
The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery

 , and
, and 
Abstract
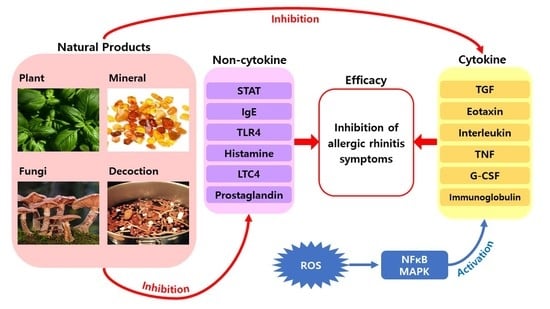
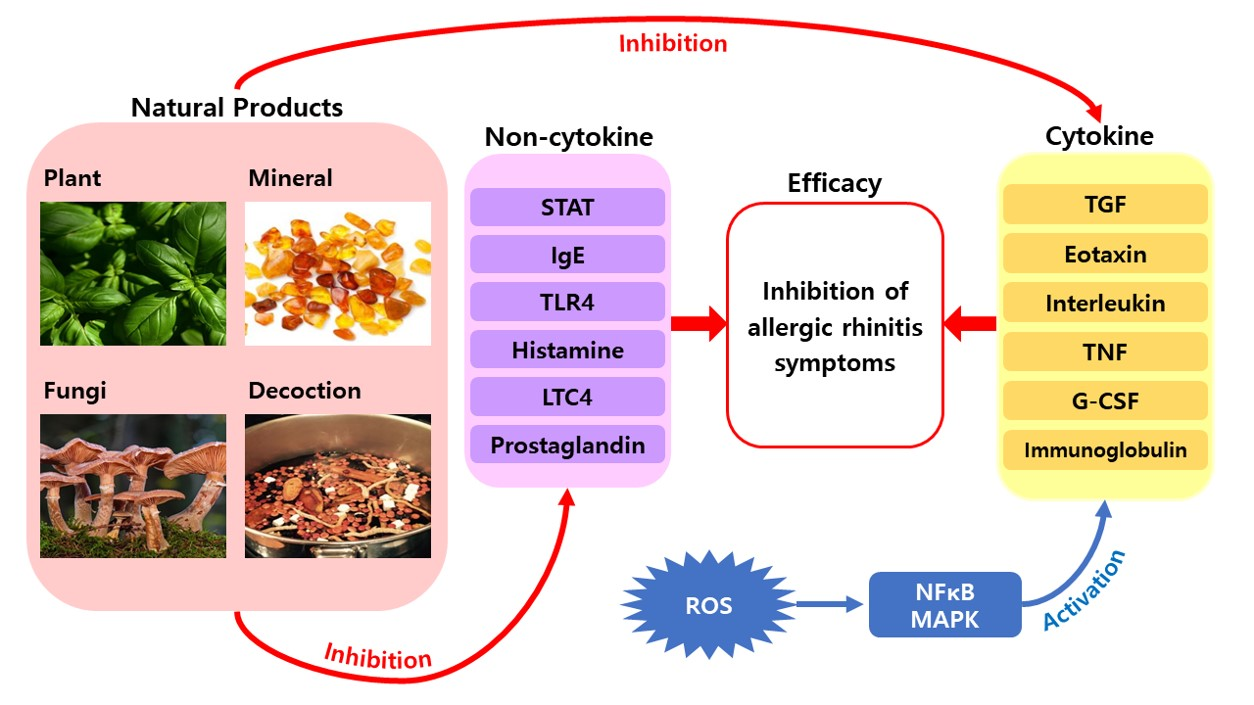
1. Introduction
2. Pathogenesis of Allergic Rhinitis
2.1. T Helper Cell Differentiation
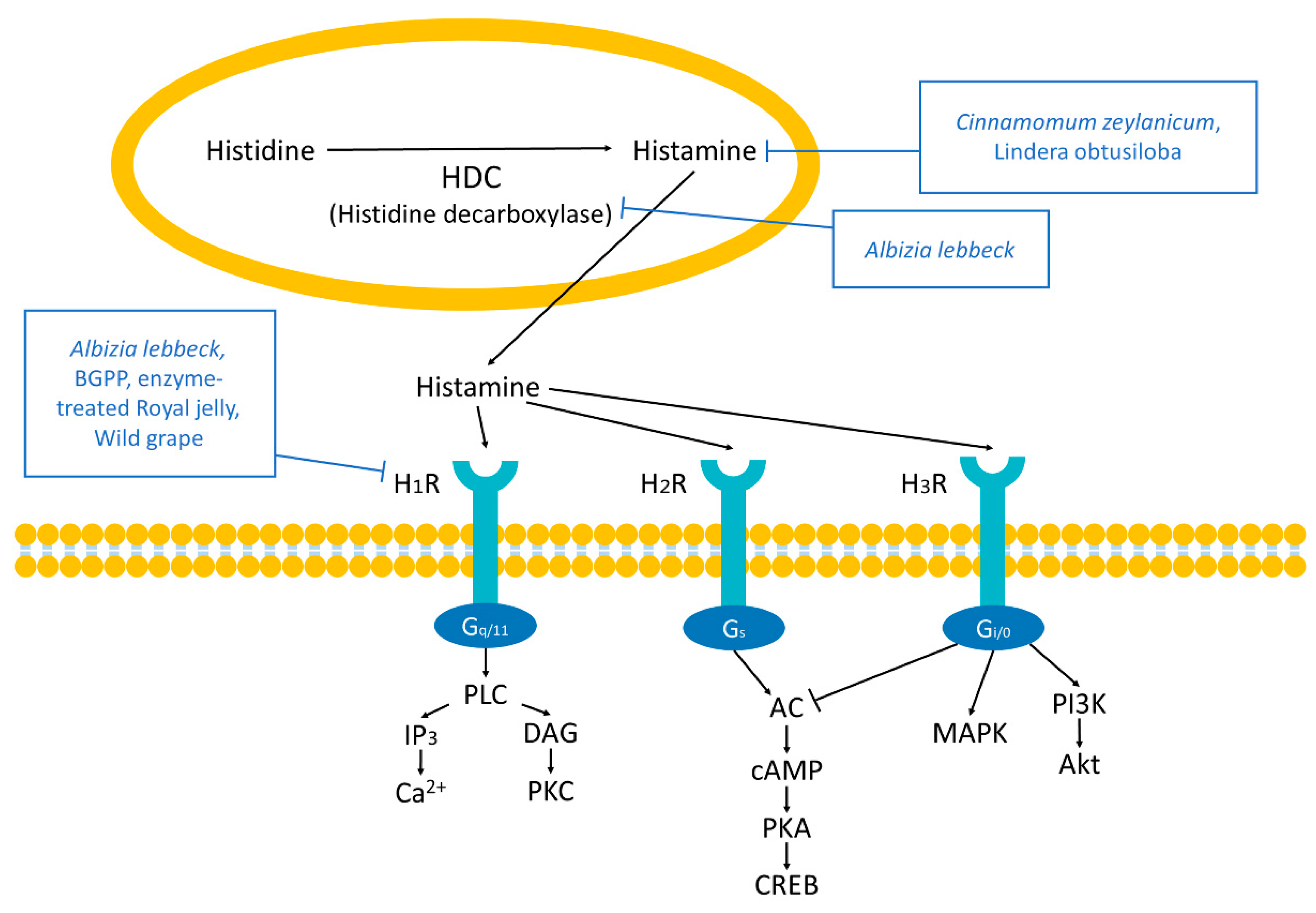
2.2. Histamine Receptors
2.3. 5-Lipooxygenase Pathway and Prostaglandin E2 Synthesis-and-Signaling Pathway
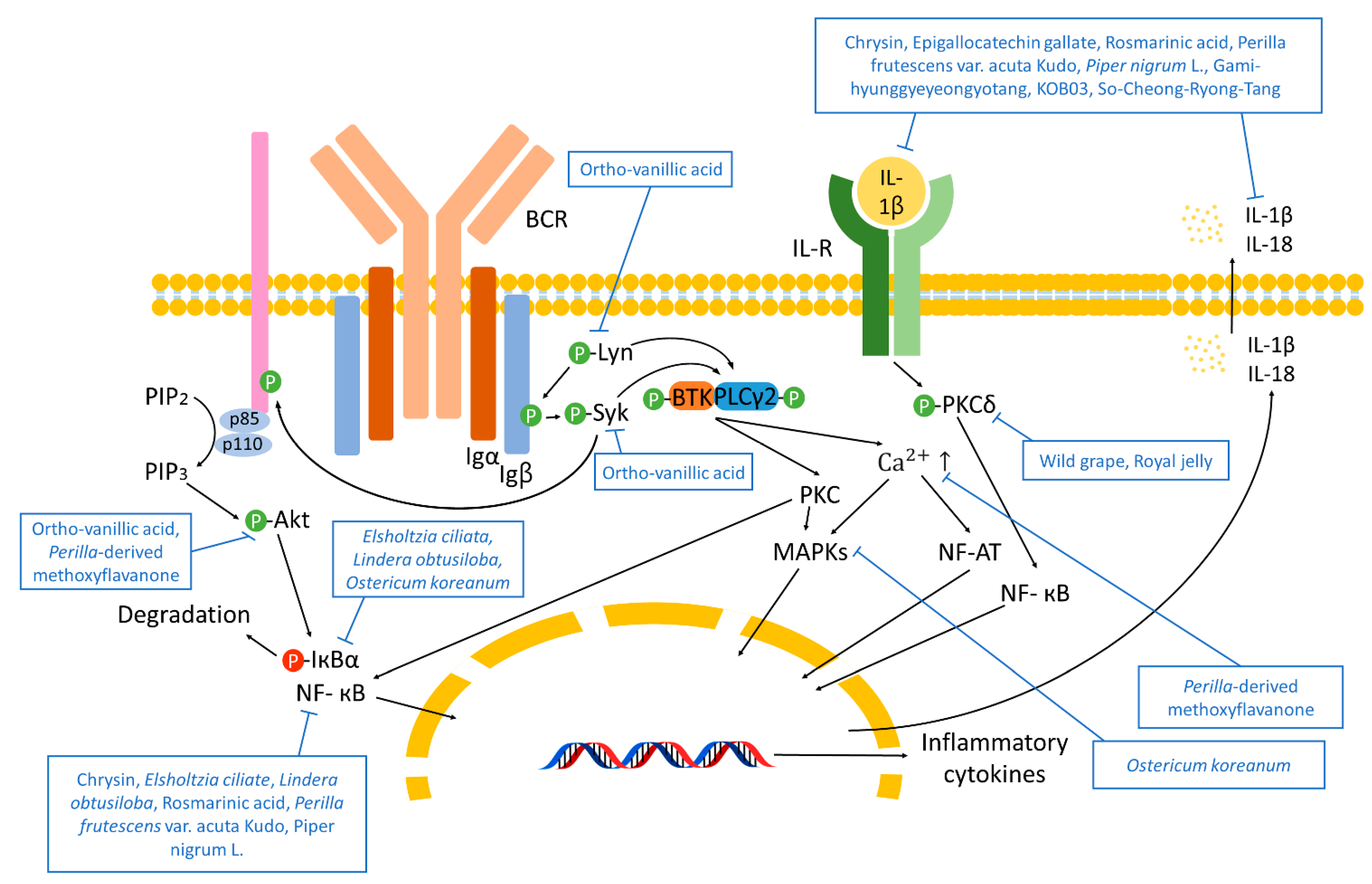
2.4. Immune Response Pathways
3. Natural Products That Inhibit Allergic Rhinitis
3.1. Plant-Originated Natural Products
3.1.1. In Vitro Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound/Extract | Source | Experimental Subject | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Albizia lebbeck bark ethanol extract | Albizia lebbeck | PMA-treated HeLa | 0.1, 1, 10 μg/mL; 24 h | Improvement of allergic nasal symptoms | ↓ H1R, HDC | [55] |
| Histamine-treated HeLa | 10 μg/mL; 24 h | |||||
| Brazilian green propolis ethanol extract | Baccharis dracunculifolia | TDI-sensitized rats | 40, 80 mg/kg; 3 weeks | Improvement of allergic nasal symptoms | ↓ H1R, p-PKCδ, IL-9, IL-4, IL-5 | [56] |
| HeLa | 25, 75, 100, 125, 200 µg/mL; 3 h | ↓ H1R, p-PKCδ, IL-9 | ||||
| RBL-2H3 | 25, 75, 100 µg/mL; 3 h | |||||
| Brazilian green propolis ethanol extract | Baccharis dracunculifolia | Cry j1-treated PBMCs | 3, 10, 30 μg/mL; 6 days | Prevention of the nasal obstruction | ↓ IL-5, IL-13 | [57] |
| Cry j1-treated peripheral leukocytes from allergic human | 3, 10, 30, 100 μg/mL; 6 days | ↓ CysLTs | ||||
| Chrysin (5,7-dihydroxyflavone) | Passiflora caerulea | (Anti-DNP IgE)-sensitized RBL-2H3 | 0.1, 1, 10 µg/mL; 30 min | Inhibition of mast cell-derived allergic inflammatory reactions | ↓ TNF-α, IL-1β, IL-4, IL-6, caspase-1, NF-κB | [58] |
| PMA/A23187-treated RBL-2H3 | ||||||
| PMA/A23187-treated HMC-1 | ||||||
| Citrus limon Burm. f. | Citrus limon (L.) Burm. f. | PBMCs from five grass pollen-allergic human | 0.01 g/mL; 7 days | Reduction of allergic responses Attenuation of nasal inflammation | ↑ IFN-γ ↓ TNF-α, IL-5 | [60] |
| Cydonia oblonga Mill. | Cydonia oblonga Mill. | PBMCs from five grass pollen-allergic human | 0.01 g/mL; 7 days | Reduction of allergic responses Attenuation of nasal inflammation | ↑ IFN-γ | [60] |
| Elsholtzia ciliata (Thunb.) Hyl. (Labiatae) water extract | Elsholtzia ciliata (Thunb.) Hyl. | PMACI-exposed HMC-1 | 1, 10, 100 μg/mL; 30 min | Inhibition of mast cell-mediated allergic reactions Treatment of allergic inflammatory diseases | ↑ IκBα ↓ TNF-α, IL-6, NF-κB, p-p38 MAPK | [61] |
| Enzyme-treated (protease-degraded) Royal Jelly | Apis cerana Fabricius | RBL-2H3 | 25, 75, 100 µg/mL; 3 h | Improvement of allergic nasal symptoms | ↓ H1R, p-PKCδ, IL-9 | [56] |
| HeLa | 25, 75, 100, 125, 200 µg/mL; 3 h | |||||
| Magnoliae Flos ethanol extract | Magnoliae Flos | hANO1-transfected HEK293T | 0.1, 0.3, 1 mg/mL; 600 s | Prevention of allergic rhinitis | [62] | |
| HEK293T, Calu-3 | 30, 100, 300 μg/mL; 600 s | |||||
| Gencydo® | Citrus limon, Cydonia oblonga | (DNP–HSA)-treated RBL-2H3 | 0.2, 0.4, 0.8 mg/mL; 10 min | Inhibition of the release of histamine and degranulation | ↓ IL-8, TNF-α | [63] |
| (DNP–HSA)-treated mice BMMCs | 0.2, 0.4, 0.8 mg/mL; 60 min | |||||
| (TNF-α/IL-4)-treated human bronchial epithelial cells | 0.2, 0.4, 0.8 mg/mL; 24 h | Inhibition of the release of eotaxin | ||||
| PMA/A23187-activated human mast cells | 0.2, 0.4, 0.8 mg/mL; 24 h | Orchestration of early- and late-phase allergic reaction | ||||
| (GM-CSF)-treated human eosinophil granulocytes | 0.2, 0.4, 0.8 mg/mL; 24 h | No impact on the behavior of (GM-CSF)-treated human eosinophils | ||||
| Lindera obtusiloba water extract | Lindera obtusiloba | PMACI-treated HMC-1 | 0.1, 1, 10, 100 μg/mL; 30 min | Reduction of histamine release Inhibition of systemic and local allergic reaction | ↑ IκBα ↓ TNF-α, IL-6, NF-κB, histamine | [64] |
| (DNP-HSA)-treated RBL-2H3 | ||||||
| (DNP-HSA)-treated RPMCs | ||||||
| Ortho-vanillic acid | Amomum xanthioides | RPMCs | 1, 10, 100 nmol/L; 24 h | Suppression of the mast cell-mediated allergic inflammatory response | ↓ TNF-α, IL-4, p-Lyn, p-Syk, p-Akt | [65] |
| OVA-treated RBL-2H3 | 1, 10, 100 nmol/L; 24 h | Suppression of histamine release | ||||
| Ostericum koreanum root methanol extract | PMA/A23187-treated RPMCs | 0.2, 0.5, 1 mg/mL; 30 min | Attenuation of disease progression as determined by nasal symptoms and histological changes of the nasal mucosa | ↓ TNF-α, IL-6, p-ERK1/2, p-p38 MAPK, p-IκBα | [66] | |
| Compound 48/80-treated RPMCs | 0.5, 1 mg/mL; 15 min | Decreased compound 48/80-induced release of histamine | ||||
| Perilla-derived methoxyflavanone | Perilla frutescens | (Anti-DNP IgE)-sensitized RBL-2H3 | 25–300 µM; 30 min | Inhibition of IgE-mediated histamine release | ↓ p-Akt, intracellular Ca2+ influx | [67] |
| 100, 300 µM/2.5 mL; 30 min | Suppression of IgE-mediated mast cell degranulation | |||||
| Perilla frutescens var. acuta Kudo 30% ethanol extract powder | Perilla frutescens var. | Mouse mast cells | 1 g/kg; 10 days | Amelioration of allergic inflammatory reactions | ↓ IL-1β, IL-6, TNF-α, COX-2, NF-κB | [68] |
| Mouse Eosinophils | 1 g/kg; 10 days | |||||
| PMACI-exposed HMC-1 | 0.01, 0.1, 1 mg/mL; 1 h | |||||
| Rosmarinic acid | Mouse mast cells | 4 g/kg; 10 days | Amelioration of allergic inflammatory reactions | ↓ IL-1β, IL-6, TNF-α, NF-κB | [68] | |
| Mouse Eosinophils | 4 g/kg; 10 days | |||||
| PMACI-exposed HMC-1 | 100 µM; 1 h | |||||
| Spinacia oleracea Linn. aqueous extract | Spinacia oleracea Linn. | (Hydrogen-peroxide)-applied SH-SY5Y neuroblastoma cells | 25 μL/mL; 24 h | Viable cell number recovery | ↓ IL-4, IL-13, Ig-E | [69] |
| Ze339 | Petasites hybridus | Human nasal epithelial cells; inferior turbinates of patients | 3 µg/mg; 24 h | Reduction of recruitment of inflammatory cells, Inhibition of pro-inflammatory cytokine and chemokine response to viral mimics | ↑ G-CSF ↓ IL-8, CCL-3, IL-6, STAT1, STAT3, STAT6 | [70] |
3.1.2. In Vivo Studies
| Compound/Extract | Source | Experimental Subject | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Albizia lebbeck bark ethanol extract | Albizia lebbeck | TDI-sensitized allergy rats | 200 mg; 3 weeks | Reduction of the amount of sneezing and nasal rubbing. | ↓ IL-4, IL-5, IL-13 | [55] |
| Artemisia vulgaris water extract | Artemisia vulgaris | Mugwort pollen-injected AR BALB/c mice | 10, 50, 100 µg/mL; 15 days | Reduction of allergic inflammation | ↑ IL-12 ↓ IL-4, IL-5, IgE2, IgE | [71] |
| Awa-tea leaves hot water extract | Setaria italica | TDI-sensitized rats | 40 mg/kg; 21 days | Inhibition of the allergic diseases | ↓ IL-9, IL-4 | [72] |
| Berberine | Berberis | (Dermatophagoides farinae (Derf)-OVA)-induced AR BALB/c mice | 10 µg/mL; 57 days | Reduction of allergic inflammation | ↑ Foxp3 | [77] |
| Brazilian green propolis ethanol extract | Baccharis dracunculifolia | TDI-sensitized rats | 40, 80 mg/kg; 3 weeks | Improvement of allergic nasal symptoms | ↓ H1R, p-PKCδ, IL-9, IL-4, IL-5 | [56] |
| Bupleurum chinense water extract | Bupleurum chinense | OVA-induced AR BALB/c mice | 50, 100, 200 mg/kg; 11 days | Attenuation of allergic responses Suppression of nasal inflammation | ↑ IL-10, IL-12, IFN-γ ↓ IL-4, IL-5, IL-13, CCL24 | [79] |
