
In Vitro Maturation of Bovine Oocytes in the Presence of Resveratrol and Ellagic Acid but Not Chlorogenic Acid Modulates Blastocyst Antioxidant Gene Expression Without Impacting Embryo Development and Oxygen Consumption

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Production and Culture of Bovine Embryos
2.2. In Vitro Production and Cryopreservation of Embryos for Extracellular Flux Analysis
2.3. Analysis of Blastocyst Real-Time Oxygen Consumption Rate
2.4. Isolation of Blastocyst Total RNA and Gene Expression Analysis
2.5. Statistical Analysis
3. Results
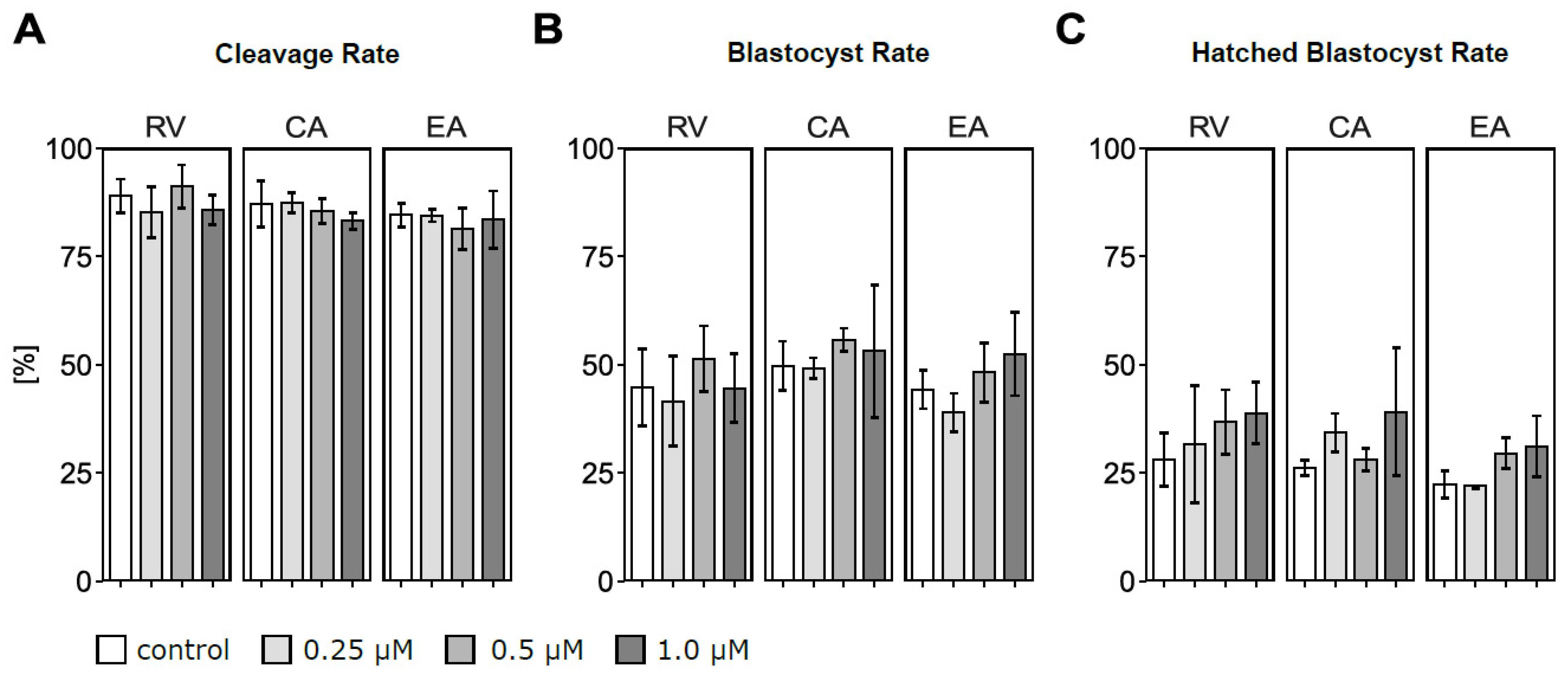
3.1. In Vitro Maturation in the Presence of Polyphenols Did Not Affect Embryo Development
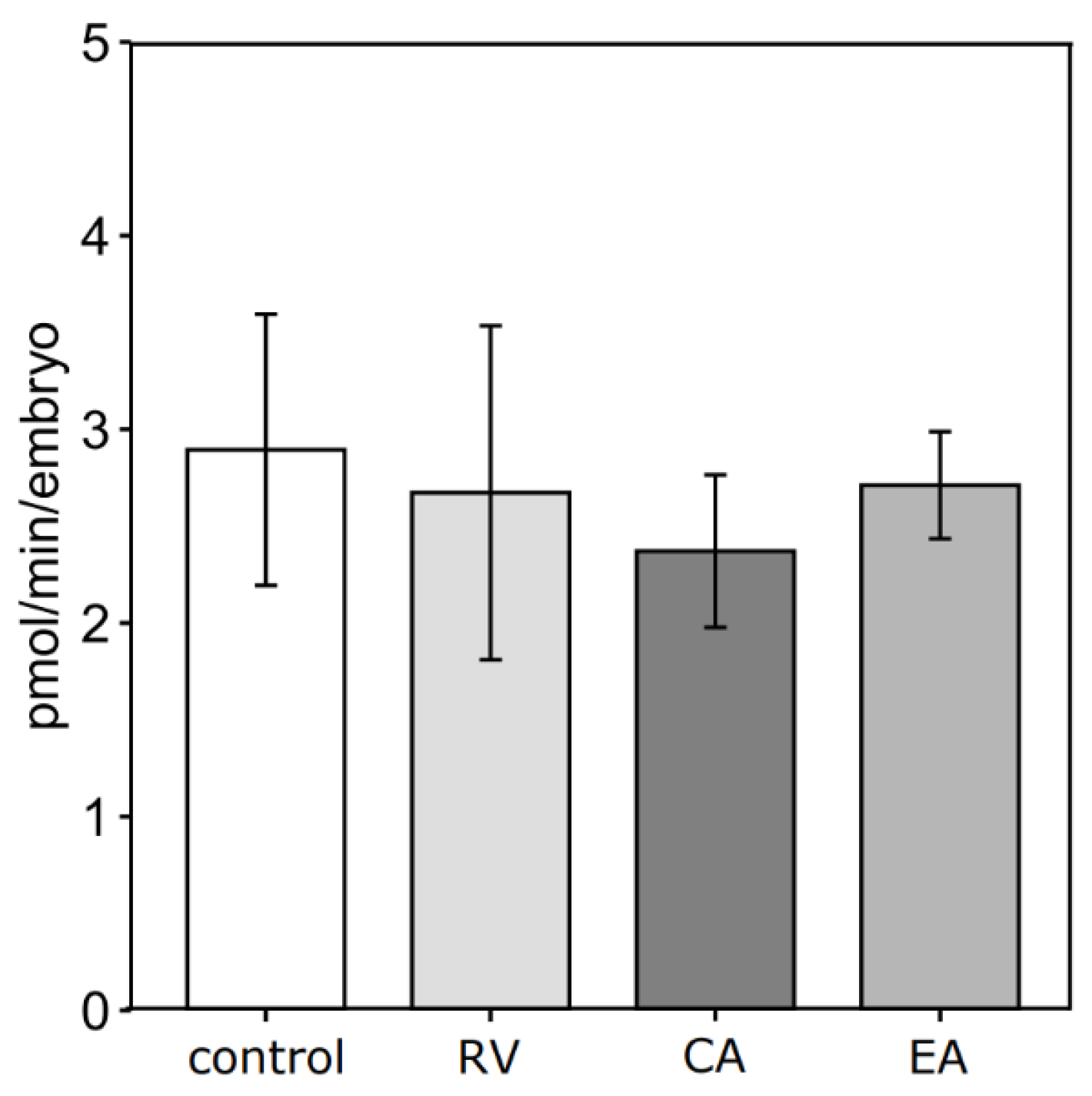
3.2. Polyphenol Supplementation During In Vitro Maturation Did Not Affect Blastocyst Oxygen Consumption Rate
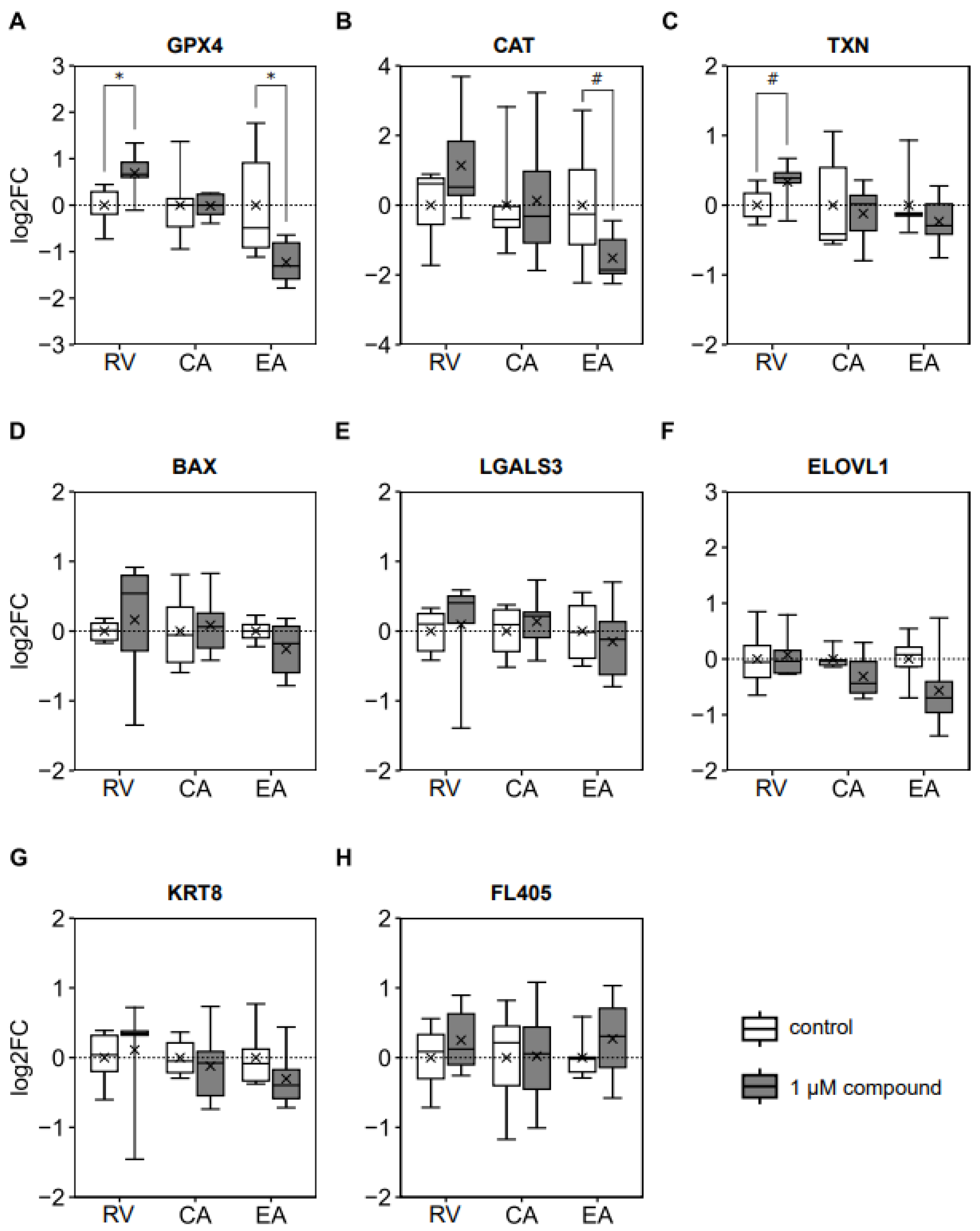
3.3. Blastocyst mRNA Expression of Genes Involved in Antioxidant Defense Was Affected by In Vitro Maturation in the Presence of Ellagic Acid and Resveratrol
4. Discussion
4.1. Embryo Development
4.2. Blastocyst Oxygen Consumption
4.3. Blastocyst Gene Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cleavage Rate (%) | Blastocyst Rate (%) | Hatched Blastocyst Rate (%) | |
|---|---|---|---|
| control | 88.9 ± 3.81 | 48.3 ± 3.26 | 33.6 ± 3.97 |
| DMSO control | 86.9 ± 1.29 | 46.2 ± 1.74 | 25.5 ± 1.72 |
| p-value | 0.647 | 0.603 | 0.135 |
| Treatment | Oocytes | Cleaved Embryos | Blastocysts | Hatched Blastocysts | |
|---|---|---|---|---|---|
| RV | control | 84 | 75 | 38 | 24 |
| 0.25 µM | 83 | 71 | 35 | 27 | |
| 0.5 µM | 82 | 75 | 42 | 30 | |
| 1 µM | 83 | 71 | 37 | 32 | |
| CA | control | 70 | 61 | 34 | 18 |
| 0.25 µM | 70 | 61 | 35 | 25 | |
| 0.5 µM | 70 | 60 | 39 | 20 | |
| 1 µM | 69 | 57 | 34 | 24 | |
| EA | control | 84 | 71 | 37 | 19 |
| 0.25 µM | 82 | 69 | 32 | 18 | |
| 0.5 µM | 83 | 67 | 39 | 24 | |
| 1 µM | 82 | 68 | 42 | 25 |
References
- Keane, J.A.; Ealy, A.D. An Overview of Reactive Oxygen Species Damage Occurring during In Vitro Bovine Oocyte and Embryo Development and the Efficacy of Antioxidant Use to Limit These Adverse Effects. Animals 2024, 14, 330. [Google Scholar] [CrossRef]
- Lizarraga, C.; Gaxiola, S.M.; Castro del Campo, N.; Enriquez-Verduzco, I.; Rodriguez-Gaxiola, M.A.; Montero-Pardo, A.; Hernandez-Carreño, P.E.; Valdez-Ramirez, A.; Diaz, D. Scoping review of primary studies evaluating Bovine in vitro oocyte maturation and embryo development. Vet. Méx. OA 2024, 11. [Google Scholar] [CrossRef]
