
Antioxidant and Anti-Inflammatory Effects of Bischofia javanica (Blume) Leaf Methanol Extracts through the Regulation of Nrf2 and TAK1

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparation of MBJ
2.2. Antibodies
2.3. High-Performance Liquid Chromatography (HPLC) Analysis
2.4. Cell Culture and Reagents
2.5. Cell Viability Assay
2.6. Nitrite Assay
2.7. Reverse Transcription-Polymerase Chain Reactions (RT-PCR)
2.8. Reverse Transcription-Quantitative Polymerase Chain Reactions (RT-qPCR)
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Luciferase Reporter Assay
2.11. Preparation of Total Cell Lysates
2.12. Immunoblotting Analysis
2.13. Animal Testing
2.14. Radical Scavenging (DPPH) Assay
2.15. Measurement of Serum Aspartate Transaminase (AST), Alanine Transaminase (ALT), and Blood Glutathione (GSH)/Glutathione Disulfide (GSSG) Ratio in an Acute Hepatitis Mouse Model
2.16. Statistical Analyses and Experimental Replicates
3. Results
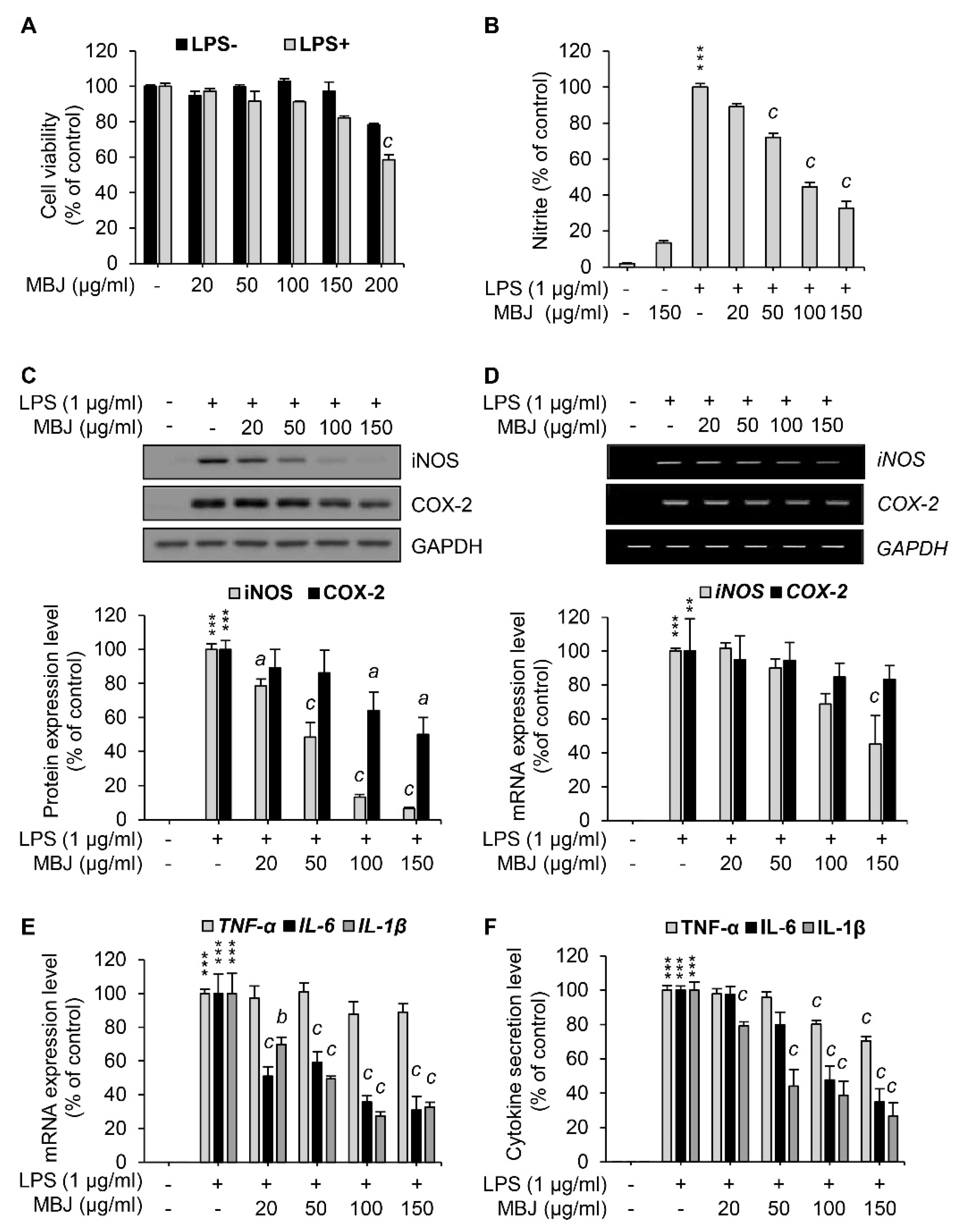
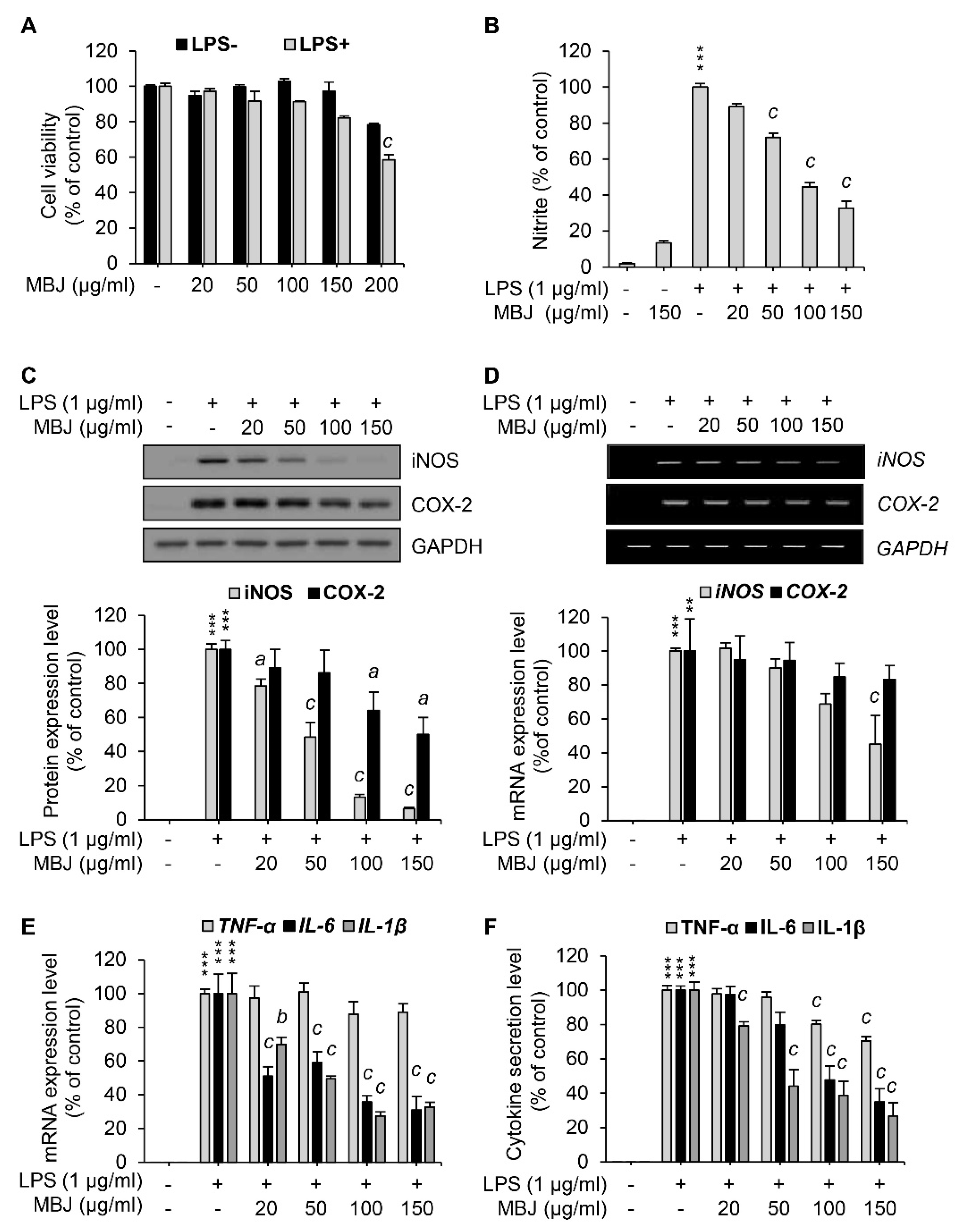
3.1. Suppression of NO Production by MBJ in Macrophages
3.2. Inhibitory Effects of MBJ on the Production of Inflammatory Mediators
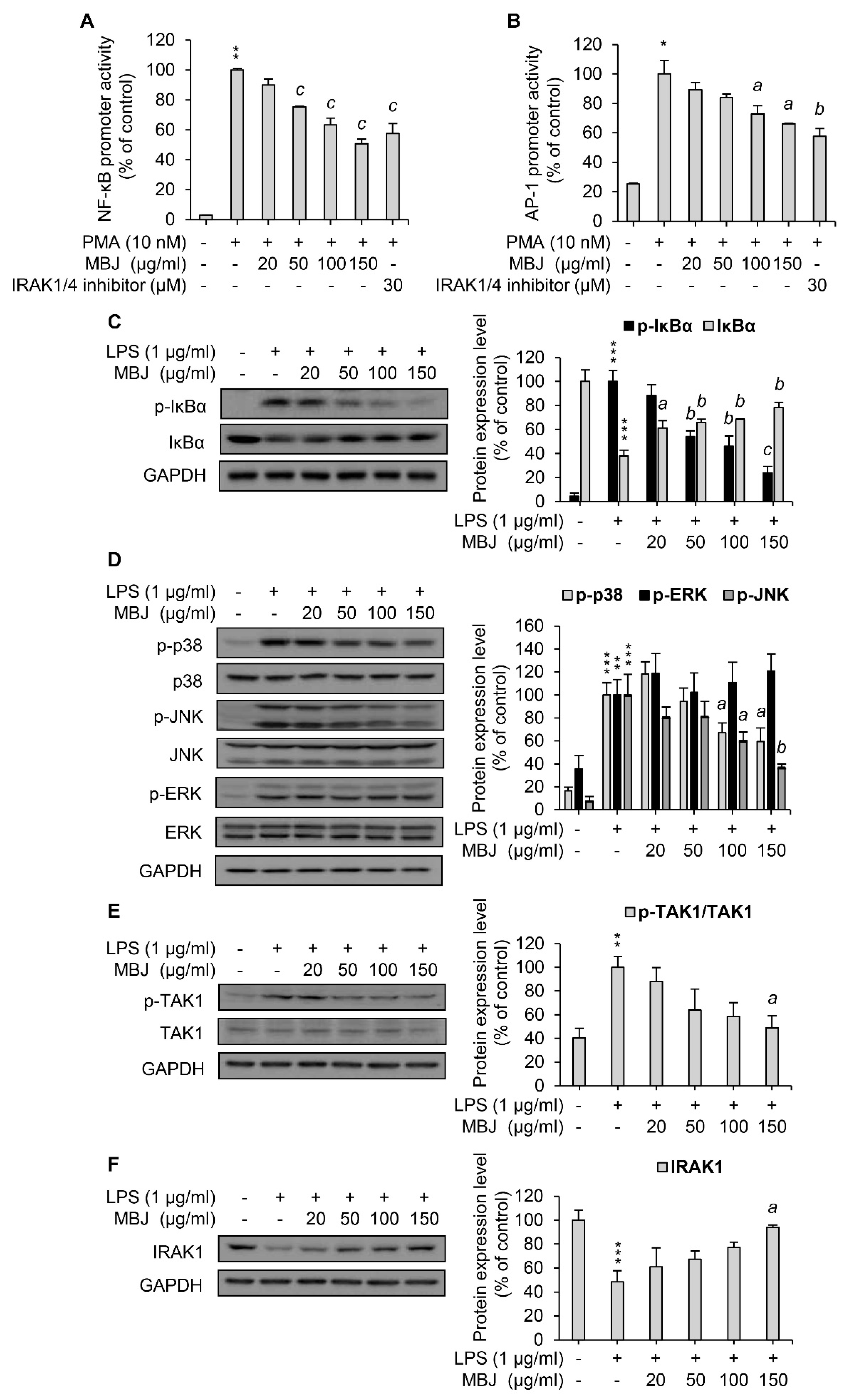
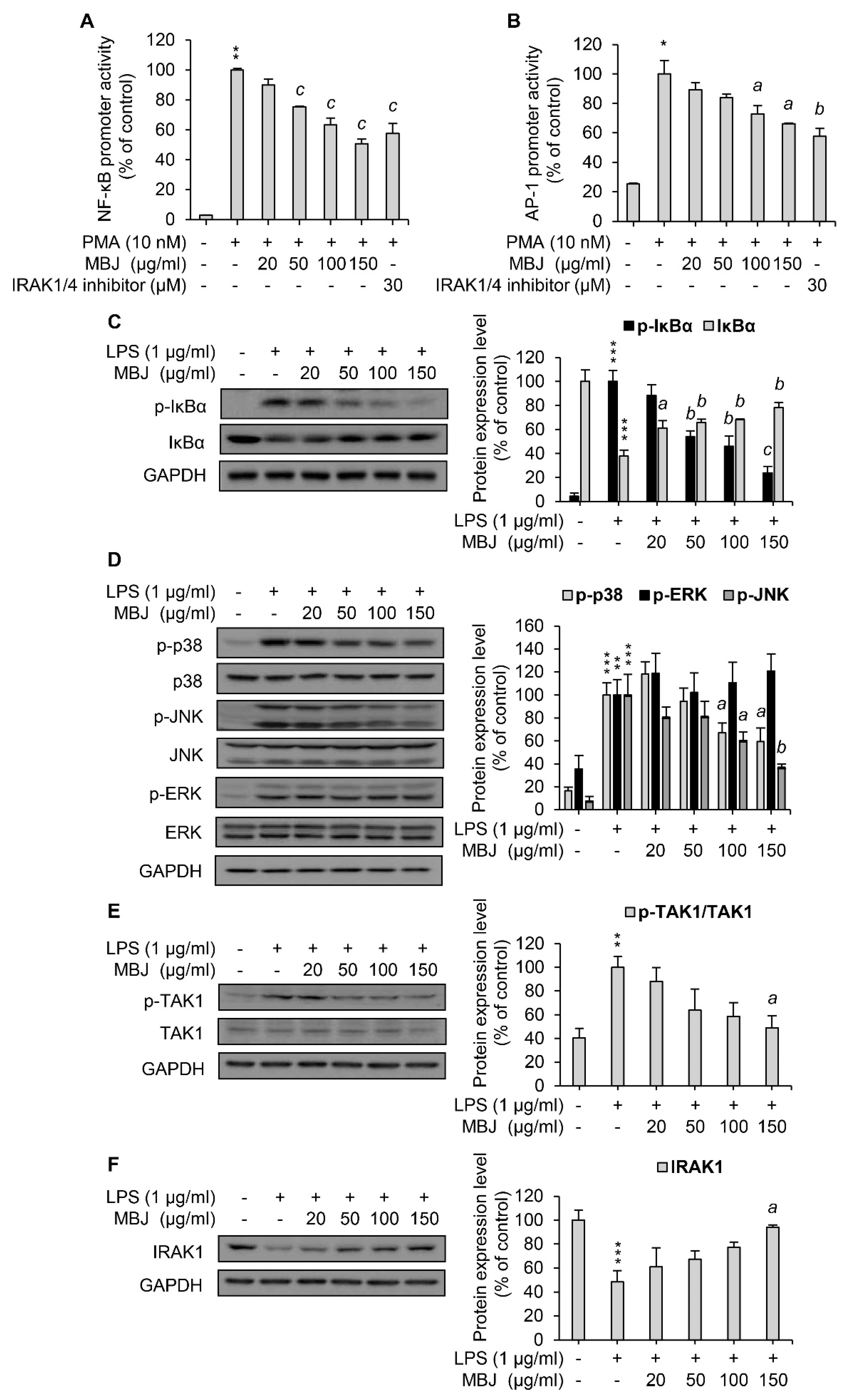
3.3. Regulation of MAPK Phosphorylation and NF-κB Activation by MBJ
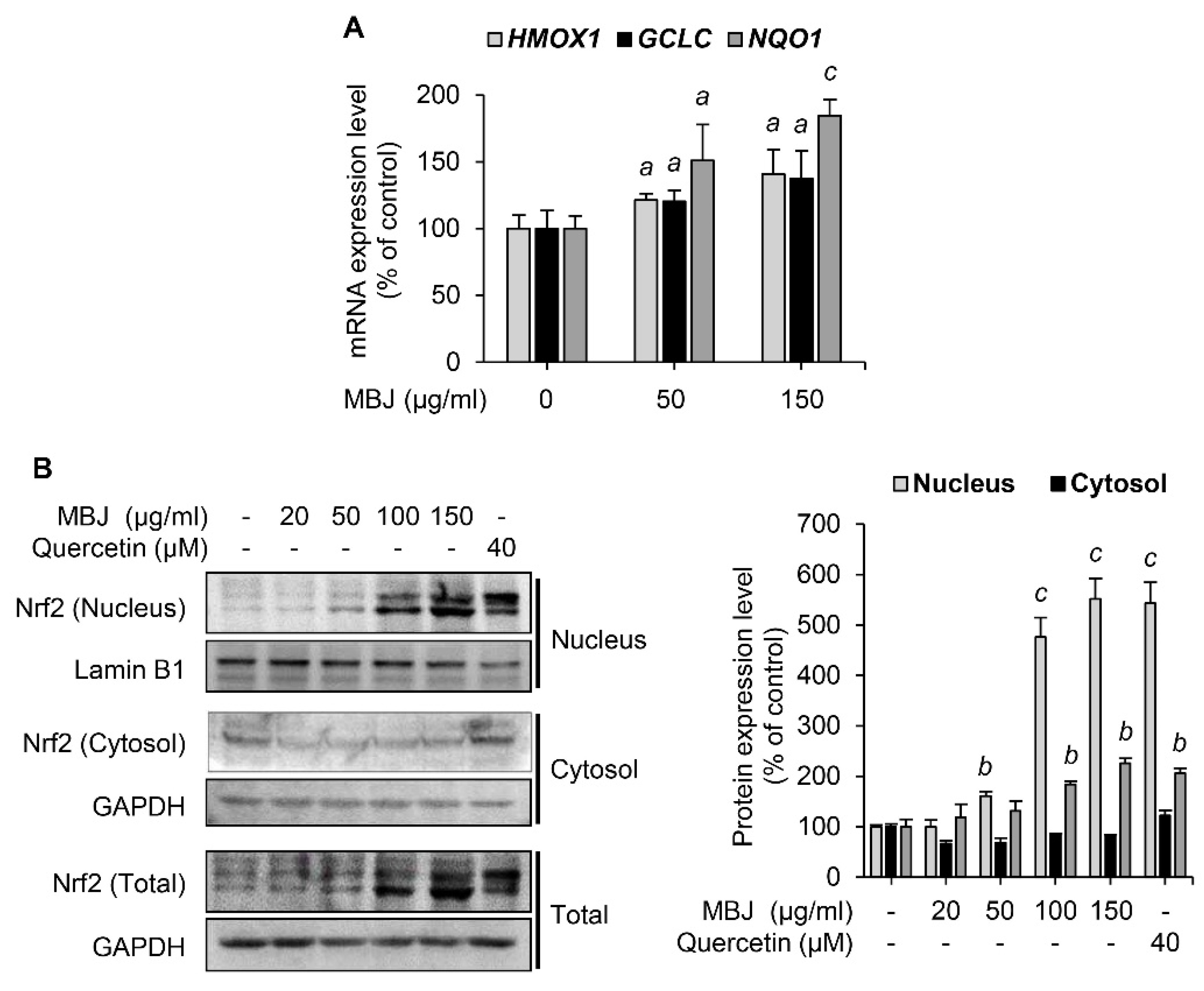
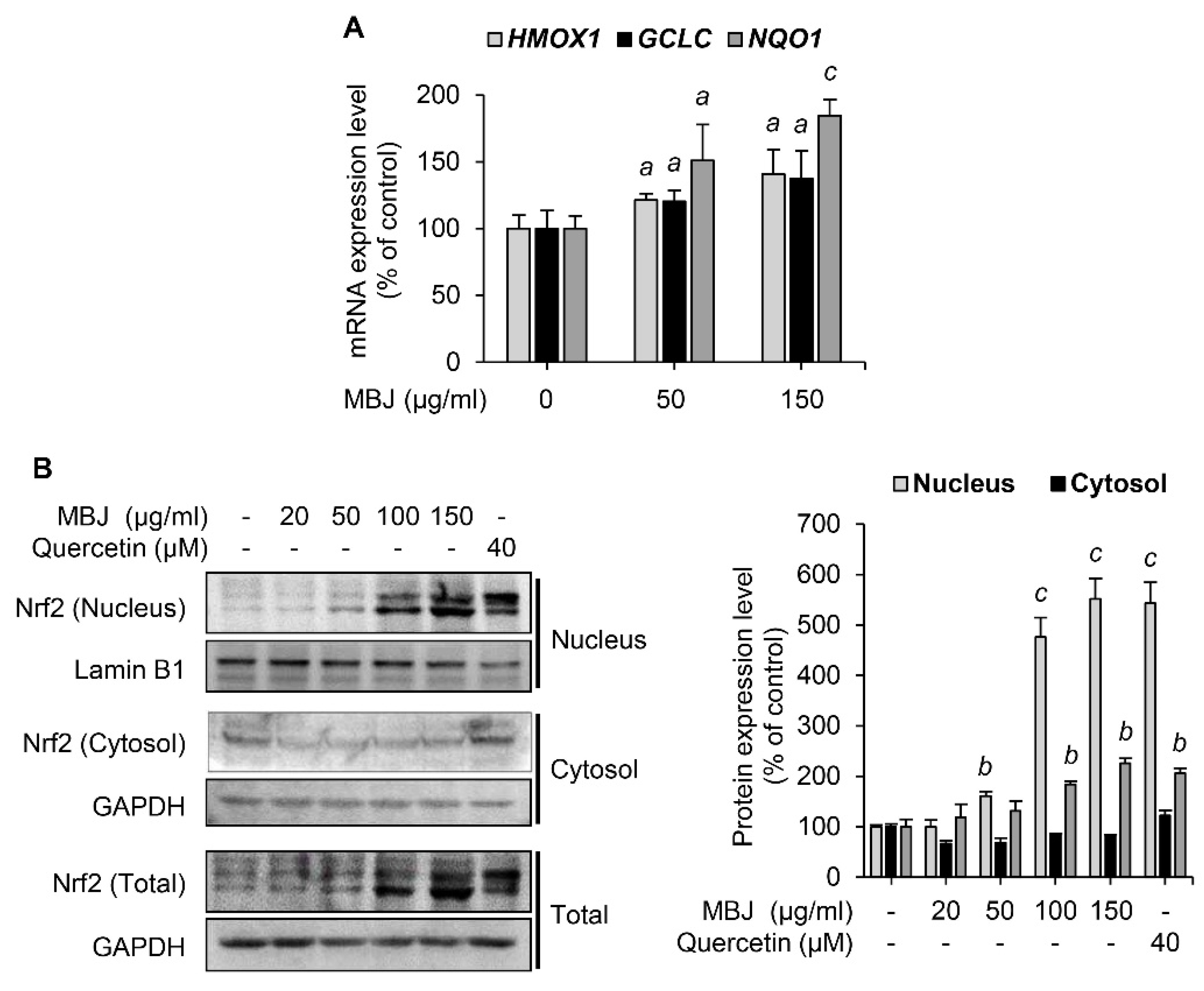
3.4. Effects of MBJ on Nrf2 Expression and Nuclear Translocation
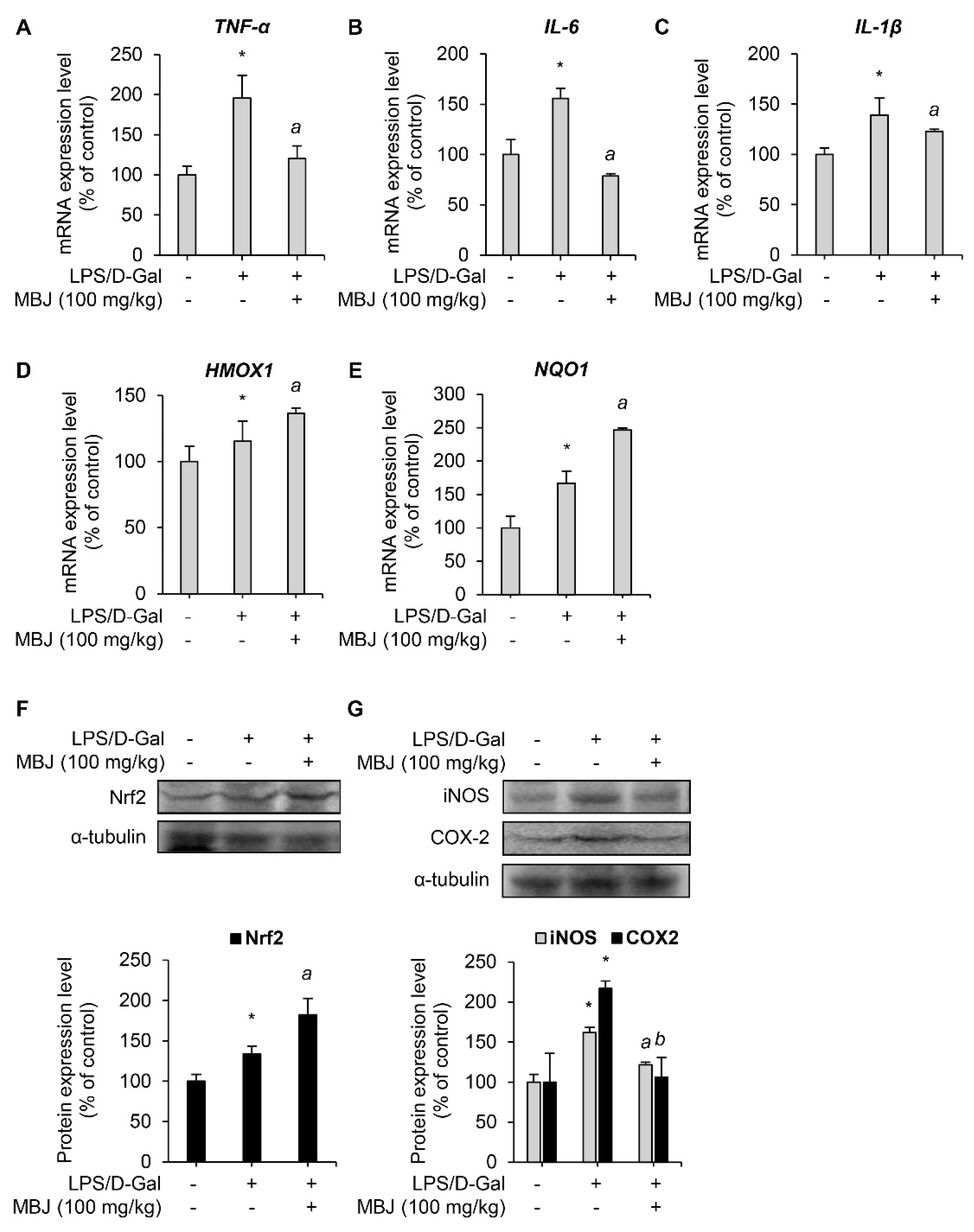
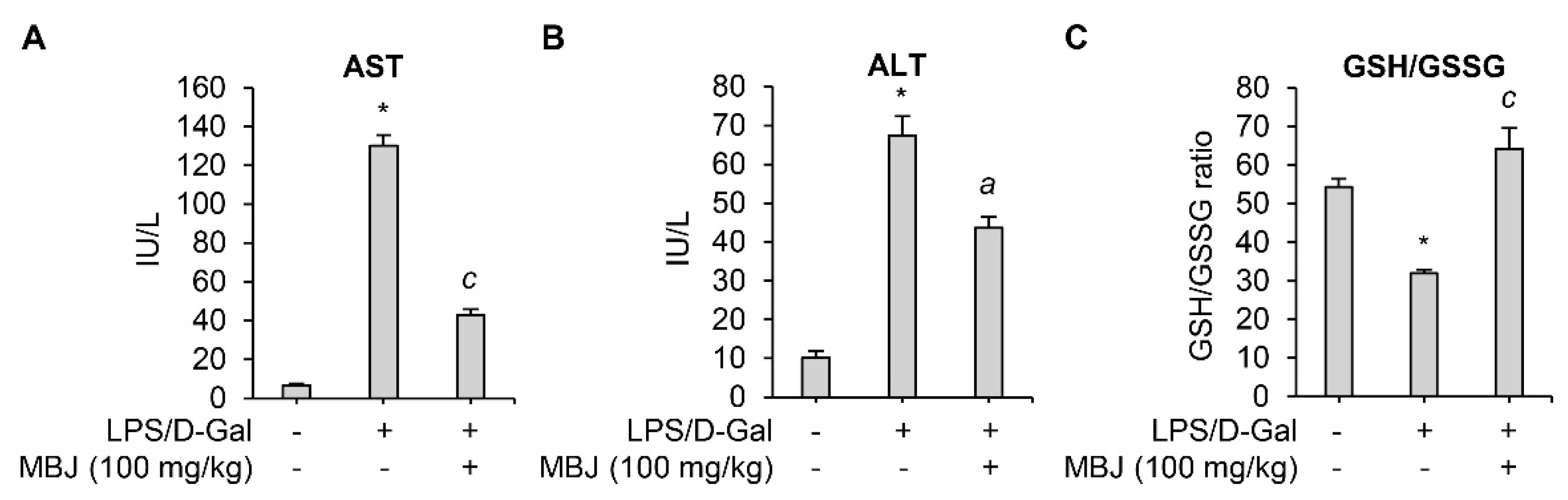
3.5. Antioxidant and Anti-Inflammatory Effects of MBJ In Vivo
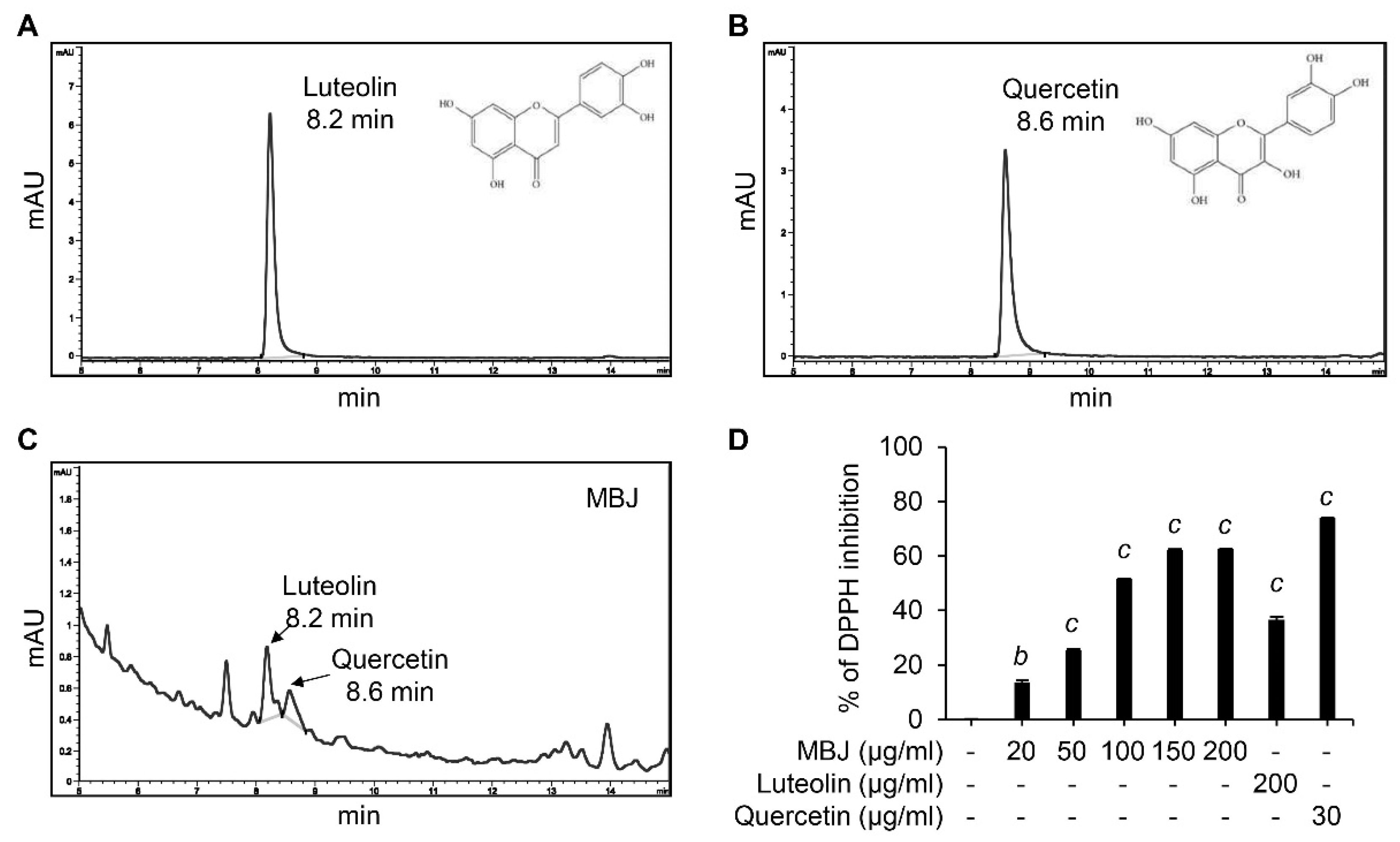
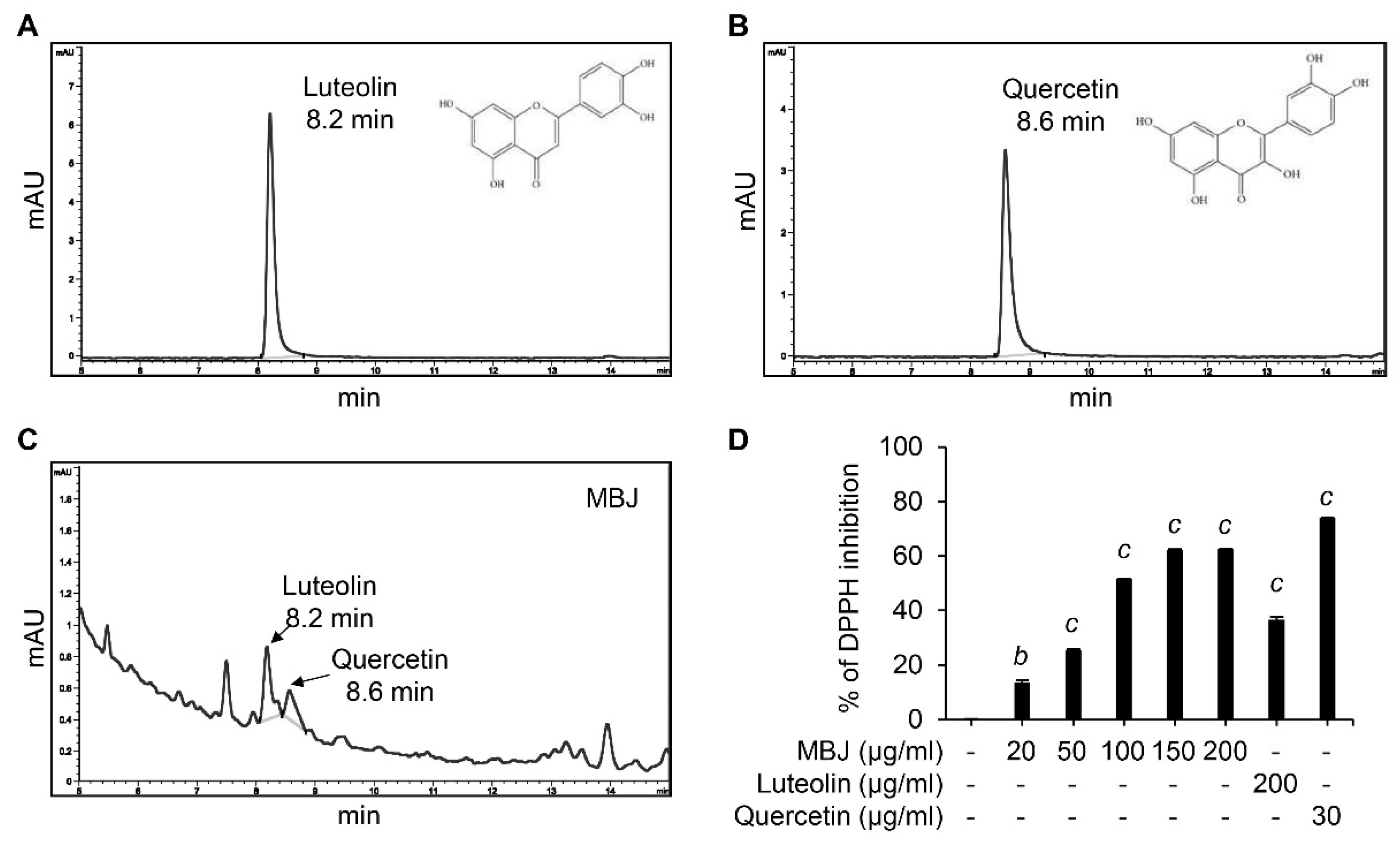
3.6. HPLC and DPPH Analyses of MBJ
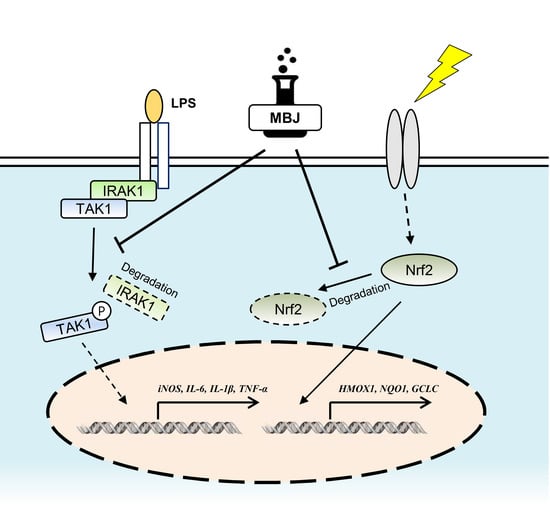
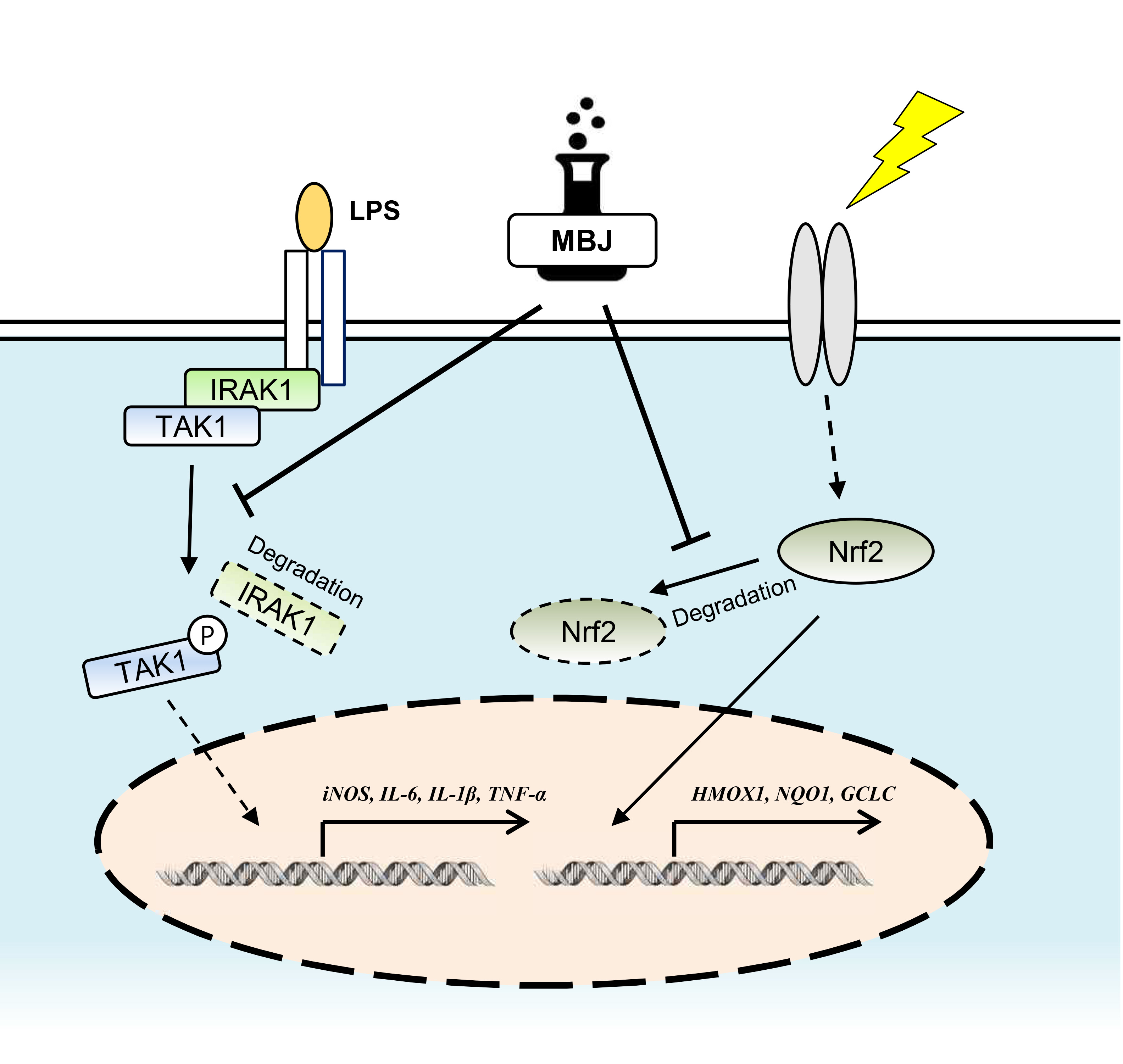
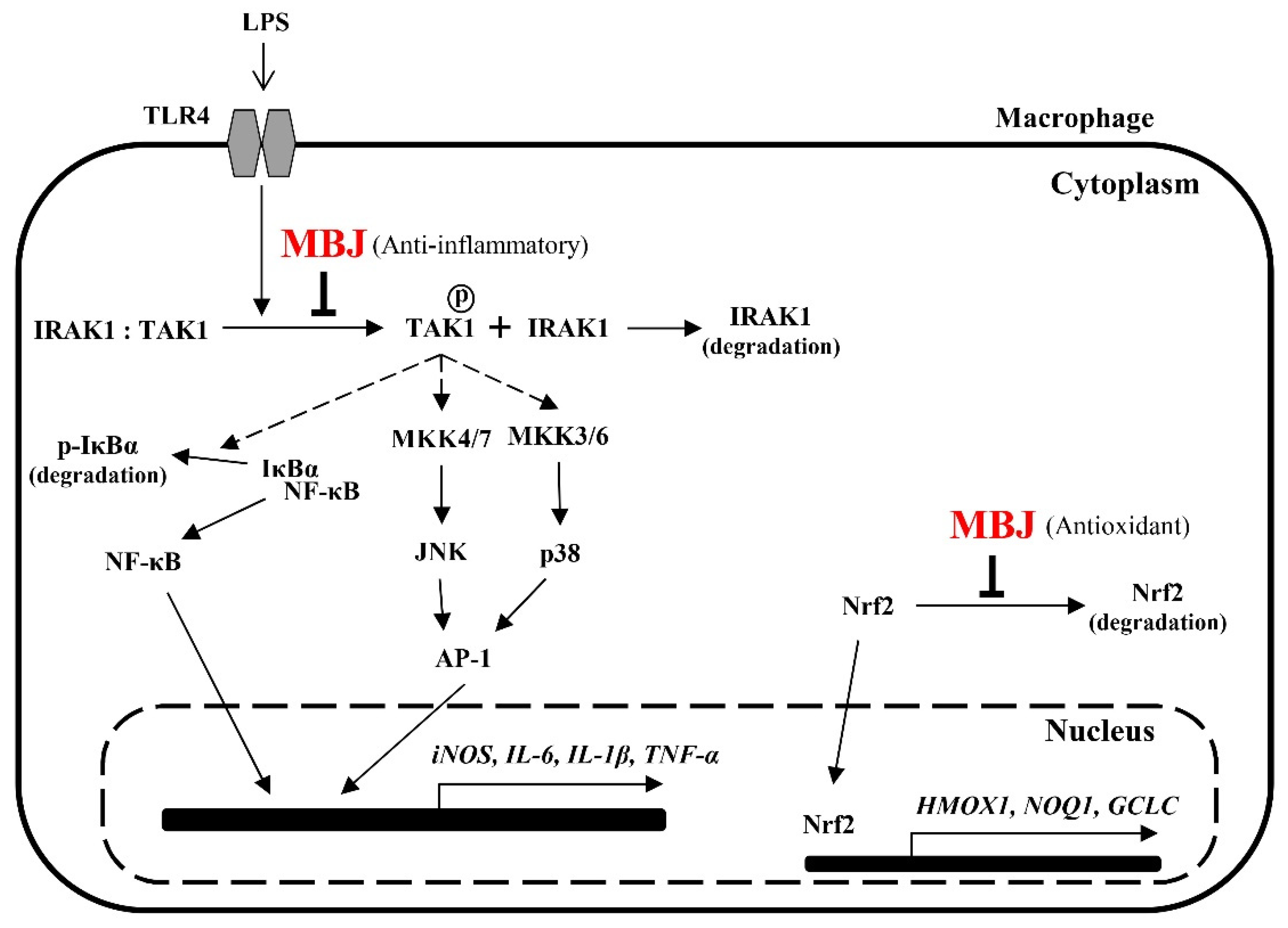
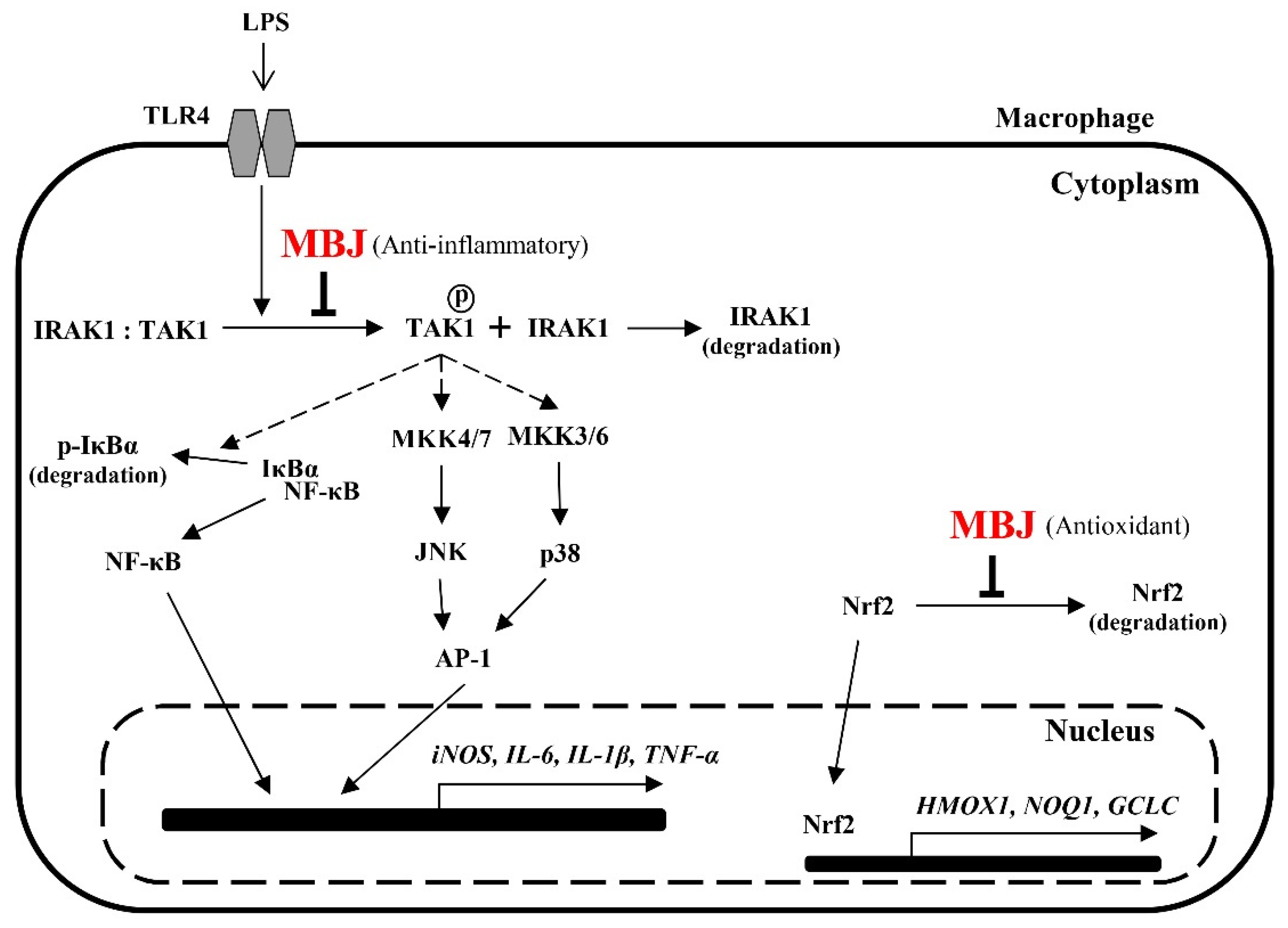
