
A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Desizgn
2.3. Determination Indices and Methods
2.3.1. Determination of Polyphenols
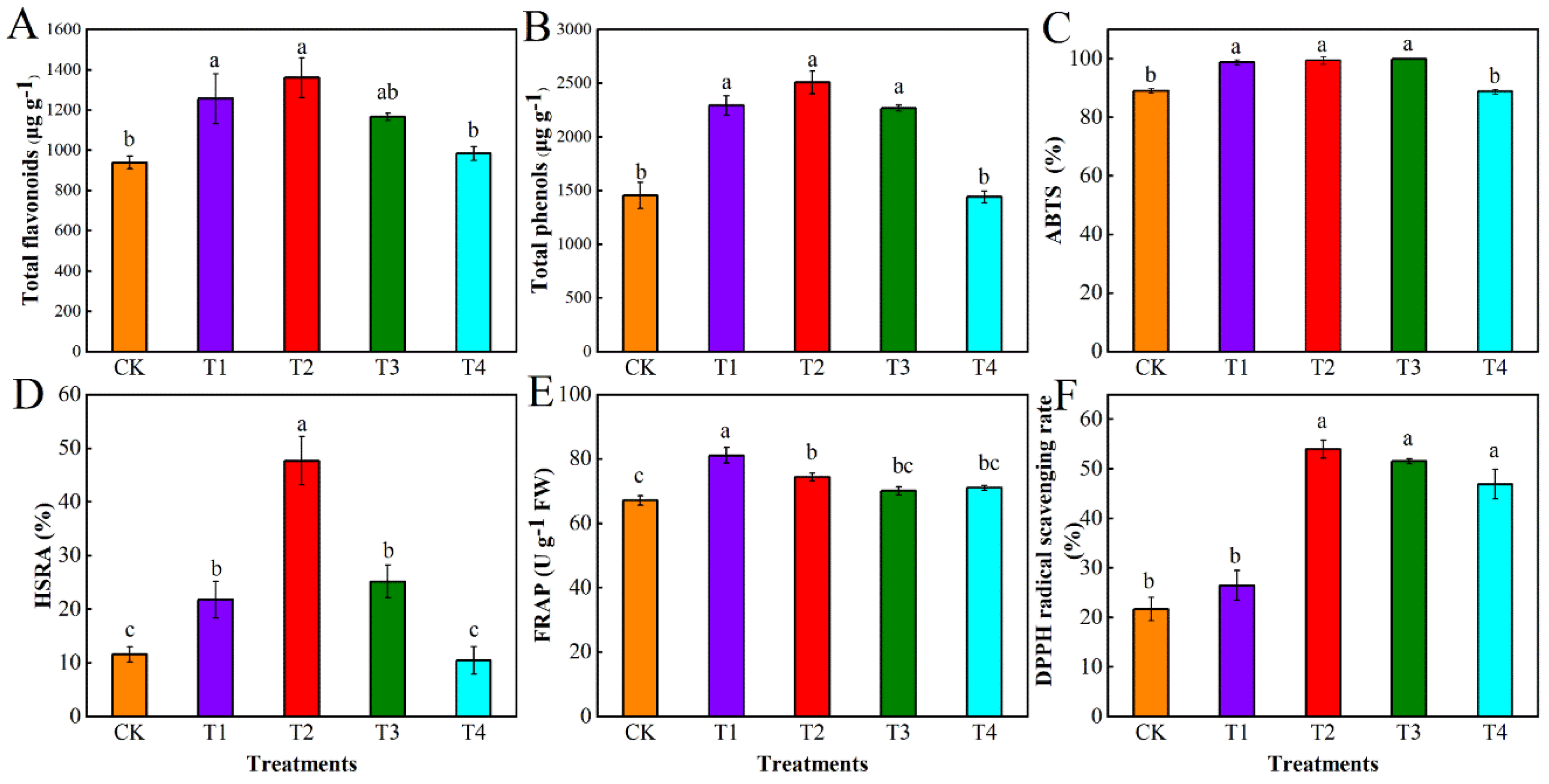
2.3.2. Determination of Antioxidant Parameters
2.3.3. Determination of Nutritional and Safety Qualities
2.4. Statistical Analysis
3. Results
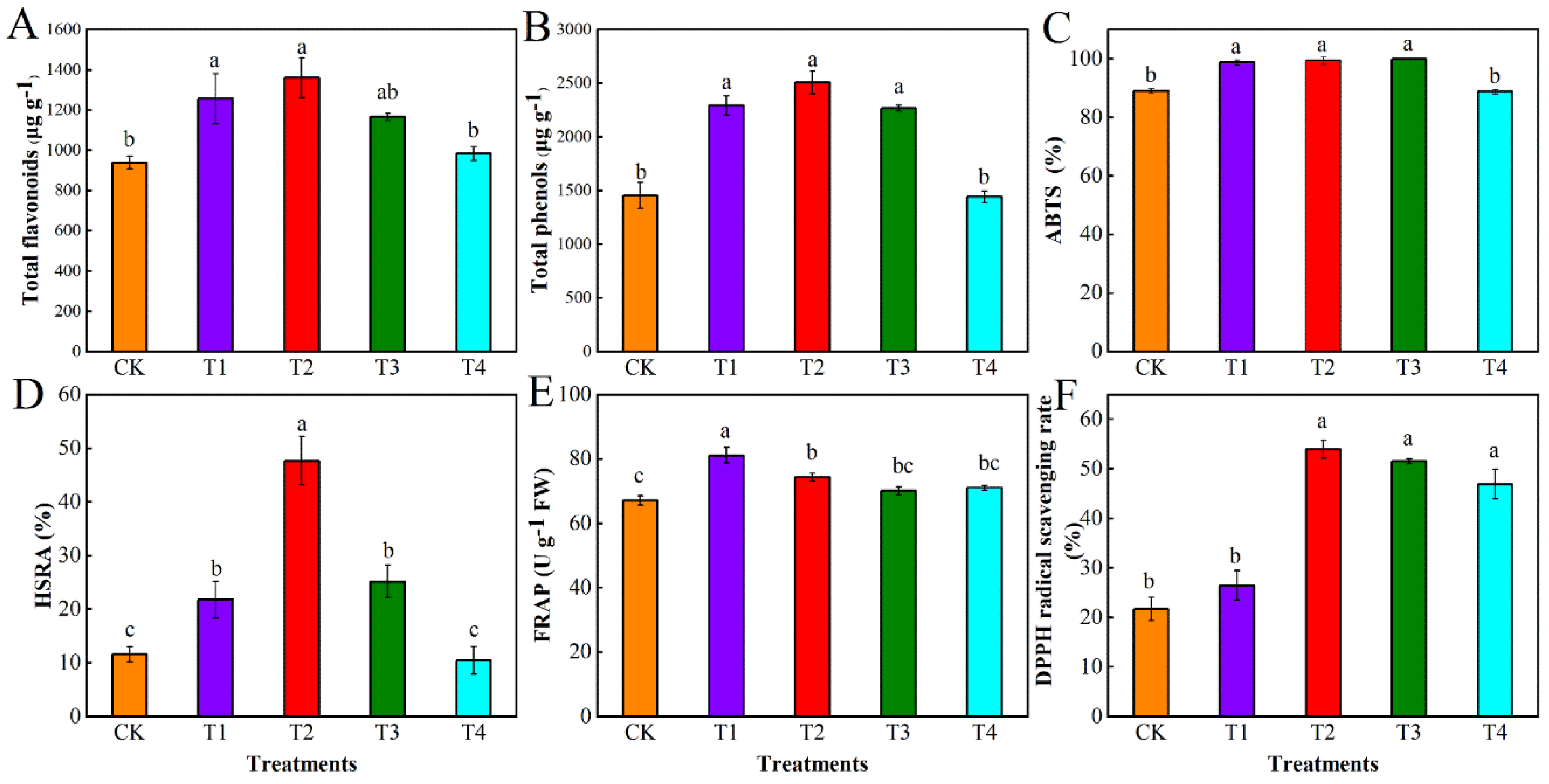
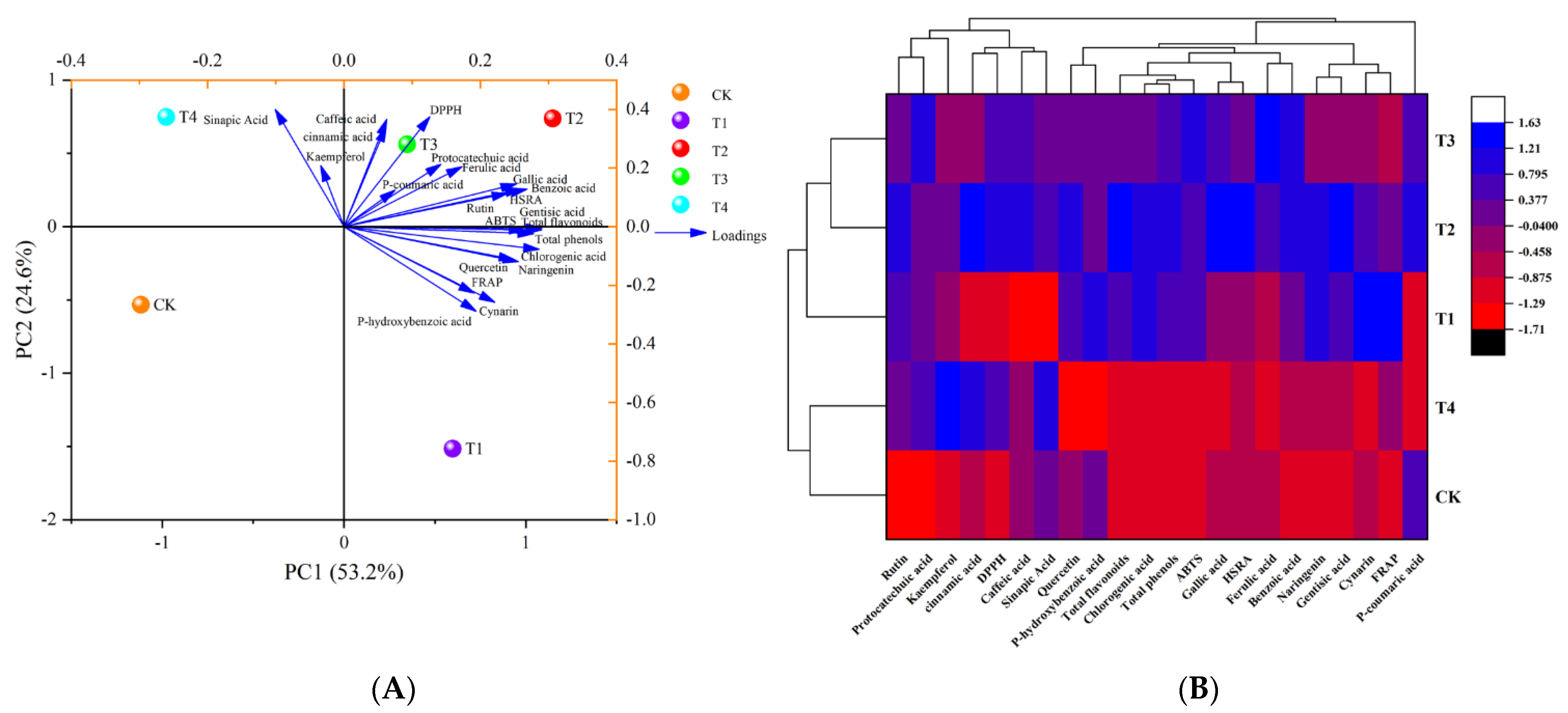
3.1. Effect of Different Levels of Water Deficit on Polyphenol Content and Antioxidant Parameters in Tomato Fruit
3.1.1. Variance Analysis
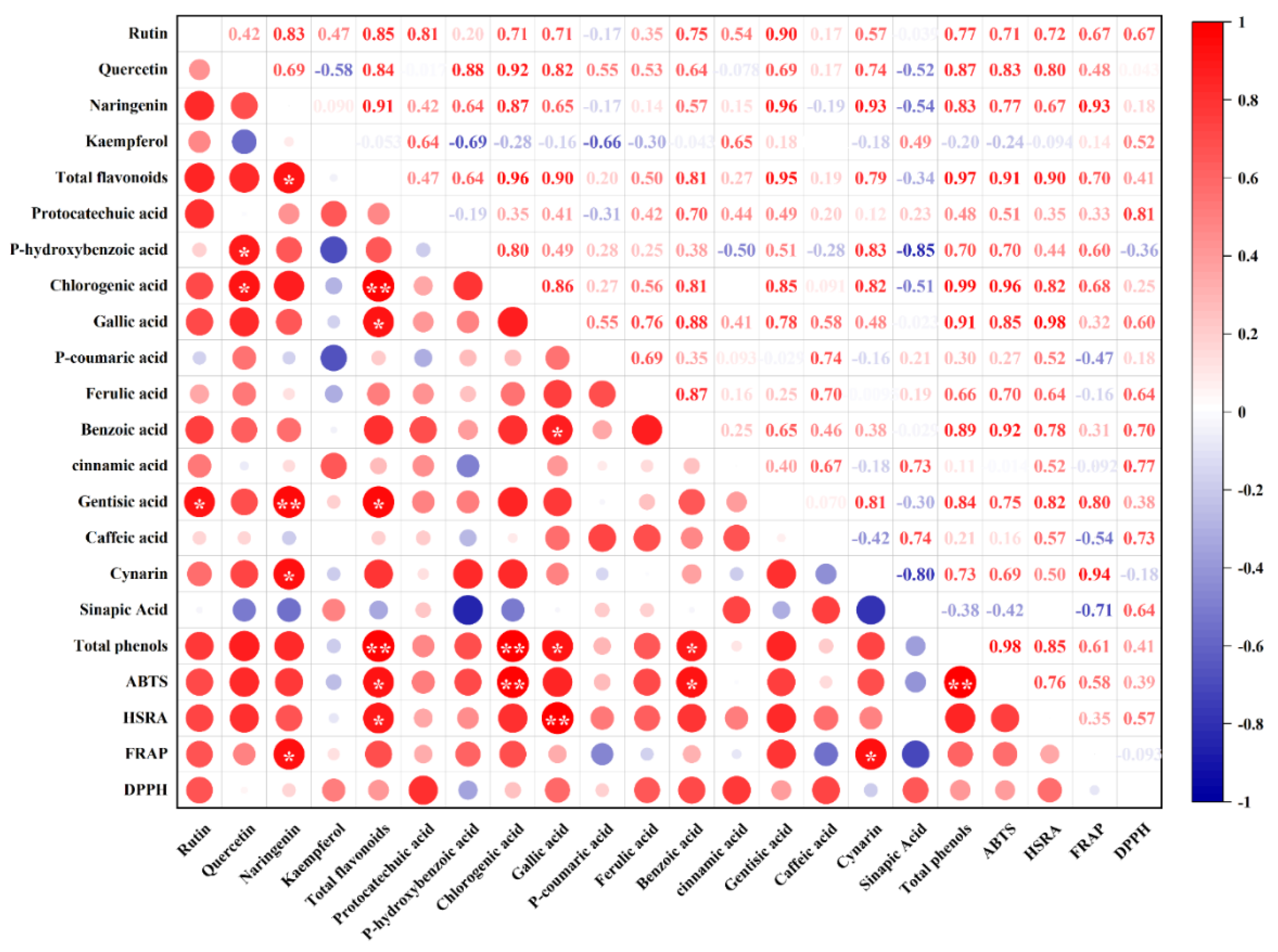
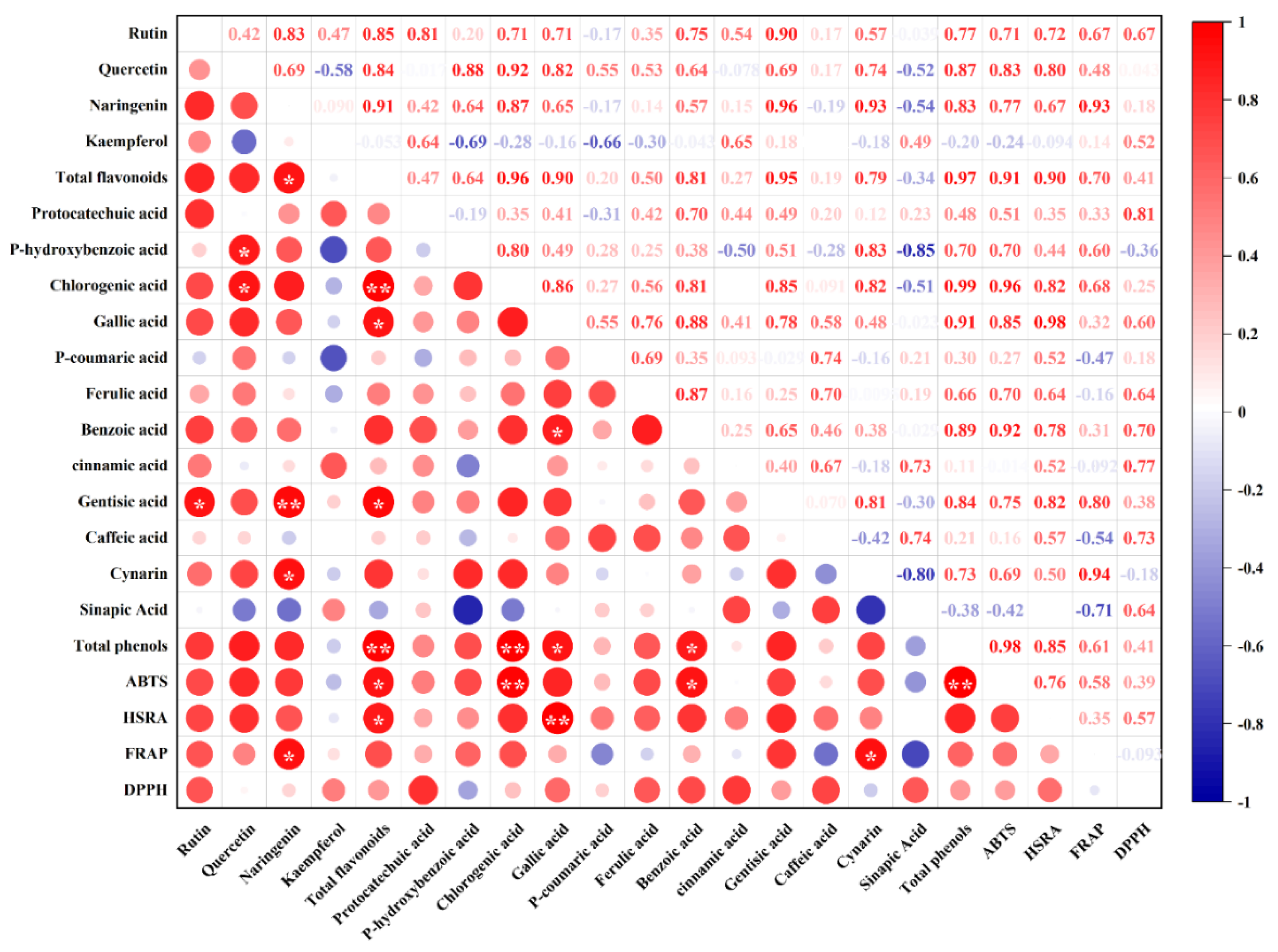
3.1.2. Correlation Analysis
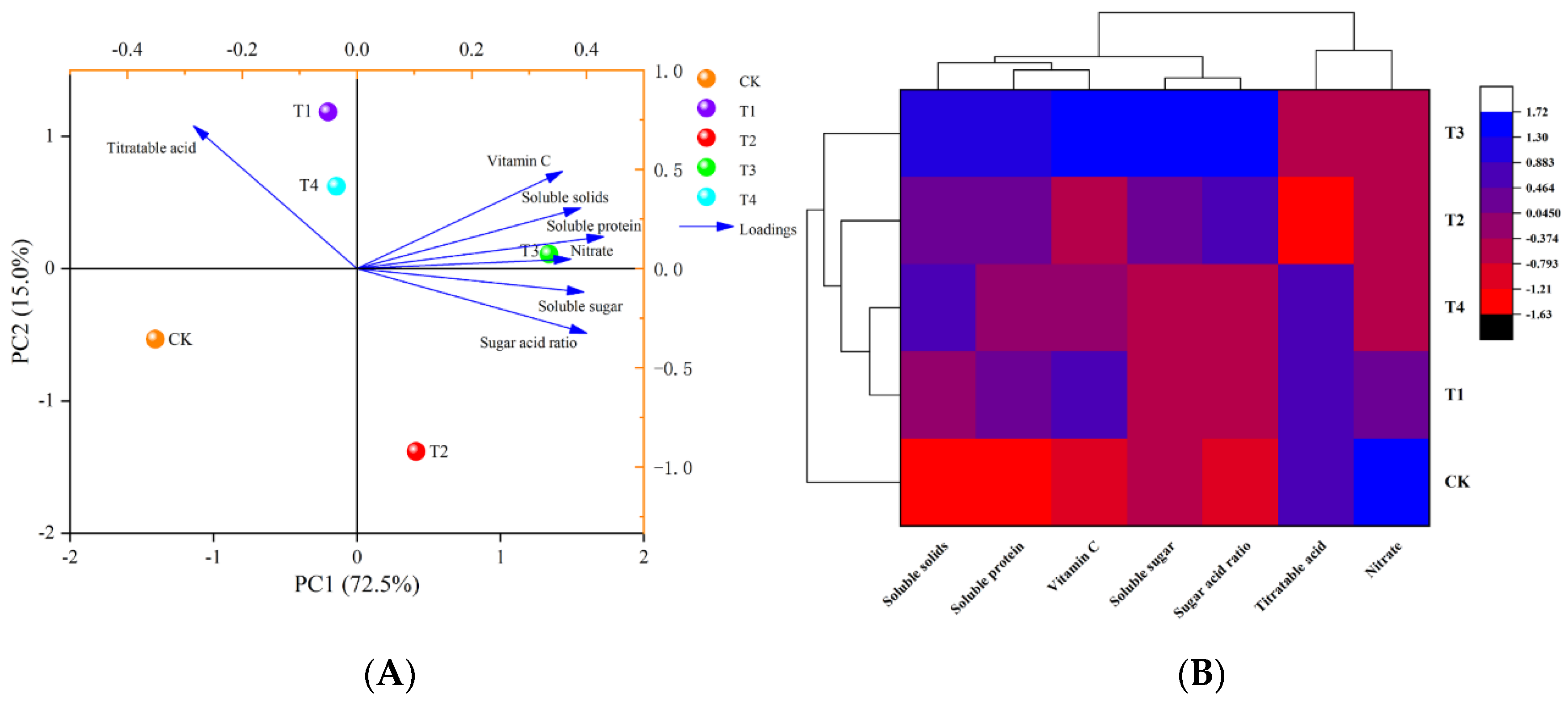
3.1.3. Principal Component Analysis and Cluster Analysis
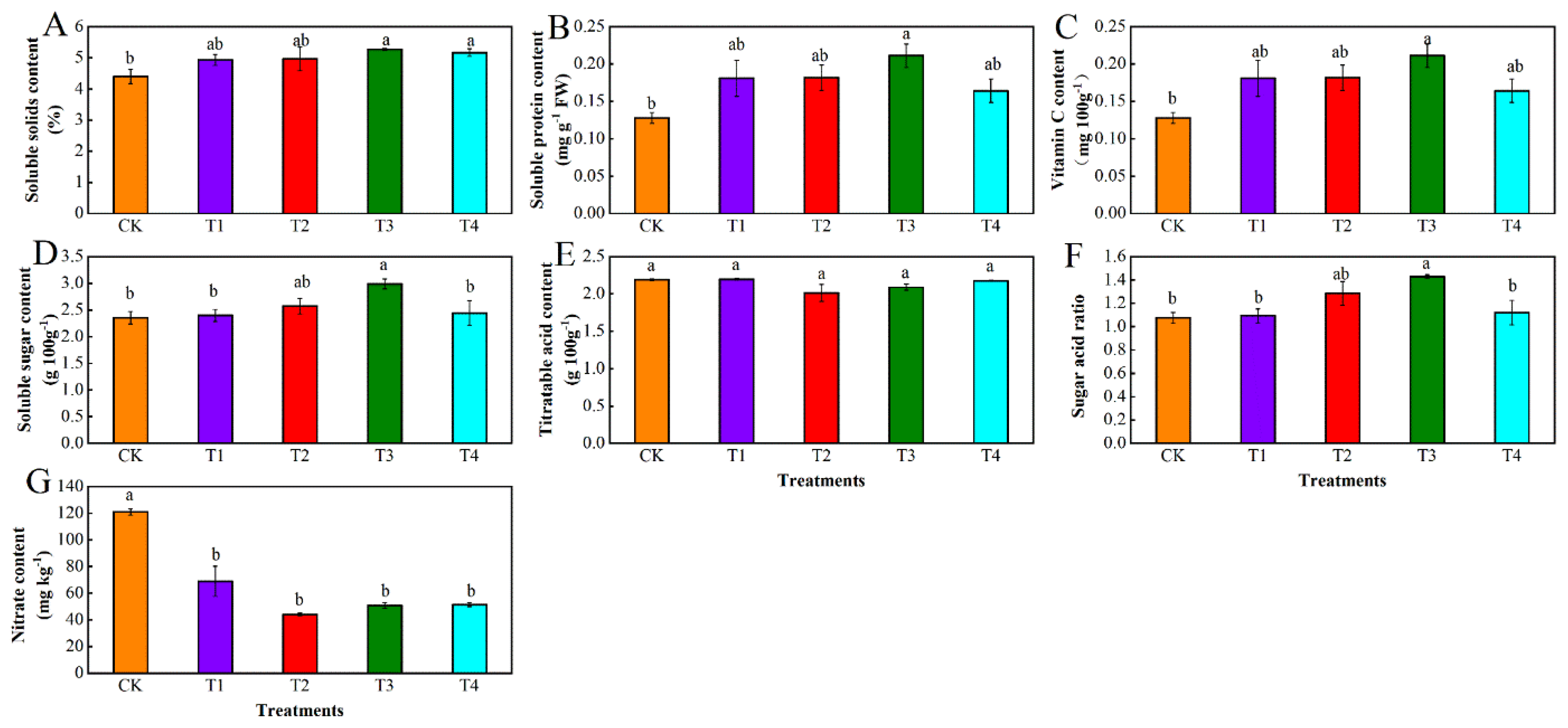
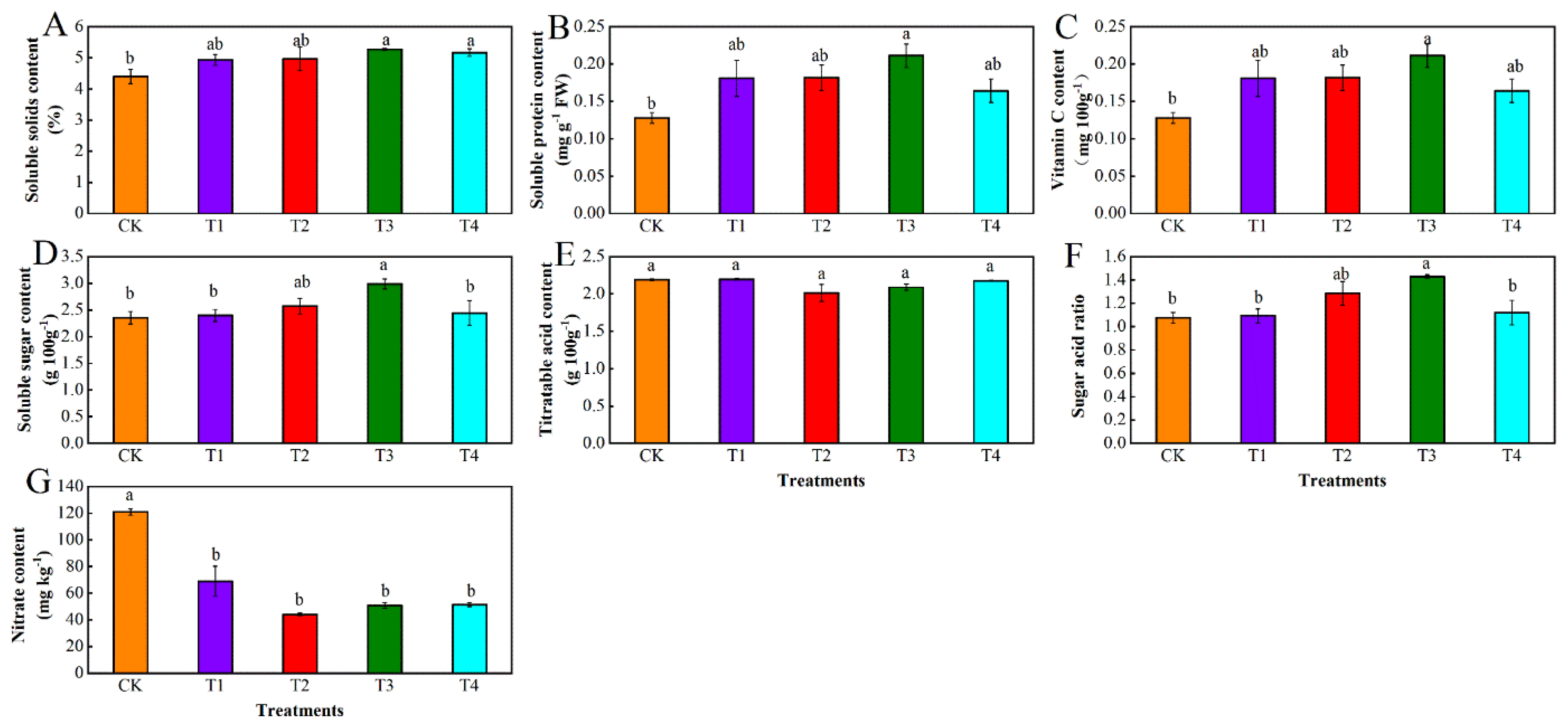
3.2. Effects of Different Levels of Water Deficit on Nutritional Quality in Tomato Fruit
3.2.1. Variance Analysis
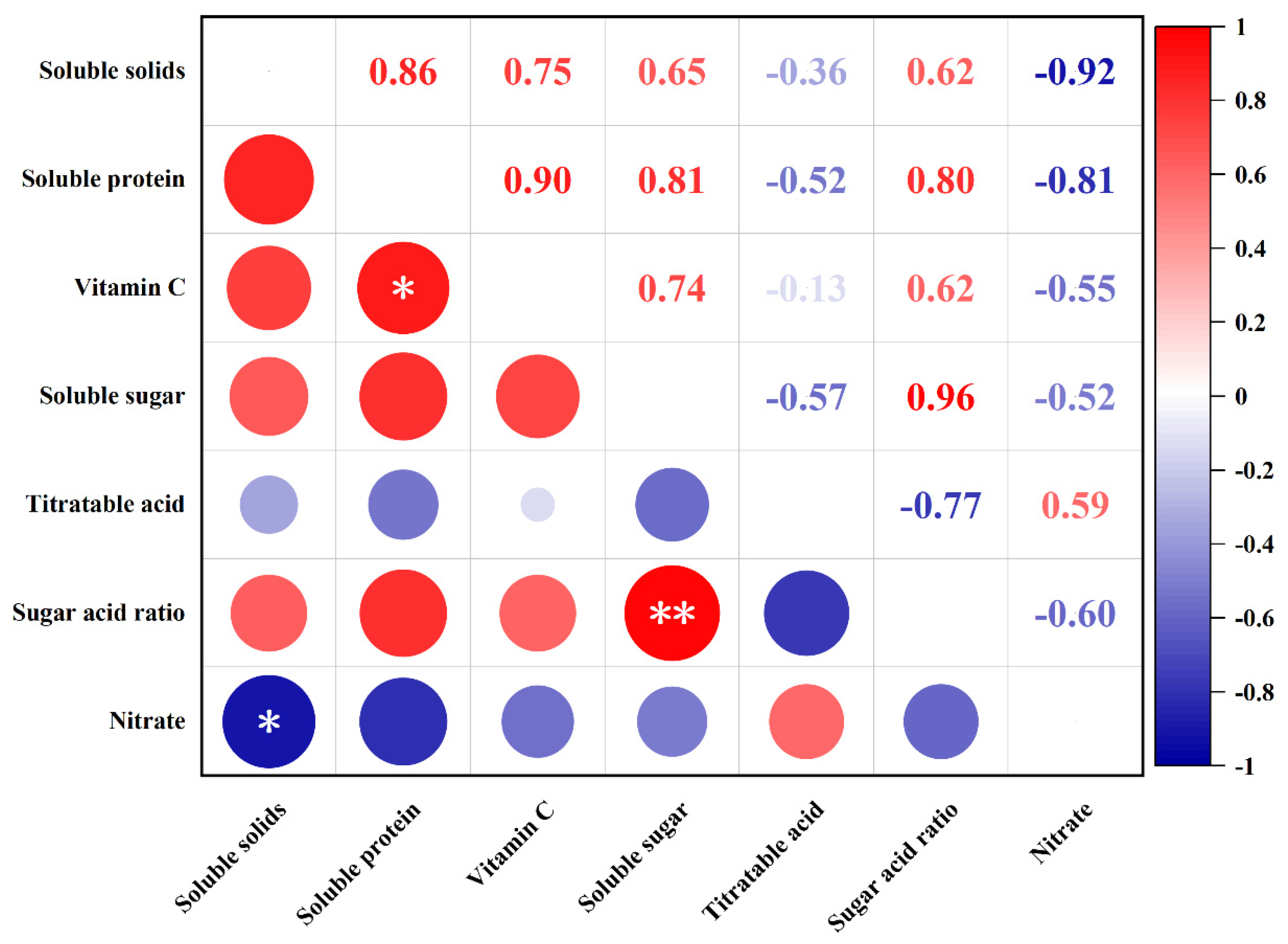
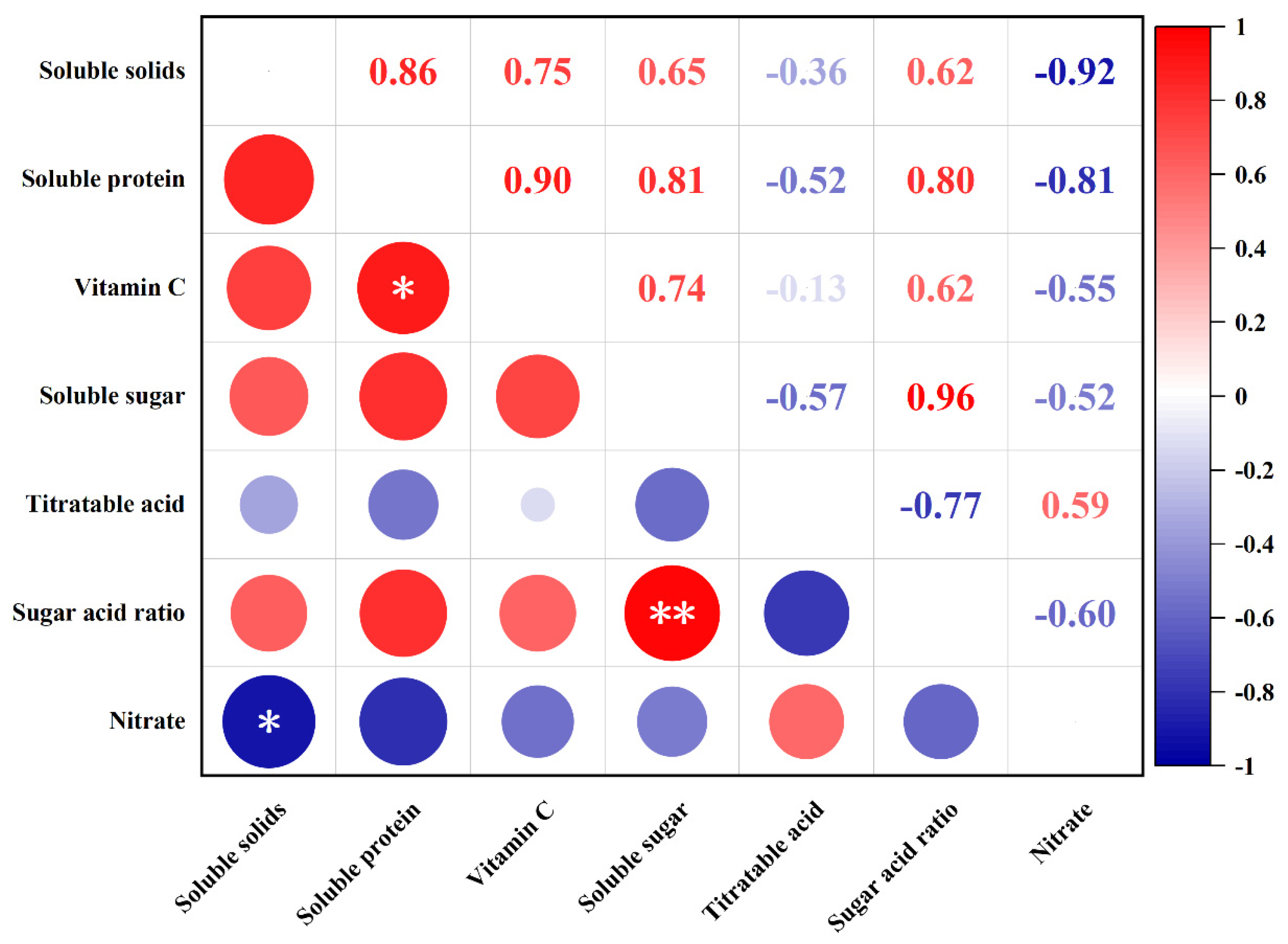
3.2.2. Correlation Analysis
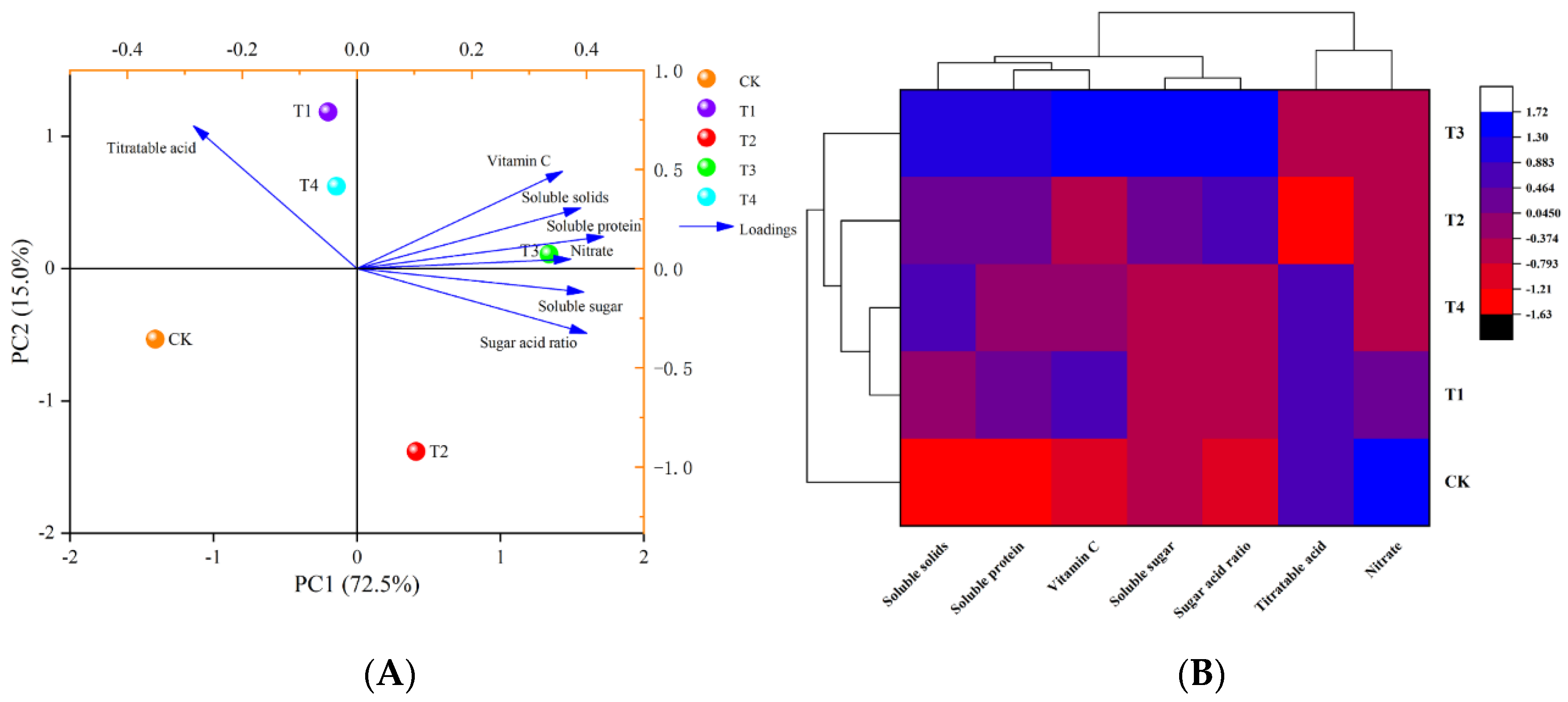
3.2.3. Principal Component Analysis and Cluster Analysis
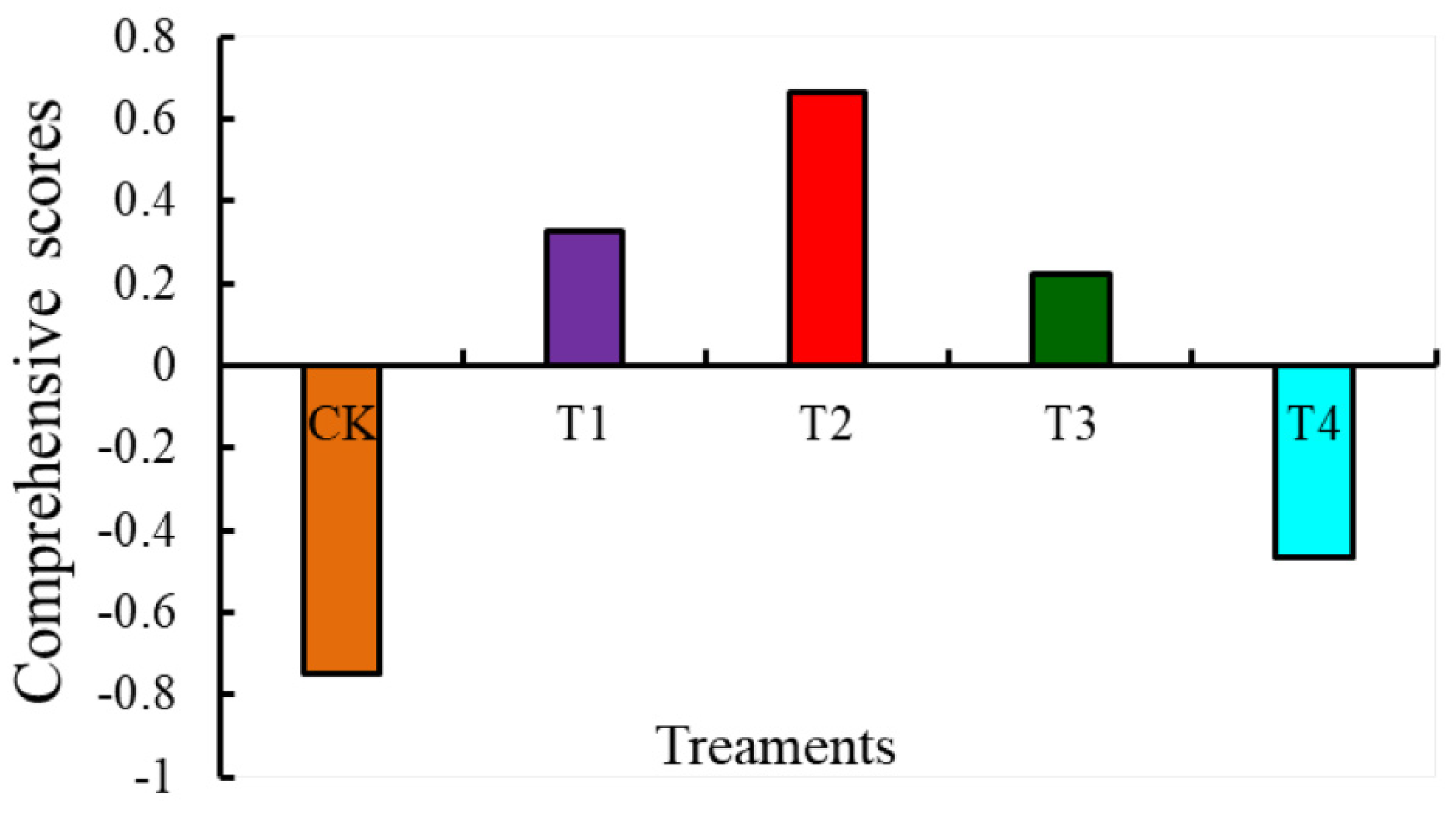
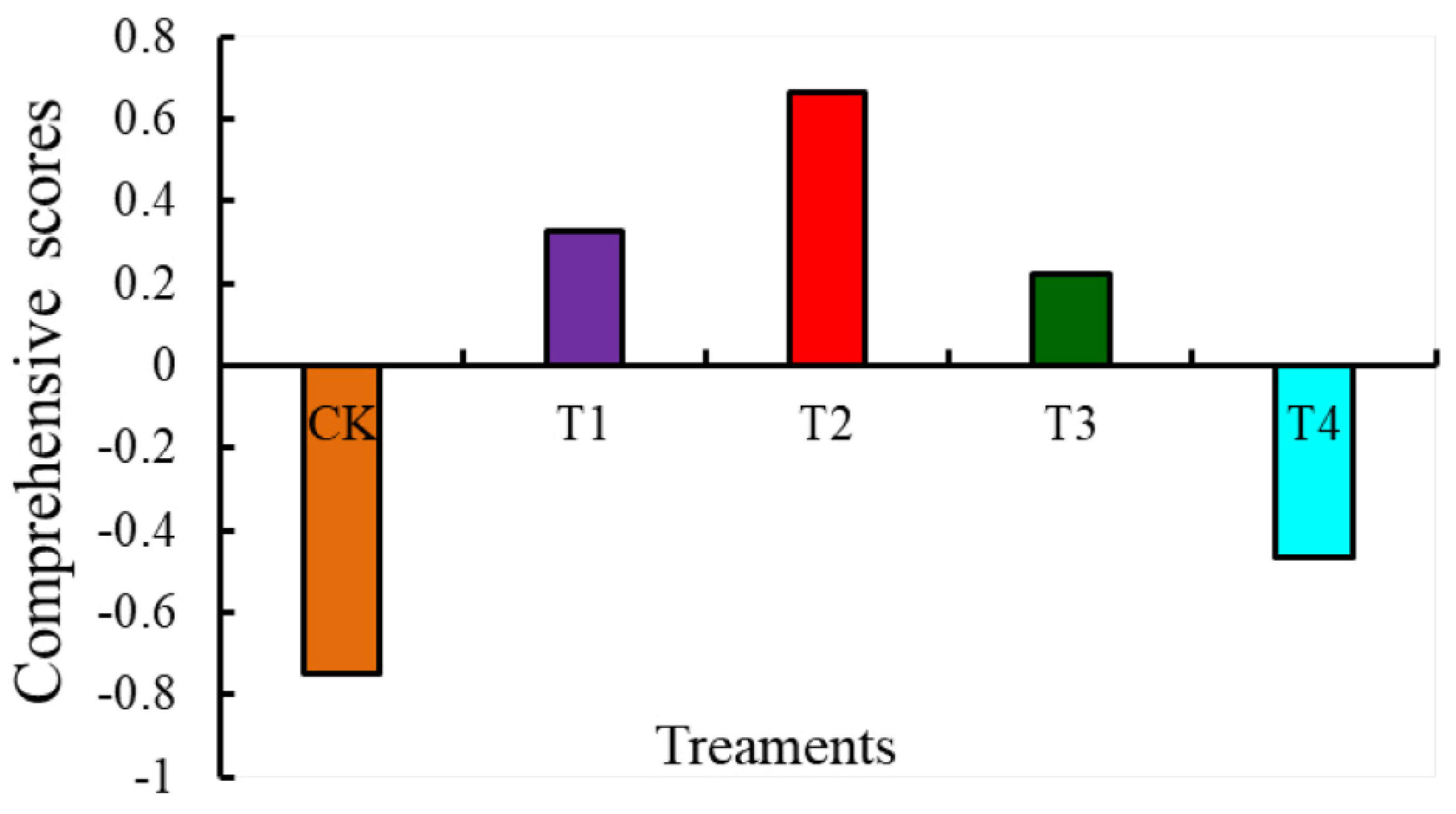
3.3. Evaluation of the Effects of Different Levels of Water-Deficit Treatments on Tomato Fruit Quality and Antioxidant Capacity Based on PCA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lahoz, I.; Pérez-de-Castro, A.; Valcárcel, M.; Macua, J.I.; Beltran, J.; Roselló, S.; Cebolla-Cornejo, J. Effect of water deficit on the agronomical performance and quality of processing tomato. Sci. Hortic. 2016, 200, 55–65. [Google Scholar]
- Mesquitaa, M.; Machadob, A.L.; dos Santosb, A.P.; da Silvac, M.V.; de Oliveirab, H.F.; Battistia, R.; Nascimentoa, A.R. Assessing the effects of deficit irrigation techniques on yield and water productivity of processing tomato. Chem. Eng. 2019, 75, 181–185. [Google Scholar]
- Szabo, K.; Diaconeasa, Z.; Cătoi, A.-F.; Vodnar, D.C. Screening of ten tomato varieties processing waste for bioactive components and their related antioxidant and antimicrobial activities. Antioxidants 2019, 8, 292. [Google Scholar]