| Chrysin (5,7-dihydroxyflavone) | Passiflora caerulea | Compound 48/80-induced systemic anaphylaxis mice | 1–100 mg/kg; 1 h | Blockage of compound 48/80-induced fatal shock | [58] | |
| PCA mice | Reduction of PCA reaction | |||||
| Cinnamomum zeylanicum bark hydroalcoholic extract | Cinnamomum zeylanicum | OVA-induced AR BALB/c mice | 3, 10, 30 µg/kg; 8 days | Attenuation of nasal inflammation | ↓ IgE, histamine | [83] |
| Dryopteris crassirhizoma ethanol extract | Dryopteris crassirhizoma | OVA-induced AR BALB/c mice | 100, 200, 400 mg/kg; 11 days | Amelioration of nasal inflammation | ↓ IgE, IgG1, IgG2 | [85] |
| Enzyme-treated (protease-degraded) Royal Jelly | Apis cerana Fabricius | TDI-sensitized rats | 40, 80 mg/kg; 3 weeks | Improvement of allergic nasal symptoms | ↓ H1R, p-PKCδ, IL-9, IL-4, IL-5 | [56] |
| Epigallocatechin gallate | Camellia sinensis | OVA-induced mice | 25, 50, 100 mg/kg; 10 days | Reduction of allergic rhinitis | ↓ IL-1β, IL-4, IL-6, COX-2 | [86] |
| Glycine max water extract | Glycine max | (2,4-TDI)-induced rhinitis guinea pigs | 30, 150, 300 µg/kg; 28 days | Suppression of nasal inflammation Decrease of allergic responses | ↓ IgE | [89] |
| Lindera obtusiloba water extract | Lindera obtusiloba | Compound 48/80-induced systemic allergy mice | 1, 10, 100 mg/kg; 1 h | Reduction of histamine release Inhibition of systemic and local allergic reaction | ↑ IκBα ↓ TNF-α, IL-6, NF-κB | [64] |
| Ortho-vanillic acid | Amomum xanthioides | OVA-induced ASA mice | 2, 10, 50 mg/kg; 14 days | Attenuation of the ovalbumin-induced ASA | ↓ TNF-α, IL-4, p-Lyn, p-Syk, p-Akt | [65] |
| IgE-mediated PCA mice | 50 mg/kg; 1 h | Attenuation of the IgE mediated cutaneous allergic reactions | ||||
| Ostericum koreanum root methanol extract | Ostericum koreanum | OVA-induced allergic rhinitis BALB/c mice | 50, 100 mg/kg; 7 days | Attenuation of disease progression as determined by nasal symptoms and histological changes of the nasal mucosa | ↑ IFN-γ ↓ OVA-specific IgE, IL-4 | [66] |
| Perilla frutescens var. acuta Kudo 30% ethanol extract powder | Perilla frutescens var. | OVA-sensitized allergic rhinitis mice | 0.01, 0.1, 1 g/kg; 10 days | Amelioration of allergic inflammatory reactions | ↓ IL-1β, IL-6, TNF-α, COX-2, NF-κB | [68] |
| Piper nigrum L. ethanol extract | Piper nigrum L. | OVA-induced AR BALB/c mice | 50, 100, 200 mg/kg; 13 days | Inhibition of allergic nasal symptoms Suppression of nasal inflammation | ↓ STAT3, NFκBp65 | [93] |
| Piper nigrum L. extract | Piper nigrum L | OVA-induced AR Swiss albino mice | 10, 20, 40 mg/kg; 7 days | Exhibits immunomodulatory and anti-inflammatory activity | ↓ IL-6, IL-1β, IgE | [94] |
| Propolis | SD rats | 200 mg/kg; 21 days | Inhibition of allergic symptoms | [101] | ||
| Rosae Multiflorae Fructus water extract | Rosae Multiflorae Fructus | OVA-induced AR BALB/c mice | 100, 200, 400 mg/kg; 13 days | Inhibition of allergic symptoms Immunomodulatory effect on allergic rhinitis | ↑ IL-12 ↓ IL-4, IL-5, IL-13 | [103] |
| Rosmarinic acid | OVA-sensitized allergic rhinitis mice | 4 mg/kg; 10 days | Amelioration of allergic inflammatory reactions | ↓ IL-1β, IL-6, TNF-α, NF-κB | [68] | |
| Spinacia oleracea Linn. aqueous extract | Spinacia oleracea Linn. | OVA-challenge asthmatic mice | 25 μL/mL; 24 h | Improvement of the asthmatic symptoms induced by OVA-challenge Reduction of the BAL’s eosinophil expression | ↓ IL-4, IL-13, Ig-E | [69] |
| Syzygium formosum leaves extract | Syzygium formosum (Wall.) Masam. | cOVA-induced food allergy mice | 80, 200 mg/kg; 13 days | Improvement of the food allergy symptoms and the inflammatory lesion in the gut. | ↓Th2 cytokines (e.g., IL-4, IL-5, IL-10), MCTP-1 | [104] |
| Xanthii Fructus ethanol extract | Xanthium sibiricum Patr. | OVA-sensitized Male BALB/C mice | 1 g/kg; 15 days | Alleviation of nasal orifices and elimination of wind-dampness | ↑ PC(40:8), L-valine | [105] |
| Wild grape hot water extract | Ampelopsis glandulosa | TDI-sensitized rats | 25, 50 mg/kg; 21 days | Alleviation of the nasal symptoms Alleviation of the eosinophilic inflammation | ↓ p-PKCδ, H1R, IL-9, IL-4, IL-5, IL-33 | [72] |
3.2. Fungi and Minerals-Originated Natural Products
3.2.1. In Vitro Studies
3.2.2. In Vivo Studies
3.3. Decoctions of Natural Products
3.3.1. In Vitro Studies
3.3.2. In Vivo Studies
| Name | Constitutions | Experimental Subject | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Biyeom-Tang (ethanol extract) | Xanthii Fructus (Xanthium strumarium L.), Trichosanthis Semen (Trichosanthes kirilowii Maxim.), Angelicae Dahuricae Radix, Menthae Herba (Mentha arvensis L.) | BMMCs from BALB/c mice | 12.5, 25, 50 μg/mL; 1 h | Inhibition of allergic and inflammatory effects | ↓ β-Hex | [115] |
| BMMCs from BALB/c mice | 6.3, 12.5, 25 μg/mL; 1 h | ↓ PGD2, LTC4 | ||||
| OVA-induced AR BALB/c mice | 50, 100, 200 mg/kg; 7 days | ↓ IL-4, IL-5, IL-10, IL-13 | ||||
| Biyuanling | Xanthium strumarium subsp. sibiricum (Patrin ex Widder) Greuter, Lonicera japonica Thunb., Glycyrrhiza uralensis Fisch., Scutellaria baicalensis Georgi | TDI-induced AR Guinea pigs | 4, 8, 16 mg/g; 7 days | Inhibition of inflammatory response | ↑ Bax ↓ IL-4, SIgE, TNF-α, SP, VCAM-1, P65 | [116] |
| Bu-Zhong-Yi-Qi-Tang | Astragali Radix, Ginseng Radix, Atractylodes macrocephala Koidz., Glycyrrhizae Radix, Cimicifugae Rhizoma, Bupleuri Radix, Aurantii Nobilis Pericarpium Citri Unshius Pericarpium, finger citron, Chinese magnolia vine fruit | (OVA-and-alum)-induced AR BALB/c mice | 10 g/100 mL; 10, 19 days | Improvement of the AR-like symptoms | ↑ IFN-γ, Foxp3 ↓ IgE, IL-4, CD4 | [117] |
| Gami-hyunggyeyeongyotang | Schizonepetae Spica (Nepeta tenuifolia Benth.), Forsythiae Fructus (Forsythia viridissima Lindl.), Saposhnikoviae Radix (Saposhnikovia divaricata (Turcz.) Schischk.), Angelicae Radix (Angelica acutiloba Kitagawa), Cnidii Rhizoma (Cnidium officinale Makino), Paeoniae Radix Alba (Paeonia lactiflora Pallas), Angelicae Dahuricae Radix (Angelica dahurica Bentham et Hooker), Bupleuri Radix (Bupleurum falcatum Linne), Scutellariae Radix (Scutellaria baicalensis Georgi), Aurantii Fructus Immaturus (Citrus aurantium Linne), Gardeniae Frutus (Gardenia jasminoides Ellis), Platycodi Radix (Platycodon grandiflorum A. De candole), Glycyrrhizae Radix (Glycyrrhiza uralensis Fischer), Ulmi Cortex (Ulmus macrocarpa Hance), Xanthii Fructus (Xanthium strumarium Linne), Magnoliae Flos (Magnolia kobus De Candolle), Rubiae Radix (Rubia akane Nakai) | OVA-induced AR BALB/c mice | 134 mg/kg; 11 days | Alleviation of the symptoms of AR | ↓ IgE, IL-5, IL-6, IL-1β, MCP-1, MIP-2, caspase-1 | [111] |
| Hyeonggaeyeongyo-Tang | Schizonepeta Spica, Forsythiae Fructus, Saposhnikoviae Radix, Angelicae Gigantis Radix, Cnidii Rhizoma, Paeoniae Radix Alba, Bupleuri Radix, Aurantii Fructus, Scutellariae Radix, Gardeniae Fructus, Angelicae Dahuricae Radix, Platycodi Radix, Glycyrrhizae Radix | OVA-induced AR BALB/c mice | 101, 202, 404 mg/kg; 14 days | Suppression of the progression of AR | ↓ IL-4, IL-13, LIF | [118] |