- Namula, Z.; Sato, Y.; Wittayarat, M.; Le, Q.A.; Nguyen, N.T.; Lin, Q.; Hirata, M.; Tanihara, F.; Otoi, T. Curcumin supplementation in the maturation medium improves the maturation, fertilisation and developmental competence of porcine oocytes. Acta Vet. Hung. 2020, 68, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Gupta, S.; Sekhon, L.; Shah, R. Redox considerations in female reproductive function and assisted reproduction: From molecular mechanisms to health implications. Antiox. Redox Signal. 2008, 10, 1375–1403. [Google Scholar] [CrossRef]
- Chiumia, D.; Hankele, A.K.; Drews, B.; Zehnder, T.; Berard, J.; Giesbertz, P.; Ulbrich, S.E.; Giller, K. Alpine and lowland grazing differentially alter the reproductive tract redox milieu and amino acid composition in cattle. Anim. Reprod. Sci. 2020, 213, 106268. [Google Scholar] [CrossRef]
- Milojevic, V.; Sinz, S.; Kreuzer, M.; Chiumia, D.; Marquardt, S.; Giller, K. Partitioning of fatty acids into tissues and fluids from reproductive organs of ewes as affected by dietary phenolic extracts. Theriogenology 2020, 144, 174–184. [Google Scholar] [CrossRef]
- Silva, B.R.; Silva, J.R.V. Mechanisms of action of non-enzymatic antioxidants to control oxidative stress during in vitro follicle growth, oocyte maturation, and embryo development. Anim. Reprod. Sci. 2023, 249, 107186. [Google Scholar] [CrossRef]
- Ruskovska, T.; Maksimova, V.; Milenkovic, D. Polyphenols in human nutrition: From the in vitro antioxidant capacity to the beneficial effects on cardiometabolic health and related inter-individual variability—An overview and perspective. Br. J. Nutr. 2020, 123, 241–254. [Google Scholar] [CrossRef]
- You, J.; Kim, J.; Lim, J.; Lee, E. Anthocyanin stimulates in vitro development of cloned pig embryos by increasing the intracellular glutathione level and inhibiting reactive oxygen species. Theriogenology 2010, 74, 777–785. [Google Scholar] [CrossRef]
- Kwak, S.S.; Cheong, S.A.; Jeon, Y.; Lee, E.; Choi, K.C.; Jeung, E.B.; Hyun, S.H. The effects of resveratrol on porcine oocyte in vitro maturation and subsequent embryonic development after parthenogenetic activation and in vitro fertilization. Theriogenology 2012, 78, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Sovernigo, T.C.; Adona, P.R.; Monzani, P.S.; Guemra, S.; Barros, F.; Lopes, F.G.; Leal, C. Effects of supplementation of medium with different antioxidants during in vitro maturation of bovine oocytes on subsequent embryo production. Reprod. Domest. Anim. 2017, 52, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, Y.; Li, Y.; Jin, Q.; Sun, J.; E, Z.; Gao, Q. Supplementation of kaempferol to in vitro maturation medium regulates oxidative stress and enhances subsequent embryonic development in vitro. Zygote 2020, 28, 59–64. [Google Scholar] [CrossRef]
- Pocar, P.; Augustin, R.; Fischer, B. Constitutive expression of CYP1A1 in bovine cumulus oocyte-complexes in vitro: Mechanisms and biological implications. Endocrinology 2004, 145, 1594–1601. [Google Scholar] [CrossRef]
