Review. Molecules 2023, 28, 5393. [Google Scholar] [CrossRef]
- Zhang, D.; Ivane, N.M.A.; Haruna, S.A.; Zekrumah, M.; Elysé, F.K.R.; Tahir, H.E.; Wang, G.; Wang, C.; Zou, X. Recent trends in the micro-encapsulation of plant-derived compounds and their specific application in meat as antioxidants and antimicrobials. Meat Sci. 2022, 191, 108842. [Google Scholar] [CrossRef]
- Mamy, D.; Boateng, I.D.; Chen, X.M. Two-pot optimization of nutrient sources to enhance antioxidants in Citrus reticulata peel powder through solid-state fermentation with Aspergillus niger CGMCC 3.6189. Food Biosci. 2024, 59, 104145. [Google Scholar] [CrossRef]
- Di Pietro, V.; Yakoub, K.M.; Caruso, G.; Lazzarino, G.; Signoretti, S.; Barbey, A.K.; Tavazzi, B.; Lazzarino, G.; Belli, A.; Amorini, A.M. Antioxidant Therapies in Traumatic Brain Injury. Antioxidants 2020, 9, 260. [Google Scholar] [CrossRef]
- Hall, E.D.; Wang, J.A.; Miller, D.M.; Cebak, J.E.; Hill, R.L. Newer pharmacological approaches for antioxidant neuroprotection in traumatic brain injury. Neuropharmacology 2019, 145, 247–258. [Google Scholar] [CrossRef]
- Keshavarz Shahbaz, S.; Koushki, K.; Izadi, O.; Penson, P.E.; Sukhorukov, V.N.; Kesharwani, P.; Sahebkar, A. Advancements in curcumin-loaded PLGA nanoparticle delivery systems: Progressive strategies in cancer therapy. J. Drug Target. 2024, 32, 1207–1232. [Google Scholar] [CrossRef]
- Lin, Y.; McClements, D.J.; Xiao, J.; Cao, Y.; Liu, X. In Vitro-In Vivo Study of the Impact of Excipient Emulsions on the Bioavailability and Antioxidant Activity of Flavonoids: Influence of the Carrier Oil Type. J. Agric. Food Chem. 2023, 71, 1488–1498. [Google Scholar] [CrossRef]
- Arslan, M.; Zou, X.B.; Tahir, H.E.; Hu, X.T.; Rakha, A.; Zareef, M.; Seweh, E.A.; Basheer, S. NIR Spectroscopy Coupled Chemometric Algorithms for Rapid Antioxidants Activity Assessment of Chinese Dates (Zizyphus jujuba Mill.). Int. J. Food Eng. 2019, 15. [Google Scholar] [CrossRef]
- Yang, R.; Mu, W.Y.; Chen, Q.Y. Urazole-Au Nanocluster as a Novel Fluorescence Probe for Curcumin Determination and Mitochondria Imaging. Food Anal. Methods 2019, 12, 1805–1812. [Google Scholar] [CrossRef]
- Santos, D.; Luzio, A.; Félix, L.; Bellas, J.; Monteiro, S.M. Oxidative stress, apoptosis and serotonergic system changes in zebrafish (Danio rerio) gills after long-term exposure to microplastics and copper. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 258, 109363. [Google Scholar] [CrossRef]
- Khan, I.; Tariq, M.; Alabbosh, K.F.; Rehman, A.; Jalal, A.; Khan, A.A.; Farooq, M.; Li, G.L.; Iqbal, B.; Ahmad, N.; et al. Soil microplastics: Impacts on greenhouse gasses emissions, carbon cycling, microbial diversity, and soil characteristics. Appl. Soil Ecol. 2024, 197, 105343. [Google Scholar] [CrossRef]
- Luo, H.; Tu, C.; He, D.; Zhang, A.; Sun, J.; Li, J.; Xu, J.; Pan, X. Interactions between microplastics and contaminants: A review focusing on the effect of aging process. Sci. Total Environ. 2023, 899, 165615. [Google Scholar] [CrossRef]
- Rondanelli, M.; Faliva, M.A.; Miccono, A.; Naso, M.; Nichetti, M.; Riva, A.; Guerriero, F.; De Gregori, M.; Peroni, G.; Perna, S. Food pyramid for subjects with chronic pain: Foods and dietary constituents as anti-inflammatory and antioxidant agents. Nutr. Res. Rev. 2018, 31, 131–151. [Google Scholar] [CrossRef]
- Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Arista-Ugalde, T.L.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. The Biological Significance of Oxidative Stress, Effects of Fruits as Natural Edible Antioxidants. Curr. Pharm. Des. 2018, 24, 4807–4824. [Google Scholar] [CrossRef]