- Añibarro-Ortega, M.; Pinela, J.; Ćirić, A.; Martins, V.; Rocha, F.; Soković, M.D.; Barata, A.M.; Carvalho, A.M.; Barros, L.; Ferreira, I.C. Valorisation of table tomato crop by-products: Phenolic profiles and in vitro antioxidant and antimicrobial activities. Food Bioprod. Process. 2020, 124, 307–319. [Google Scholar]
- Laranjeira, T.; Costa, A.; Faria-Silva, C.; Ribeiro, D.; de Oliveira, J.M.P.F.; Simões, S.; Ascenso, A. Sustainable valorization of tomato by-products to obtain bioactive compounds: Their potential in inflammation and cancer management. Molecules 2022, 27, 1701. [Google Scholar]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar]
- Vinha, A.F.; Alves, R.C.; Barreira, S.V.; Castro, A.; Costa, A.S.; Oliveira, M.B.P. Effect of peel and seed removal on the nutritional value and antioxidant activity of tomato (Lycopersicon esculentum L.) fruits. LWT-Food Sci. Technol. 2014, 55, 197–202. [Google Scholar]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting free radicals in oxidative stress-related human diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar]
- Toor, R.K.; Savage, G.P.; Lister, C.E. Release of antioxidant components from tomatoes determined by an in vitro digestion method. Int. J. Food Sci. Nutr. 2009, 60, 119–129. [Google Scholar]
- Chandra, H.M.; Ramalingam, S. Antioxidant potentials of skin, pulp, and seed fractions of commercially important tomato cultivars. Food Sci. Biotechnol. 2011, 20, 15–21. [Google Scholar]
- García-Flores, L.A.; Medina, S.; Gómez, C.; Wheelock, C.E.; Cejuela, R.; Martínez-Sanz, J.M.; Oger, C.; Galano, J.-M.; Durand, T.; Hernández-Sáez, Á. Aronia–citrus juice (polyphenol-rich juice) intake and elite triathlon training: A lipidomic approach using representative oxylipins in urine. Food Funct. 2018, 9, 463–475. [Google Scholar]
- Frei, B.; Birlouez-Aragon, I.; Lykkesfeldt, J. Authors’ perspective: What is the optimum intake of vitamin C in humans? Crit. Rev. Food Sci. Nutr. 2012, 52, 815–829. [Google Scholar]
- Kim, H.-J.; Yang, T.; Choi, S.; Wang, Y.-J.; Lin, M.-Y.; Liceaga, A.M. Supplemental intracanopy far-red radiation to red LED light improves fruit quality attributes of greenhouse tomatoes. Sci. Hortic. 2020, 261, 108985. [Google Scholar]
- Shahryari, R.; Mollasadeghi, V. Correlation study of some traits affecting yield and yield components of wheat genotypes in terms of normal irrigation and end drought stress. Adv. Environ. Biol. 2011, 5, 523–552. [Google Scholar]