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Jalabi, W.; Shpargel, K.B.; Farabaugh, K.T.; Dutta, R.; Yin, X.; Kidd, G.J.; Bergmann, C.C.; Stohlman, S.A.; Trapp, B.D. Lipopolysaccharide-induced microglial activation and neuroprotection against experimental brain injury is independent of hematogenous TLR4. J. Neurosci. 2012, 32, 11706–11715. [Google Scholar] [CrossRef] [Green Version]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Jia, Z.; Zhu, H. Regulation of Nrf2 Signaling. React. Oxyg. Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef]
- Dunne, A.; Carpenter, S.; Brikos, C.; Gray, P.; Strelow, A.; Wesche, H.; Morrice, N.; O’Neill, L.A. IRAK1 and IRAK4 promote phosphorylation, ubiquitination, and degradation of MyD88 adaptor-like (Mal). J. Biol. Chem. 2010, 285, 18276–18282. [Google Scholar] [CrossRef] [Green Version]
- Landstrom, M. The TAK1-TRAF6 signalling pathway. Int. J. Biochem. Cell Biol. 2010, 42, 585–589. [Google Scholar] [CrossRef]
- Sakurai, H.; Miyoshi, H.; Mizukami, J.; Sugita, T. Phosphorylation-dependent activation of TAK1 mitogen-activated protein kinase kinase kinase by TAB1. FEBS Lett. 2000, 474, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Suppression of Inflammatory Responses by Black Rice Extract in RAW 264.7 Macrophage Cells via Downregulation of NF-kB and AP-1 Signaling Pathways. Asian Pac. J. Cancer Prev. 2015, 16, 4277–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunaweera, N.; Raju, R.; Gyengesi, E.; Münch, G. Plant polyphenols as inhibitors of NF-κB induced cytokine production—A potential anti-inflammatory treatment for Alzheimer’s disease? Front. Mol. Neurosci. 2015, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Alhouayek, M.; Muccioli, G.G. COX-2-derived endocannabinoid metabolites as novel inflammatory mediators. Trends Pharmacol. Sci. 2014, 35, 284–292. [Google Scholar] [CrossRef]
- Lin, C.-C.; Edelson, B.T. New insights into the role of IL-1β in experimental autoimmune encephalomyelitis and multiple sclerosis. J. Immunol. 2017, 198, 4553–4560. [Google Scholar] [CrossRef] [Green Version]