- Mani, S.; Dubey, R.; Lai, I.C.; Babu, M.A.; Tyagi, S.; Swargiary, G.; Mody, D.; Singh, M.; Agarwal, S.; Iqbal, D.; et al. Oxidative Stress and Natural Antioxidants: Back and Forth in the Neurological Mechanisms of Alzheimer’s Disease. J. Alzheimers Dis. 2023, 96, 877–912. [Google Scholar] [CrossRef]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Shariati, M.A.; Rebezov, M.; Rengasamy, K.R.R.; Mubarak, M.S. Superoxide dismutase: An updated review on its health benefits and industrial applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 7282–7300. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, B.M.; Kim, H.S. Potential protective roles of curcumin against cadmium-induced toxicity and oxidative stress. J. Toxicol. Environ. Health B Crit. Rev. 2021, 24, 95–118. [Google Scholar] [CrossRef]
- do Carmo, M.A.V.; Granato, D.; Azevedo, L. Antioxidant/pro-oxidant and antiproliferative activities of phenolic-rich foods and extracts: A cell-based point of view. Adv. Food Nutr. Res. 2021, 98, 253–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, W.; Li, H.; Cui, X.; Ma, J.; Wang, R.; Xu, Y.; Li, M.; Bai, X.; Wang, D.; et al. 4-octyl itaconate alleviates cisplatin-induced ferroptosis possibly via activating the NRF2/HO-1 signalling pathway. J. Cell Mol. Med. 2024, 28, e18207. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chu, T.; Yu, T.; Li, N.; Wang, C.; Li, C.; Zhang, Y.; Meng, H.; Nie, G. Design of Diselenide-Bridged Hyaluronic Acid Nano-antioxidant for Efficient ROS Scavenging to Relieve Colitis. ACS Nano 2022, 16, 13037–13048. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Nirumand, M.C.; Hajialyani, M.; Rahimi, R.; Farzaei, M.H.; Zingue, S.; Nabavi, S.M.; Bishayee, A. Dietary Plants for the Prevention and Management of Kidney Stones: Preclinical and Clinical Evidence and Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 765. [Google Scholar] [CrossRef]
- Cox, K.D.; Covernton, G.A.; Davies, H.L.; Dower, J.F.; Juanes, F.; Dudas, S.E. Human Consumption of Microplastics. Environ. Sci. Technol. 2019, 53, 7068–7074. [Google Scholar] [CrossRef]
- Gasmi, A.; Nasreen, A.; Lenchyk, L.; Lysiuk, R.; Peana, M.; Shapovalova, N.; Piscopo, S.; Komisarenko, M.; Shanaida, M.; Smetanina, K.; et al. An Update on Glutathione’s Biosynthesis, Metabolism, Functions, and Medicinal Purposes. Curr. Med. Chem. 2024, 31, 4579–4601. [Google Scholar] [CrossRef]
- Savage, R.A.; Zafar, N.; Yohannan, S.; Miller, J.M.M. Melatonin. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2025. [Google Scholar]
- Shao, K.; Zou, R.; Zhang, Z.; Mandemaker, L.D.B.; Timbie, S.; Smith, R.D.; Durkin, A.M.; Dusza, H.M.; Meirer, F.; Weckhuysen, B.M.; et al. Advancements in Assays for Micro- and Nanoplastic Detection: Paving the Way for Biomonitoring and Exposomics Studies. Annu. Rev. Pharmacol. Toxicol. 2025, 65, 567–585. [Google Scholar] [CrossRef]
- Hu, C.J.; Garcia, M.A.; Nihart, A.; Liu, R.; Yin, L.; Adolphi, N.; Gallego, D.F.; Kang, H.; Campen, M.J.; Yu, X. Microplastic presence in dog and human testis and its potential association with sperm count and weights of testis and epididymis. Toxicol. Sci. 2024, 200, 235–240. [Google Scholar] [CrossRef]
- Praveena, S.M.; Shamsul Ariffin, N.I.; Nafisyah, A.L. Microplastics in Malaysian bottled water brands: Occurrence and potential human exposure. Environ. Pollut. 2022, 315, 120494. [Google Scholar] [CrossRef]
