

The Instantaneous Effect and Its Mechanism of Transcranial Direct Current Stimulation on Working Memory Based on Delta and Gamma Band Electroencephalography Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
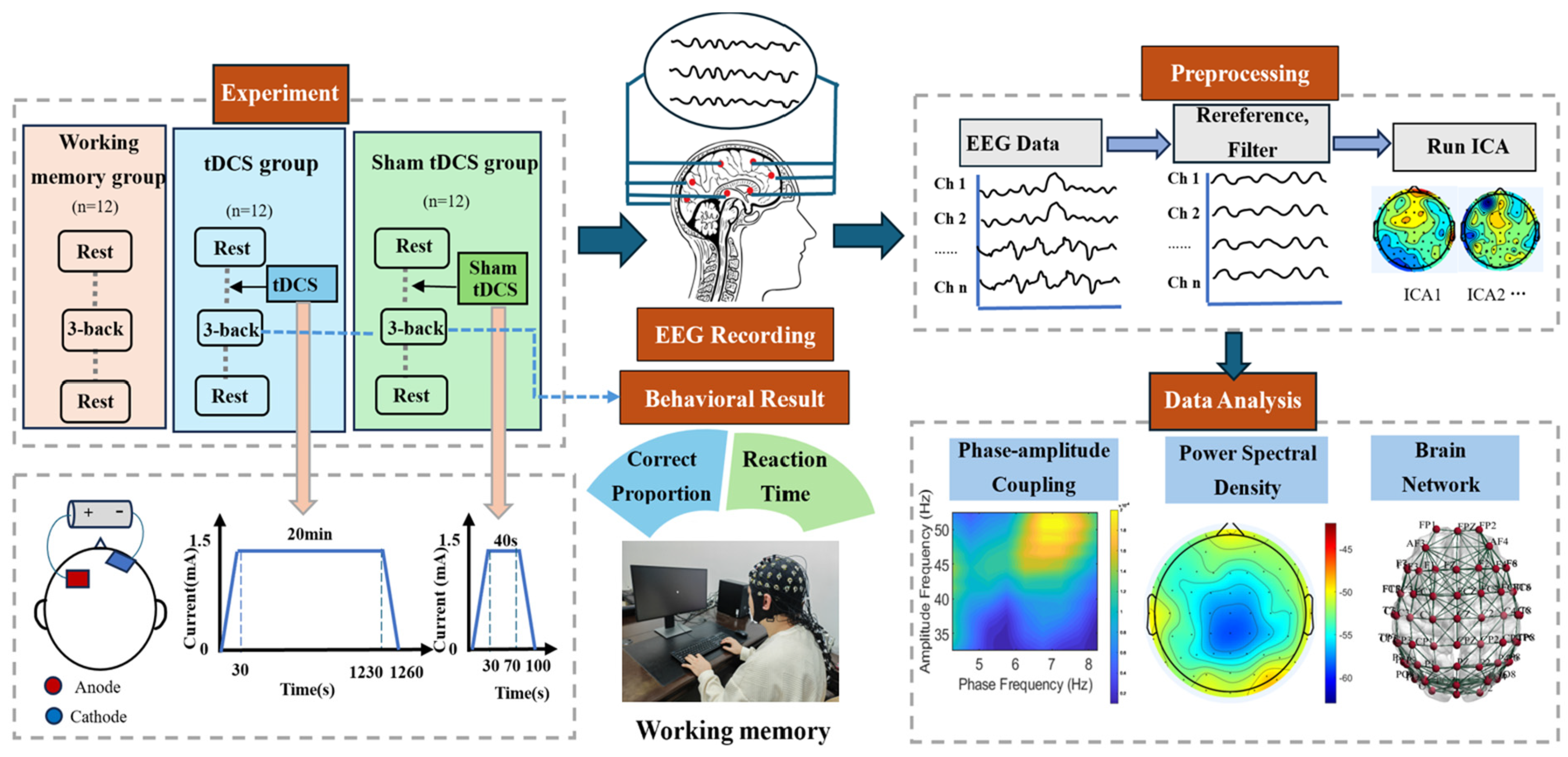
2.1. Experiment
2.1.1. Participant
2.1.2. Experiment Design
2.1.3. tDCS
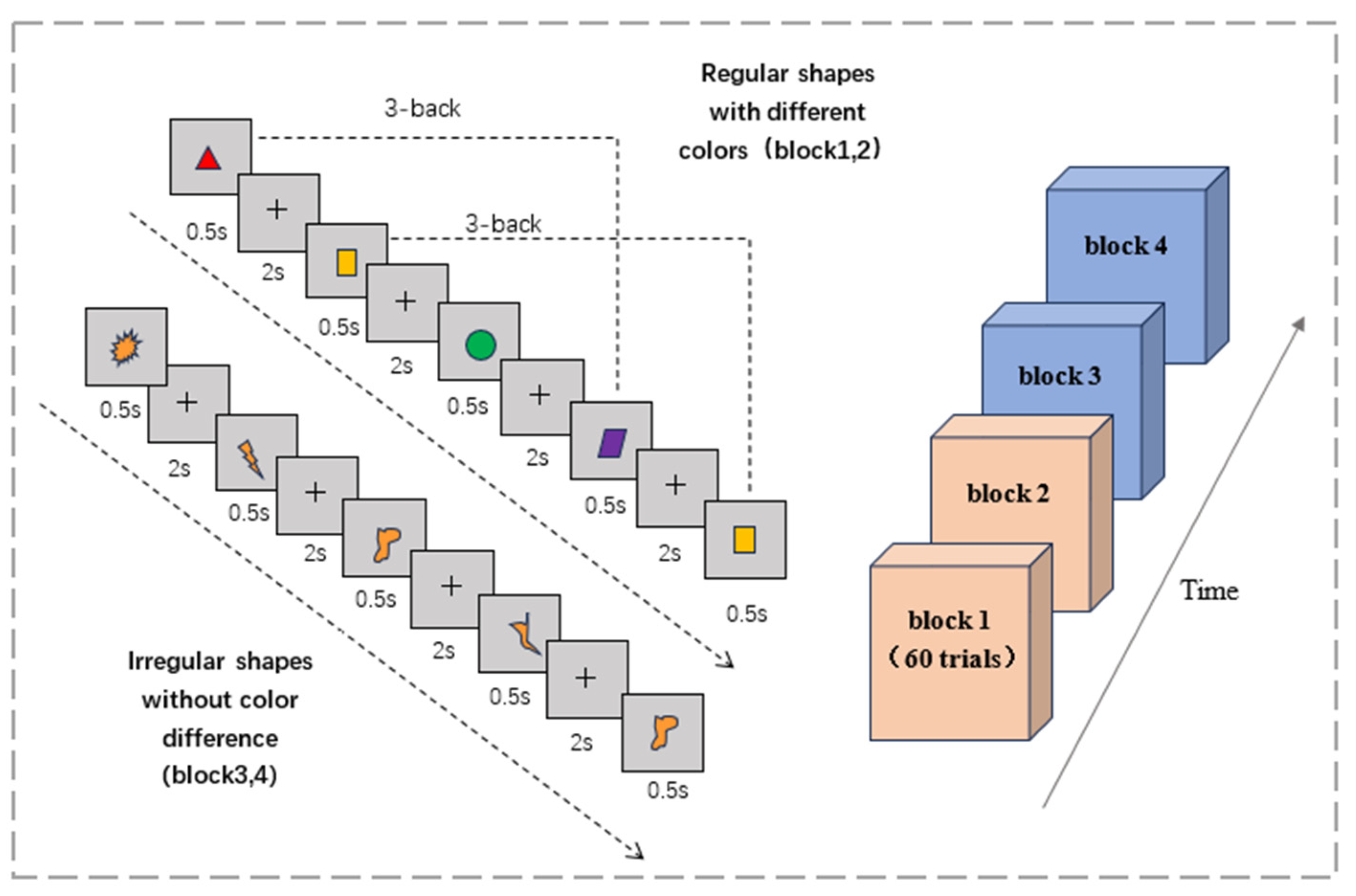
2.1.4. Three-Back Task
2.2. Data Acquisition and Preprocessing
2.3. Analysis
3. Results
3.1. EEG Feature of Working Memory Task States
3.1.1. Data Analysis Based on Phase–Amplitude Coupling
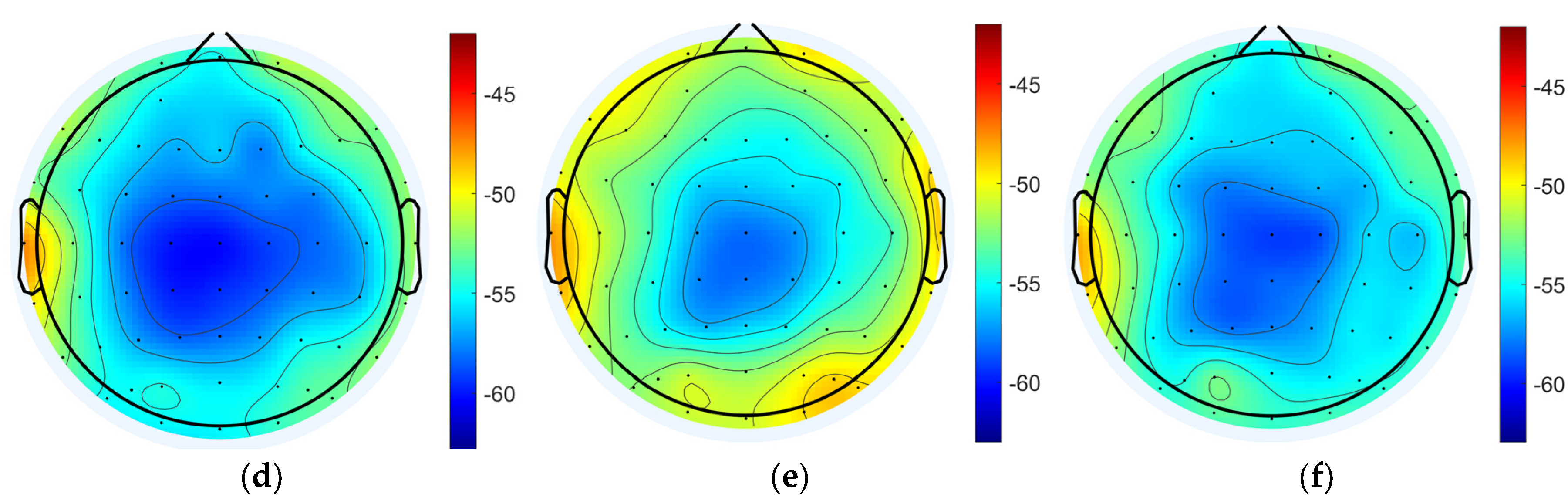
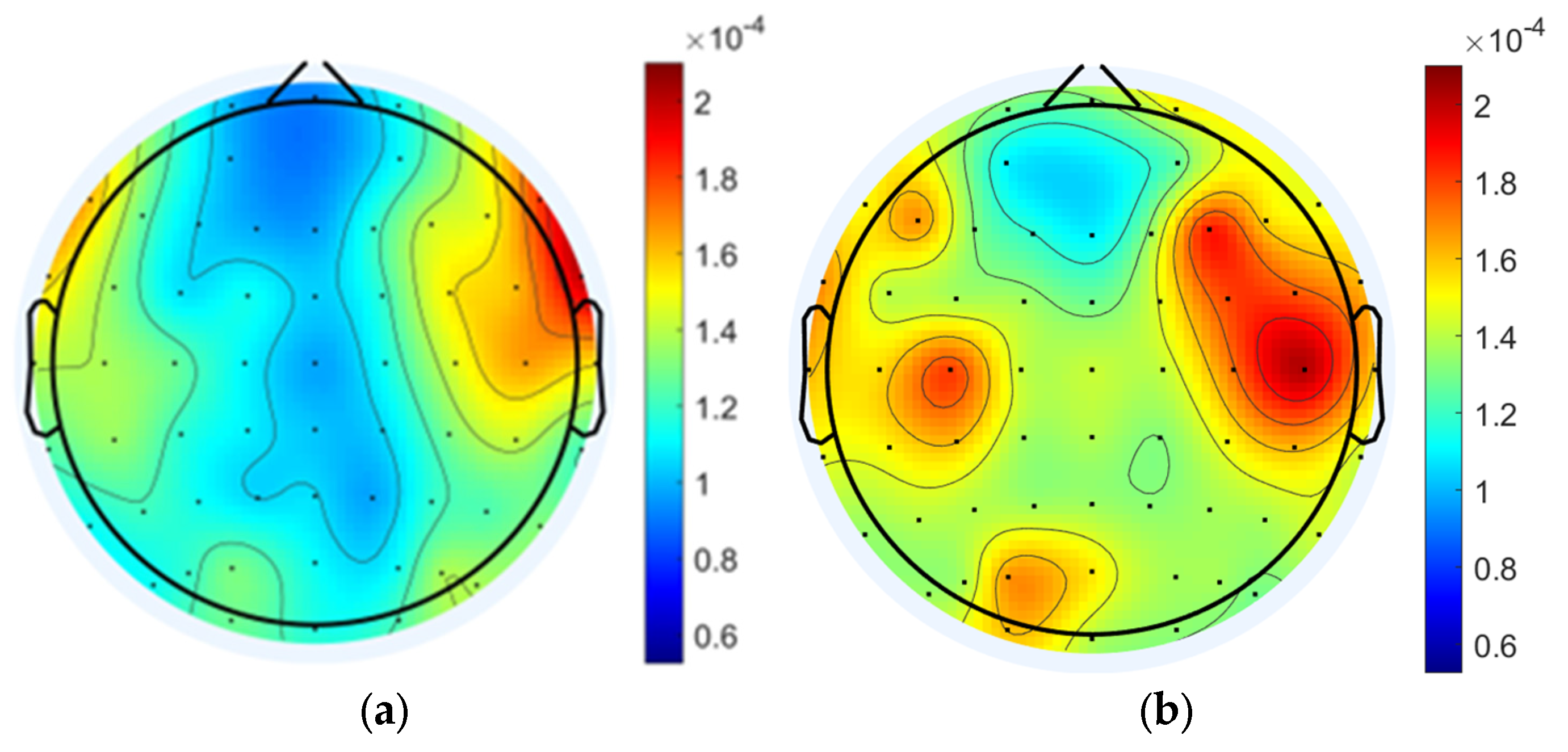
3.1.2. Data Analysis Based on Power Spectral Density
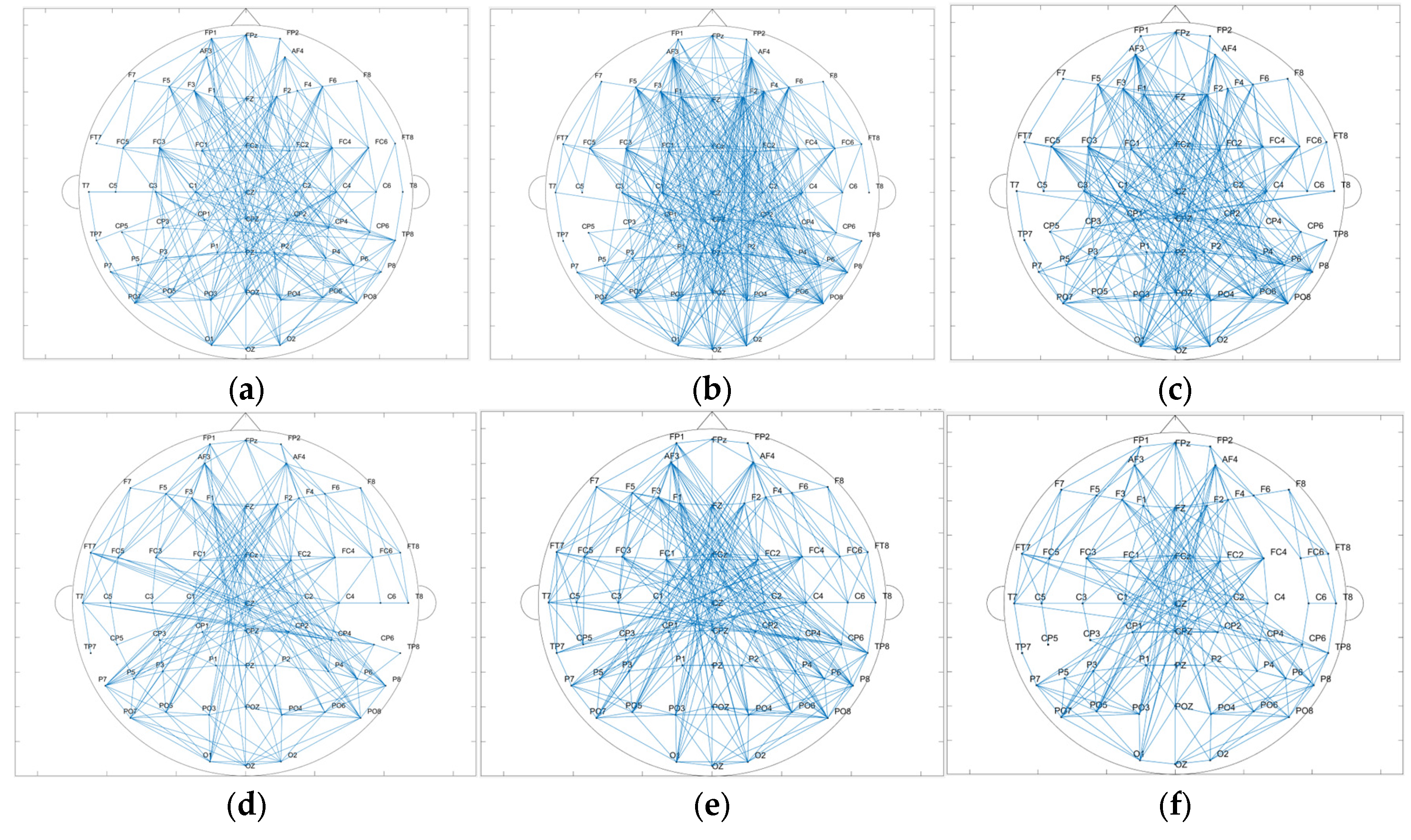
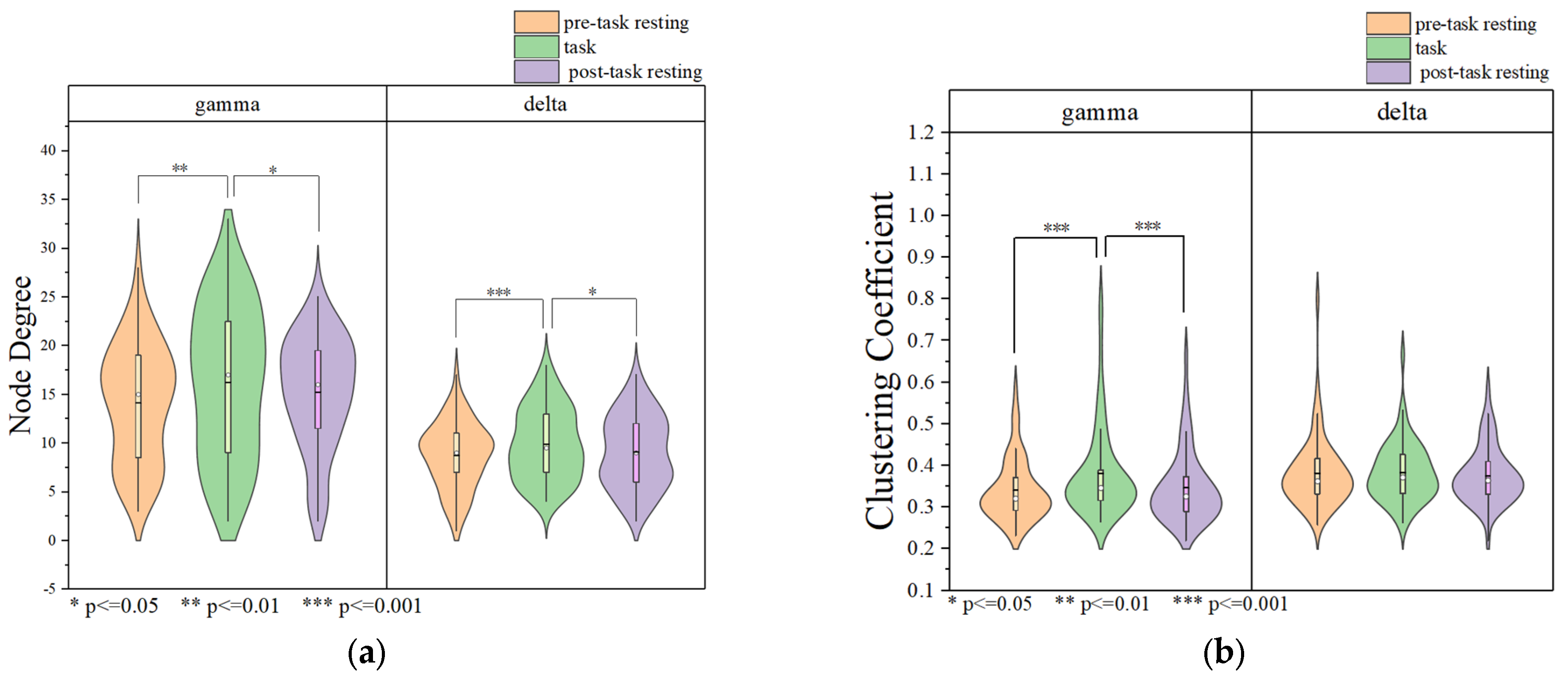
3.1.3. Data Analysis Based on Brain Network
3.2. EEG Feature and Behavioral Result of Working Memory Task in True/Sham tDCS Group
3.2.1. Three-Back Behaviorral Result
3.2.2. Data Analysis Based on Phase–Amplitude Coupling
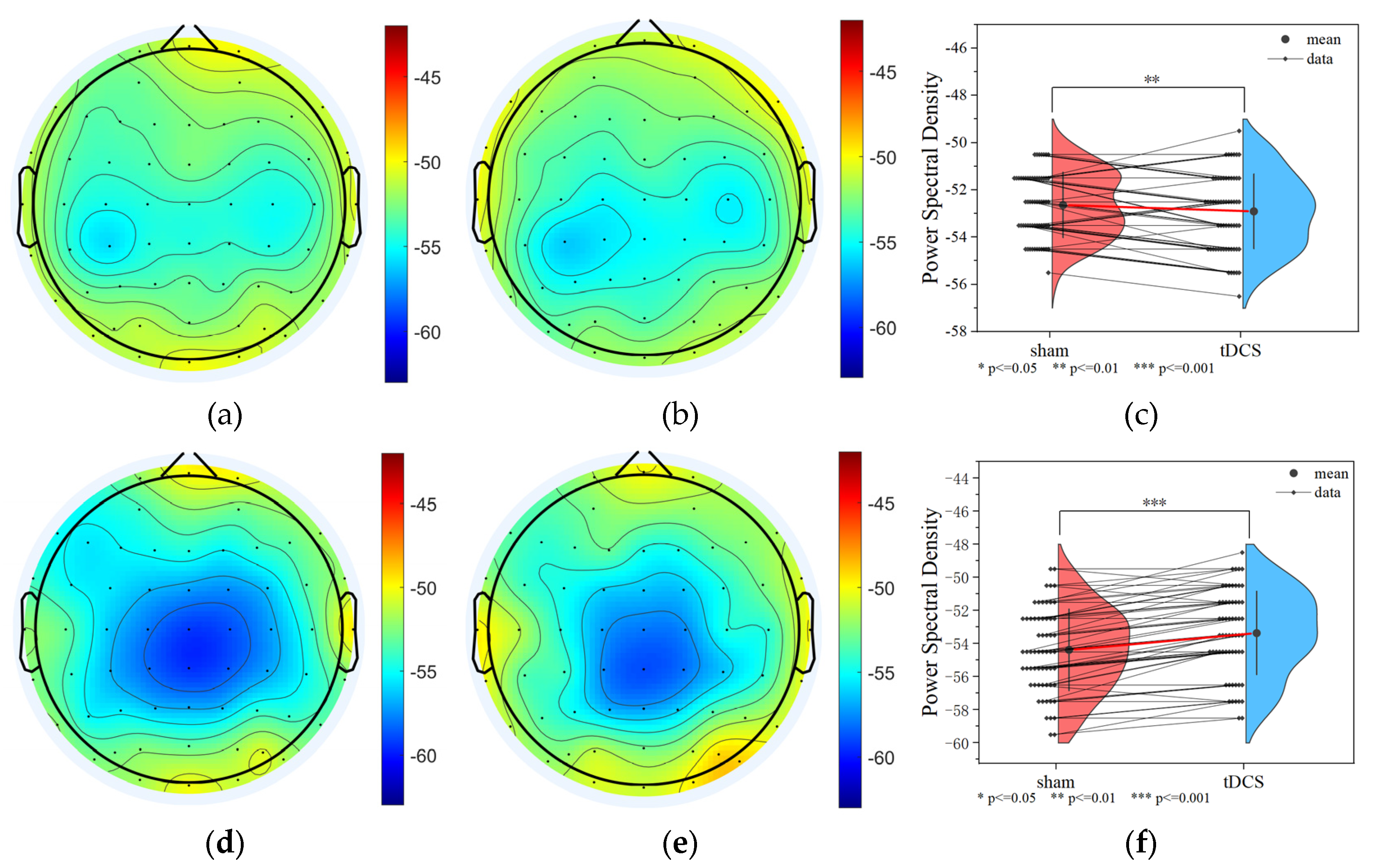
3.2.3. Data Analysis Based on Power Spectral Density
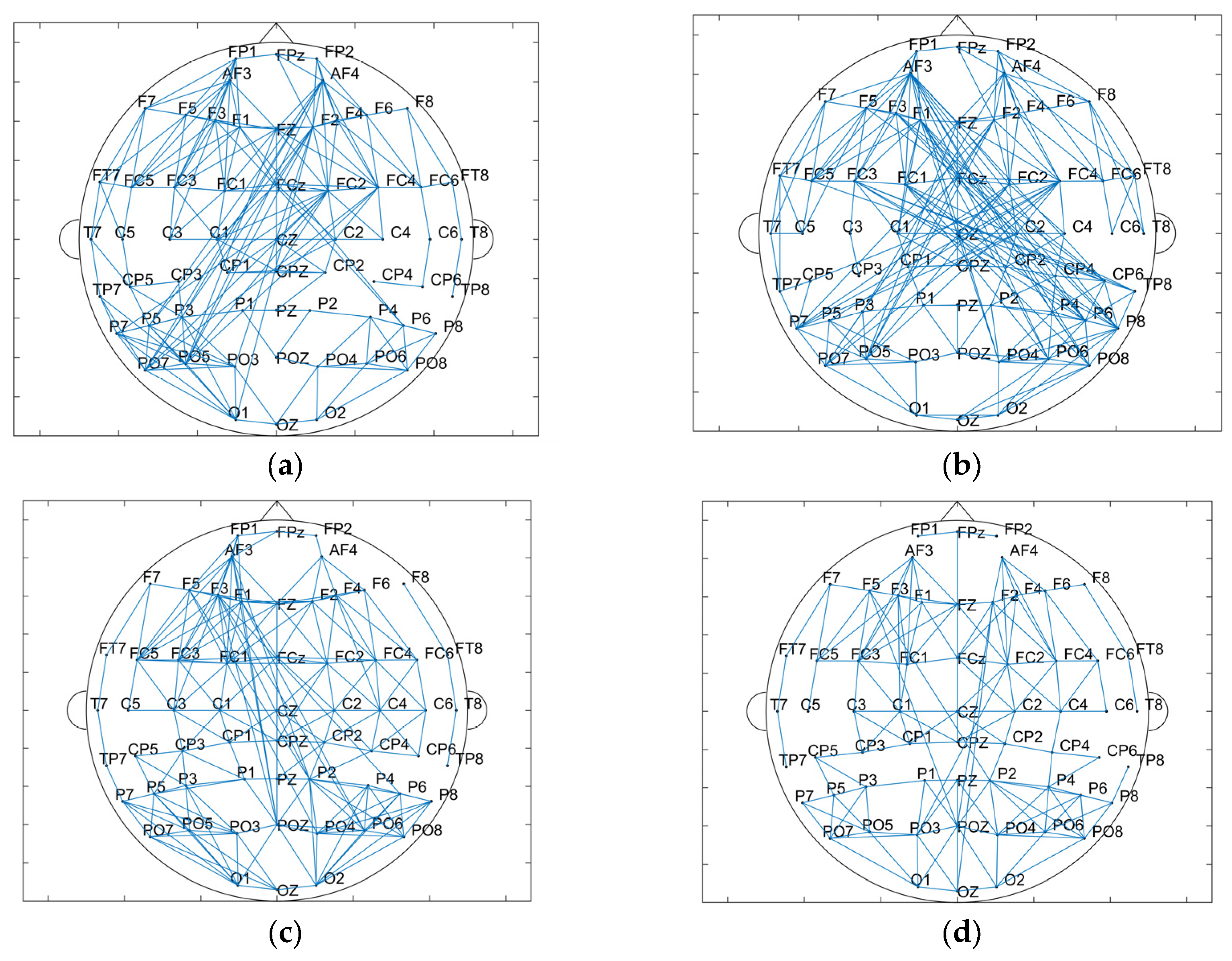
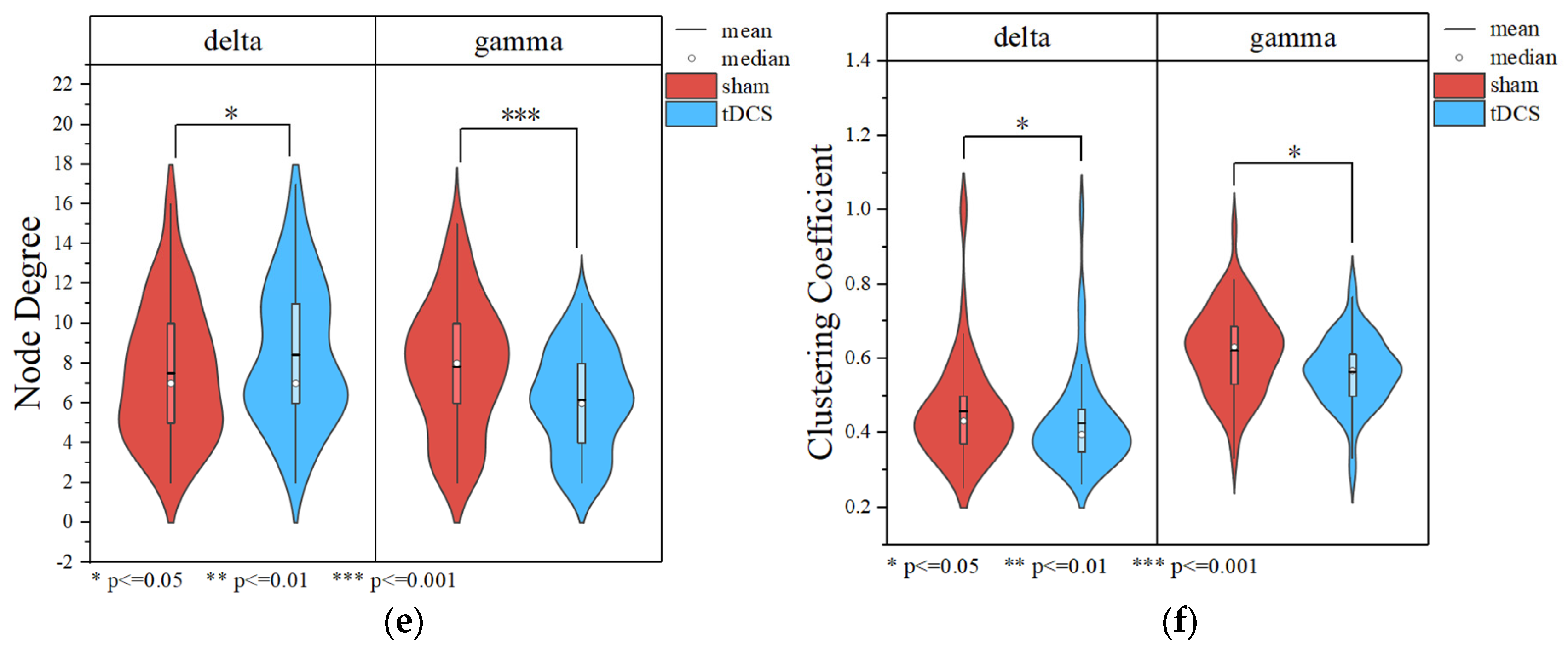
3.2.4. Data Analysis Based on the Brain Network
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bressler, S.L.; Menon, V. Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Ilardi, C.R.; Chieffi, S.; Iachini, T.; Iavarone, A. Neuropsychology of posteromedial parietal cortex and conversion factors from Mild Cognitive Impairment to Alzheimer’s disease: Systematic search and state-of-the-art review. Aging Clin. Exp. Res. 2022, 34, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Gebreegziabhere, Y.; Habatmu, K.; Mihretu, A.; Cella, M.; Alem, A. Cognitive impairment in people with schizophrenia: An umbrella review. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 1139–1155. [Google Scholar] [CrossRef]