- Wu, Z.L.; Yang, Y.; Chen, Y.; Xia, G.L.; Zhang, R.Q. Effects of subcutaneous administration of daidzein on blastocyst implantation in rats. Food Chem. Toxicol. 2005, 43, 167–172. [Google Scholar] [CrossRef]
- Lee, K.; Wang, C.; Chaille, J.M.; Machaty, Z. Effect of resveratrol on the development of porcine embryos produced in vitro. J. Reprod. Dev. 2010, 56, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tian, X.; Zhang, L.; He, C.; Ji, P.; Li, Y.; Tan, D.; Liu, G. Beneficial effect of resveratrol on bovine oocyte maturation and subsequent embryonic development after in vitro fertilization. Fertil. Steril. 2014, 101, 577–586. [Google Scholar] [CrossRef]
- Nunes, H.P.; Furnas, S.; Dinis, M.; Borba, A.; da Silva, J.M. Effects of chlorogenic acid on the maturation and fertilisation of bovine oocytes and their embryonic development with a comparative bovine granulosa cell co-culture. Anim. Reprod. 2017, 14, 1057–1061. [Google Scholar] [CrossRef]
- Lee, H.B.; Lee, S.E.; Park, M.J.; Han, D.H.; Lim, E.S.; Ryu, B.; Kim, E.Y.; Park, S.P. Ellagic acid treatment during in vitro maturation of porcine oocytes improves development competence after parthenogenetic activation and somatic cell nuclear transfer. Theriogenology 2024, 215, 214–223. [Google Scholar] [CrossRef]
- Stringfellow, D.A.; Givens, M.D. Manual of the International Embryo Transfer Society: A Procedural Guide and General Information for the Use of Embryo Transfer Technology Emphasizing Sanitary Procedures, 4th ed.; International Embryo Transfer Society: Champaign, IL, USA, 2010. [Google Scholar]
- Gardner, D.K.; Lane, M.; Spitzer, A.; Batt, P.A. Enhanced rates of cleavage and development for sheep zygotes culture to the blastocyst stage in vitro in the absence of serum and somatic cells: Amino acids, vitamins, and culturing embryos in groups stimulated development. Biol. Reprod. 1994, 50, 390–400. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Torres, V.; Muñoz, L.; Urrego, R.; Echeverry, J.J.; Lopez, A. Resveratrol during in vitro maturation improves the quality of bovine oocyte and enhances embryonic development in vitro. Reprod. Fertil. Dev. 2017, 29, 199. [Google Scholar] [CrossRef]
- Wein, S.; Beyer, B.; Gohlke, A.; Blank, R.; Metges, C.C.; Wolffram, S. Systemic absorption of catechins after intraruminal or intraduodenal application of a green tea extract in cows. PLoS ONE 2016, 11, e0159428. [Google Scholar] [CrossRef]