- Besbes, M.; Chahed, J.; Hamdane, A. Water Security, Food Security and the National Water Dependency. In National Water Security; Springer: Berlin/Heidelberg, Germany, 2019; pp. 219–255. [Google Scholar]
- Farouk, S.; El-Metwally, I. Synergistic responses of drip-irrigated wheat crop to chitosan and/or silicon under different irrigation regimes. Agric. Water Manag. 2019, 226, 105807. [Google Scholar]
- Yin, J.; Gentine, P.; Zhou, S.; Sullivan, S.C.; Wang, R.; Zhang, Y.; Guo, S. Large increase in global storm runoff extremes driven by climate and anthropogenic changes. Nat. Commun. 2018, 9, 4389. [Google Scholar]
- Ali, S.; Liu, Y.; Ishaq, M.; Shah, T.; Ilyas, A.; Din, I.U. Climate change and its impact on the yield of major food crops: Evidence from Pakistan. Foods 2017, 6, 39. [Google Scholar]
- Keabetswe, L.; Shao, G.C.; Cui, J.; Lu, J.; Stimela, T. A combination of biochar and regulated deficit irrigation improves tomato fruit quality: A comprehensive quality analysis. Folia Hortic. 2019, 31, 181–193. [Google Scholar]
- Ripoll, J.; Urban, L.; Staudt, M.; Lopez-Lauri, F.; Bidel, L.P.; Bertin, N. Water shortage and quality of fleshy fruits—Making the most of the unavoidable. J. Exp. Bot. 2014, 65, 4097–4117. [Google Scholar]
- Pérez-Álvarez, E.P.; Intrigliolo, D.S.; Almajano, M.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Effects of water deficit irrigation on phenolic composition and antioxidant activity of Monastrell grapes under semiarid conditions. Antioxidants 2021, 10, 1301. [Google Scholar]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The roles of mitochondrial reactive oxygen species in cellular signaling and stress response in plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin. Molecules 2018, 23, 535. [Google Scholar]
- Xoconostle-Cazares, B.; Ramirez-Ortega, F.A.; Flores-Elenes, L.; Ruiz-Medrano, R. Drought tolerance in crop plants. J. Plant Physiol. 2010, 5, 241–256. [Google Scholar]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar]
- Lopez, G.; Echeverria, G.; Bellvert, J.; Mata, M.; Behboudian, M.H.; Girona, J.; Marsal, J. Water stress for a short period before harvest in nectarine: Yield, fruit composition, sensory quality, and consumer acceptance of fruit. Sci. Hortic. 2016, 211, 1–7. [Google Scholar]