- Shaftel, S.S.; Griffin, W.S.; O’Banion, M.K. The role of interleukin-1 in neuroinflammation and Alzheimer disease: An evolving perspective. J. Neuroinflammation 2008, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Horiuchi, T.; Mitoma, H.; Harashima, S.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-alpha: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef] [Green Version]
- Sastri, B.N. The Wealth of India. A Dictionary of Indian Raw Materials and Industrial Products. Raw Materials; Council of Scientific and Industrial Research: New Delhi, India, 1950. [Google Scholar]
- Rijal, A. Surviving on Knowledge: Ethnobotany of Chepang community from mid-hills of Nepal. Ethnobot. Res. Appl. 2011, 9, 181–215. [Google Scholar] [CrossRef] [Green Version]
- Saha, G.; Biswas, R.; Das, A. Survey of medicinal plants in the Gorumara National Park, Jalpaiguri, West Bengal, India. Pleione 2013, 7, 127–137. [Google Scholar]
- Rajbongshi, P.; Zaman, K.; Boruah, S.; Das, S. A review on traditional use and phytopharmacological potential of Bischofia javanica Blume. Int. J. Pharm. Sci. Rev. Res. 2014, 24, 24–29. [Google Scholar]
- Chopra, R.; Nayar, S.; Chopra, I.; Asolkar, L.; Kakkar, K. Glossary of Indian Medicinal Plants; Council of Scientific and Industrial Research: New Delhi, India, 1956. [Google Scholar]
- Cambie, R.C.; Ash, J. Fijian Medicinal Plants; CSIRO: Clayton, Australia, 1994.
- Sutharson, L.; Prasanna, K.; Lila, K.; Shila, E.; Rajan, J. Free radical scavenging activity of Leaves of Bischofia javanica Blume and Fraxinus Floribunda wallich. Pharmacologyonline 2009, 1, 1324–1332. [Google Scholar]
- Dunstan, C.A.; Noreen, Y.; Serrano, G.; Cox, P.A.; Perera, P.; Bohlin, L. Evaluation of some Samoan and Peruvian medicinal plants by prostaglandin biosynthesis and rat ear oedema assays. J. Ethnopharmacol. 1997, 57, 35–56. [Google Scholar] [CrossRef]
- Barstow, M. Bischofia javanica. The IUCN Red List of Threatened Species 2019: e.T61956399A61956472. Available online: https://www.iucnredlist.org/species/61956399/61956472 (accessed on 30 March 2020).
