
Neural Entrainment to Musical Pulse in Naturalistic Music Is Preserved in Aging: Implications for Music-Based Interventions

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.3. Behavioral Battery
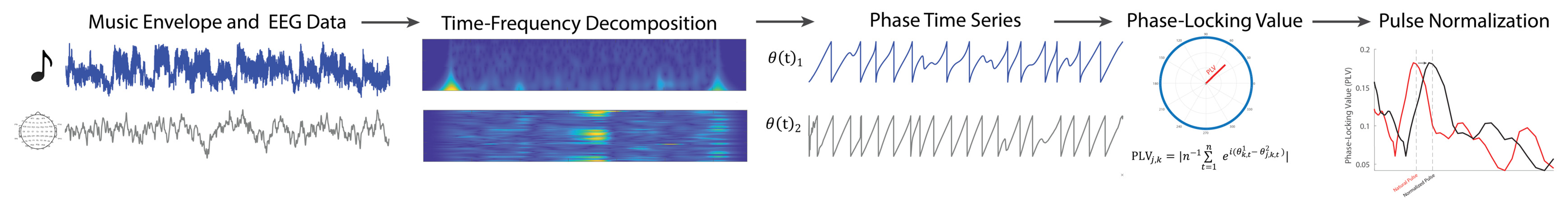
2.4. Audiovisual Stimulation
2.5. EEG Recording
2.6. EEG Preprocessing
2.7. Music-Feature Analysis: Estimating Pulse Frequencies and Identifying Stable Epochs
2.8. Neural Entrainment to Rhythm: Phase-Locking Values
2.9. Pulse Normalization of Phase-Locking Values
2.10. Linear Mixed-Effects Models
3. Results
3.1. Behavioral Battery Results
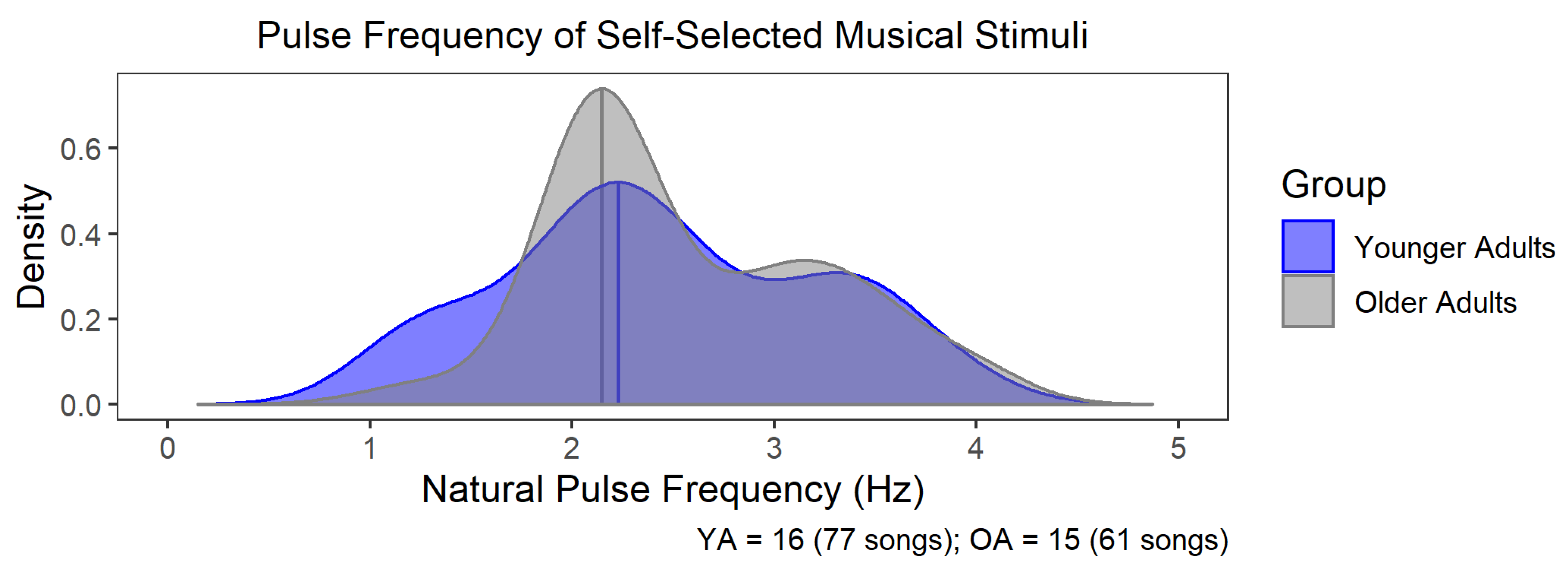
3.2. Natural Pulse Frequency of Self-Selected Music Did Not Differ between OA and YA
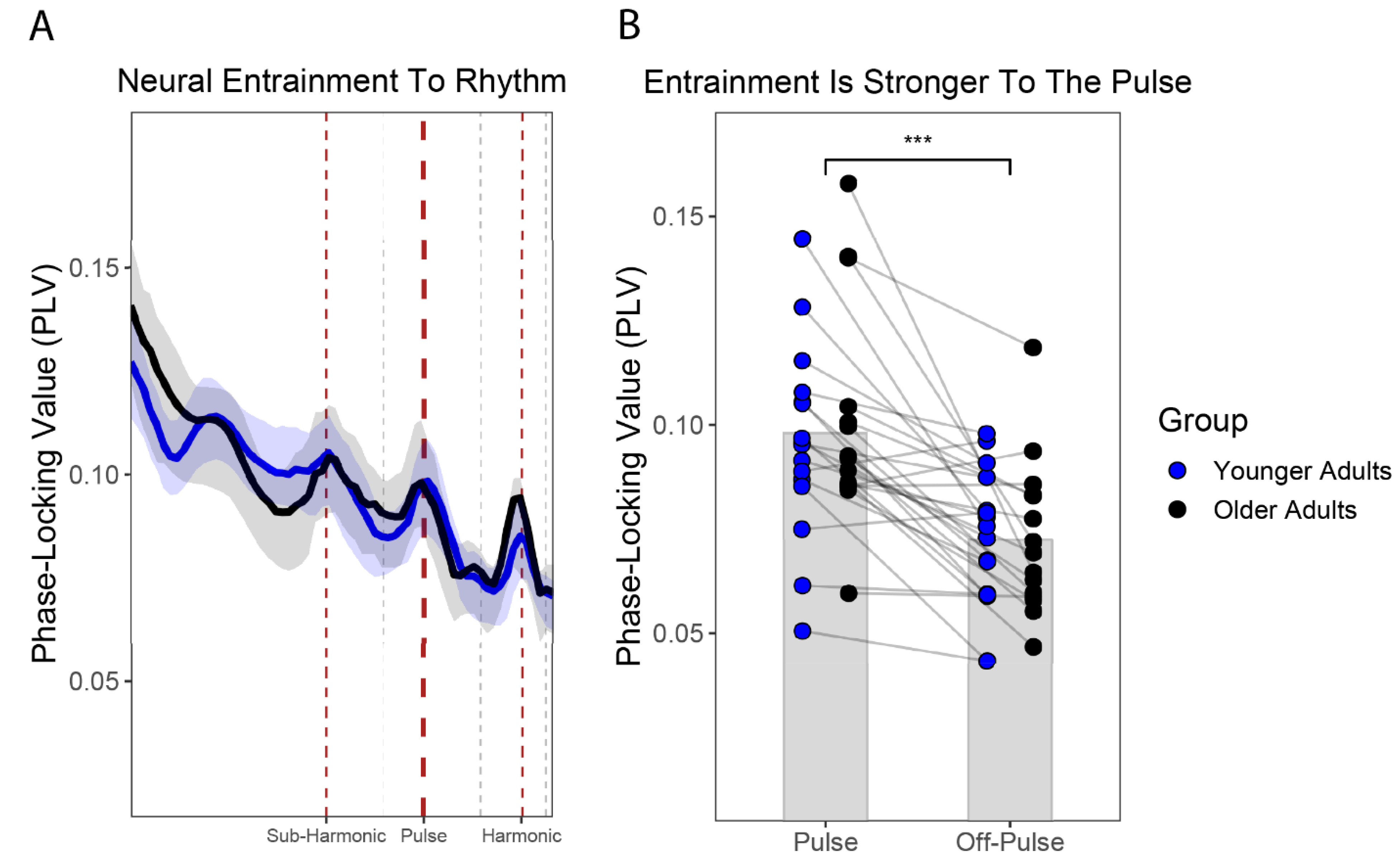
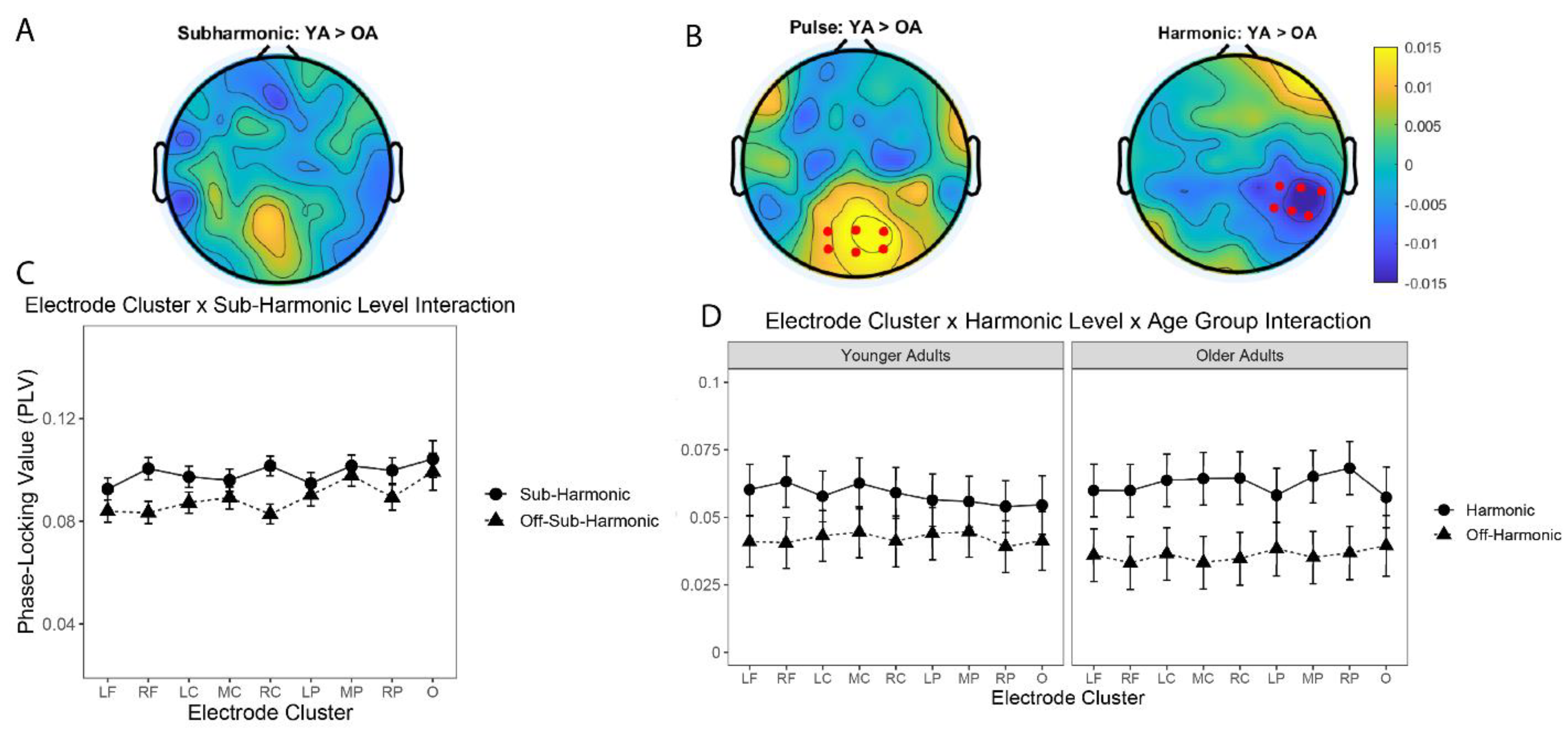
3.3. Neural Entrainment at the Pulse Level Did Not Differ between OA and YA
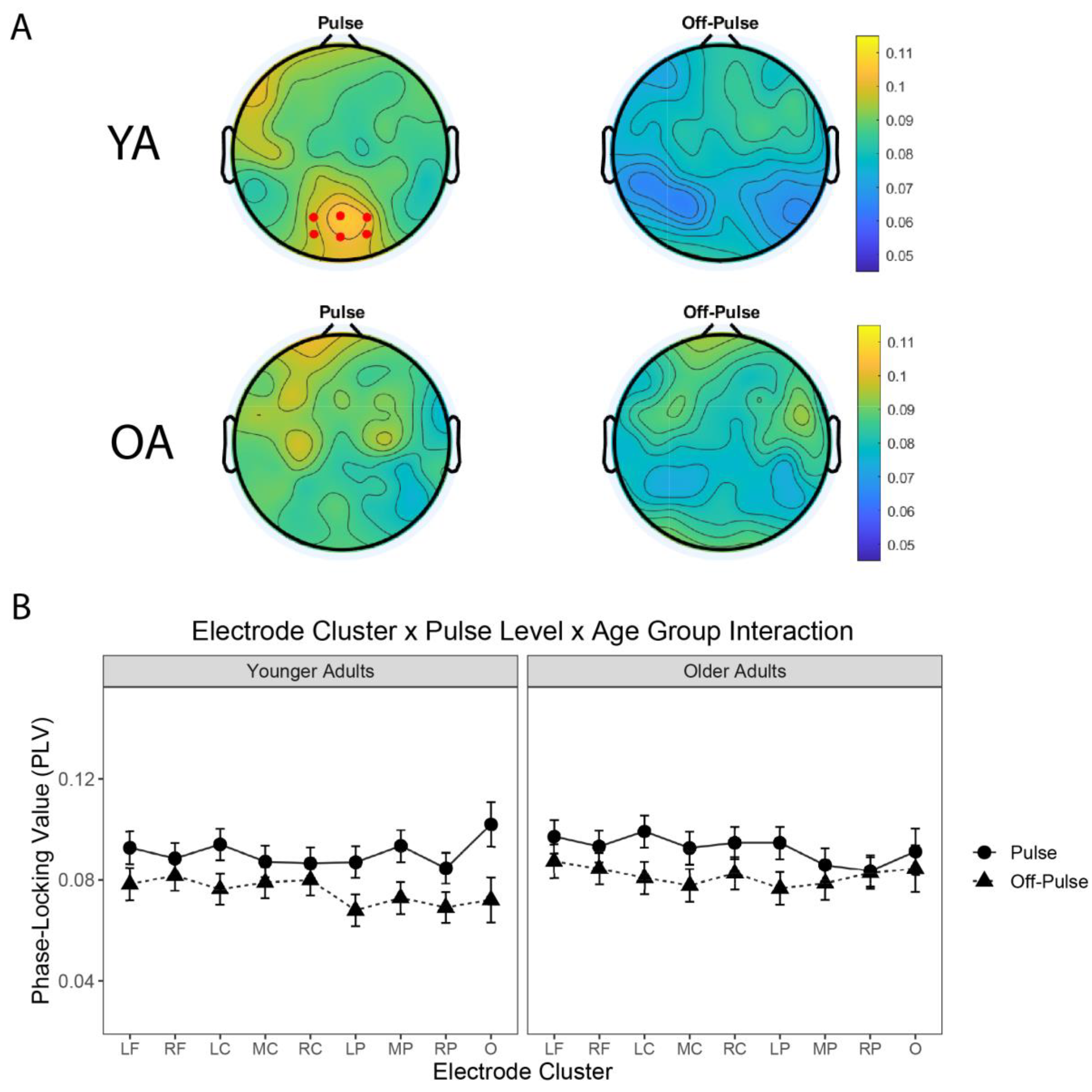
3.4. Neural Entrainment to the Pulse Differed across Electrode Clusters and Age Groups
3.5. Neural Entrainment at Sub-Harmonic and Harmonic Levels Did Not Differ between OA and YA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheever, T.; Taylor, A.; Finkelstein, R.; Edwards, E.; Thomas, L.; Bradt, J.; Holochwost, S.J.; Johnson, J.K.; Limb, C.; Patel, A.D.; et al. NIH/Kennedy Center Workshop on Music and the Brain: Finding Harmony. Neuron 