- Yang, B.; Yao, H.; Zhang, J.; Li, Y.; Ju, Y.; Zhao, X.; Sun, X.; Fang, Y. Effect of regulated deficit irrigation on the content of soluble sugars, organic acids and endogenous hormones in Cabernet Sauvignon in the Ningxia region of China. Food Chem. 2020, 312, 126020. [Google Scholar]
- Cáceres-Mella, A.; Talaverano, M.I.; Villalobos-González, L.; Ribalta-Pizarro, C.; Pastenes, C. Controlled water deficit during ripening affects proanthocyanidin synthesis, concentration and composition in Cabernet Sauvignon grape skins. Plant Physiol. Biochem. 2017, 117, 34–41. [Google Scholar]
- Nie, Z.; Wan, C.; Chen, C.; Chen, J. Comprehensive evaluation of the postharvest antioxidant capacity of Majiayou pomelo harvested at different maturities based on PCA. Antioxidants 2019, 8, 136. [Google Scholar]
- Devarajan, R.; Jayaraman, J.K.; Somasundaram, S.M.; Ragupathy, S.; Raman, P.; Sathiamoorthy, K.; Subbaraya, U. Genetic diversity in fresh fruit pulp mineral profile of 100 Indian Musa accessions. Food Chem. 2021, 361, 130080. [Google Scholar]
- Zhang, Z.; Mao, C.; Shi, Z.; Kou, X. The amino acid metabolic and carbohydrate metabolic pathway play important roles during salt-stress response in tomato. Front. Plant Sci. 2017, 8, 1231. [Google Scholar]
- Liu, M.; Yu, H.; Zhao, G.; Huang, Q.; Lu, Y.; Ouyang, B. Profiling of drought-responsive microRNA and mRNA in tomato using high-throughput sequencing. BMC Genom. 2017, 18, 481. [Google Scholar]
- Tian, S.-L.; Lu, B.-Y.; Gong, Z.-H.; Shah, S.N.M. Effects of drought stress on capsanthin during fruit development and ripening in pepper (Capsicum annuum L.). Agric. Water Manag. 2014, 137, 46–51. [Google Scholar]
- Saltveit, M.E. Effect of ethylene on quality of fresh fruits and vegetables. Postharvest Biol. Technol. 1999, 15, 279–292. [Google Scholar]
- Arya, S.; Mahajan, M.; Jain, P. Non-spectrophotometric methods for the determination of Vitamin C. Anal. Chim. Acta 2000, 417, 1–14. [Google Scholar]
- Wang, J.; Zhang, J.; Li, J.; Dawuda, M.M.; Ali, B.; Wu, Y.; Yu, J.; Tang, Z.; Lyu, J.; Xiao, X. Exogenous application of 5-aminolevulinic acid promotes coloration and improves the quality of tomato fruit by regulating carotenoid metabolism. Front. Plant Sci. 2021, 12, 683868. [Google Scholar]
