



The Impact of Some Modulators of the Renin–Angiotensin System on the Scopolamine-Induced Memory Loss Mice Model
 ,
,  ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
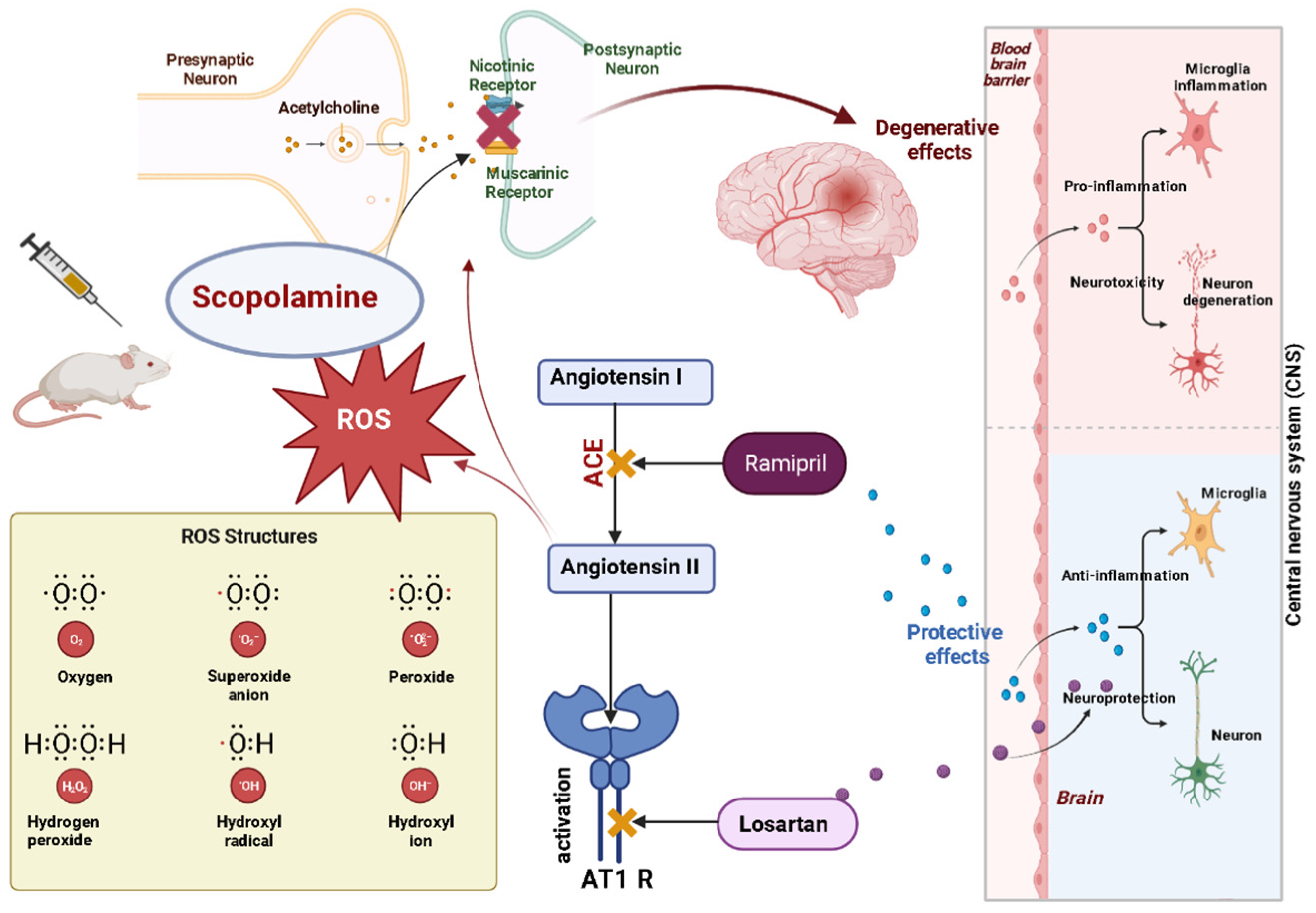
1. Introduction
2. Materials and Methods
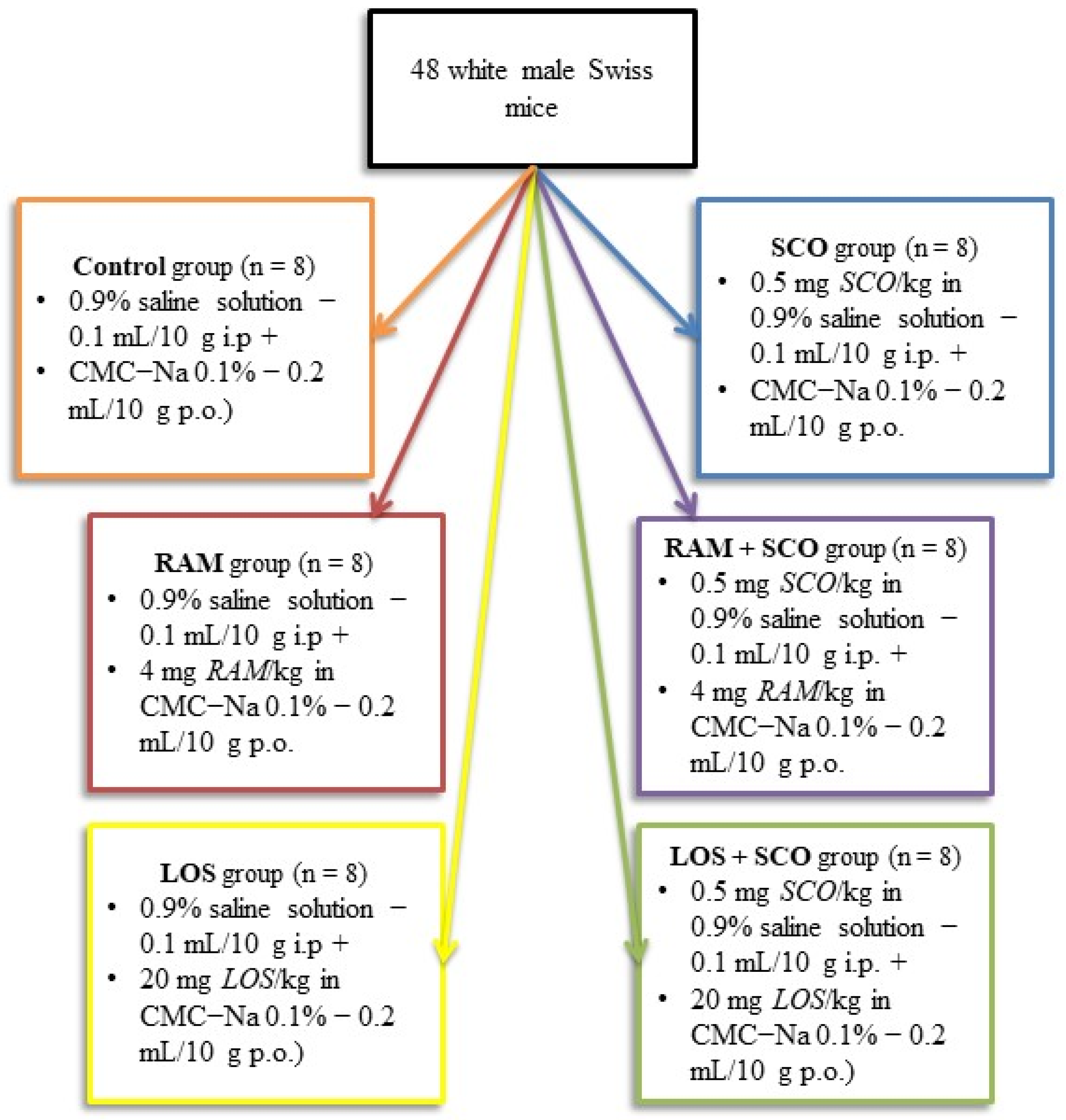
2.1. Animals and Housing
2.2. Substances and Treatments
2.3. Samples Collection and Preparation
2.4. Biochemical Analysis
2.5. Statistical Analysis
3. Results
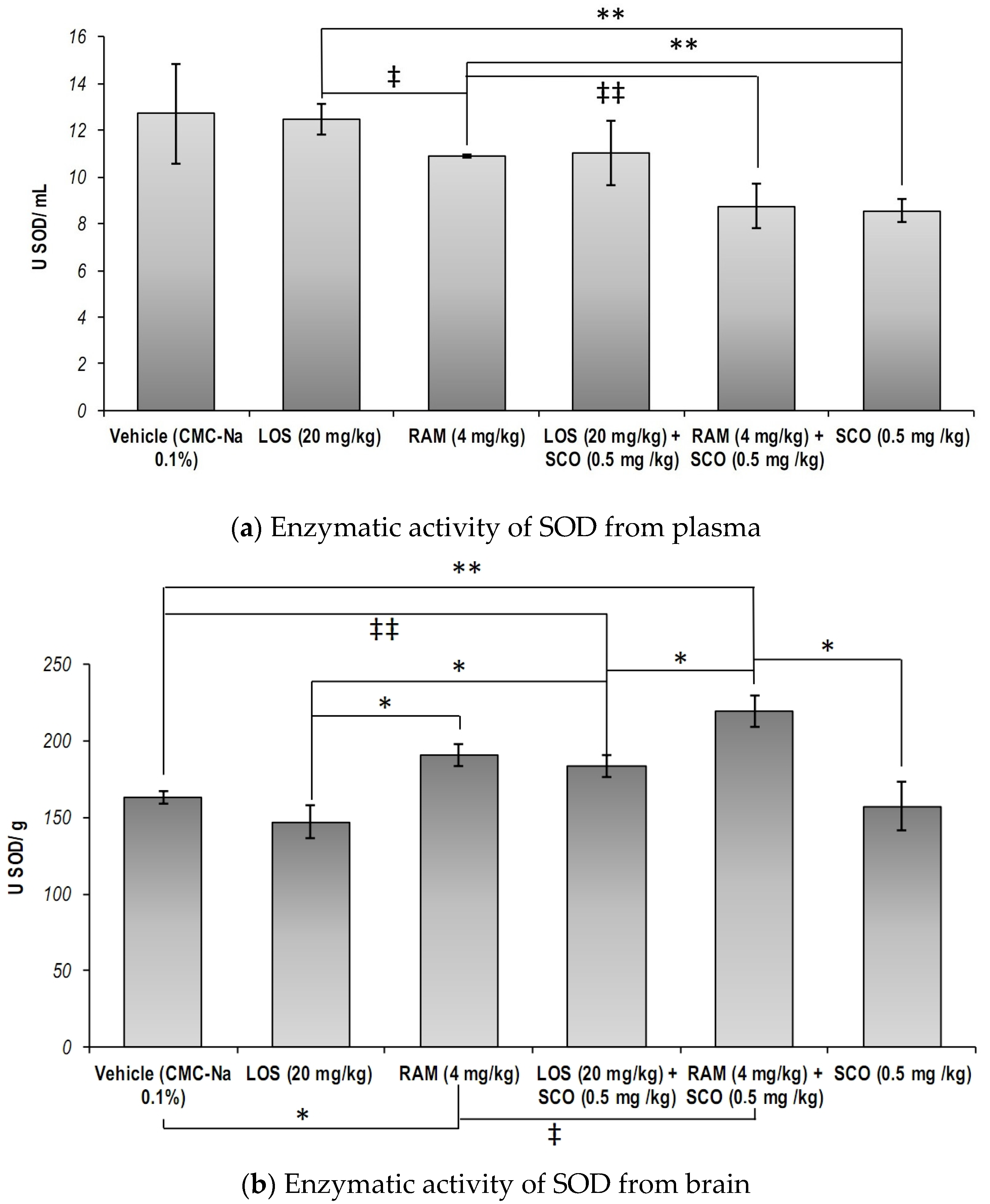
3.1. The Effect of RAS Modulation in Scopolamine Induced Memory Loss on the Systemic and Central Nervous System Total Superoxide Dismutase Specific Activity
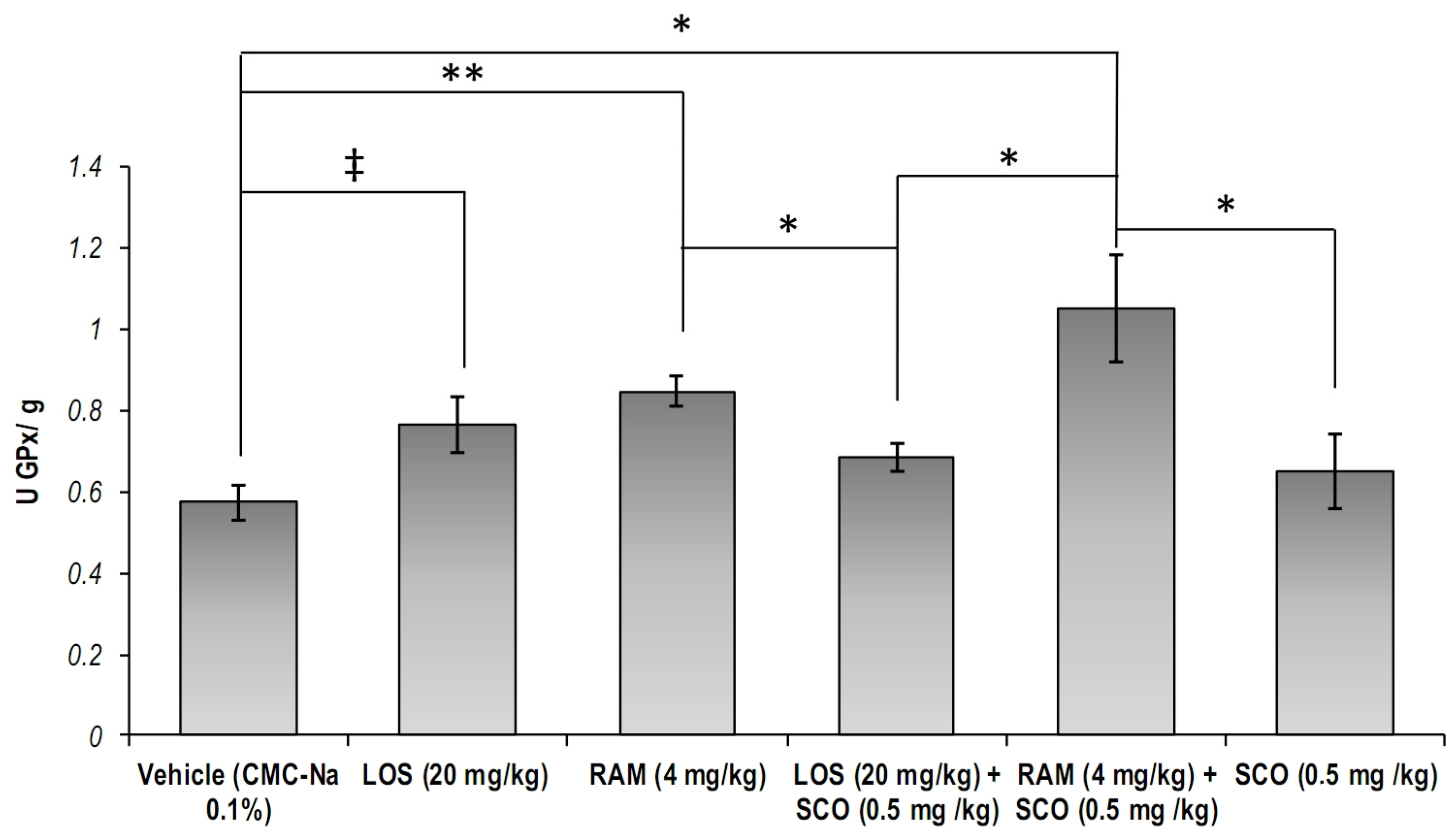
3.2. The Effect of RAS Modulation in Scopolamine Induced Memory Loss on the Systemic and Central Nervous System Cellular Glutathione Peroxidase Specific Activity
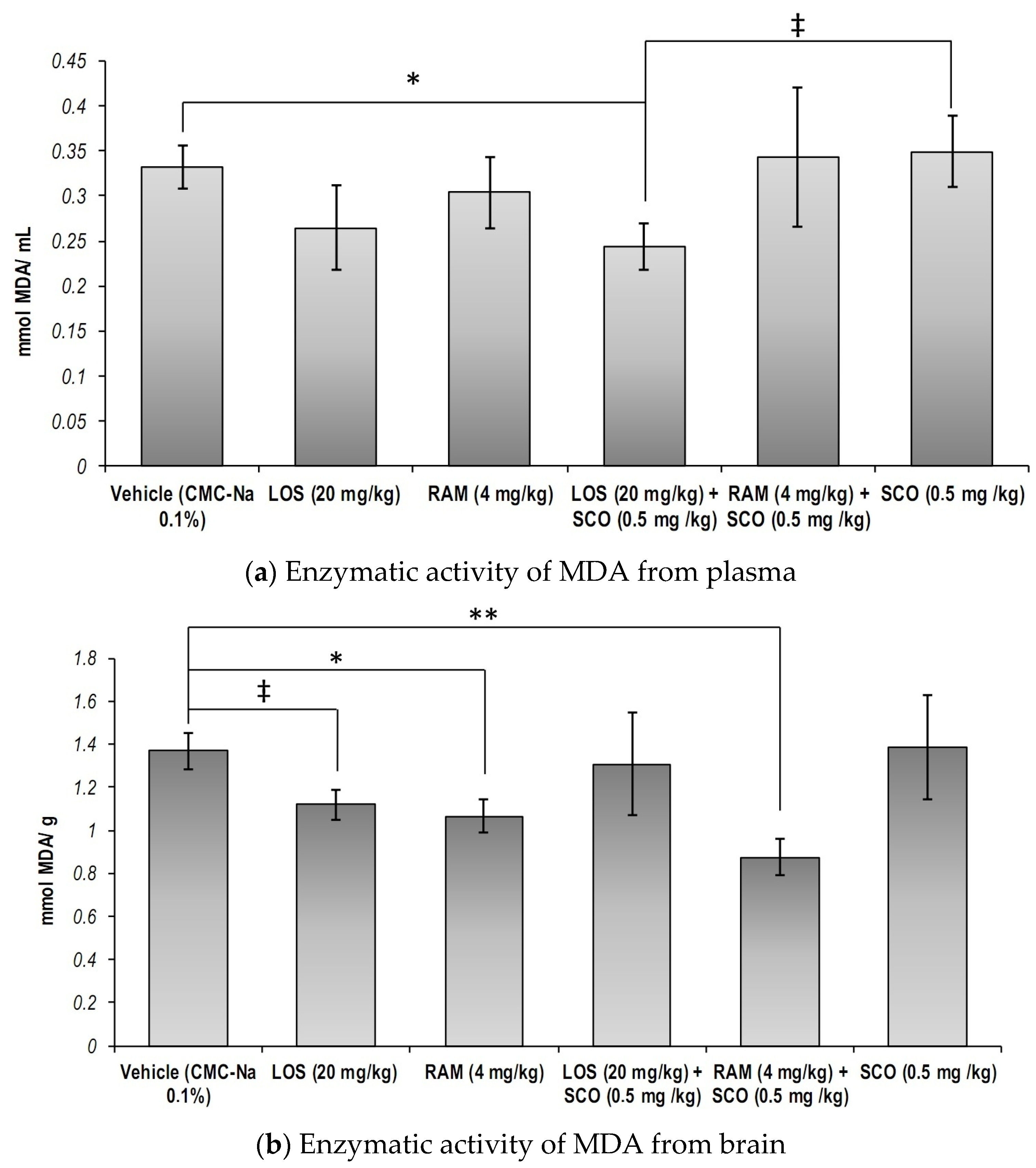
3.3. The Effect of RAS Modulation in Scopolamine-Induced Memory Loss on the Systemic and Central Nervous System Malondialdehyde Concentration
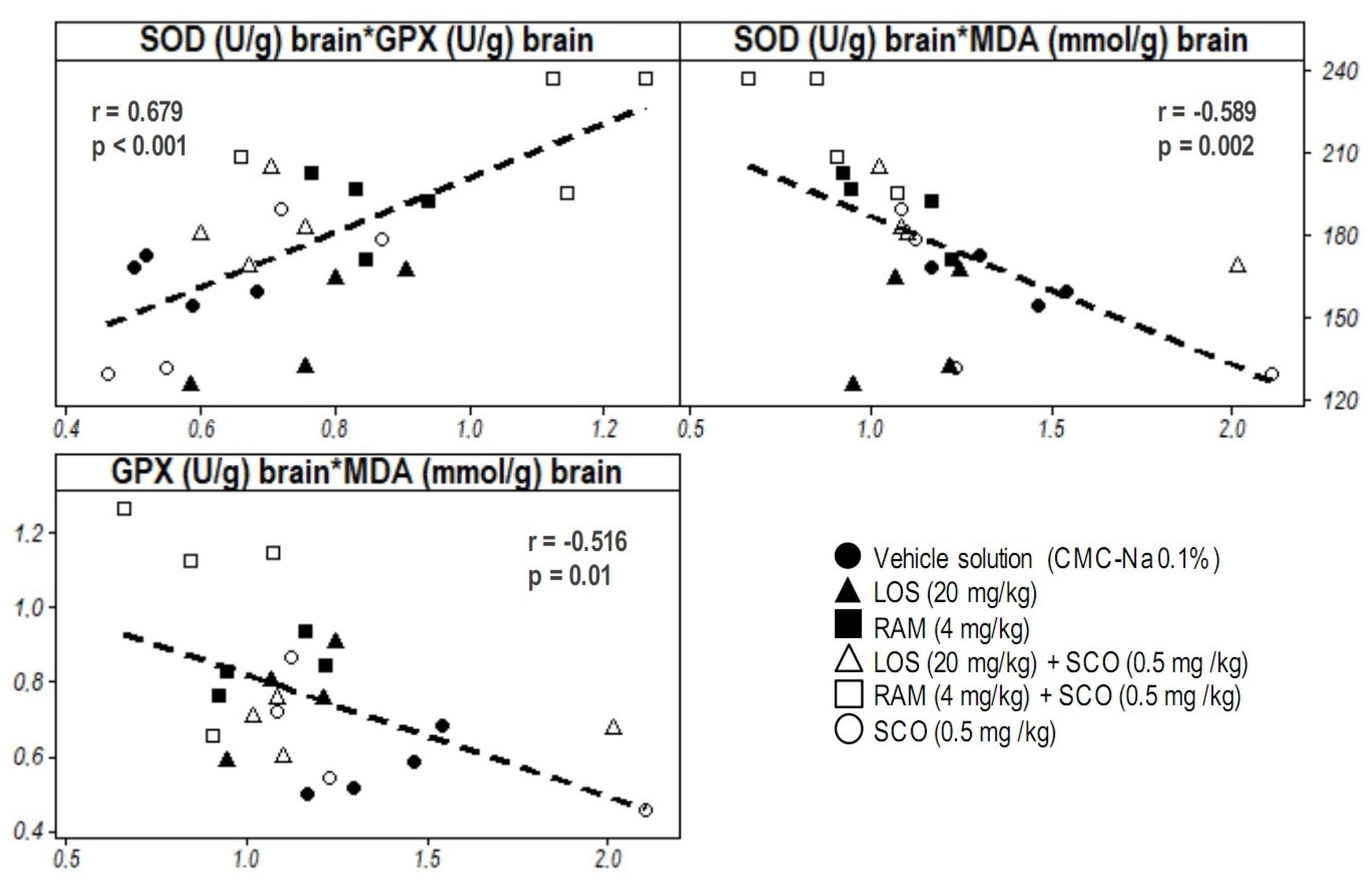
3.4. Correlative Analysis of Molecular Biomarkers and Discussion on the Relevance of Systemic and Central Nervous System Oxidative Stress in RAS Modulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flood, J.F.; Cherkin, A. Scopolamine Effects on Memory Retention in Mice: A Model of Dementia? Behav. Neural Biol. 1986, 45, 169–184. [Google Scholar] [CrossRef]
- Ababei, D.C.; Lefter, R.; Ciobica, A.; Antioh, I.; Barbu, R.M.; Arcan, O.D.; Chiriac, S.B.; Zbârcea, C.; Bild, W.; Bild, V. Effects of Renin-Angiotensin System Blockade on Anxiety and Memory in Mice. Farmacia 2017, 65, 381–389. [Google Scholar]