- Torres, V.; Hamdi, M.; Millán de la Blanca, M.G.; Urrego, R.; Echeverri, J.; López-Herrera, A.; Rizos, D.; Gutiérrez-Adán, A.; Sánchez-Calabuig, M.J. Resveratrol-cyclodextrin complex affects the expression of genes associated with lipid metabolism in bovine in vitro produced embryos. Reprod. Domest. Anim. 2018, 53, 850–858. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Machado-Simões, J.; Moniz, I.; Soares, M.; Carvalho, A.; Diniz, P.; Ramalho-Santos, J.; Sousa, A.P.; Lopes-da-Costa, L.; Almeida-Santos, T. Chemical reversion of age-related oocyte dysfunction fails to enhance embryo development in a bovine model of postovulatory aging. J. Assist. Reprod. Genet. 2024, 41, 1997–2009. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Synergistic effects of resveratrol and melatonin on in vitro maturation of porcine oocytes and subsequent embryo development. Theriogenology 2018, 114, 191–198. [Google Scholar] [CrossRef]
- Liu, M.J.; Sun, A.G.; Zhao, S.G.; Liu, H.; Ma, S.Y.; Li, M.; Huai, Y.X.; Zhao, H.; Liu, H.B. Resveratrol improves in vitro maturation of oocytes in aged mice and humans. Fertil. Steril. 2018, 109, 900–907. [Google Scholar] [CrossRef]
- Wang, X.F.; Zhu, X.X.; Liang, X.W.; Xu, H.Y.; Liao, Y.Y.; Lu, K.H.; Lu, S.S. Effects of resveratrol on in vitro maturation of porcine oocytes and subsequent early embryonic development following somatic cell nuclear transfer. Reprod. Domest. Anim. 2019, 54, 1195–1205. [Google Scholar] [CrossRef]
- Piras, A.R.; Menéndez-Blanco, I.; Soto-Heras, S.; Catalá, M.G.; Izquierdo, D.; Bogliolo, L.; Paramio, M.T. Resveratrol supplementation during in vitro maturation improves embryo development of prepubertal goat oocytes selected by brilliant cresyl blue staining. J. Reprod. Dev. 2019, 65, 113–120. [Google Scholar] [CrossRef]
- Piras, A.R.; Ariu, F.; Falchi, L.; Zedda, M.T.; Pau, S.; Schianchi, E.; Paramio, M.; Bogliolo, L. Resveratrol treatment during maturation enhances developmental competence of oocytes after prolonged ovary storage at 4 °C in the domestic cat model. Theriogenology 2020, 144, 152–157. [Google Scholar] [CrossRef]
- Zabihi, A.; Shabankareh, H.K.; Hajarian, H.; Foroutanifar, S. In vitro maturation medium supplementation with resveratrol improves cumulus cell expansion and developmental competence of Sanjabi sheep oocytes. Livest. Sci. 2021, 243, 104378. [Google Scholar] [CrossRef]
- Kabeer, S.W.; Riaz, A.; Ul-Rahman, A.; Shahbakht, R.M.; Anjum, A.; Khera, H.u.R.A.; Haider, A.; Riaz, F.; Yasin, R.; Yaseen, M.; et al. Effect of different concentrations of resveratrol on nuclear maturation and in-vitro development competence of oocytes of Nili Ravi buffalo. Trop. Anim. Health Prod. 2024, 56, 105. [Google Scholar] [CrossRef]
- Takeo, S.; Sato, D.; Kimura, K.; Monji, Y.; Kuwayama, T.; Kawahara-Miki, R.; Iwata, H. Resveratrol improves the mitochondrial function and fertilization outcome of bovine oocytes. J. Reprod. Dev. 2014, 60, 92–99. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Tanihara, F.; Do, L.; Sato, Y.; Taniguchi, M.; Takagi, M.; Van Nguyen, T.; Otoi, T. Chlorogenic acid supplementation during in vitro maturation improves maturation, fertilization and developmental competence of porcine oocytes. Reprod. Domest. Anim. 2017, 52, 969–975. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Wittayarat, M.; Do, L.T.K.; Nguyen, T.V.; Nii, M.; Namula, Z.; Kunihara, T.; Tanihara, F.; Hirata, M.; Otoi, T. Effects of chlorogenic acid (CGA) supplementation during in vitro maturation culture on the development and quality of porcine embryos with electroporation treatment after in vitro fertilization. Anim. Sci. J. 2018, 89, 1207–1213. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Do, L.T.K.; Somfai, T.; Otoi, T.; Taniguchi, M.; Kikuchi, K. Presence of chlorogenic acid during in vitro maturation protects porcine oocytes from the negative effects of heat stress. Anim. Sci. J. 2019, 90, 1530–1536. [Google Scholar] [CrossRef]
- Thongkittidilok, C.; Le, Q.A.; Lin, Q.; Takebayashi, K.; Do, T.K.L.; Namula, Z.; Hirata, M.; Tanihara, F.; Otoi, T. Effects of individual or in-combination antioxidant supplementation during in vitro maturation culture on the developmental competence and quality of porcine embryos. Reprod. Domest. Anim. 2022, 57, 314–320. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Do, L.T.K.; Nguyen, N.T.; Kikuchi, K.; Somfai, T.; Otoi, T. The effects of an in vitro oocyte maturation system and chlorogenic acid supplementation during embryo culture on the development of porcine cloned embryos derived from native Vietnamese Ban pigs. Vet. Med. Int. 2023, 17, 5702970. [Google Scholar] [CrossRef]
- Kawasaki, K.; Hirai, M.; Ishiki, Y.; Nagahama, A.; Konno, T.; Yamanaka, K.; Tatemoto, H. The strong anti-hyaluronidase effect of ellagic acid markedly decreases polyspermy during in vitro fertilization, resulting in sustainment of the developmental potency in porcine oocytes. Theriogenology 2024, 215, 95–102. [Google Scholar] [CrossRef]
- Perez-Pasten, R.; Martinez-Galero, E.; Chamorro-Cevallos, G. Quercetin and naringenin reduce abnormal development of mouse embryos produced by hydroxyurea. J. Pharm. Pharmacol. 2010, 62, 1003–1009. [Google Scholar] [CrossRef]
- Banihosseini, S.Z.; Novin, M.G.; Nazarian, H.; Piryaei, A.; Parvardeh, S.; Eini, F. Quercetin improves developmental competence of mouse oocytes by reducing oxidative stress during in vitro maturation. Ann. Anim. Sci. 2018, 18, 87–98. [Google Scholar] [CrossRef]
- Leese, H.J.; Guerif, F.; Allgar, V.; Brison, D.R.; Lundin, K.; Sturmey, R.G. Biological optimization, the Goldilocks Principle, and how much is lagom in the preimplantation embryo. Mol. Reprod. Dev. 2016, 83, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Lewis, N.; Adeniyi, T.; Leese, H.J.; Brison, D.R.; Sturmey, R.G. Application of extracellular flux analysis for determining mitochondrial function in mammalian oocytes and early embryos. Sci. Rep. 2019, 9, 16778. [Google Scholar] [CrossRef]