- Wang, Y.; Wang, Y.; Ji, K.; Dai, S.; Hu, Y.; Sun, L.; Li, Q.; Chen, P.; Sun, Y.; Duan, C. The role of abscisic acid in regulating cucumber fruit development and ripening and its transcriptional regulation. Plant Physiol. Biochem. 2013, 64, 70–79. [Google Scholar]
- Cataldo, D.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar]
- Cilla, A.; Alegria, A.; Attanzio, A.; Garcia-Llatas, G.; Tesoriere, L.; Livrea, M.A. Dietary phytochemicals in the protection against oxysterol-induced damage. Chem. Phys. Lipids 2017, 207, 192–205. [Google Scholar]
- Lu, B.; Zhao, Y. Photooxidation of phytochemicals in food and control: A review. Ann. N. Y. Acad. Sci. 2017, 1398, 72–82. [Google Scholar]
- Ju, Y.-L.; Xu, G.-Q.; Yue, X.-F.; Zhao, X.-F.; Tu, T.-Y.; Zhang, J.-X.; Fang, Y.-L. Effects of regulated deficit irrigation on amino acid profiles and their derived volatile compounds in Cabernet Sauvignon (Vitis vinifera L.) grapes and wines. Molecules 2018, 23, 1983. [Google Scholar]
- Slimestad, R.; Verheul, M. Review of flavonoids and other phenolics from fruits of different tomato (Lycopersicon esculentum Mill.) cultivars. J. Sci. Food Agric. 2009, 89, 1255–1270. [Google Scholar]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic profiling of the host response of tomato (Solanum lycopersicum) following infection by Ralstonia solanacearum. Int. J. Mol. Sci. 2019, 20, 3945. [Google Scholar]
- Leng, F.; Zhou, J.; Wang, C.; Sun, L.; Zhang, Y.; Li, Y.; Wang, L.; Wang, S.; Zhang, X.; Xie, Z. Post-veraison different frequencies of water deficit strategies enhance Reliance grapes quality under root restriction. Food Chem. 2022, 390, 133181. [Google Scholar]
- Sokół-Łętowska, A.; Oszmiański, J.; Wojdyło, A. Antioxidant activity of the phenolic compounds of hawthorn, pine and skullcap. Food Chem. 2007, 103, 853–859. [Google Scholar]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar]
- Popović, B.; Štajner, D.; Ždero-Pavlović, R.; Tumbas-Šaponjac, V.; Čanadanović-Brunet, J.; Orlović, S. Water stress induces changes in polyphenol profile and antioxidant capacity in poplar plants (Populus spp.). Plant Physiol. Biochem. 2016, 105, 242–250. [Google Scholar]