- Szyszkowska, J.; Bala, A. The impact of depressive symptomology, rumination and objective memory performance on subjective cognitive complaints. Int. J. Psychophysiol. 2023, 190, 1–7. [Google Scholar] [CrossRef]
- Begemann, M.J.; Brand, B.A.; Ćurčić-Blake, B.; Aleman, A.; Sommer, I.E. Efficacy of non-invasive brain stimulation on cognitive functioning in brain disorders: A meta-analysis. Psychol. Psychol. Med. 2020, 50, 2465–2486. [Google Scholar] [CrossRef]
- Chase, H.W.; Boudewyn, M.A.; Carter, C.S.; Phillips, M.L. Transcranial direct current stimulation: A roadmap for research, from mechanism of action to clinical implementation. Mol. Psychiatry 2020, 25, 397–407. [Google Scholar] [CrossRef]
- Grover, S.; Nguyen, J.A.; Reinhart, R.M.G. Synchronizing Brain Rhythms to Improve Cognition. Annu. Rev. Med. 2021, 72, 29–43. [Google Scholar] [CrossRef]
- Naka, M.; Matsuzawa, D.; Ishii, D.; Hamada, H.; Uchida, T.; Sugita, K.; Sutoh, C.; Shimizu, E. Differential effects of high-definition transcranial direct current stimulation on verbal working memory performance according to sensory modality. Neurosci. Lett. 2018, 687, 131–136. [Google Scholar] [CrossRef]
- Martin, D.M.; Rushby, J.A.; De Blasio, F.M.; Wearne, T.; Osborne-Crowley, K.; Francis, H.; Xu, M.; Loo, C.; McDonald, S. The effect of tDCS electrode montage on attention and working memory. Neuropsychologia 2022, 179, 108462. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Smoot, M.R.; Mehta, R.K. Anodal tDCS augments and preserves working memory beyond time-on-task deficits. Sci. Rep. 2021, 11, 19134. [Google Scholar] [CrossRef]
- Talsma, L.J.; Kroese, H.A.; Slagter, H.A. Boosting Cognition: Effects of Multiple-Session Transcranial Direct Current Stimulation on Working Memory. J. Cogn. Neurosci. 2017, 29, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Cerreta Adelle, G.B.; Mruczek Ryan, E.B.; Berryhill Marian, E. Predicting Working Memory Training Benefits from Transcranial Direct Current Stimulation Using Resting-State fMRI. Front. Psychol. 2020, 11, 570030. [Google Scholar] [CrossRef] [PubMed]