- Ciobica, A.; Olteanu, Z.; Padurariu, M.; Hritcu, L. The Effects of Pergolide on Memory and Oxidative Stress in a Rat Model of Parkinson’s Disease. J. Physiol. Biochem. 2012, 68, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Bild, W.; Ciobica, A. Angiotensin-(1-7) Central Administration Induces Anxiolytic-like Effects in Elevated plus Maze and Decreased Oxidative Stress in the Amygdala. J. Affect. Disord. 2013, 145, 165–171. [Google Scholar] [CrossRef]
- Bild, W.; Hritcu, L.; Stefanescu, C.; Ciobica, A. Inhibition of Central Angiotensin II Enhances Memory Function and Reduces Oxidative Stress Status in Rat Hippocampus. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 43, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ciobica, A.; Bild, V.; Hritcu, L.; Padurariu, M.; Bild, W. Effects of Angiotensin II Receptor Antagonists on Anxiety and Some Oxidative Stress Markers in Rat. Cent. Eur. J. Med. 2011, 6, 331–340. [Google Scholar] [CrossRef]
- Braszko, J.J. AT2 but Not AT1 Receptor Antagonism Abolishes Angiotensin II Increase of the Acquisition of Conditioned Avoidance Responses in Rats. Behav. Brain Res. 2002, 131, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Inaba, S.; Iwai, M.; Furuno, M.; Tomono, Y.; Kanno, H.; Senba, I.; Okayama, H.; Mogi, M.; Higaki, J.; Horiuchi, M. Continuous Activation of Renin-Angiotensin System Impairs Cognitive Function in Renin/Angiotensinogen Transgenic Mice. Hypertension 2009, 53, 356–362. [Google Scholar] [CrossRef]
- Maul, B.; Von Bohlen Und Halbach, O.; Becker, A.; Sterner-Kock, A.; Voigt, J.P.; Siems, W.E.; Grecksch, G.; Walther, T. Impaired Spatial Memory and Altered Dendritic Spine Morphology in Angiotensin II Type 2 Receptor-Deficient Mice. J. Mol. Med. 2008, 86, 563–571. [Google Scholar] [CrossRef]
- Walther, T.; Voigt, J.P.; Fukamizu, A.; Fink, H.; Bader, M. Learning and Anxiety in Angiotensin-Deficient Mice. Behav. Brain Res. 1999, 100, 1–4. [Google Scholar] [CrossRef]