- Overstrom, E.W.; Duby, R.T.; Dobrinsky, J.R.; Roche, J.F.; Boland, M.P. Viability and oxidative metabolism of the bovine blastocyst. Theriogenology 1992, 37, 269. [Google Scholar] [CrossRef]
- Venturas, M.; Shah, J.S.; Yang, X.; Sanchez, T.H.; Conway, W.; Sakkas, D.; Needleman, D.J. Metabolic state of human blastocysts measured by fluorescence lifetime imaging microscopy. Hum. Reprod. 2022, 37, 411–427. [Google Scholar] [CrossRef]
- Almansa-Ordonez, A.; Bellido, R.; Vassena, R.; Barragan, M.; Zambelli, F. Oxidative stress in reproduction: A mitochondrial perspective. Biology 2020, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Q.; Pang, Y.W.; Hao, H.S.; Du, W.H.; Zhao, X.M.; Zhu, H.B. Effects of epigallocatechin-3-gallate on bovine oocytes matured in vitro. Asian-Australas. J. Anim. Sci. 2018, 31, 1420–1430. [Google Scholar] [CrossRef]
- Wang, Z.G.; Fu, C.Q.; Yu, S.D. Green tea polyphenols added to IVM and IVC media affect transcript abundance, apoptosis, and pregnancy rates in bovine embryos. Theriogenology 2013, 79, 186–192. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, H.; Wang, Z.; Zhang, C.; Bian, Y.; Liu, X.; Zhang, C.; Zhang, X.; Zhao, Y. Quercetin promotes in vitro maturation of oocytes from humans and aged mice. Cell Death Dis. 2020, 11, 965. [Google Scholar] [CrossRef]
- Hawkes, H.J.; Karlenius, T.C.; Tonissen, K.F. Regulation of the human thioredoxin gene promoter and its key substrates: A study of functional and putative regulatory elements. Biochim. Biophys. Acta 2014, 1840, 303–314. [Google Scholar] [CrossRef]
- Divyajanani, S.; Harithpriya, K.; Ganesan, K.; Ramkumar, K.M. Dietary polyphenols remodel DNA methylation patterns of NRF2 in chronic disease. Nutrients 2023, 15, 3347. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative stress, antioxidant capabilities, and bioavailability: Ellagic acid or urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Pasqualetti, V.; Locato, V.; Fanali, C.; Mulinacci, N.; Cimini, S.; Morgia, A.M.; Pasqua, G.; de Gara, L. Comparison between in vitro chemical and ex vivo biological assays to evaluate antioxidant capacity of botanical extracts. Antioxidants 2021, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Caruso, F.; Tanski, J.; Villegas-Estrada, A.; Rossi, M. Structural basis for antioxidant activity of trans-resveratrol: Ab initio calculations and crystal and molecular structure. J. Agric. Food Chem. 2004, 52, 7279–7285. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Nguyen, V.; Taine, E.G.; Meng, D.; Cui, T.; Tan, W. Chlorogenic acid: A systematic review on the biological functions, mechanistic actions, and therapeutic potentials. Nutrients 2024, 16, 924. [Google Scholar] [CrossRef] [PubMed]