| KOB03 | Atractylodis Rhizoma Alba (Atractylodes japonica Koidz), Astragalus Radix (Astragalus membranaceus Bunge), Saposhnikoviae Radix (Saposhnikovia divaricata Schischkin), Osterici Radix (Ostericum koreanum maximowicz), Scutellariae Radix (Scutellaria baicalensis Georgi) | OVA-induced AR BALB/c mice | 100, 200 mg/kg; 7 days | Regulation of allergic inflammation | ↓ TNF-α, IL-1β, IL-6, IL-8 | [122] |
| OVA-induced AR SD rats | ||||||
| KOB03 | Atractylodis Rhizoma White (Atractylodes japonica Koidz), Astragalus Root (Astragalus membranaceus Bunge), Saposhnikovia Root (Saposhnikovia divaricata Schischkin), Ostericum Root (Ostericum koreanum maximowicz), Scutellaria Root (Scutellaria baicalensis Georgi) | OVA-induced AR BALB/c mouse model | 500 mg/kg; 8 days | Inhibition of nasal inflammation | ↓ IgE, LTC4, IL-4, and IL-1β | [123] |
| Mahuang Fuzi Xixin Decoction | Ephedrae, Aconiti Lateralis Radix, Asarum | OVA-induced AR SPF Wistar rats | 1.52 g/mL; 10 days | Alleviation of the symptoms of AR | ↓ IgE, histamine, IFN-γ, IL-4, GATA3, STAT6 | [124] |
| Pyeongwee-San extract (KMP6) | Atractylodes japonica Koidz. ex Kitam., Machilus thunbergii Siebold & Zucc., Citrus reticulata Blanco, Zizyphus jujube, Glycyrrhiza uralensis Fisch., Zingiber officinale Roscoe | OVA-induced AR BALB/c mice | 1 mg/mL; 10 days | Amelioration of AR | ↑ IFN-γ ↓ IgE, IL-4, TNF-α, IL-1β, MIP-2, ICAM-1, COX-2 | [125] |
| Qi-fang-bi-min-tang (QFBMT) | Astragali Radix, Ramulus Cinnamomi, Saposhnikoviae Radix, Magnoliae Flos, Atractylodis Macrocephalae Rhizoma, Zingiberis Rhizoma, Paeoniae Radix Alba, Glycyrrhizae Radix | OVA-induced AR SD rats | 5 × 106 MSCs, 2% QFBMT; 19 days | Inhibition of AR symptoms combined with MSCs | ↓ IFN-γ, IL-17, IL-4, TNF-α, IgE | [126] |
| OVA-induced AR BALB/c mice | QFBMT + MSCs: 5 × 106 MSCs, 2% | Inhibition of AR symptoms combined with MSCs | ↓ TNF-α, IL-6, IFN-γ, IL-4, IL-17 | |||
| QFBMT: 20%; 19 days | Inhibition of AR | |||||
| Senn-kinn-naidaku-sann | Ginseng Radix, Angelicae Radix, Astragali Radix, Cnidii Rhizoma, Sinomeni Caulis et Rhizoma, Platycodonis Radix, Magnoliae Cortex, Angelicae Dahuricae Radix, Glycyrrhizae Radix | OVA-induced AR C3H/HeN mice | 100, 1000 mg/kg; 7 days | Alleviation of the symptoms of AR | ↑ IFN-γ | [113] |
| OVA-induced AR C3H/HeJ mice | 100, 1000 mg/kg; 7 days | No enhancement of AR | ↓ IgE, IgG1, IL-4, IL-5 | |||
| Shenqi | Angelicae Dahuricae Radix (Angelica dahurica (Hoffm.) Benth. & Hook.f. ex Franch. & Sav.), Scutellariae Radix (Scutellaria baicalensis Georgi), Lonicerae Japonicae Flos (Lonicera japonica Thunb.), Menthae Haplocalycis Herba (Mentha canadensis L.), Astragali Radix (Astragalus propinquus Schischkin), Codonopsis Radix (Codonopsis pilosula subsp. tangshen (Oliv.) D.Y.Hong), Isatidis Folium (Isatis tinctoria L.), Taraxaci Herba (Taraxacum mongolicum Hand.-Mazz.), Agrimoniae Herba (Agrimonia pilosa Ledeb.), Asiasari Radix et Rhizome (Asarum sieboldii Miq.) | OVA-sensitized primary spleen lymphocytes of Wistar rats | 50, 100, 200 μg/mL; 7 days | Alleviation of symptoms of AR | ↑ T-bet ↓ GATA3, p-STAT6, SOCS1 | [114] |
| OVA-induced AR Wistar rats | 0.9, 2.7, 8.1 g/kg; 7 days | ↑ IFN-γ, T-bet ↓ IL-4, IgE, GATA3, p-STAT6, SOCS1 | ||||
| So-Cheong-Ryong-Tang | Asarum sieboldii Miq., Glycyrrhiza uralensis Fisch., Ephedra sinica Stapf, Cinnamomum cassia Blume, Peony root, Pinellia ternata (Thunb.) Makino, Schisandra chinensis (Turcz.) Baill., Zingiber officinale Roscoe | OVA-induced AR BALB/c mice | 20 mg/mL; 14 days | Suppression of the progression of AR | ↓ IL-4, IL-6, IL-1β, TNF-α, LIF | [127] |
| So-Cheong-Ryong-Tang | Pinelliae Tuber (Pinellia ternata (Thunb.) Makino), Ephedrae Herba (Ephedra sinica Stapf), Schisandrae Fructus (Schisandra chinensis (Turcz.) Baill.), Paeoniae Radix (Paeonia lactiflora Pall.), Glycyrrhizae Radix (Glycyrrhiza uralensis Fisch.), Zingiberis Siccatum Rhizoma (Zingiber officinale Roscoe), Asiasari Radix (Asarum sieboldii Miq.), Cinnamomi Cortex (Cinnamomum cassia (L.) J.Presl) | (OVA-and-alum)-induced AR BALB/c mice | 1 g/kg; 7 days | Alleviation of the symptoms of AR | ↑ IgG2a, IFN-γ ↓ IgE, IL-4, IgG1, IL-5 | [128] |
| Tong Qiao | Angelicae Dahuricae Radix, Gleditsia sinensis Lam., Magnoliae Flos | OVA-induced AR SD rats | 10 µL/kg; 7,15 days | Alleviation of the symptoms of AR | ↓ eotaxin, IL-5, IL-13 | [129] |
| Xingbi gel | Cynanchi Paniculati Radix (Cynanchum paniculatum (Bunge) Kitag. ex H.Hara), Cicadidae Periostracum (cicada slough), Bovis Calculus (bezoar), Borneolum Syntheticum (borneol) | OVA-induced AR guinea pigs | 50 µL/kg; 12 days | Alleviation of the symptoms of AR | ↓ LTE4, IgE | [130] |
| Yiqi Wenyang Fang | Astragalus membranaceus (Fisch.) Bunge, Codonopsis pilosula (Franch.) Nannf., Zingiberis Rhizoma, Cinnamomi Ramulus, Ephedra sinica Stapf, Schisandra chinensis (Turcz.) Baill. | OVA-induced AR SD rats | 1.6 g/mL; 28 days | Inhibition of inflammatory response Alleviation of the symptoms of AR | ↓ IL-10, TGF-β1, IL-4, IL-13 | [131] |
3.4. Clinical Trials
4. Discussion
4.1. Mechanisms of Natural Products Regulating AR Symptoms
4.1.1. T Helper Cell Differentiation
4.1.2. Histamine Receptors
4.1.3. 5-Lipooxygenase Pathway and Prostaglandin E2 Synthesis-and-Signaling Pathway
4.1.4. Immune Response Pathways
4.2. Comparison Analysis
4.3. Limitations
4.4. Significance
5. Methods
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartle, J. Patient education in the effective management of hay fever. Nurs. Stand. 2016, 30, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Brożek, J.L.; Bousquet, J.; Agache, I.; Agarwal, A.; Bachert, C.; Bosnic-Anticevich, S.; Brignardello-Petersen, R.; Canonica, G.W.; Casale, T.; Chavannes, N.H. Allergic Rhinitis and its Impact on Asthma (ARIA) guidelines—2016 revision. J. Allergy Clin. Immunol. 2017, 140, 950–958. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef]
- Bousquet, J.; Van Cauwenberge, P.; Khaltaev, N. Allergic rhinitis and its impact on asthma. J. Allergy Clin. Immunol. 2001, 108, S147–S334. [Google Scholar] [CrossRef]
- May, J.R.; Dolen, W.K. Management of allergic rhinitis: A review for the community pharmacist. Clin. Ther. 2017, 39, 2410–2419. [Google Scholar] [CrossRef]
- Seidman, M.D.; Gurgel, R.K.; Lin, S.Y.; Schwartz, S.R.; Baroody, F.M.; Bonner, J.R.; Dawson, D.E.; Dykewicz, M.S.; Hackell, J.M.; Han, J.K. Clinical practice guideline: Allergic rhinitis. Otolaryngol.–Head Neck Surg. 2015, 152, S1–S43. [Google Scholar] [CrossRef]
- Greiner, A.N.; Hellings, P.W.; Rotiroti, G.; Scadding, G.K. Allergic rhinitis. Lancet 2011, 378, 2112–2122. [Google Scholar] [CrossRef]
- Mygind, N. Allergic rhinitis. In History of Allergy; Karger Publishers: Basel, Switzerland, 2014; Volume 100, pp. 62–68. [Google Scholar]
- Dipiro, J.T.; Talbert, R.L.; Yee, G.C.; Matzke, G.R.; Wells, B.G.; Posey, L.M. Pharmacotherapy: A Pathophysiologic Approach, ed; McGraw-Hill Medical: New York, NY, USA, 2014. [Google Scholar]
- Lipworth, B.J.; Jackson, C.M. Safety of inhaled and intranasal corticosteroids. Drug Saf. 2000, 23, 11–33. [Google Scholar] [CrossRef]
- Lin, L.; Luo, L.; Zhong, M.; Xie, T.; Liu, Y.; Li, H.; Ni, J. Gut microbiota: A new angle for traditional herbal medicine research. RSC Adv. 2019, 9, 17457–17472. [Google Scholar] [CrossRef]