2018, 97, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Global Council on Brain Health. Music on Our Minds: The Rich Potential of Music to Promote Brain Health and Mental Well-Being; Global Council on Brain Health: Washington, DC, USA, 2020. [Google Scholar]
- Mammarella, N.; Fairfield, B.; Cornoldi, C. Does music enhance cognitive performance in healthy older adults? The Vivaldi effect. Aging Clin. Exp. Res. 2007, 19, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.; Dowson, B.; McDermott, O.; Schneider, J.; Fernandes, L. Music-based interventions in the acute setting for patients with dementia: A systematic review. Eur. Geriatr. Med. 2020, 11, 929–943. [Google Scholar] [CrossRef]
- van der Steen, J.T.; Smaling, H.J.; van der Wouden, J.C.; Bruinsma, M.S.; Scholten, R.J.; Vink, A.C. Music-based therapeutic interventions for people with dementia. Cochrane Database Syst. Rev. 2018, 7, CD003477. [Google Scholar] [CrossRef]
- Vasionytė, I.; Madison, G. Musical intervention for patients with dementia: A meta-analysis. J. Clin. Nurs. 2013, 22, 1203–1216. [Google Scholar] [CrossRef]
- Vink, A.; Hanser, S. Music-Based Therapeutic Interventions for People with Dementia: A Mini-Review. Medicines 2018, 5, 109. [Google Scholar] [CrossRef]
- Loui, P. Neuroscientific Insights for Improved Outcomes in Music-based Interventions. Music Sci. 2020, 3, 205920432096506. [Google Scholar] [CrossRef]