- Tahmaz, H.; Yüksel Küskü, D.; Söylemezoğlu, G. Phenolic compounds and antioxidant capacities in grape berry skin, seed and stems of six wine grape varieties grown in Turkey. Acta Sci. Pol. Hortorum Cultus 2021, 20, 15–25. [Google Scholar]
- Dorai, M.; Papadopoulos, A.; Gosselin, A. Influence of electric conductivity management on greenhouse tomato yield and fruit quality. Agronomie 2001, 21, 367–383. [Google Scholar]
- Ripoll, J.; Urban, L.; Brunel, B.; Bertin, N. Water deficit effects on tomato quality depend on fruit developmental stage and genotype. J. Plant Physiol. 2016, 190, 26–35. [Google Scholar]
- Terry, L.A.; Chope, G.A.; Bordonaba, J.G. Effect of water deficit irrigation and inoculation with Botrytis cinerea on strawberry (Fragaria x ananassa) fruit quality. J. Agric. Food Chem. 2007, 55, 10812–10819. [Google Scholar]
- Davies, J.N.; Hobson, G.E.; McGlasson, W. The constituents of tomato fruit—The influence of environment, nutrition, and genotype. Crit. Rev. Food Sci. Nutr. 1981, 15, 205–280. [Google Scholar]
- Ripoll, J.; Urban, L.; Bertin, N. The potential of the MAGIC TOM parental accessions to explore the genetic variability in tomato acclimation to repeated cycles of water deficit and recovery. Front. Plant Sci. 2016, 6, 1172. [Google Scholar]
- Gao, S.; Wang, Y.; Yu, S.; Huang, Y.; Liu, H.; Chen, W.; He, X. Effects of drought stress on growth, physiology and secondary metabolites of Two Adonis species in Northeast China. Sci. Hortic. 2020, 259, 108795. [Google Scholar]
- Cheng, L.; Han, M.; Yang, L.-m.; Li, Y.; Sun, Z.; Zhang, T. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crops Prod. 2018, 122, 473–482. [Google Scholar]
- Ju, Y.-l.; Yue, X.-f.; Zhao, X.-f.; Zhao, H.; Fang, Y.-L. Physiological, micro-morphological and metabolomic analysis of grapevine (Vitis vinifera L.) leaf of plants under water stress. Plant Physiol. Biochem. 2018, 130, 501–510. [Google Scholar]
- Wang, X.; Xing, Y. Evaluation of the effects of irrigation and fertilization on tomato fruit yield and quality: A principal component analysis. Sci. Rep. 2017, 7, 350. [Google Scholar]
- Massot, C.; Génard, M.; Stevens, R.; Gautier, H. Fluctuations in sugar content are not determinant in explaining variations in vitamin C in tomato fruit. Plant Physiol. Biochem. 2010, 48, 751–757. [Google Scholar]