- Yun, X.; Liang, L.; Tian, J.; Li, N.; Chen, Z.; Zheng, Y.; Duan, S.; Zhang, L. Raman-guided exploration of placental microplastic exposure: Unraveling the polymeric tapestry and assessing developmental implications. J. Hazard. Mater. 2024, 477, 135271. [Google Scholar] [CrossRef] [PubMed]
- Chartres, N.; Cooper, C.B.; Bland, G.; Pelch, K.E.; Gandhi, S.A.; BakenRa, A.; Woodruff, T.J. Effects of Microplastic Exposure on Human Digestive, Reproductive, and Respiratory Health: A Rapid Systematic Review. Environ. Sci. Technol. 2024, 58, 22843–22864. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wu, C.; Wang, Y.; Wang, L.; Tian, P.; Shang, K.X.; Zhao, J.; Wang, G. Lactobacillus plantarum reduces polystyrene microplastic induced toxicity via multiple pathways: A potentially effective and safe dietary strategy to counteract microplastic harm. J. Hazard. Mater. 2025, 489, 137669. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Chen, K.F.; Andrew Lin, K.Y.; Su, H.P.; Wu, D.N.; Lin, C.H. Evaluation of toxicity of polystyrene microplastics under realistic exposure levels in human vascular endothelial EA.hy926 cells. Chemosphere 2023, 313, 137582. [Google Scholar] [CrossRef]
- Obeng, E.M.; Hodge, C.; You, J. Microplastic pollution: A review of specific blood-tissue barrier breaches and health effects. Environ. Pollut. 2025, 376, 126416. [Google Scholar] [CrossRef]
- Kadac-Czapska, K.; Ośko, J.; Knez, E.; Grembecka, M. Microplastics and Oxidative Stress-Current Problems and Prospects. Antioxidants 2024, 13, 579. [Google Scholar] [CrossRef]
- Nguyen, V.; Taine, E.G.; Meng, D.; Cui, T.; Tan, W. Chlorogenic Acid: A Systematic Review on the Biological Functions, Mechanistic Actions, and Therapeutic Potentials. Nutrients 2024, 16, 924. [Google Scholar] [CrossRef]
- Jiang, J.; Shu, Z.; Qiu, L. Adverse effects and potential mechanisms of polystyrene microplastics (PS-MPs) on the blood-testis barrier. Environ. Geochem. Health 2024, 46, 238. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef]
- Shi, X.; Xu, T.; Gao, M.; Bi, Y.; Wang, J.; Yin, Y.; Xu, S. Combined exposure of emamectin benzoate and microplastics induces tight junction disorder, immune disorder and inflammation in carp midgut via lysosome/ROS/ferroptosis pathway. Water Res. 2024, 257, 121660. [Google Scholar] [CrossRef]
- Li, B.; Shao, H.; Gao, L.; Li, H.; Sheng, H.; Zhu, L. Nano-drug co-delivery system of natural active ingredients and chemotherapy drugs for cancer treatment: A review. Drug Deliv. 2022, 29, 2130–2161. [Google Scholar] [CrossRef] [PubMed]
- Shetty, K.; Bhandari, A.; Yadav, K.S. Nanoparticles incorporated in nanofibers using electrospinning: A novel nano-in-nano delivery system. J. Control. Release 2022, 350, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, T.; Ruttala, H.B.; Munusamy, S.; Chakraborty, N.; Kim, J.O. Nano drug delivery systems for antisense oligonucleotides (ASO) therapeutics. J. Control. Release 2022, 352, 861–878. [Google Scholar] [CrossRef]
- Jermy, B.R.; Al-Jindan, R.Y.; Ravinayagam, V.; El-Badry, A.A. Anti-blastocystosis activity of antioxidant coated ZIF-8 combined with mesoporous silicas MCM-41 and KIT-6. Sci. Rep. 2022, 12, 6403. [Google Scholar] [CrossRef]
- Sun, X.; Li, F.; Yuan, L.; Bing, Z.; Li, X.; Yang, K. pH-responsive resveratrol-loaded ZIF-8 nanoparticles modified with tannic acid for promoting colon cancer cell apoptosis. J. Biomed. Mater. Res. B Appl. Biomater. 2024, 112, e35320. [Google Scholar] [CrossRef]
- Sheik, A.; Rethinasabapathy, M.; Kodiveri Muthukaliannan, G.; Safarkhani, M.; Kang, H.; Kim, D.; Alhammadi, M.; Jung, E.; Huh, Y.S. ZIF-8 nanocarriers synthesized by co-encapsulating resveratrol and cellulase for biomedical applications. Int. J. Biol. Macromol. 2024, 283, 137756. [Google Scholar] [CrossRef]