- Le, H.T.T.; Cho, Y.C.; Cho, S. Methanol extract of Guettarda speciosa Linn. inhibits the production of inflammatory mediators through the inactivation of Syk and JNK in macrophages. Int. J. Mol. Med. 2018, 41, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO. 10993–5:2009 Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. In International Organization for Standardization; ISO: Geneva, Switzerland, 2009. [Google Scholar]
- Hou, G.Q.; Guo, C.; Song, G.H.; Fang, N.; Fan, W.J.; Chen, X.D.; Yuan, L.; Wang, Z.Q. Lipopolysaccharide (LPS) promotes osteoclast differentiation and activation by enhancing the MAPK pathway and COX-2 expression in RAW264.7 cells. Int. J. Mol. Med. 2013, 32, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanos, C.; Freudenberg, M.A.; Reutter, W. Galactosamine-induced sensitization to the lethal effects of endotoxin. Proc. Natl. Acad. Sci. USA 1979, 76, 5939–5943. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Yang, Y.; Shen, Y.; Li, L.; Huang, J.; Tang, L.; Zhang, L. Luzindole attenuates LPS/d-galactosamine-induced acute hepatitis in mice. Innate Immun. 2020, 26, 319–327. [Google Scholar] [CrossRef]
- Wu, Z.; Han, M.; Chen, T.; Yan, W.; Ning, Q. Acute liver failure: Mechanisms of immune-mediated liver injury. Liver Int. 2010, 30, 782–794. [Google Scholar] [CrossRef]
- Peng, X.; Yang, Y.; Tang, L.; Wan, J.; Dai, J.; Li, L.; Huang, J.; Shen, Y.; Lin, L.; Gong, X. Therapeutic benefits of apocynin in mice with lipopolysaccharide/D-galactosamine-induced acute liver injury via suppression of the late stage pro-apoptotic AMPK/JNK pathway. Biomed. Pharmacother. 2020, 125, 110020. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Tirkey, N.; Bharrhan, S.; Chanana, V.; Rishi, P.; Chopra, K. Inhibition of oxidative stress and cytokine activity by curcumin in amelioration of endotoxin-induced experimental hepatoxicity in rodents. Clin. Exp. Immunol. 2006, 145, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Yang, H.; Noureddin, M.; Iglesia-Ara, A.; Xia, M.; Wagner, C.; Luka, Z.; Mato, J.M.; Lu, S.C. Changes in S-adenosylmethionine and GSH homeostasis during endotoxemia in mice. Lab. Investig. 