- Kolskår, K.K.; Richard, G.; Alnæs, D.; Dørum, E.S.; Sanders, A.M.; Ulrichsen, K.M.; Sánchez, J.M.; Ihle-Hansen, H.; Nordvik, J.E.; Westlye, L.T. Reliability, sensitivity, and predictive value of fMRI during multiple object tracking as a marker of cognitive training gain in combination with tDCS in stroke survivors. Hum. Brain Mapp. 2021, 42, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Pergher, V.; Au, J.; Alizadeh Shalchy, M.; Santarnecchi, E.; Seitz, A.; Jaeggi, S.M.; Battelli, L. The benefits of simultaneous tDCS and working memory training on transfer outcomes: A systematic review and meta-analysis. Brain Stimul. 2022, 15, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Narmashiri, A.; Akbari, F. The effects of transcranial direct current stimulation (tDCS) on the cognitive functions: A systematic review and meta-analysis. Neuropsychol. Rev. 2023, 35, 126–152. [Google Scholar] [CrossRef]
- Müller, D.; Habel, U.; Brodkin, E.S.; Weidler, C. High-definition transcranial direct current stimulation (HD-tDCS) for the enhancement of working memory—A systematic review and meta-analysis of healthy adults. Brain Stimul. 2022, 15, 1475–1485. [Google Scholar] [CrossRef]
- Hyde, J.; Carr, H.; Kelley, N.; Seneviratne, R.; Reed, C.; Parlatini, V.; Garner, M.; Solmi, M.; Rosson, S.; Cortese, S.; et al. Efficacy of neurostimulation across mental disorders: Systematic review and meta-analysis of 208 randomized controlled trials. Mol. Psychiatry 2022, 7, 2709–2719. [Google Scholar] [CrossRef]
- Nejati, V.; Salehinejad, M.A.; Nitsche, M.A.; Najian, A.; Javadi, A.H. Transcranial direct current stimulation improves executive dysfunctions in ADHD: Implications for inhibitory control, interference control, working memory, and cognitive flexibility. J. Atten. Disord. 2020, 24, 1928–1943. [Google Scholar] [CrossRef]