- Padurariu, M.; Ciobica, A.; Hritcu, L.; Stoica, B.; Bild, W.; Stefanescu, C. Changes of Some Oxidative Stress Markers in the Serum of Patients with Mild Cognitive Impairment and Alzheimer’s Disease. Neurosci. Lett. 2010, 469, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Real, A.; Rey, P.; Soto-Otero, R.; Mendez-Alvarez, E.; Labandeira-Garcia, J.L. Angiotensin-Converting Enzyme Inhibition Reduces Oxidative Stress and Protects Dopaminergic Neurons in a 6-Hydroxydopamine Rat Model of Parkinsonism. J. Neurosci. Res. 2005, 81, 865–873. [Google Scholar] [CrossRef]
- Firouzabadi, N.; Ghazanfari, N.; Shoushtari, A.A.; Erfani, N.; Fathi, F.; Bazrafkan, M.; Bahramali, E. Genetic Variants of Angiotensin-Converting Enzyme Are Linked to Autism: A Case-Control Study. PLoS ONE 2016, 11, e0153667. [Google Scholar] [CrossRef]
- Wahlbeck, K.; Rimon, R. CSF Ace Correlates with Duration of Illness in Schizophrenia. Schizophr. Res. 1998, 29, 103. [Google Scholar] [CrossRef]
- Wahlbeck, K.; Ahokas, A.; Nikkilä, H.; Miettinen, K.; Rimón, R. A Longitudinal Study of Cerebrospinal Fluid Angiotensin-Converting Enzyme in Neuroleptic-Treated Schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1997, 21, 591–599. [Google Scholar] [CrossRef]
- Rygiel, K. Can Angiotensin-Converting Enzyme Inhibitors Impact Cognitive Decline in Early Stages of Alzheimer’s Disease? An Overview of Research Evidence in the Elderly Patient Population. J. Postgrad. Med. 2016, 62, 242–248. [Google Scholar] [CrossRef]
- Goel, R.; Bhat, S.A.; Hanif, K.; Nath, C.; Shukla, R. Angiotensin II Receptor Blockers Attenuate Lipopolysaccharide-Induced Memory Impairment by Modulation of NF-ΚB-Mediated BDNF/CREB Expression and Apoptosis in Spontaneously Hypertensive Rats. Mol. Neurobiol. 2018, 55, 1725–1739. [Google Scholar] [CrossRef]
- Arumugam, S.; Sreedhar, R.; Thandavarayan, R.A.; Karuppagounder, V.; Krishnamurthy, P.; Suzuki, K.; Nakamura, M.; Watanabe, K. Angiotensin Receptor Blockers: Focus on Cardiac and Renal Injury. Trends Cardiovasc. Med. 2016, 26, 221–228. [Google Scholar] [CrossRef]