- Bertin, N.; Génard, M. Tomato quality as influenced by preharvest factors. Sci. Hortic. 2018, 233, 264–276. [Google Scholar]
- Patras, A.; Brunton, N.P.; Downey, G.; Rawson, A.; Warriner, K.; Gernigon, G. Application of principal component and hierarchical cluster analysis to classify fruits and vegetables commonly consumed in Ireland based on in vitro antioxidant activity. J. Food Compos. Anal. 2011, 24, 250–256. [Google Scholar]
- Hossain, M.B.; Patras, A.; Barry-Ryan, C.; Martin-Diana, A.B.; Brunton, N.P. Application of principal component and hierarchical cluster analysis to classify different spices based on in vitro antioxidant activity and individual polyphenolic antioxidant compounds. J. Funct. Foods 2011, 3, 179–189. [Google Scholar]
- Zhou, D.-C.; Zheng, G.; Jia, L.-Y.; He, X.; Zhang, C.-F.; Wang, C.-Z.; Yuan, C.-S. Comprehensive evaluation on anti-inflammatory and anti-angiogenic activities in vitro of fourteen flavonoids from Daphne Genkwa based on the combination of efficacy coefficient method and principal component analysis. J. Ethnopharmacol. 2021, 268, 113683. [Google Scholar]
- Jin, N.; Jin, L.; Luo, S.; Tang, Z.; Liu, Z.; Wei, S.; Liu, F.; Zhao, X.; Yu, J.; Zhong, Y. Comprehensive Evaluation of Amino Acids and Polyphenols in 69 Varieties of Green Cabbage (Brassica oleracea L. var. capitata L.) Based on Multivariate Statistical Analysis. Molecules 2021, 26, 5355. [Google Scholar]
- Cao, X.; Jiang, F.; Wang, X.; Zang, Y.; Wu, Z. Comprehensive evaluation and screening for chilling-tolerance in tomato lines at the seedling stage. Euphytica 2015, 205, 569–584. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treaments | CK | T1 | T2 | T3 | T4 | |
|---|---|---|---|---|---|---|
| Flavonoids | Rutin | 160.65 ± 25.66 b | 371.63 ± 37.97 a | 429.93 ± 40.94 a | 337.22± 9.41 a | 331.47 ± 22.47 a |
| Quercetin | 633.07 ± 18.24 ab | 716.21 ± 85.74 a | 764.43 ± 69.00 a | 674.87 ± 15.35 ab | 499.19 ± 41.65 b | |
| Naringenin | 144.66 ± 13.74 a | 166.06 ± 4.20 a | 163.39 ± 8.96 a | 152.52 ± 4.37 a | 150.09 ± 1.70 a | |
| Kaempferol | 1.53 ± 0.31 b | 2.33 ± 0.38 b | 2.58 ± 0.66 b | 2.16 ± 0.25 b | 3.89 ± 0.35 a | |
| Phenols | Protocatechuic acid | 8.69 ± 0.19 b | 33.66 ± 4.34 a | 37.45 ± 3.76 a | 44.96 ± 6.04 a | 41.95 ± 4.72 a |