- Srivastava, R.; Srivastava, S.; Singh, S.P. Comparative study of anti-allergic activity of two poly-herbal formulations in ova-challenged allergic rhinitis mice model. Orient. Pharm. Exp. Med. 2019, 19, 37–47. [Google Scholar] [CrossRef]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural products for human health: An historical overview of the drug discovery approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef]
- David, B.; Wolfender, J.-L.; Dias, D.A. The pharmaceutical industry and natural products: Historical status and new trends. Phytochem. Rev. 2015, 14, 299–315. [Google Scholar] [CrossRef]
- Platts-Mills, T.A. The role of immunoglobulin E in allergy and asthma. Am. J. Respir. Crit. Care Med. 2001, 164, S1–S5. [Google Scholar] [CrossRef] [PubMed]
- Ishizaka, K.; Ishizaka, T.; Lee, E.H. Biologic function of the Fc fragments of E myeloma protein. Immunochemistry 1970, 7, 687–702. [Google Scholar] [CrossRef]
- Metzger, H.; Alcaraz, G.; Hohman, R.; Kinet, J.; Pribluda, V.; Quarto, R. The receptor with high affinity for immunoglobulin E. Annu. Rev. Immunol. 1986, 4, 419–470. [Google Scholar] [CrossRef]
- Bieber, T.; De La Salle, H.; Wollenberg, A.; Hakimi, J.; Chizzonite, R.; Ring, J.; Hanau, D.; De la Salle, C. Human epidermal Langerhans cells express the high affinity receptor for immunoglobulin E (Fc epsilon RI). J. Exp. Med. 1992, 175, 1285–1290. [Google Scholar] [CrossRef]
- Wang, B.; Rieger, A.; Kilgus, O.; Ochiai, K.; Maurer, D.; Födinger, D.; Kinet, J.; Stingl, G. Epidermal Langerhans cells from normal human skin bind monomeric IgE via Fc epsilon RI. J. Exp. Med. 1992, 175, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.; Liu, X.; Fang, L.; Chen, X.; Guo, T.; Zhang, J. The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease. Cell. Mol. Immunol. 2010, 7, 182–189. [Google Scholar] [CrossRef]
- Hallberg, D.; Pernow, B. Effect of substance P on various vascular beds in the dog. Acta Physiol. Scand. 1975, 93, 277–285. [Google Scholar] [CrossRef]
- Johnson, A.R.; Erdös, E.G. Release of histamine from mast cells by vasoactive peptides. Proc. Soc. Exp. Biol. Med. 1973, 142, 1252–1256. [Google Scholar] [CrossRef]
- Hägermark, O.; Hökfelt, T.; Pernow, B. Flare and itch induced by substance P in human skin. Nation 1978, 12, 13. [Google Scholar] [CrossRef]
- Taylor-Clark, T. Histamine in allergic rhinitis. In Histamine in Inflammation; Springer: Boston, MA, USA, 2010; pp. 33–41. [Google Scholar]
- Huang, H.; Li, Y.; Liang, J.; Finkelman, F.D. Molecular Regulation of Histamine Synthesis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Schwartz, J.C.; Arrang, J.M.; Garbarg, M.; Pollard, H.; Ruat, M. Histaminergic transmission in the mammalian brain. Physiol. Rev. 1991, 71, 1–51. [Google Scholar] [CrossRef]
- Hill, S.J. Distribution, properties, and functional characteristics of three classes of histamine receptor. Pharm. Rev. 1990, 42, 45–83. [Google Scholar] [PubMed]
- Hill, S.J.; Ganellin, C.R.; Timmerman, H.; Schwartz, J.C.; Shankley, N.P.; Young, J.M.; Schunack, W.; Levi, R.; Haas, H.L. International Union of Pharmacology. XIII. Classification of histamine receptors. Pharm. Rev. 1997, 49, 253–278. [Google Scholar] [PubMed]
- Debacker, M.; Gommeren, W.; Moereels, H.; Nobels, G.; Vangompel, P.; Leysen, J.E.; Luyten, W.H. Genomic cloning, heterologous expression and pharmacological characterization of a human histamine H1 receptor. Biochem. Biophys. Res. Commun. 1993, 197, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Umemori, H.; Inoue, T.; Kume, S.; Sekiyama, N.; Nagao, M.; Itoh, H.; Nakanishi, S.; Mikoshiba, K.; Yamamoto, T. Activation of the G Protein Gq/11 Through Tyrosine Phosphorylation of the α Subunit. Science 1997, 276, 1878–1881. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.M.; Cripps, A.W.; West, N.P.; Cox, A.J. Modulation of allergic inflammation in the nasal mucosa of allergic rhinitis sufferers with topical pharmaceutical agents. Front. Pharmacol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Houston, C.; Wenzel-Seifert, K.; Bürckstümmer, T.; Seifert, R. The human histamine H2-receptor couples more efficiently to Sf9 insect cell Gs-proteins than to insect cell Gq-proteins: Limitations of Sf9 cells for the analysis of receptor/Gq-protein coupling. J. Neurochem. 2002, 80, 678–696. [Google Scholar] [CrossRef]
- Shayo, C.; Davio, C.; Brodsky, A.; Mladovan, A.G.; Legnazzi, B.L.; Rivera, E.; Baldi, A. Histamine Modulates the Expression of c-fos through Cyclic AMP Production via the H2 Receptor in the Human Promonocytic Cell Line U937. Mol. Pharmacol. 1997, 51, 983–990. [Google Scholar] [CrossRef]
- Hegyi, K.; Falus, A.; Toth, S. Elevated CREB activity in embryonic fibroblasts of gene-targeted histamine deficient mice. Inflamm. Res. 2007, 56, 339–344. [Google Scholar] [CrossRef]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of histamine in modulating the immune response and inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef]
- Wijtmans, M.; Leurs, R.; Esch, I.d. Histamine H3 receptor ligands break ground in a remarkable plethora of therapeutic areas. Expert Opin. Investig. Drugs 2007, 16, 967–985. [Google Scholar] [CrossRef]
- Bongers, G.; Bakker, R.A.; Leurs, R. Molecular aspects of the histamine H3 receptor. Biochem. Pharmacol. 2007, 73, 1195–1204. [Google Scholar] [CrossRef]
- Nieto-Alamilla, G.; Márquez-Gómez, R.; García-Gálvez, A.-M.; Morales-Figueroa, G.-E.; Arias-Montaño, J.-A. The Histamine H3 Receptor: Structure, Pharmacology, and Function. Mol. Pharmacol. 2016, 90, 649–673. [Google Scholar] [CrossRef]
- Xu, H.; Gu, L.-N.; Yang, Q.-Y.; Zhao, D.-Y.; Liu, F. MiR-221 promotes IgE-mediated activation of mast cells degranulation by PI3K/Akt/PLCγ/Ca2+ pathway. J. Bioenerg. Biomembr. 2016, 48, 293–299. [Google Scholar] [CrossRef]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef]
- Son, Y.; Kim, S.; Chung, H.-T.; Pae, H.-O. Reactive oxygen species in the activation of MAP kinases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 528, pp. 27–48. [Google Scholar]
- Quadri, S.S.; Culver, S.A.; Li, C.; Siragy, H.M. Interaction of the renin angiotensin and cox systems in the kidney. Front. Biosci. (Sch. Ed.) 2016, 8, 215. [Google Scholar]
- Nasrallah, R.; Hassouneh, R.; Hébert, R.L. PGE2, kidney disease, and cardiovascular risk: Beyond hypertension and diabetes. J. Am. Soc. Nephrol. 2016, 27, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.F.; Chang, L.C.; Huang, L.J.; Kuo, S.C.; Lee, H.Y.; Lu, M.C.; Wang, J.P. The influence of acetylshikonin, a natural naphthoquinone, on the production of leukotriene B4 and thromboxane A2 in rat neutrophils. Eur. J. Pharm. 2009, 607, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Pinho, B.R.; Sousa, C.; Valentao, P.; Oliveira, J.M.; Andrade, P.B. Modulation of basophils’ degranulation and allergy-related enzymes by monomeric and dimeric naphthoquinones. PLoS ONE 2014, 9, e90122. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef]
- Miller, D.K.; Gillard, J.W.; Vickers, P.J.; Sadowski, S.; Leveille, C.; Mancini, J.A.; Charleson, P.; Dixon, R.A.; Ford-Hutchinson, A.W.; Fortin, R.; et al. Identification and isolation of a membrane protein necessary for leukotriene production. Nature 1990, 343, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Colazzo, F.; Gelosa, P.; Tremoli, E.; Sironi, L.; Castiglioni, L. Role of the Cysteinyl Leukotrienes in the Pathogenesis and Progression of Cardiovascular Diseases. Mediat. Inflamm. 2017, 2017, 2432958. [Google Scholar] [CrossRef]