- Koelsch, S. Brain correlates of music-evoked emotions. Nat. Rev. Neurosci. 2014, 15, 170–180. [Google Scholar] [CrossRef]
- Loui, P.; Przysinda, E. Music and the Brain: Areas and Networks. In Routledge Companion Music Cognition; Routledge: London, UK, 2017; pp. 13–24. [Google Scholar]
- Quinci, M.A.; Belden, A.; Goutama, V.; Gong, D.; Hanser, S.; Donovan, N.J.; Geddes, M.; Loui, P. Music-Based Intervention Connects Auditory and Reward Systems. bioRxiv 2021. bioRxiv:2021.07.02.450867. [Google Scholar]
- Wang, D.; Belden, A.; Hanser, S.B.; Geddes, M.R.; Loui, P. Resting-State Connectivity of Auditory and Reward Systems in Alzheimer’s Disease and Mild Cognitive Impairment. Front. Hum. Neurosci. 2020, 14, 280. [Google Scholar] [CrossRef]
- Sutcliffe, R.; Du, K.; Ruffman, T. Music Making and Neuropsychological Aging: A Review. Neurosci. Biobehav. Rev. 2020, 113, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, L.; Moussard, A.; Bigand, E.; Tillmann, B. Music and the Aging Brain. In The Oxford Handbook of Music and the Brain; Thaut, M.H., Hodges, D.A., Eds.; Oxford University Press: Oxford, UK, 2019; pp. 622–644. [Google Scholar]
- Tichko, P.; Kim, J.C.; Large, E.; Loui, P. Integrating music-based interventions with Gamma-frequency stimulation: Implications for healthy ageing. Eur. J. Neurosci. 2020, 55, 15059. [Google Scholar] [CrossRef] [PubMed]