- Bouchenaki, H.; Danigo, A.; Bernard, A.; Bessaguet, F.; Richard, L.; Sturtz, F.; Balayssac, D.; Magy, L.; Demiot, C. Ramipril Alleviates Oxaliplatin-Induced Acute Pain Syndrome in Mice. Front. Pharmacol. 2021, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ababei, D.C.; Bild, V.; Ciobică, A.; Lefter, R.M.; Rusu, R.N.; Bild, W. A Comparative Study on the Memory-Enhancing Actions of Oral Renin-Angiotensin System Altering Drugs in Scopolamine-Treated Mice. Am. J. Alzheimer’s Dis. Other Dement. 2019, 34, 329–336. [Google Scholar] [CrossRef]
- Ababei, D.C.; Beșchea Chiriac, S.; Bild, W.; Solcan, C.; Luca, A.; Rusu, R.N.; Bulea, D.; Bild, V. Synergistic Effects of the Doxepin-Candesartan Combination on the Thermoalgesic Sensibility in Mice. Farmacia 2017, 65, 726–730. [Google Scholar]
- Cojocariu, R.; Ciobica, A.; Balmus, I.M.; Guenne, S.; Trifan, A.; Stanciu, C.; Hricu, L.; Lefter, R. Antioxidant Capacity and Behavioral Relevance of a Polyphenolic Extract of Chrysanthellum Americanum in a Rat Model of Irritable Bowel Syndrome. Oxidative Med. Cell. Longev. 2019, 2019, 3492767. [Google Scholar] [CrossRef]
- Cojocariu, R.O.; Balmus, I.M.; Lefter, R.; Hritcu, L.; Ababei, D.C.; Ciobica, A.; Copaci, S.; Mot, S.E.L.; Copolovici, L.; Copolovici, D.M.; et al. Camelina Sativa Methanolic and Ethanolic Extract Potential in Alleviating Oxidative Stress, Memory Deficits, and Affective Impairments in Stress Exposure-Based Irritable Bowel Syndrome Mouse Models. Oxidative Med. Cell. Longev. 2020, 2020, 9510305. [Google Scholar] [CrossRef] [PubMed]
- Ciobica, A.S.; Lefter, R.M.; Balmus, M.I. P2-003: The Interaction between Decreased Short-term Spatial Memory and Increased Oxidative Stress in a Scopolamine-induced Rat Model of Alzheimer’s Disease. Alzheimer’s Dement. 2015, 11, P480–P481. [Google Scholar] [CrossRef]