2008, 88, 1121–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Walker, L.M.; Hinson, J.A.; Mayeux, P.R. Oxidant stress in rat liver after lipopolysaccharide administration: Effect of inducible nitric-oxide synthase inhibition. J. Pharmacol. Exp. Ther. 2000, 293, 968–972. [Google Scholar]
- Lee, E.-J.; Cheon, C.-I.; Kim, J.-S.; Yoo, H.; Sung, M.-K. Evaluation of plant phenolic compounds as anti-oxidative and anti-inflammatory agents. Cancer Prev. Res. 2008, 13, 18–25. [Google Scholar]
- Park, C.M.; Song, Y.-S. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-κB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. Nutr. Res. Pract. 2013, 7, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, D.R.; Acharya, A.K.; Pradhan, M.; Chhetri, R.; Sherpa, S.D.; Lepcha, D.L. Nutraceutical potential of two edible wild fruits, Bischofia javanica Blume and Ficus cunia Buch.-Ham. ex Roxb. from Sikkim Himalaya. Int. J. Food Sci. Nutr. 2017, 2, 1–9. [Google Scholar]
- Indra, R.; Bachheti, R.; Archana, J. Chemical composition, mineral and nutritional value of wild Bischofia javanica seed. Int. Food Res. J. 2013, 20, 1747–1751. [Google Scholar]
- Sutharson Lingadurai, S.R.; Joseph, R.V.; Nath, L.K. Antileukemic activity of the leaf extract of Bischofia javanica blume on human leukemic cell lines. Indian J. Pharmacol. 2011, 43, 143. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.R.; Chowdhury, K.H.; Hanif, N.B.; Sayeed, M.A.; Mouah, J.; Mahmud, I.; Kamal, A.M.; Chy, M.N.U.; Adnan, M. An integrated exploration of pharmacological potencies of Bischofia javanica (Blume) leaves through experimental and computational modeling. Heliyon 2020, 6, e04895. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A. Islands, Plants, and Polynesians: An Introduction to Polynesian Ethnobotany: Proceedings of a Symposium; Dioscorides Press: Portland, OR, USA, 1991. [Google Scholar]