| P-hydroxybenzoic acid | 18.95 ± 0.82 ab | 22.74 ± 1.28 a | 20.01 ± 3.05 a | 19.16 ± 1.65 ab | 13.78 ± 1.19 b | |
| Chlorogenic acid | 1195.52 ± 112.65 b | 1811.66 ± 60.22 a | 1871.26 ± 82.64 a | 1651.91 ± 58.10 a | 1087.77 ± 68.75 b | |
| Gallic acid | 31.25 ± 2.69 c | 50.08 ± 6.99 b | 88.64 ± 5.41 a | 63.83 ± 7.77 b | 28.09 ± 1.79 c | |
| P-coumaric acid | 0.52 ± 0.10 a | 0.33 ± 0.06 a | 0.54 ± 0.18 a | 0.50 ± 0.01 a | 0.31 ± 0.07 a | |
| Ferulic acid | 9.92 ± 0.24 a | 9.44 ± 3.12 a | 14.61 ± 4.82 a | 17.40 ± 1.04 a | 8.50 ± 1.72 a | |
| Benzoic acid | 165.74 ± 28.56 b | 302.62 ± 84.04 ab | 399.91 ± 43.28 a | 427.1 ± 28.53 a | 220.18 ± 23.42 b | |
| Cinnamic acid | 1.30 ± 0.22 c | 1.13 ± 0.04 c | 2.95 ± 0.70 a | 1.57 ± 0.31 bc | 2.54 ± 0.24 ab | |
| Gentisic acid | 16.87 ± 2.70 c | 53.32 ± 5.35 a | 62.23 ± 6.30 a | 33.63 ± 4.85 b | 30.86 ± 4.07 bc | |
| Caffeic acid | 0.53 ± 0.02 ab | 0.43 ± 0.02 b | 0.62 ± 0.02 a | 0.58 ± 0.09 ab | 0.53 ± 0.04 ab | |
| Cynarin | 3.92 ± 0.39 b | 7.15 ± 0.80 a | 5.76 ± 0.73 ab | 4.31 ± 0.83 b | 3.63 ± 0.60 b | |
| Sinapic acid | 2.07 ± 0.26 b | 1.54 ± 0.17 ab | 2.23 ± 0.40 ab | 2.15 ± 0.21 ab | 2.42 ± 0.08 a | |
| Component Number | Eigenvalues | % of Variance | Cumulative % |
|---|---|---|---|
| 1 | 14.59 | 50.30 | 50.30 |
| 2 | 6.81 | 23.49 | 73.78 |
| 3 | 4.63 | 15.96 | 89.75 |
| 4 | 2.97 | 10.25 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, N.; Jin, L.; Wang, S.; Meng, X.; Ma, X.; He, X.; Zhang, G.; Luo, S.; Lyu, J.; Yu, J. A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity. Antioxidants 2022, 11, 1585. https://doi.org/10.3390/antiox11081585
Jin N, Jin L, Wang S, Meng X, Ma X, He X, Zhang G, Luo S, Lyu J, Yu J. A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity. Antioxidants. 2022; 11(8):1585. https://doi.org/10.3390/antiox11081585
Chicago/Turabian StyleJin, Ning, Li Jin, Shuya Wang, Xin Meng, Xianglan Ma, Xianxia He, Guobing Zhang, Shilei Luo, Jian Lyu, and Jihua Yu. 2022. "A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity" Antioxidants 11, no. 8: 1585. https://doi.org/10.3390/antiox11081585
APA StyleJin, N., Jin, L., Wang, S., Meng, X., Ma, X., He, X., Zhang, G., Luo, S., Lyu, J., & Yu, J. (2022). A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity. Antioxidants, 11(8), 1585. https://doi.org/10.3390/antiox11081585