- Vuust, P.; Heggli, O.A.; Friston, K.J.; Kringelbach, M.L. Music in the brain. Nat. Rev. Neurosci. 2022, 23, 287–305. [Google Scholar] [CrossRef]
- Alluri, V.; Toiviainen, P.; Burunat, I.; Kliuchko, M.; Vuust, P.; Brattico, E. Connectivity patterns during music listening: Evidence for action-based processing in musicians: Connectivity Patterns During Music Listening. Hum. Brain Mapp. 2017, 38, 2955–2970. [Google Scholar] [CrossRef] [PubMed]
- Aydogan, G.; Flaig, N.; Ravi, S.N.; Large, E.W.; McClure, S.M.; Margulis, E.H. Overcoming Bias: Cognitive Control Reduces Susceptibility to Framing Effects in Evaluating Musical Performance. Sci. Rep. 2018, 8, 6229. [Google Scholar] [CrossRef]
- Janata, P. The Neural Architecture of Music-Evoked Autobiographical Memories. Cereb. Cortex. 2009, 19, 2579–2594. [Google Scholar] [CrossRef]
- Juslin, P.N.; Laukka, P. Expression, Perception, and Induction of Musical Emotions: A Review and a Questionnaire Study of Everyday Listening. J. New Music Res. 2004, 33, 217–238. [Google Scholar] [CrossRef]
- Sloboda, J.A.; O’Neill, S.A.; Ivaldi, A. Functions of Music in Everyday Life: An Exploratory Study Using the Experience Sampling Method. Music. Sci. 2001, 5, 9–32. [Google Scholar] [CrossRef]
- Blood, A.J.; Zatorre, R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef]