- Kay, A.B. Asthma and inflammation. J. Allergy Clin. Immunol. 1991, 87, 893–910. [Google Scholar] [CrossRef]
- Libby, P. Role of inflammation in atherosclerosis associated with rheumatoid arthritis. Am. J. Med. 2008, 121, S21–S31. [Google Scholar] [CrossRef]
- Ishihara, K.; Hirano, T. IL-6 in autoimmune disease and chronic inflammatory proliferative disease. Cytokine Growth Factor Rev. 2002, 13, 357–368. [Google Scholar] [CrossRef]
- Neurath, M.F.; Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev. 2011, 22, 83–89. [Google Scholar] [CrossRef]
- Stenvinkel, P.; Alvestrand, A. Review articles: Inflammation in end-stage renal disease: Sources, consequences, and therapy. Semin. Dial. 2002, 15, 329–337. [Google Scholar] [CrossRef]
- Vane, J.; Botting, R. Anti-inflammatory drugs and their mechanism of action. Inflamm. Res. 1998, 47, 78–87. [Google Scholar] [CrossRef]
- Gremese, E.; Ferraccioli, G.F. Benefit/risk of cyclosporine in rheumatoid arthritis. Clin. Exp. Rheumatol. 2004, 22, S101–S107. [Google Scholar]
- Jang, J.-Y.; Lee, M.-J.; You, B.-R.; Jin, J.-S.; Lee, S.-H.; Yun, Y.-R.; Kim, H.J. Allium hookeri root extract exerts anti-inflammatory effects by nuclear factor-κB down-regulation in lipopolysaccharide-induced RAW264. 7 cells. BMC Complement. Altern Med. 2017, 17, 126. [Google Scholar]
- Kleemann, R.; Verschuren, L.; Morrison, M.; Zadelaar, S.; van Erk, M.J.; Wielinga, P.Y.; Kooistra, T. Anti-inflammatory, anti-proliferative and anti-atherosclerotic effects of quercetin in human in vitro and in vivo models. Atherosclerosis 2011, 218, 44–52. [Google Scholar] [CrossRef]
- Seelinger, G.; Merfort, I.; Schempp, C.M. Anti-oxidant, anti-inflammatory and anti-allergic activities of luteolin. Planta Med. 2008, 74, 1667–1677. [Google Scholar] [CrossRef]
- Baricevic, D.; Sosa, S.; Della Loggia, R.; Tubaro, A.; Simonovska, B.; Krasna, A.; Zupancic, A. Topical anti-inflammatory activity of Salvia officinalis L. leaves: The relevance of ursolic acid. J. Ethnopharmacol. 2001, 75, 125–132. [Google Scholar] [CrossRef]