- Opene, M.; Kurantsin-Mills, J.; Husain, S.; Ibe, B.O. Sickle erythrocytes and platelets augment lung leukotriene synthesis with downregulation of anti-inflammatory proteins: Relevance in the pathology of the acute chest syndrome. Pulm. Circ. 2014, 4, 482–495. [Google Scholar] [CrossRef]
- Brodie, E.J.; Infantino, S.; Low, M.S.Y.; Tarlinton, D.M. Lyn, Lupus, and (B) Lymphocytes, a Lesson on the Critical Balance of Kinase Signaling in Immunity. Front. Immunol. 2018, 9, 401. [Google Scholar] [CrossRef]
- Reth, M. Antigen receptor tail clue. Nature 1989, 338, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Kim, S.J.; Lee, H.-S.; Kwon, O.-S. PKCδ mediates NF-κB inflammatory response and downregulates SIRT1 expression in liver fibrosis. Int. J. Mol. Sci. 2019, 20, 4607. [Google Scholar] [CrossRef]
- Carpenter, L.; Cordery, D.; Biden, T.J. Protein kinase Cdelta activation by interleukin-1beta stabilizes inducible nitric-oxide synthase mRNA in pancreatic beta-cells. J. Biol. Chem. 2001, 276, 5368–5374. [Google Scholar] [CrossRef]
- Willingham, S.B.; Allen, I.C.; Bergstralh, D.T.; Brickey, W.J.; Huang, M.T.-H.; Taxman, D.J.; Duncan, J.A.; Ting, J.P.-Y. NLRP3 (NALP3, Cryopyrin) Facilitates In Vivo Caspase-1 Activation, Necrosis, and HMGB1 Release via Inflammasome-Dependent and -Independent Pathways. J. Immunol. 2009, 183, 2008–2015. [Google Scholar] [CrossRef] [PubMed]
- Nurul, I.M.; Mizuguchi, H.; Shahriar, M.; Venkatesh, P.; Maeyama, K.; Mukherjee, P.K.; Hattori, M.; Choudhuri, M.S.; Takeda, N.; Fukui, H. Albizia lebbeck suppresses histamine signaling by the inhibition of histamine H1 receptor and histidine decarboxylase gene transcriptions. Int. Immunopharmacol. 2011, 11, 1766–1772. [Google Scholar] [CrossRef]
- Shaha, A.; Mizuguchi, H.; Kitamura, Y.; Fujino, H.; Yabumoto, M.; Takeda, N.; Fukui, H. Effect of Royal Jelly and Brazilian Green Propolis on the Signaling for Histamine H1 Receptor and Interleukin-9 Gene Expressions Responsible for the Pathogenesis of the Allergic Rhinitis. Biol. Pharm. Bull. 2018, 41, 1440–1447. [Google Scholar] [CrossRef]
- Tani, H.; Hasumi, K.; Tatefuji, T.; Hashimoto, K.; Koshino, H.; Takahashi, S. Inhibitory activity of Brazilian green propolis components and their derivatives on the release of cys-leukotrienes. Bioorg. Med. Chem. 2010, 18, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.; Lee, S.; Kim, S.H. Chrysin suppresses mast cell-mediated allergic inflammation: Involvement of calcium, caspase-1 and nuclear factor-kappaB. Toxicol. Appl. Pharm. 2011, 254, 56–64. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, A.; Baars, E. Citrus/Cydonia Comp. Use in General Practice. A Survey among Anthroposophic Physicians; Louis Bolk Instituut: Driebergen, The Netherlands, 2001; p. 50. [Google Scholar]
- Baars, E.W.; Jong, M.C.; Boers, I.; Nierop, A.F.; Savelkoul, H.F. A comparative in vitro study of the effects of separate and combined products of Citrus e fructibus and Cydonia e fructibus on immunological parameters of seasonal allergic rhinitis. Mediat. Inflamm. 2012, 2012, 109829. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, H.H.; Yoo, J.S.; Lee, H.S.; Kwon, T.K.; Shin, T.Y.; Kim, S.H. Elsholtzia ciliata inhibits mast cell-mediated allergic inflammation: Role of calcium, p38 mitogen-activated protein kinase and nuclear factor-{kappa}B. Exp. Biol. Med. (Maywood) 2011, 236, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Nam, Y.R.; Nam, J.H. Flos Magnoliae Inhibits Chloride Secretion via ANO1 Inhibition in Calu-3 Cells. Am. J. Chin. Med. 2018, 46, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Grundemann, C.; Papagiannopoulos, M.; Lamy, E.; Mersch-Sundermann, V.; Huber, R. Immunomodulatory properties of a lemon-quince preparation (Gencydo(R)) as an indicator of anti-allergic potency. Phytomedicine 2011, 18, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Suh, W.M.; Park, S.B.; Lee, S.; Kim, H.H.; Suk, K.; Son, J.H.; Kwon, T.K.; Choi, H.G.; Lee, S.H.; Kim, S.H. Suppression of mast-cell-mediated allergic inflammation by Lindera obtusiloba. Exp. Biol. Med. (Maywood) 2011, 236, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Je, I.G.; Kim, M.J.; Kang, B.C.; Choi, Y.A.; Baek, M.C.; Lee, B.; Choi, J.K.; Park, H.R.; Shin, T.Y.; et al. 2-Hydroxy-3-methoxybenzoic acid attenuates mast cell-mediated allergic reaction in mice via modulation of the FcepsilonRI signaling pathway. Acta Pharm. Sin. 2017, 38, 90–99. [Google Scholar] [CrossRef]
- Jung, H.W.; Jung, J.K.; Park, Y.K. Antiallergic effect of Ostericum koreanum root extract on ovalbumin-induced allergic rhinitis mouse model and mast cells. Asian Pac. J. Allergy Immunol. 2011, 29, 338–348. [Google Scholar]
- Kamei, R.; Fujimura, T.; Matsuda, M.; Kakihara, K.; Hirakawa, N.; Baba, K.; Ono, K.; Arakawa, K.; Kawamoto, S. A flavanone derivative from the Asian medicinal herb (Perilla frutescens) potently suppresses IgE-mediated immediate hypersensitivity reactions. Biochem. Biophys. Res. Commun. 2017, 483, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.A.; Park, C.S.; Ahn, H.J.; Park, Y.S.; Kim, H.M. Effect of Perilla frutescens var. acuta Kudo and rosmarinic acid on allergic inflammatory reactions. Exp. Biol. Med. (Maywood) 2011, 236, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.C.; Park, C.H.; Lee, H.J.; Kim, S.O.; Kim, T.H.; Lee, S.H. Amelioration of asthmatic inflammation by an aqueous extract of Spinacia oleracea Linn. Int. J. Mol. Med. 2010, 25, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Steiert, S.A.; Zissler, U.M.; Chaker, A.M.; Esser-von-Bieren, J.; Dittlein, D.; Guerth, F.; Jakwerth, C.A.; Piontek, G.; Zahner, C.; Drewe, J.; et al. Anti-inflammatory effects of the petasin phyto drug Ze339 are mediated by inhibition of the STAT pathway. Biofactors 2017, 43, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Bao, M.J.; Zhu, J.P.; Yao, H.Y.; Xie, Y.C.; Guan, Y.; Li, F.F.; Dong, X.W.; Zheng, Y.M.; Xie, Q.M. Oral administration of allergen extracts from mugwort pollen desensitizes specific allergen-induced allergy in mice. Vaccine 2012, 30, 1437–1444. [Google Scholar] [CrossRef]
- Islam, R.; Mizuguchi, H.; Shaha, A.; Nishida, K.; Yabumoto, M.; Ikeda, H.; Fujino, H.; Kitamura, Y.; Fukui, H.; Takeda, N. Effect of wild grape on the signaling of histamine H1 receptor gene expression responsible for the pathogenesis of allergic rhinitis. J. Med. Investig. 2018, 65, 242–250. [Google Scholar] [CrossRef]
- Hu, Z.; Jiao, Q.; Ding, J.; Liu, F.; Liu, R.; Shan, L.; Zeng, H.; Zhang, J.; Zhang, W. Berberine induces dendritic cell apoptosis and has therapeutic potential for rheumatoid arthritis. Arthritis Rheum. 2011, 63, 949–959. [Google Scholar] [CrossRef]
- Cui, G.; Qin, X.; Zhang, Y.; Gong, Z.; Ge, B.; Zang, Y.Q. Berberine differentially modulates the activities of ERK, p38 MAPK, and JNK to suppress Th17 and Th1 T cell differentiation in type 1 diabetic mice. J. Biol. Chem. 2009, 284, 28420–28429. [Google Scholar] [CrossRef]