- Salimpoor, V.N.; van den Bosch, I.; Kovacevic, N.; McIntosh, A.R.; Dagher, A.; Zatorre, R.J. Interactions Between the Nucleus Accumbens and Auditory Cortices Predict Music Reward Value. Science 2013, 340, 216–219. [Google Scholar] [CrossRef]
- Pereira, C.S.; Teixeira, J.; Figueiredo, P.; Xavier, J.; Castro, S.L.; Brattico, E. Music and Emotions in the Brain: Familiarity Matters. PLoS ONE 2011, 6, e27241. [Google Scholar] [CrossRef] [PubMed]
- Leggieri, M.; Thaut, M.H.; Fornazzari, L.; Schweizer, T.A.; Barfett, J.; Munoz, D.G.; Fischer, C.E. Music Intervention Approaches for Alzheimer’s Disease: A Review of the Literature. Front. Neurosci. 2019, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, G.; Macdonald, R. The effects of music choice on task performance: A study of the impact of self-selected and experimenter-selected music on driving game performance and experience. Music. Sci. 2009, 13, 357–386. [Google Scholar] [CrossRef]
- Arnal, L.H.; Doelling, K.B.; Poeppel, D. Delta–Beta Coupled Oscillations Underlie Temporal Prediction Accuracy. Cereb. Cortex. 2015, 25, 3077–3085. [Google Scholar] [CrossRef]
- Fujioka, T.; Trainor, L.J.; Large, E.W.; Ross, B. Internalized Timing of Isochronous Sounds Is Represented in Neuromagnetic Beta Oscillations. J. Neurosci. 2012, 32, 1791–1802. [Google Scholar] [CrossRef]