- Zhu, L.; Yi, X.; Zhao, J.; Yuan, Z.; Wen, L.; Pozniak, B.; Obminska-Mrukowicz, B.; Tian, Y.; Tan, Z.; Wu, J. Betulinic acid attenuates dexamethasone-induced oxidative damage through the JNK-P38 MAPK signaling pathway in mice. Biomed. Pharmacother. 2018, 103, 499–508. [Google Scholar] [CrossRef]
- Madajka, M.; Korda, M.; White, J.; Malinski, T. Effect of aspirin on constitutive nitric oxide synthase and the biovailability of NO. Thromb. Res. 2003, 110, 317–321. [Google Scholar] [CrossRef]
- Ogata, S.; Kubota, Y.; Yamashiro, T.; Takeuchi, H.; Ninomiya, T.; Suyama, Y.; Shirasuna, K. Signaling pathways regulating IL-1alpha-induced COX-2 expression. J. Dent. Res. 2007, 86, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.N.; Pahan, K. Signals for the induction of nitric oxide synthase in astrocytes. Neurochem. Int. 2006, 49, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Rhodus, N.L.; Cheng, B.; Myers, S.; Bowles, W.; Ho, V.; Ondrey, F. A comparison of the pro-inflammatory, NF-kappaB-dependent cytokines: TNF-alpha, IL-1-alpha, IL-6, and IL-8 in different oral fluids from oral lichen planus patients. Clin. Immunol. 2005, 114, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, Y.; Jin, Z.; Jiang, H.; Wen, J. Silica-Induced TNF-α and TGF-β 1 Expression in RAW264. 7 Cells are Dependent on Src-ERK/AP-1 Pathways. Toxicol. Mech. Methods 2009, 19, 51–58. [Google Scholar] [CrossRef]
- Silvers, A.L.; Bachelor, M.A.; Bowden, G.T. The role of JNK and p38 MAPK activities in UVA-induced signaling pathways leading to AP-1 activation and c-Fos expression. Neoplasia 2003, 5, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Chen, T.; Liu, F. Betulinic acid alleviates myocardial hypoxia/reoxygenation injury via inducing Nrf2/HO-1 and inhibiting p38 and JNK pathways. Eur. J. Pharmacol. 2018, 838, 53–59. [Google Scholar] [CrossRef]