- Marks, M.J.; O’Connor, M.F.; Artman, L.D.; Burch, J.B.; Collins, A.C. Chronic Scopolamine Treatment and Brain Cholinergic Function. Pharmacol. Biochem. Behav. 1984, 20, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Kohl, R.L.; Sandoz, G.R.; Reschke, M.F.; Calkins, D.S.; Richelson, E. Facilitation of Adaptation and Acute Tolerance to Stressful Sensory Input by Doxepin and Scopolamine Plus Amphetamine. J. Clin. Pharmacol. 1993, 33, 1092–1103. [Google Scholar] [CrossRef]
- Geng, C.; Mao, C.; Wu, L.; Cheng, Y.; Liu, R.; Chen, B.; Chen, L.; Zhang, L.; Xu, Z. Cholinergic Signal Activated Renin Angiotensin System Associated with Cardiovascular Changes in the Ovine Fetus. J. Perinat. Med. 2010, 38, 71–76. [Google Scholar] [CrossRef][Green Version]
- Cosarderelioglu, C.; Nidadavolu, L.S.; George, C.J.; Oh, E.S.; Bennett, D.A.; Walston, J.D.; Abadir, P.M. Brain Renin–Angiotensin System at the Intersect of Physical and Cognitive Frailty. Front. Neurosci. 2020, 14, 586314. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Sakamoto, T.; Komatsu, K.; Fujishima, H.; Morii, T.; Narita, T.; Takahashi, T.; Yamada, Y. Reduction of Circulating Superoxide Dismutase Activity in Type 2 Diabetic Patients with Microalbuminuria and Its Modulation by Telmisartan Therapy. Hypertens. Res. 2011, 34, 1302–1308. [Google Scholar] [CrossRef]
- Kiran, V.U.; Rajaiah, N.V.; Krishna, D.R.; Reddy, Y.N. Effect of Losartan and Ramipril on Oxidative stress and Anti-oxidant Status in South Indian Hypertensive Patients. Int. J. Pharmacol. 2010, 6, 916–920. [Google Scholar] [CrossRef]
- Mitu, F.; Cojocaru, E.; Gorga, A.; Leon, M.-M. Blocantii Receptorilor Pentru Angiotensina I Și II Și Rezultatele Cardiovasculare. Intern. Med. 2014, 11, 41–53. [Google Scholar]
- Susanne, H.; Culman, J.; Boser, M.; Qadri, F.; Unger, T. Effect of angiotensin AT2 and muscarinic receptor blockade on osmotically induced vasopressin release. Eur. J. Pharmacol. 1996, 300, 119–123. [Google Scholar]