- Nozaradan, S.; Peretz, I.; Mouraux, A. Selective Neuronal Entrainment to the Beat and Meter Embedded in a Musical Rhythm. J. Neurosci. 2012, 32, 17572–17581. [Google Scholar] [CrossRef]
- Fujioka, T.; Ross, B.; Trainor, L.J. Beta-Band Oscillations Represent Auditory Beat and Its Metrical Hierarchy in Perception and Imagery. J. Neurosci. 2015, 35, 15187–15198. [Google Scholar] [CrossRef]
- Harding, E.E.; Sammler, D.; Henry, M.J.; Large, E.W.; Kotz, S.A. Cortical tracking of rhythm in music and speech. NeuroImage 2019, 185, 96–101. [Google Scholar] [CrossRef]
- Stefanics, G.; Hangya, B.; Hernadi, I.; Winkler, I.; Lakatos, P.; Ulbert, I. Phase Entrainment of Human Delta Oscillations Can Mediate the Effects of Expectation on Reaction Speed. J. Neurosci. 2010, 30, 13578–13585. [Google Scholar] [CrossRef]
- Will, U.; Berg, E. Brain wave synchronization and entrainment to periodic acoustic stimuli. Neurosci. Lett. 2007, 424, 55–60. [Google Scholar] [CrossRef]
- Woods, K.J.; Sampaio, G.; James, T.; Przysinda, E.; Spencer, A.E.; Morillon, B.; Loui, P. Stimulating music supports attention in listeners with attentional difficulties. bioRxiv 2021, 30. [Google Scholar] [CrossRef]