- Jones, K.T.; Johnson, E.L.; Berryhill, M.E. Frontoparietal theta-gamma interactions track working memory enhancement with training and Tdcs. Neuroimage 2020, 211, 116615. [Google Scholar] [CrossRef]
- Voegtle, A.; Reichert, C.; Hinrichs, H.; Sweeney-Reed, C.M. Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing. Brain Sci. 2022, 12, 1545. [Google Scholar] [CrossRef]
- Antonenko, D.; Fromm, A.E.; Thams, F.; Kuzmina, A.; Backhaus, M.; Knochenhauer, E.; Li, S.-C.; Grittner, U.; Flöel, A. Cognitive training and brain stimulation in patients with cognitive impairment: A randomized controlled trial. Alzheimer’s Res. Ther. 2024, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, R.; Nawaz, R.; Wood, G.; Yap, V.V. Exploring the Effects of EEG-Based Alpha Neurofeedback on Working Memory Capacity in Healthy Participants. Bioengineering 2023, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Beuckelaer, A.D.; Zhou, R. EEG correlates of neutral working memory training induce attentional control improvements in test anxiety. Biol. Psychol. 2022, 174, 108407. [Google Scholar] [CrossRef]
- Harmony, T. The functional significance of delta oscillations in cognitive processing. Front. Integr. Neurosci. 2013, 7, 83. [Google Scholar] [CrossRef]
- Kumar, S.; Gander, P.E.; Berger, J.I.; Billig, A.J.; Nourski, K.V.; Oya, H.; Kawasaki, H.; Howard, M.A.; Griffiths, T.D. Oscillatory correlates of auditory working memory examined with human electrocorticography. Neuropsychologia 2020, 150, 107691. [Google Scholar] [CrossRef]
- Cassidy, C.M.; Van Snellenberg, J.X.; Benavides, C.; Slifstein, M.; Wang, Z.; Moore, H.; Abi-Dargham, A.; Horga, G. Dynamic connectivity between brain networks supports working memory: Relationships to dopamine release and schizophrenia. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 4377–4388. [Google Scholar] [CrossRef]