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II Revisited: New Roles in Inflammation, Immunology and Aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef]
- Kucuk, M.; Kaya, M.; Kalayci, R.; Cimen, V.; Kudat, H.; Arican, N.; Elmas, I.; Korkut, F. Effects of Losartan on the Blood-Brain Barrier Permeability in Long-Term Nitric Oxide Blockade-Induced Hypertensive Rats. Life Sci. 2002, 71, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Abiodun, O.A.; Ola, M.S. Role of Brain Renin Angiotensin System in Neurodegeneration: An Update. Saudi J. Biol. Sci. 2020, 27, 905–912. [Google Scholar] [CrossRef]
- Campos, J.; Pacheco, R. Involvement of Dopaminergic Signaling in the Cross Talk between the Renin-Angiotensin System and Inflammation. Semin. Immunopathol. 2020, 42, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.A.; Ishrat, T.; Pillai, B.; Fouda, A.Y.; Sayed, M.A.; Eldahshan, W.; Waller, J.L.; Ergul, A.; Fagan, S.C. RAS Modulation Prevents Progressive Cognitive Impairment after Experimental Stroke: A Randomized, Blinded Preclinical Trial. J. Neuroinflamm. 2018, 15, 229. [Google Scholar] [CrossRef]
- Mirzahosseini, G.; Ismael, S.; Ahmed, H.A.; Ishrat, T. Manifestation of Renin Angiotensin System Modulation in Traumatic Brain Injury. Metab. Brain Dis. 2021, 36, 1079–1086. [Google Scholar] [CrossRef]
- He, D.H.; Zhang, L.M.; Lin, L.M.; Ning, R.B.; Wang, H.J.; Xu, C.S.; Lin, J.X. Long-Term Prehypertension Treatment with Losartan Effectively Prevents Brain Damage and Stroke in Stroke-Prone Spontaneously Hypertensive Rats. Int. J. Mol. Med. 2014, 33, 301–309. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Saavedra, J.M.; Sánchez-Lemus, E.; Benicky, J. Blockade of Brain Angiotensin II AT1 Receptors Ameliorates Stress, Anxiety, Brain Inflammation and Ischemia: Therapeutic Implications. Psychoneuroendocrinology 2011, 36, 1–18. [Google Scholar] [CrossRef]