- Qin, X.; Guo, B.T.; Wan, B.; Fang, L.; Lu, L.; Wu, L.; Zang, Y.Q.; Zhang, J.Z. Regulation of Th1 and Th17 cell differentiation and amelioration of experimental autoimmune encephalomyelitis by natural product compound berberine. J. Immunol. 2010, 185, 1855–1863. [Google Scholar] [CrossRef]
- Ma, X.; Jiang, Y.; Wu, A.; Chen, X.; Pi, R.; Liu, M.; Liu, Y. Berberine attenuates experimental autoimmune encephalomyelitis in C57 BL/6 mice. PLoS ONE 2010, 5, e13489. [Google Scholar] [CrossRef]
- Kim, B.Y.; Park, H.R.; Jeong, H.G.; Kim, S.W. Berberine reduce allergic inflammation in a house dust mite allergic rhinitis mouse model. Rhinology 2015, 53, 353–358. [Google Scholar] [CrossRef]
- Sun, H.-X. Haemolytic activities and adjuvant effect of Bupleurum chinense saponins on the immune responses to ovalbumin in mice. Vaccine 2006, 24, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Piao, C.H.; Hyeon, E.; Fan, Y.; Choi, D.W.; Jung, S.Y.; Jang, B.H.; Shin, H.S.; Song, C.H.; Chai, O.H. Preventive Effect of Bupleurum chinense on Nasal Inflammation via Suppressing T Helper Type 2, Eosinophil and Mast Cell Activation. Am. J. Chin. Med. 2019, 47, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Kirtikar, K.; Basu, B. Indian Medicinal Plants; LM Basu: Allahabad, India, 1975. [Google Scholar]
- Warrier, P.K. Indian Medicinal Plants: A Compendium of 500 Species; Orient Blackswan: Andhra Pradesh, India, 1993; Volume 5. [Google Scholar]
- Khory, R.N.; Katrak, N.N. Materica Medica of India and Their Therapeutics; Times of India Press: Mumbai, India, 1903. [Google Scholar]
- Aswar, U.M.; Kandhare, A.D.; Mohan, V.; Thakurdesai, P.A. Anti-allergic effect of intranasal administration of type-A procyanidin polyphenols based standardized extract of cinnamon bark in ovalbumin sensitized BALB/c mice. Phytother. Res. 2015, 29, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-H.; Bae, J.-H.; Hong, D.-P.; Choi, K.-D.; Kim, S.-C.; Her, E.; Kim, S.-H.; Kang, C.-D. Dryopteris crassirhizoma has anti-cancer effects through both extrinsic and intrinsic apoptotic pathways and G0/G1 phase arrest in human prostate cancer cells. J. Ethnopharmacol. 2010, 130, 248–254. [Google Scholar] [CrossRef]
- Piao, C.H.; Kim, T.G.; Bui, T.T.; Song, C.H.; Shin, D.U.; Eom, J.E.; Lee, S.Y.; Shin, H.S.; Chai, O.H. Ethanol extract of Dryopteris crassirhizoma alleviates allergic inflammation via inhibition of Th2 response and mast cell activation in a murine model of allergic rhinitis. J. Ethnopharmacol. 2019, 232, 21–29. [Google Scholar] [CrossRef]
- Fu, M.; Fu, S.; Ni, S.; Zou, L.; Liu, Y.; Hong, T. Anti-inflammatory effect of epigallocatechin gallate in a mouse model of ovalbumin-induced allergic rhinitis. Int. Immunopharmacol. 2017, 49, 102–108. [Google Scholar] [CrossRef]
- Mateos-Aparicio, I.; Cuenca, A.R.; Villanueva-Suárez, M.; Zapata-Revilla, M. Soybean, a promising health source. Nutr. Hosp. 2008, 23, 305–312. [Google Scholar]
- Lund, C.O.; Mortensen, A.; Nilas, L.; Breinholt, V.M.; Larsen, J.-J.; Ottesen, B. Estrogen and phytoestrogens: Effect on eNOS expression and in vitro vasodilation in cerebral arteries in ovariectomized Watanabe heritable hyperlipidemic rabbits. Eur. J. Obstet. Gynecol. Reprod. Biol. 2007, 130, 84–92. [Google Scholar] [CrossRef]
- Katayanagi, Y.; Yasui, K.; Ikemoto, H.; Taguchi, K.; Fukutomi, R.; Isemura, M.; Nakayama, T.; Imai, S. The clinical and immunomodulatory effects of green soybean extracts. Food Chem. 2013, 138, 2300–2305. [Google Scholar] [CrossRef]
- Bang, J.S.; Choi, H.M.; Sur, B.-J.; Lim, S.-J.; Kim, J.Y.; Yang, H.-I.; Yoo, M.C.; Hahm, D.-H.; Kim, K.S. Anti-inflammatory and antiarthritic effects of piperine in human interleukin 1β-stimulated fibroblast-like synoviocytes and in rat arthritis models. Arthritis Res. Ther. 2009, 11, R49. [Google Scholar] [CrossRef]
- Butt, M.S.; Pasha, I.; Sultan, M.T.; Randhawa, M.A.; Saeed, F.; Ahmed, W. Black pepper and health claims: A comprehensive treatise. Crit. Rev. Food Sci. Nutr. 2013, 53, 875–886. [Google Scholar] [CrossRef]
- Meghwal, M.; Goswami, T. Piper nigrum and piperine: An update. Phytother. Res. 2013, 27, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Piao, C.H.; Hyeon, E.; Fan, Y.; Van Nguyen, T.; Jung, S.Y.; Choi, D.W.; Lee, S.Y.; Shin, H.S.; Song, C.H.; et al. The protective role of Piper nigrum fruit extract in an ovalbumin-induced allergic rhinitis by targeting of NFkappaBp65 and STAT3 signalings. Biomed. Pharm. 2019, 109, 1915–1923. [Google Scholar] [CrossRef]
- Aswar, U.; Shintre, S.; Chepurwar, S.; Aswar, M. Antiallergic effect of piperine on ovalbumin-induced allergic rhinitis in mice. Pharm. Biol. 2015, 53, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.D.S.; Seixas, F.R.M.S.; Aquino Neto, F.R.d. Própolis: 100 anos de pesquisa e suas perspectivas futuras. Química Nova 2002, 25, 321–326. [Google Scholar] [CrossRef]
- Wollenweber, E.; Hausen, B.; Greenaway, W. Phenolic constituents and sensitizing properties of propolis, poplar balsam and balsam of Peru. Bull. Liaison-Groupe Polyphen. 1990, 15, 112–120. [Google Scholar]
- Dimov, V.; Ivanovska, N.; Bankova, V.; Popov, S. Immunomodulatory action of propolis: IV. Prophylactic activity against gram-negative infections and adjuvant effect of the water-soluble derivative. Vaccine 1992, 10, 817–823. [Google Scholar] [CrossRef]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Shimizu, T.; Hino, A.; Tsutsumi, A.; Park, Y.K.; Watanabe, W.; Kurokawa, M. Anti-influenza virus activity of propolis in vitro and its efficacy against influenza infection in mice. Antivir. Chem. Chemother. 2008, 19, 7–13. [Google Scholar] [CrossRef]
- Park, Y.K.; Koo, M.H.; Abreu, J.A.; Ikegaki, M.; Cury, J.A.; Rosalen, P.L. Antimicrobial activity of propolis on oral microorganisms. Curr. Microbiol. 1998, 36, 24–28. [Google Scholar] [CrossRef]
- Yasar, M.; Savranlar, Y.; Karaman, H.; Sagit, M.; Silici, S.; Ozcan, I. Effects of propolis in an experimental rat model of allergic rhinitis. Am. J. Otolaryngol. 2016, 37, 287–293. [Google Scholar] [CrossRef]
- Song, C.H.; Bui, T.T.; Piao, C.H.; Shin, H.S.; Shon, D.-H.; Han, E.-H.; Kim, H.T.; Chai, O.H. Rosae Multiflorae fructus hot water extract inhibits a murine allergic asthma via the suppression of Th2 cytokine production and histamine release from mast cells. J. Med. Food 2016, 19, 853–859. [Google Scholar] [CrossRef]
- Bui, T.T.; Kwon, D.A.; Choi, D.W.; Jung, S.Y.; Lee, S.Y.; Piao, C.H.; Hyeon, E.; Fan, Y.; Yeon, S.H.; Son, R.H.; et al. Rosae multiflorae fructus extract and its four active components alleviate ovalbumin-induced allergic inflammatory responses via regulation of Th1/Th2 imbalance in BALB/c rhinitis mice. Phytomedicine 2019, 55, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.N.; Lomunova, M.; Vu, T.P.D.; Le, B.V.; Kim, Y.H.; Kang, J.S.; Hwang, I. Anti-allergic effects of the ethanol extract of Syzygium formosum (Wall.) Masam leaves and its immunoregulatory mechanisms. J. Ethnopharmacol. 2018, 211, 171–179. [Google Scholar] [CrossRef]