- Wang, C.; Deng, L.; Hong, M.; Akkaraju, G.R.; Inoue, J.; Chen, Z.J. TAK1 is a ubiquitin-dependent kinase of MKK and IKK. Nature 2001, 412, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Xiao, N.; Xiao, H.; Zhou, H.; Yu, M.; Gu, J.; Li, X. Beta-TrCP-mediated IRAK1 degradation releases TAK1-TRAF6 from the membrane to the cytosol for TAK1-dependent NF-kappaB activation. Mol. Cell. Biol. 2012, 32, 3990–4000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paupe, V.; Dassa, E.P.; Goncalves, S.; Auchere, F.; Lonn, M.; Holmgren, A.; Rustin, P. Impaired nuclear Nrf2 translocation undermines the oxidative stress response in Friedreich ataxia. PLoS ONE 2009, 4, e4253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Liu, L.; Li, X.; Zhang, L.; Lv, J.; Guo, X.; Chen, H.; Zhao, T. Neuroprotective effects of an Nrf2 agonist on high glucose-induced damage in HT22 cells. Biol. Res. 2019, 52, 53. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Ha, J.; Park, J.; Kang, E.; Jeon, S.-H.; Han, S.B.; Ningsih, S.; Paik, J.H.; Cho, S. Antioxidant and Anti-Inflammatory Effects of Bischofia javanica (Blume) Leaf Methanol Extracts through the Regulation of Nrf2 and TAK1. Antioxidants 2021, 10, 1295. https://doi.org/10.3390/antiox10081295
Lee S, Ha J, Park J, Kang E, Jeon S-H, Han SB, Ningsih S, Paik JH, Cho S. Antioxidant and Anti-Inflammatory Effects of Bischofia javanica (Blume) Leaf Methanol Extracts through the Regulation of Nrf2 and TAK1. Antioxidants. 2021; 10(8):1295. https://doi.org/10.3390/antiox10081295
Chicago/Turabian StyleLee, Sewoong, Jain Ha, Jiyoung Park, Eunjeong Kang, Sung-Hyun Jeon, Sang Beom Han, Sri Ningsih, Jin Hyub Paik, and Sayeon Cho. 2021. "Antioxidant and Anti-Inflammatory Effects of Bischofia javanica (Blume) Leaf Methanol Extracts through the Regulation of Nrf2 and TAK1" Antioxidants 10, no. 8: 1295. https://doi.org/10.3390/antiox10081295
APA StyleLee, S., Ha, J., Park, J., Kang, E., Jeon, S.-H., Han, S. B., Ningsih, S., Paik, J. H., & Cho, S. (2021). Antioxidant and Anti-Inflammatory Effects of Bischofia javanica (Blume) Leaf Methanol Extracts through the Regulation of Nrf2 and TAK1. Antioxidants, 10(8), 1295. https://doi.org/10.3390/antiox10081295