- Saavedra, J.M. Beneficial Effects of Angiotensin II Receptor Blockers in Brain Disorders. Pharmacol. Res. 2017, 125, 91–103. [Google Scholar] [CrossRef]
- Osherovich, L. New Muscle for ARB Strategy. Sci.-Bus. Exch. 2011, 4, 587. [Google Scholar] [CrossRef]
- Spurney, C.F.; Sali, A.; Guerron, A.D.; Iantorno, M.; Yu, Q.; Gordish-Dressman, H.; Rayavarapu, S.; Van Der Meulen, J.; Hoffman, E.P.; Nagaraju, K. Losartan Decreases Cardiac Muscle Fibrosis and Improves Cardiac Function in Dystrophin-Deficient Mdx Mice. J. Cardiovasc. Pharmacol. Ther. 2011, 16, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Mao, C.; Hu, Y.; Rui, C.; Xu, Z.; Zhang, L. Cardiovascular Effects of Losartan and Its Relevant Clinical Application. Curr. Med. Chem. 2009, 16, 3841–3857. [Google Scholar] [CrossRef] [PubMed]
- Tamuli, S.; Kakati, S.; Das, S.; Singh, K.D.; Ghosh, S.K. Comparative Studies of Efficacy and Effects on Oxidative Stress of Amlodipine and Ramipril in the Hypertensive Patients of North East India. Int. J. Pharm. Pharm. Sci. 2015, 7, 118–121. [Google Scholar]
- Singh, B.; Mourya, A.; Sah, S.P.; Kumar, A. Protective Effect of Losartan and Ramipril against Stress Induced Insulin Resistance and Related Complications: Anti-Inflammatory Mechanisms. Eur. J. Pharmacol. 2017, 801, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Dinicolantonio, J.J.; Lavie, C.J.; O’Keefe, J.H. Not All Angiotensin-Converting Enzyme Inhibitors Are Equal: Focus on Ramipril and Perindopril. Postgrad. Med. 2013, 125, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Jawaid, T.; Jahan, S.; Kamal, M. A Comparative Study of Neuroprotective Effect of Angiotensin Converting Enzyme Inhibitors against Scopolamine-Induced Memory Impairments in Rats. J. Adv. Pharm. Technol. Res. 2015, 6, 130–135. [Google Scholar] [CrossRef]