- Baud, O.; Greene, A.E.; Li, J.; Wang, H.; Volpe, J.J.; Rosenberg, P.A. Glutathione peroxidase-catalase cooperativity is required for resistance to hydrogen peroxide by mature rat oligodendrocytes. J. Neurosci. 2004, 24, 1531–1540. [Google Scholar] [CrossRef]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Ho, Y.S.; Xiong, Y.; Ma, W.; Spector, A.; Ho, D.S. Mice lacking catalase develop normally but show differential sensitivity to oxidant tissue injury. J. Biol. Chem. 2004, 279, 32804–32812. [Google Scholar] [CrossRef]
- Glorieux, C.; Zamocky, M.; Sandoval, J.M.; Verrax, J.; Calderon, P.B. Regulation of catalase expression in healthy and cancerous cells. Free Radic. Biol. Med. 2015, 87, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Boumela, I.; Assou, S.; Aouacheria, A.; Haouzi, D.; Dechaud, H.; De Vos, J.; Handyside, A.; Hamamah, S. Involvement of BCL2 family members in the regulation of human oocyte and early embryo survival and death: Gene expression and beyond. Reproduction 2011, 141, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Li, C.; Gao, F.; Fan, Y.; Zeng, F.; Meng, L.; Li, L.; Zhang, S.; Wie, H. Epigallocatechin-3-gallate promotes the in vitro maturation and embryo development following IVF of porcine oocytes. Drug Des. Devel. Ther. 2021, 15, 1013–1020. [Google Scholar] [CrossRef]
- Rizos, D.; Gutierrez-Adan, A.; Perez-Garnelo, S.; de la Fuente, J.; Boland, M.P.; Lonergan, P. Bovine embryo culture in the presence or absence of serum: Implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol. Reprod. 2003, 68, 236–243. [Google Scholar] [CrossRef]
- Lee, K.S.; Kim, E.Y.; Jeon, K.; Cho, S.G.; Han, Y.J.; Yang, B.C.; Lee, S.S.; Ko, M.S.; Riu, K.J.; Lee, H.T.; et al. 3,4-Dihydroxyflavone acts as an antioxidant and antiapoptotic agent to support bovine embryo development in vitro. J. Reprod. Dev. 2011, 57, 127–134. [Google Scholar] [CrossRef]
- Jeon, Y.; Kwak, S.S.; Cheong, S.A.; Seong, Y.H.; Hyun, S.H. Effect of trans-epsilon-viniferin on in vitro porcine oocyte maturation and subsequent developmental competence in preimplantation embryos. J. Vet. Med. Sci. 2013, 75, 1277–1286. [Google Scholar] [CrossRef]
- Machado, G.M.; Caixeta, E.S.; Lucci, C.M.; Rumpf, R.; Franco, M.M.; Dode, M.A.N. Post-hatching development of bovine embryos in vitro: The effects of tunnel preparation and gender. Zygote 2012, 20, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, N.; Salilew-Wondim, D.; Gad, A.; Tesfaye, D.; Phatsara, C.; Tholen, E.; Looft, C.; Schellander, K.; Hoelker, M. Bovine blastocysts with developmental competence to term share similar expression of developmentally important genes although derived from different culture environments. Reproduction 2011, 142, 551–564. [Google Scholar] [CrossRef]