- McKeon, S.D.; Calabro, F.; Thorpe, R.V.; de la Fuente, A.; Foran, W.; Parr, A.C.; Jones, S.R.; Luna, B. Age-related differences in transient gamma band activity during working memory maintenance through adolescence. Neuroimage 2023, 274, 120112. [Google Scholar] [CrossRef]
- Miller, E.K.; Lundqvist, M.; Bastos, A.M. Working memory 2.0. Neuron 2018, 100, 463–475. [Google Scholar] [CrossRef]
- Carver, F.W.; Rubinstein, D.Y.; Gerlich, A.H.; Fradkin, S.I.; Holroyd, T.; Coppola, R. Prefrontal high gamma during a magnetoencephalographic working memory task. Hum. Brain Mapp. 2018, 40, 1774–1785. [Google Scholar] [CrossRef]
- Hoy, K.E.; Bailey, N.W.; Arnold, S.L.; Fitzgerald, P.B. The effect of transcranial direct current stimulation on gamma activity and working memory in schizophrenia. Psychiatry Res. 2015, 228, 191–196. [Google Scholar] [CrossRef]
- Barbey, A.K.; Koenigs, M.; Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex 2013, 49, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, S.; Loo, C.K.; Bai, S.; Dokos, S.; Martin, D.M. Focalised stimulation using high definition transcranial direct current stimulation (HD-tDCS) to investigate declarative verbal learning and memory functioning. Neuroimage 2015, 117, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Vanderhasselt, M.A. Working memory improvement with non-invasive brain stimulation of the dorsolateral prefrontal cortex: A systematic review and meta-analysis. Brain Cogn. 2014, 86, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cancino, R.; Heng, J.; Delorme, A.; Kreutz-Delgado, K.; Sotero, R.C.; Makeig, S. Measuring transient phase-amplitude coupling using local mutual information. Neuroimage 2019, 185, 361–378. [Google Scholar] [CrossRef]
- Robert, M.G.R.; John, A.N. Working memory revived in older adults by synchronizing rhythmic brain circuits. Nat. Neurosci. 2019, 22, 820–827. [Google Scholar]
- Hülsemann, M.J.; Naumann, E.; Rasch, B. Quantification of Phase-Amplitude Coupling in Neuronal Oscillations: Comparison of Phase-Locking Value, Mean Vector Length, Modulation Index, and Generalized-Linear-Modeling-Cross-Frequency-Coupling. Front. Neurosci. 2019, 13, 573. [Google Scholar] [CrossRef]