- Brunetti, L.; Wang, L.; Wassef, A.; Gong, Y.; Brinker, A.; Buckley, B.; Lipsky, P.E.; Ondar, P.; Poiani, G.; Zhao, L.; et al. Pharmacokinetics and Pharmacodynamics of Anthocyanins after Administration of Tart Cherry Juice to Individuals with Gout. Mol. Nutr. Food Res. 2023, 67, e2200550. [Google Scholar] [CrossRef]
- Yang, D.; Zhu, J.; Zhou, X.; Pan, D.; Nan, S.; Yin, R.; Lei, Q.; Ma, N.; Zhu, H.; Chen, J.; et al. Polystyrene micro- and nano-particle coexposure injures fetal thalamus by inducing ROS-mediated cell apoptosis. Environ. Int. 2022, 166, 107362. [Google Scholar] [CrossRef]
- Mohan, M.; Gaonkar, A.A.; Pandyanda Nanjappa, D.; K, K.; Vittal, R.; Chakraborty, A.; Chakraborty, G. Screening for microplastics in drinking water and its toxicity profiling in zebrafish. Chemosphere 2023, 341, 139882. [Google Scholar] [CrossRef]
- Zeng, G.; Li, J.; Wang, Y.; Su, J.; Lu, Z.; Zhang, F.; Ding, W. Polystyrene microplastic-induced oxidative stress triggers intestinal barrier dysfunction via the NF-κB/NLRP3/IL-1β/MCLK pathway. Environ. Pollut. 2024, 345, 123473. [Google Scholar] [CrossRef]
- Li, J.; Yin, K.; Hou, L.; Zhang, Y.; Lu, H.; Ma, C.; Xing, M. Polystyrene microplastics mediate inflammatory responses in the chicken thymus by Nrf2/NF-κB pathway and trigger autophagy and apoptosis. Environ. Toxicol. Pharmacol. 2023, 100, 104136. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, G.; Tian, L.; Kong, C.; Gao, D.; Chen, Y.; Junaid, M.; Wang, J. Neuro- and hepato-toxicity of polystyrene nanoplastics and polybrominated diphenyl ethers on early life stages of zebrafish. Sci. Total Environ. 2023, 857, 159567. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, L.; Sun, B.; Ding, Y.; Lv, Y.; Wu, Q.; Zhao, S.; Zhang, X.; Shen, T. Transgenerational effects of Nanoplastics and bisphenol A on Zebrafish lipid metabolism: Disruption of the gut Microbiota-liver axis via mTOR pathway. Aquat. Toxicol. 2025, 284, 107401. [Google Scholar] [CrossRef] [PubMed]
- Del Piano, F.; Almroth, B.C.; Lama, A.; Piccolo, G.; Addeo, N.F.; Paciello, O.; Martino, G.; Esposito, S.; Mercogliano, R.; Pirozzi, C.; et al. Subchronic oral exposure to polystyrene microplastics affects hepatic lipid metabolism, inflammation, and oxidative balance in gilthead seabream (Sparus aurata). Ecotoxicol. Environ. Saf. 2024, 279, 116455. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic effects on bioaccumulation, hematological parameters, oxidative stress, immune responses and neurotoxicity in fish exposed to microplastics: A review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef]
- Liang, B.; Zhong, Y.; Huang, Y.; Lin, X.; Liu, J.; Lin, L.; Hu, M.; Jiang, J.; Dai, M.; Wang, B.; et al. Underestimated health risks: Polystyrene micro- and nanoplastics jointly induce intestinal barrier dysfunction by ROS-mediated epithelial cell apoptosis. Part. Fibre Toxicol. 2021, 18, 20. [Google Scholar] [CrossRef]
- Meng, X.; Yin, K.; Zhang, Y.; Wang, D.; Lu, H.; Hou, L.; Zhao, H.; Xing, M. Polystyrene microplastics induced oxidative stress, inflammation and necroptosis via NF-κB and RIP1/RIP3/MLKL pathway in chicken kidney. Toxicology 2022, 478, 153296. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Impact Category | Key Findings | Target Organs/Systems | References |
|---|---|---|---|
| Oxidative Stress |
| Colon, Liver | [27,28] |
| Toxicological Effects |
| Liver, Lungs, Kidneys | [28,33,35] |
| Inflammation | ↑ Pro-inflammatory cytokines (IL-6, TNF-α) and ↓ goblet cells in colon tissue.