- Chen, H.W.; Chen, J.J.; Yu, S.L.; Li, H.N.; Yang, P.C.; Su, C.M.; Au, H.K.; Chang, C.W.; Chien, L.W.; Chen, C.S.; et al. Transcriptome analysis in blastocyst hatching by cDNA microarray. Hum. Reprod. 2005, 20, 2492–2501. [Google Scholar] [CrossRef]
| Gene | Accession Number | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon Length (bp) |
|---|---|---|---|---|
| ACTB | NM_173979.3 | GATCTGGCACCACACCTTCT | AGAGACAGCACAGCCTGGAT | 174 |
| BAX | NM_173894.1 | GCCCTTTTGCTTCAGGGTTT | ACAGCTGCGATCATCCTCTG | 179 |
| CAT | NM_001035386.2 | CTGGGACCCAACTATCTCCA | AAGTGGGTCCTGTGTTCCAG | 179 |
| ELOVL1 | NM_001034703 | GTACTTCGTCCTCTCACTGG | GCCAGGTGTAGGAACTTAGCC | 158 |
| FL405 | AY308069.1 | ACCAGCTCAATCTGCCTCCG | CCGTGGGCAATCATAAGGGC | 146 |
| GPX4 | NM_174770.4 | ACCCTCTGTGGAAATGGATG | GAAGGCTTCTCGGAACACAG | 228 |
| H3-3A | NM_001014389.2 | GTACTGTGGCACTCCGTGAA | GATAGGCCTCACTTGCCTCC | 168 |
| KRT8 | NM_001033610.1 | GAATGTGCCTTATGACCTGCC | GGCTGTAGTTGAAGCCAGGG | 179 |
| LGALS3 | NM_001102341.2 | GAATGTGCCTTATGACCTGCC | TGGAAGGCGACATCATTCCC | 132 |
| TXN | NM_173968.3 | ATCGATTGCACTGTCAGGTCGC | TCTCTCCTGCACTGTTCAAGGC | 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giller, K.; Schmid, D.; Serbetci, I.; Meleán, M.; Greve, S.; von Meyenn, F.; Bollwein, H.; Herrera, C. In Vitro Maturation of Bovine Oocytes in the Presence of Resveratrol and Ellagic Acid but Not Chlorogenic Acid Modulates Blastocyst Antioxidant Gene Expression Without Impacting Embryo Development and Oxygen Consumption. Antioxidants 2025, 14, 621. https://doi.org/10.3390/antiox14060621
Giller K, Schmid D, Serbetci I, Meleán M, Greve S, von Meyenn F, Bollwein H, Herrera C. In Vitro Maturation of Bovine Oocytes in the Presence of Resveratrol and Ellagic Acid but Not Chlorogenic Acid Modulates Blastocyst Antioxidant Gene Expression Without Impacting Embryo Development and Oxygen Consumption. Antioxidants. 2025; 14(6):621. https://doi.org/10.3390/antiox14060621
Chicago/Turabian StyleGiller, Katrin, Dominique Schmid, Idil Serbetci, Manuel Meleán, Sarah Greve, Ferdinand von Meyenn, Heinrich Bollwein, and Carolina Herrera. 2025. "In Vitro Maturation of Bovine Oocytes in the Presence of Resveratrol and Ellagic Acid but Not Chlorogenic Acid Modulates Blastocyst Antioxidant Gene Expression Without Impacting Embryo Development and Oxygen Consumption" Antioxidants 14, no. 6: 621. https://doi.org/10.3390/antiox14060621
APA StyleGiller, K., Schmid, D., Serbetci, I., Meleán, M., Greve, S., von Meyenn, F., Bollwein, H., & Herrera, C. (2025). In Vitro Maturation of Bovine Oocytes in the Presence of Resveratrol and Ellagic Acid but Not Chlorogenic Acid Modulates Blastocyst Antioxidant Gene Expression Without Impacting Embryo Development and Oxygen Consumption. Antioxidants, 14(6), 621. https://doi.org/10.3390/antiox14060621