- Kesic, S.; Kalauzi, A.; Radulovacki, M.; Carley, D.W.; Saponjic, J. Coupling changes in cortical and pontine sigma and theta frequency oscillations following monoaminergic lesions in rat. Sleep Breath 2011, 15, 35–47. [Google Scholar] [CrossRef]
- Tort, A.B.L.; Kramer, M.A.; Thorn, C.; Gibson, D.J.; Kubota, Y.; Graybiel, A.M.; Kopell, N.J. Dynamic cross-frequency couplings of local field potential oscillations in rat striatum and hippocampus during performance of a T-maze task. Proc. Natl. Acad. Sci. USA 2008, 105, 20517–20522. [Google Scholar] [CrossRef]
- Ye, H.; Li, G.; Sheng, X.; Zhu, X. Phase-amplitude coupling between low-frequency scalp EEG and high-frequency intracranial EEG during working memory task. J. Neural Eng. 2022, 19, 026043. [Google Scholar] [CrossRef]
- Cheng, C.H.; Hung, C.C.; Chao, Y.P.; Nouchi, R.; Wang, P.N. Subjective cognitive decline exhibits alterations of resting-state phase-amplitude coupling in precuneus. Clin. Neurophysiol. 2023, 156, 281–289. [Google Scholar] [CrossRef]
- Kumar, S.; Zomorrodi, R.; Ghazala, Z.; Goodman, M.S.; Blumberger, D.M.; Cheam, A.; Fischer, C.; Daskalakis, Z.J.; Mulsant, B.H.; Pollock, B.G.; et al. Extent of Dorsolateral Prefrontal Cortex Plasticity and Its Association with Working Memory in Patients with Alzheimer Disease. JAMA Psychiatry 2017, 74, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- D’Esposito, M.; Postle, B.R. The cognitive neuroscience of working memory. Annu. Rev. Psychol. 2014, 66, 115–142. [Google Scholar] [CrossRef] [PubMed]
- Souza-Couto, D.; Bretas, R.; Aversi-Ferreira, T.A. Neuropsychology of the parietal lobe: Luria’s and contemporary conceptions. Front. Neurosci. 2023, 17, 1226226. [Google Scholar] [CrossRef]