- Nade, V.S.; Kawale, L.A.; Valte, K.D.; Shendye, N.V. Cognitive Enhancing Effect of Angiotensin-Converting Enzyme Inhibitors and Angiotensin Receptor Blockers on Learning and Memory. Indian J. Pharmacol. 2015, 47, 263–269. [Google Scholar] [CrossRef]
- Carnovale, C.; Perrotta, C.; Baldelli, S.; Cattaneo, D.; Montrasio, C.; Barbieri, S.S.; Pompilio, G.; Vantaggiato, C.; Clementi, E.; Pozzi, M. Antihypertensive Drugs and Brain Function: Mechanisms Underlying Therapeutically Beneficial and Harmful Neuropsychiatric Effects. Cardiovasc. Res. 2022, 119, 647–667. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ababei, D.-C.; Balmus, I.-M.; Bild, W.; Ciobica, A.S.; Lefter, R.M.; Rusu, R.-N.; Stanciu, G.D.; Cojocaru, S.; Hancianu, M.; Bild, V. The Impact of Some Modulators of the Renin–Angiotensin System on the Scopolamine-Induced Memory Loss Mice Model. Brain Sci. 2023, 13, 1211. https://doi.org/10.3390/brainsci13081211
Ababei D-C, Balmus I-M, Bild W, Ciobica AS, Lefter RM, Rusu R-N, Stanciu GD, Cojocaru S, Hancianu M, Bild V. The Impact of Some Modulators of the Renin–Angiotensin System on the Scopolamine-Induced Memory Loss Mice Model. Brain Sciences. 2023; 13(8):1211. https://doi.org/10.3390/brainsci13081211
Chicago/Turabian StyleAbabei, Daniela-Carmen, Ioana-Miruna Balmus, Walther Bild, Alin Stelian Ciobica, Radu Marian Lefter, Răzvan-Nicolae Rusu, Gabriela Dumitrita Stanciu, Sabina Cojocaru, Monica Hancianu, and Veronica Bild. 2023. "The Impact of Some Modulators of the Renin–Angiotensin System on the Scopolamine-Induced Memory Loss Mice Model" Brain Sciences 13, no. 8: 1211. https://doi.org/10.3390/brainsci13081211
APA StyleAbabei, D.-C., Balmus, I.-M., Bild, W., Ciobica, A. S., Lefter, R. M., Rusu, R.-N., Stanciu, G. D., Cojocaru, S., Hancianu, M., & Bild, V. (2023). The Impact of Some Modulators of the Renin–Angiotensin System on the Scopolamine-Induced Memory Loss Mice Model. Brain Sciences, 13(8), 1211. https://doi.org/10.3390/brainsci13081211