| Colon, Lungs, Kidneys | [27,33,35] |
| Bioaccumulation |
| Brain, Kidneys, Blood, Multi-organ | [26,31,33,36] |
| Category | Representative Compounds | Mechanism of Action | References |
|---|---|---|---|
| Enzymatic Antioxidants | Superoxide Dismutase (SOD) | Catalyzes the conversion of superoxide radicals (O2−) → H2O2 + O2
| [93,94,97] |
| Catalase (CAT) | Decomposes H2O2 → H2O + O2; Prevents hydroxyl radical formation | [14,95,96] | |
| Glutathione Peroxidase (GPx) | Reduces lipid hydroperoxides and H2O2 using glutathione (GSH) as an electron donor | [14,95,96] | |
| Non-Enzymatic Antioxidants | Vitamin C, Vitamin E | Direct free radical scavenging
| [98,99,100] |
| Carotenoids | Quenches singlet oxygen and scavenges radicals
| [98,99,101] | |
| Flavonoids, Polyphenols | Neutralize radicals via phenolic hydroxyl groups
| [98,99,100] |
| Antioxidant | Sources | Bioavailability | Dose–Effect | Limitations | Mechanisms | In Vitro Evidence | In Vivo Evidence | References |
|---|---|---|---|---|---|---|---|---|
| Vitamin C | Citrus fruits, leafy greens | High (water soluble, readily absorbed) |
|
|
|
|
| [133,134,135,157,158] |
| Curcumin | Turmeric rhizomes | Low (lipophilic, requires lipid co-ingestion) |
|
|
|
|
| [111,136,159,160,161] |
| Quercetin (QSN) | Apples, onions, tea (nanoformulations) | Moderate (nanoformulations improve targeting) |
|
|
|
|
| [126,137,138,162] |
| Resveratrol | Grapes, red wine, nuts | Low (rapid metabolism, significant first-pass effect) |
|
|
|
|
| [139,140,150,151] |
| Melatonin (MT) | Plants | High bioavailability in plants; efficiently absorbed and distributed in tissues Efficiently absorbed and distributed in tissues |
|
|
|
|
| [58,142,143,144,163] |
| Glutathione (GlyNAC) | Endogenous (glycine + NAC precursors) | Limited oral absorption (requires precursor supplementation) |
|
|
|
|
| [148,149,164] |
| Astaxanthin | Algae, salmon, krill | Moderate (lipophilic, requires dietary fats) |
|
|
|
|
| [152,153,154,165,166,167] |
| Beta-carotene | Carrots, leafy greens | Dependent on dietary fats (variable conversion efficiency) |
|
|
|
|
| [155,156,168,169] |
| Combination Therapy (e.g., Vitamin E + Selenium) | Nuts, fish, whole grains | Synergistic effects (absorption mechanisms require optimization) |
|
|
|
|
| [102,103,147,157] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wang, Y.; Zhang, J.; Feng, G.; Miao, S.; Lu, R.; Tian, X.; Ye, Y. Antioxidant Intervention Against Microplastic Hazards. Antioxidants 2025, 14, 797. https://doi.org/10.3390/antiox14070797
Wang Z, Wang Y, Zhang J, Feng G, Miao S, Lu R, Tian X, Ye Y. Antioxidant Intervention Against Microplastic Hazards. Antioxidants. 2025; 14(7):797. https://doi.org/10.3390/antiox14070797
Chicago/Turabian StyleWang, Zhihua, Yunting Wang, Jian Zhang, Guoquan Feng, Shuhan Miao, Rongzhu Lu, Xinyu Tian, and Yang Ye. 2025. "Antioxidant Intervention Against Microplastic Hazards" Antioxidants 14, no. 7: 797. https://doi.org/10.3390/antiox14070797
APA StyleWang, Z., Wang, Y., Zhang, J., Feng, G., Miao, S., Lu, R., Tian, X., & Ye, Y. (2025). Antioxidant Intervention Against Microplastic Hazards. Antioxidants, 14(7), 797. https://doi.org/10.3390/antiox14070797