- Hill, A.T.; Rogasch, N.C.; Fitzgerald, P.B.; Hoy, K.E. Effects of single versus dual-site High-Definition transcranial direct current stimulation (HD-tDCS) on cortical reactivity and working memory performance in healthy subjects. Brain Stimul. 2018, 11, 1033–1043. [Google Scholar] [CrossRef]
- Hunt, M.J.; Kopell, N.J.; Traub, R.D. Aberrant Network Activity in Schizophrenia. Trends Neurosci. 2017, 40, 371–382. [Google Scholar] [CrossRef]
- Pellegrini, M.; Zoghi, M.; Jaberzadeh, S. The effects of transcranial direct current stimulation on corticospinal and cortico-cortical excitability and response variability: Conventional versus high-definition montages. Neurosci. Res. 2021, 166, 12–25. [Google Scholar] [CrossRef]
- Yang, B.; Xiaoyu, X.; Jiannan, K.; Yi, Y.; Jianghong, H. TDCS modulates cortical excitability in patients with disorders of consciousness. NeuroImage. Clin. 2017, 15, 702–709. [Google Scholar]
- Wong, P.L.; Yang, Y.R.; Huang, S.F.; Wang, R.Y. Effects of DLPFC tDCS Followed by Treadmill Training on Dual-Task Gait and Cortical Excitability in Parkinson’s Disease: A Randomized Controlled Trial. Neurorehabilit. Neural Repair 2024, 38, 680–692. [Google Scholar] [CrossRef]
- Ikeda, T.; Takahashi, T.; Hiraishi, H.; Saito, D.N.; Kikuchi, M. Anodal Transcranial Direct Current Stimulation Induces High Gamma-Band Activity in the Left Dorsolateral Prefrontal Cortex During a Working Memory Task: A Double-Blind, Randomized, Crossover Study. Front. Hum. Neurosci. 2019, 24, 136. [Google Scholar] [CrossRef]
- Keeser, D.; Padberg, F.; Reisinger, E.; Pogarell, O.; Kirsch, V.; Palm, U.; Karch, S.; Möller, H.-J.; Nitsche, M.A.; Mulert, C. Prefrontal direct current stimulation modulates resting EEG and event-related potentials in healthy subjects: A standardized low resolution tomography (sLORETA) study. Neuroimage 2011, 55, 644–657. [Google Scholar] [CrossRef]