- Zhuang, Y.; Qin, K.; Yu, B.; Liu, X.; Cai, B.; Cai, H. A metabolomics research based on UHPLC-ESI-Q-TOF-MS coupled with metabolic pathway analysis: Treatment effects of stir-frying Xanthii Fructus on allergic rhinitis in mice model. Biomed. Chromatogr. 2018, 32, e4352. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Kobayashi, M.; Uchida, T.; Shimizu, E.; Higashio, H.; Ohno, M.; Uesugi, S.; Kimura, K.I. Anti-allergy activities of Kuji amber extract and kujigamberol. Fitoterapia 2018, 127, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Taniguchi, M.; Baba, K. Antitumor and antimetastatic effects on liver of triterpenoid fractions of Ganoderma lucidum: Mechanism of action and isolation of an active substance. Anticancer Res. 2002, 22, 3309–3318. [Google Scholar] [PubMed]
- Maruyama, H.; Yamazaki, K.; Murofushi, S.; Konda, C.; Ikekawa, T. Antitumor activity of Sarcodon aspratus (Berk.) S. Ito and Ganoderma lucidum (Fr.) Karst. J. Pharm. 1989, 12, 118–123. [Google Scholar] [CrossRef]
- Min, B.S.; Gao, J.J.; Nakamura, N.; Hattori, M. Triterpenes from the spores of Ganoderma lucidum and their cytotoxicity against meth-A and LLC tumor cells. Chem. Pharm. Bull. (Tokyo) 2000, 48, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, N.; Nabe, T.; Shimazu, M.; Yoshino, S.; Kohno, S. Effect of Ganoderma lucidum on pollen-induced biphasic nasal blockage in a guinea pig model of allergic rhinitis. Phytother. Res. 2012, 26, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Im, Y.S.; Lee, B.; Kim, E.Y.; Min, J.H.; Song, D.U.; Lim, J.M.; Eom, J.W.; Cho, H.J.; Sohn, Y.; Jung, H.S. Antiallergic effect of gami-hyunggyeyeongyotang on ovalbumin-induced allergic rhinitis in mouse and human mast cells. J. Chin. Med. Assoc. 2016, 79, 185–194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang, F.; Tang, Q.; Tian, Y.; Fan, Q.; Huang, Y.; Tan, X. Network pharmacology-based prediction of the active ingredients and potential targets of Mahuang Fuzi Xixin decoction for application to allergic rhinitis. J. Ethnopharmacol. 2015, 176, 402–412. [Google Scholar] [CrossRef]
- Morikura, I.; Murata, A.; Aoi, N.; Shimizu, Y.; Fuchiwaki, T.; Prokopakis, E.; Kawauchi, H. Japanese traditional medicine, Senn-kinn-naidaku-sann up-regulates Toll-like receptor 4 and reduces murine allergic rhinitis. Rhinology 2014, 52, 252–259. [Google Scholar] [CrossRef]
- Shao, Y.Y.; Zhou, Y.M.; Hu, M.; Li, J.Z.; Chen, C.J.; Wang, Y.J.; Shi, X.Y.; Wang, W.J.; Zhang, T.T. The Anti-Allergic Rhinitis Effect of Traditional Chinese Medicine of Shenqi by Regulating Mast Cell Degranulation and Th1/Th2 Cytokine Balance. Molecules 2017, 22, 504. [Google Scholar] [CrossRef]
- Jeong, K.T.; Kim, S.G.; Lee, J.; Park, Y.N.; Park, H.H.; Park, N.Y.; Kim, K.J.; Lee, H.; Lee, Y.J.; Lee, E. Anti-allergic effect of a Korean traditional medicine, Biyeom-Tang on mast cells and allergic rhinitis. BMC Complement. Altern. Med. 2014, 14, 54. [Google Scholar] [CrossRef]
- Xiang, M.; Wu, L.; Su, H.; Han, B.; Liu, H.; Xiao, X.; Yin, X.; Fan, Y.; Zhang, L.; Huang, Y.; et al. Biyuanling suppresses the toluene-2, 4-diisocyanate induced allergic rhinitis in guinea pigs. Oncotarget 2018, 9, 12620–12629. [Google Scholar] [CrossRef]
- Xie, M.Q.; Liu, J.; Long, Z.; Tian, D.F.; Zhao, C.Q.; Yang, P.C. Modulation of immune tolerance with a Chinese traditional prescription inhibits allergic rhinitis in mice. N. Am. J. Med. Sci. 2011, 3, 503–507. [Google Scholar] [CrossRef]
- Hong, S.H.; Kim, S.R.; Choi, H.S.; Ku, J.M.; Seo, H.S.; Shin, Y.C.; Ko, S.G. Effects of Hyeonggaeyeongyo-tang in ovalbumin-induced allergic rhinitis model. Mediat. Inflamm. 2014, 2014, 418705. [Google Scholar] [CrossRef]
- Kim, C.M.; Heo, M.Y.; Kim, H.P.; Sin, K.S.; Pachaly, P. Pharmacological activities of water extracts ofUmbelliferae plants. Arch. Pharmacal Res. 1991, 14, 87. [Google Scholar] [CrossRef]
- Na, H.J.; Jeong, H.J.; Hong, S.H.; Hwang, C.Y.; Cha, W.S.; Kim, H.M. Effect of Boo Yong-Tang on Mast Cell-Mediated Allergic Reaction. Immunopharmacol. Immunotoxicol. 2004, 26, 445–454. [Google Scholar] [CrossRef]
- Shin, S. In vitro effects of essential oils fromOstericum koreanum against antibiotic-resistantSalmonella spp. Arch. Pharmacal Res. 2005, 28, 765–769. [Google Scholar] [CrossRef]
- Won Jung, H.; Jung, J.K.; Weon Cho, C.; Kang, J.S.; Park, Y.K. Antiallergic effect of KOB03, a polyherbal medicine, on mast cell-mediated allergic responses in ovalbumin-induced allergic rhinitis mouse and human mast cells. J. Ethnopharmacol. 2012, 142, 684–693. [Google Scholar] [CrossRef]
- Jung, H.W.; Jung, J.K.; Park, Y.K. Comparison of the efficacy of KOB03, ketotifen, and montelukast in an experimental mouse model of allergic rhinitis. Int. Immunopharmacol. 2013, 16, 254–260. [Google Scholar] [CrossRef]
- Ren, M.; Tang, Q.; Chen, F.; Xing, X.; Huang, Y.; Tan, X. Mahuang Fuzi Xixin Decoction Attenuates Th1 and Th2 Responses in the Treatment of Ovalbumin-Induced Allergic Inflammation in a Rat Model of Allergic Rhinitis. J. Immunol. Res. 2017, 2017, 8254324. [Google Scholar] [CrossRef]
- Oh, H.A.; Kim, H.M.; Jeong, H.J. Alleviation of allergic rhinitis symptoms with Pyeongwee-San extract (KMP6). Immunopharmacol. Immunotoxicol. 2012, 34, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Kong, Y.; Li, J.; Wang, Y.; Li, M.; Wang, Y.; Wang, Y.; Ren, F.; Ni, J.; Li, Y.; et al. Mesenchymal stem cells combined with traditional Chinese medicine (qi-fang-bi-min-tang) alleviates rodent allergic rhinitis. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ku, J.M.; Hong, S.H.; Kim, S.R.; Choi, H.S.; Seo, H.S.; Jang, B.H.; Ko, S.G.; Shin, Y.C. Anti-allergic effects of So-Cheong-Ryong-Tang in ovalbumin-induced allergic rhinitis model. Eur. Arch. Otorhinolaryngol. 2016, 273, 123–131. [Google Scholar] [CrossRef]
- Mo, J.H.; Lee, S.E.; Wee, J.H.; Lee, J.E.; Rhee, C.S.; Lee, C.H.; Kim, D.Y. Anti-allergic effects of So-Cheong-Ryong-Tang, a traditional Korean herbal medicine, in an allergic rhinitis mouse model. Eur. Arch. Otorhinolaryngol. 2013, 270, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Y.; Liu, X.; Dai, L.B.; Zhou, S.H. Effect of Tong Qiao drops on the expression of eotaxin, IL-13 in the nasal mucosa of rats with allergic rhinitis. J. Chin. Med. Assoc. 2012, 75, 524–529. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ai, S.; Zheng, J.; Chu, K.D.; Zhang, H.S. Effects of Xingbi gel on leukotriene E4 and immunoglobulin E production and nasal eosinophilia in a guinea pig model for allergic rhinitis. Asian Pac. J. Allergy Immunol. 2015, 33, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, Y.; Yan, S.; Yan, D. Yiqi Wenyang Fang ameliorates allergic rhinitis through inhibiting inflammatory response and promoting the expression of Foxp3. Int. J. Immunopathol. Pharm. 2016, 29, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Principal Investigator: Pedro Guardia, D.H.V.M.; Principal Investigator: Joaquín Quiralte, D.H.V.D.R.; Principal Investigator: Luis Angel Navarro, D.H.L.A.; Principal Investigator: Santiago Nevot, D.H.M. Safety Clinical Trial with Depigopid 50% grasses/50% Olea Europaea (2000 dpp/mL) or depiGoid 50% Grasses/50% Parietaria Judaica (2000 dpp/mL). Available online: https://www.clinicaltrials.gov/ct2/show/NCT01734265?term=NCT01734265&draw=2&rank=1 (accessed on 29 January 2020).
- Merck Sharp & Dohme Corp. Safety Study of mk-3641 and mk-7243 Co-Administered in Adult Participants with Ragweed and Grass Pollen Induced Allergic Rhinitis (p08607, mk-3641-006). Available online: https://www.clinicaltrials.gov/ct2/show/NCT02256553?term=NCT02256553&draw=2&rank=1 (accessed on 3 March 2020).