- Eriksson, J.; Vogel, E.K.; Lansner, A.; Bergström, F.; Nyberg, L. Neurocognitive Architecture of Working Memory. Neuron 2015, 88, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Thams, F.; Külzow, N.; Flöel, A.; Antonenko, D. Modulation of network centrality and gray matter microstructure using multi-session brain stimulation and memory training. Hum. Brain Mapp. 2022, 43, 3416–3426. [Google Scholar] [CrossRef] [PubMed]
- Licata, A.E.; Zhao, Y.; Herrmann, O.; Hillis, A.E.; Desmond, J.; Onyike, C.; Tsapkini, K. Sex differences in effects of tDCS and language treatments on brain functional connectivity in primary progressive aphasia. NeuroImage. Clin. 2023, 37, 103329. [Google Scholar] [CrossRef]
- Kurtin, D.L.; Violante, I.R.; Zimmerman, K.; Leech, R.; Hampshire, A.; Patel, M.C.; Carmichael, D.W.; Sharp, D.J.; Li, L.M. Investigating the interaction between white matter and brain state on tDCS-induced changes in brain network activity. Brain Stimul. 2021, 14, 1261–1270. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, P.; Yu, H. The Instantaneous Effect and Its Mechanism of Transcranial Direct Current Stimulation on Working Memory Based on Delta and Gamma Band Electroencephalography Characteristics. Brain Sci. 2025, 15, 579. https://doi.org/10.3390/brainsci15060579
Lu P, Yu H. The Instantaneous Effect and Its Mechanism of Transcranial Direct Current Stimulation on Working Memory Based on Delta and Gamma Band Electroencephalography Characteristics. Brain Sciences. 2025; 15(6):579. https://doi.org/10.3390/brainsci15060579
Chicago/Turabian StyleLu, Pengyi, and Hongli Yu. 2025. "The Instantaneous Effect and Its Mechanism of Transcranial Direct Current Stimulation on Working Memory Based on Delta and Gamma Band Electroencephalography Characteristics" Brain Sciences 15, no. 6: 579. https://doi.org/10.3390/brainsci15060579
APA StyleLu, P., & Yu, H. (2025). The Instantaneous Effect and Its Mechanism of Transcranial Direct Current Stimulation on Working Memory Based on Delta and Gamma Band Electroencephalography Characteristics. Brain Sciences, 15(6), 579. https://doi.org/10.3390/brainsci15060579