- Merck Sharp & Dohme Corp. Efficacy and Safety Study of Ragwitek™ (mk-3641) in Children with Ragweed-Induced Rhinoconjunctivitis with or without Asthma (mk-3641-008). Available online: https://www.clinicaltrials.gov/ct2/show/NCT02478398?term=NCT02478398&draw=1&rank=1 (accessed on 6 September 2019).
- Pfaar, O.; van Twuijver, E.; Boot, J.D.; Opstelten, D.J.; Klimek, L.; van Ree, R.; Diamant, Z.; Kuna, P.; Panzner, P. A randomized DBPC trial to determine the optimal effective and safe dose of a SLIT-birch pollen extract for the treatment of allergic rhinitis: Results of a phase II study. Allergy 2016, 71, 99–107. [Google Scholar] [CrossRef]
- Study Chair: Walter G. Canonica, P.D.A.a.R.D.U.o.G. Sublivac Fix Phleum Pratense dt/drf. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01682070?term=NCT01682070&draw=1&rank=1 (accessed on 29 May 2019).
- Principal Investigator: Peter Couroux, M.I. Efficacy and Safety of Sublivac Phleum for Immunotherapy of Grass Pollen-Allergy. Available online: https://www.clinicaltrials.gov/ct2/show/NCT02556801?term=NCT02556801&draw=1&rank=1 (accessed on 13 September 2019).
- Wood, R.A.; Tgias, A.; Wildfire, J.; Visness, C.M.; Matsui, E.C.; Gruchalla, R.; Hershey, G.; Liu, A.H.; O’Connor, G.T.; Pongracic, J.A.; et al. Development of cockroach immunotherapy by the Inner-City Asthma Consortium. J. Allergy Clin. Immunol. 2014, 133, 846–852.e6. [Google Scholar] [CrossRef]
- Study Chair: Robert Wood, M.D.J.H.C.s.C.D.o.A.I. Subcutaneous Immunotherapy for Mouse in Adults (Scitmo). Available online: https://www.clinicaltrials.gov/ct2/show/NCT02532179?term=NCT02532179&draw=1&rank=1 (accessed on 7 March 2020).
- Iwasaki, M.; Saito, K.; Takemura, M.; Sekikawa, K.; Fujii, H.; Yamada, Y.; Wada, H.; Mizuta, K.; Seishima, M.; Ito, Y. TNF-α contributes to the development of allergic rhinitis in mice. J. Allergy Clin. Immunol. 2003, 112, 134–140. [Google Scholar] [CrossRef]
- Jung, H.W.; Jung, J.K.; Kim, Y.H.; Kang, J.S.; Park, Y.K. Effect of KOB03, a polyherbal medicine, on ovalbumin-induced allergic rhinitis in guinea pigs. Chin. Med. 2012, 7, 27. [Google Scholar] [CrossRef]
- Tanaka, K.-I.; Okamoto, Y.; Nagaya, Y.; Nishimura, F.; Takeoka, A.; Hanada, S.; Kohno, S.; Kawai, M. A nasal allergy model developed in the guinea pig by intranasal application of 2, 4-toluene diisocyanate. Int. Arch. Allergy Immunol. 1988, 85, 392–397. [Google Scholar] [CrossRef]





| Classification | Compound | Source | Cell Line/Animal Model | Dose/Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Mineral | Kuji amber methanol extract | Succinum | RBL-2H3 | 2 μL (0–25 μg/mL); 30 min | Inhibition of the degranulation of RBL-2H3 cells | ↓ p-ERK1/2, LTC4 | [106] |
| Mineral | Kujigamberol (15,20-dinor-5,7,9-labdatrien-18-ol) | Succinum | RBL-2H3 | 2 μL (0–25 μg/mL); 30 min | Inhibition of the degranulation of RBL-2H3 cells | ↓ p-ERK1/2, LTC4 | [106] |
| Classification | Compound | Source | Cell Line/Animal Model | Dose/Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Fungi | Ganoderma lucidum powder suspended in water | Ganoderma lucidum | Cedar pollen extracts-sensitized AR guinea pigs | 100, 1000 mg/5 mL/kg; 8 weeks | Inhibition of nasal inflammation | ↓ CysLT1 | [110] |
| Mineral | Kuji amber methanol extract | Succinum | OVA-induced rhinitis guinea pigs | 20 μg; 10 min, 1 h | Inhibition of nasal blockade | ↓ p-ERK1/2, LTC4 | [106] |
| Mineral | Kujigamberol (15,20-dinor-5,7,9-labdatrien-18-ol) | Succinum | OVA-induced rhinitis guinea pigs | 20 μg; 10 min, 1 h | Inhibition of nasal blockade | ↓ p-ERK1/2, LTC4 | [106] |
| Name | Constitutions | Experimental Subject | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Gami-hyunggyeyeongyotang | Schizonepetae Spica (Nepeta tenuifolia Benth.), Forsythiae Fructus (Forsythia viridissima Lindl.), Saposhnikoviae Radix (Saposhnikovia divaricata (Turcz.) Schischk.), Angelicae Radix (Angelica acutiloba Kitagawa), Cnidii Rhizoma (Cnidium officinale Makino), Paeoniae Radix Alba (Paeonia lactiflora Pallas), Angelicae Dahuricae Radix (Angelica dahurica Bentham et Hooker), Bupleuri Radix (Bupleurum falcatum Linne), Scutellariae Radix (Scutellaria baicalensis Georgi), Aurantii Fructus Immaturus (Citrus aurantium Linne), Gardeniae Frutus (Gardenia jasminoides Ellis), Platycodi Radix (Platycodon grandiflorum A. De candole), Glycyrrhizae Radix (Glycyrrhiza uralensis Fischer), Ulmi Cortex (Ulmus macrocarpa Hance), Xanthii Fructus (Xanthium strumarium Linne), Magnoliae Flos (Magnolia kobus De Candolle), Rubiae Radix (Rubia akane Nakai) | HMC-1 | 1, 10, 100 μg/mL; 1 h | Alleviation of the symptoms of AR | ↓ IgE, IL-5, IL-6, IL-1β, MCP-1, MIP-2, caspase-1 | [111] |
| Mahuang Fuzi Xixin Decoction | Ephedrae Herba (Ephedra sinica Stapf, Ephedra intermedia Schrenk & C.A. Mey. or Ephedra equisetina Bunge), Aconiti Lateralis Radix (Aconitum carmichaelii Debeaux), Asiasari Radix (Asarum heterotropoides f. mandshuricum (Maxim.) Kitag.) | IgE-activated RBL-2H3 | M136: 3, 10, 30 μM; 1 h | Anti-inflammatory effect | ↓ PGE2 | [112] |
| M188: 5, 10 μM; 1 h | ||||||
| M234: 2, 5 μM; 1 h | ||||||
| M289: 3, 10, 30 μM; 1 h | ||||||
| Senn-kinn-naidaku-sann | Ginseng Radix, Angelicae Radix, Astragali Radix, Cnidii Rhizoma, Sinomeni Caulis et Rhizoma, Platycodonis Radix, Magnoliae Cortex, Angelicae Dahuricae Radix, Glycyrrhizae Radix | RAW264.7 | 0.01, 0.1, 1, 10 μg/mL; 24 h | Inhibition of inflammatory response | ↑ TLR4, IL-12 | [113] |
| Shenqi | Angelicae Dahuricae Radix (Angelica dahurica (Hoffm.) Benth. & Hook.f. ex Franch. & Sav.), Scutellariae Radix (Scutellaria baicalensis Georgi), Lonicerae Japonicae Flos (Lonicera japonica Thunb.), Menthae Haplocalycis Herba (Mentha canadensis L.), Astragali Radix (Astragalus propinquus Schischkin), Codonopsis Radix (Codonopsis pilosula subsp. tangshen (Oliv.) D.Y.Hong), Isatidis Folium (Isatis tinctoria L.), Taraxaci Herba (Taraxacum mongolicum Hand.-Mazz.), Agrimoniae Herba (Agrimonia pilosa Ledeb.), Asiasari Radix et Rhizome (Asarum sieboldii Miq.) | RBL-2H | 50, 100, 200 μg/mL; 1 h | Alleviation of symptoms of AR | ↓ β-Hex, histamine | [114] |
| Classification | Compound/Extract | Source | Phase | Patients | Status | Registration Number | Reference |
|---|---|---|---|---|---|---|---|
| Plant | Depigoid 50% Grasses/50% Olea europaea (2000DPP/mL), Depigoid 50% Grasses/50% Parietaria judaica (2000DPP/mL) | Grasses, Olea europaea L., Parietaria judaica L. | Phase 2 | 63 | Completed | NCT01734265 | [132] |
| Plant | MK-3641 12 Amb a 1-U (short ragweed extract) MK-7243 2800 BAU (Timothy grass extract) | Ambrosia artemisiifolia L., Phleum pratense L. | Phase 4 | 102 | Completed | NCT02256553 | [133] |
| Plant | Short ragweed pollen allergen extract | Ambrosia artemisiifolia L. | Phase 3 | 1025 | Completed | NCT02478398 | [134] |
| Plant | SUBLIVAC FIX Birch | Betula verrucose Ehrh. | Phase 2 | 269 | Completed | NCT01639768 | [135] |
| Plant | SUBLIVAC FIX Phleum pratense | Phleum pratense L. | Phase 2 | 266 | Completed | NCT01682070 | [136] |
| Plant | SUBLIVAC FIX Phleum Pratense | Phleum pratense L. | Phase 2 | 168 | Completed | NCT02556801 | [137] |
| Animal | German cockroach (Blattella germanica) allergenic extract | Blattella germanica | Phase 1 | 10 | Completed | NCT01221285 | [138] |
| Animal | Mouse Allergenic Extract | Mus musculus | Phase 1 Phase 2 | 12 | Completed | NCT02532179 | [139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.; Jeong, I.; Cho, J.; Shin, C.; Kim, K.-I.; Shim, B.-S.; Ko, S.-G.; Kim, B. The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery. Antioxidants 2021, 10, 1524. https://doi.org/10.3390/antiox10101524
Lim S, Jeong I, Cho J, Shin C, Kim K-I, Shim B-S, Ko S-G, Kim B. The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery. Antioxidants. 2021; 10(10):1524. https://doi.org/10.3390/antiox10101524
Chicago/Turabian StyleLim, Suhyun, Iwah Jeong, Jonghyeok Cho, Chaewon Shin, Kwan-Il Kim, Bum-Sang Shim, Seong-Gyu Ko, and Bonglee Kim. 2021. "The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery" Antioxidants 10, no. 10: 1524. https://doi.org/10.3390/antiox10101524
APA StyleLim, S., Jeong, I., Cho, J., Shin, C., Kim, K.-I., Shim, B.-S., Ko, S.-G., & Kim, B. (2021). The Natural Products Targeting on Allergic Rhinitis: From Traditional Medicine to Modern Drug Discovery. Antioxidants, 10(10), 1524. https://doi.org/10.3390/antiox10101524