- Ding, N.; Patel, A.D.; Chen, L.; Butler, H.; Luo, C.; Poeppel, D. Temporal modulations in speech and music. Neurosci. Biobehav. Rev. 2017, 81, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Large, E.W.; Herrera, J.A.; Velasco, M.J. Neural Networks for Beat Perception in Musical Rhythm. Front. Syst. Neurosci. 2015, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Doelling, K.B.; Poeppel, D. Cortical entrainment to music and its modulation by expertise. Proc. Natl. Acad. Sci. USA 2015, 112, E6233–E6242. [Google Scholar] [CrossRef]
- Nozaradan, S.; Peretz, I.; Missal, M.; Mouraux, A. Tagging the Neuronal Entrainment to Beat and Meter. J. Neurosci. 2011, 31, 10234–10240. [Google Scholar] [CrossRef] [PubMed]
- Tal, I.; Large, E.W.; Rabinovitch, E.; Wei, Y.; Schroeder, C.E.; Poeppel, D.; Zion Golumbic, E. Neural Entrainment to the Beat: The “Missing-Pulse” Phenomenon. J. Neurosci. 2017, 37, 6331–6341. [Google Scholar] [CrossRef]
- Lerud, K.D.; Almonte, F.V.; Kim, J.C.; Large, E.W. Mode-locking neurodynamics predict human auditory brainstem responses to musical intervals. Hear. Res. 2014, 308, 41–49. [Google Scholar] [CrossRef]
- Skoe, E.; Krizman, J.; Spitzer, E.; Kraus, N. The auditory brainstem is a barometer of rapid auditory learning. Neuroscience 2013, 243, 104–114. [Google Scholar] [CrossRef]
- Skoe, E.; Kraus, N. Hearing It Again and Again: On-Line Subcortical Plasticity in Humans. PLoS ONE 2010, 5, e13645. [Google Scholar] [CrossRef]
- Tichko, P.; Skoe, E. Frequency-dependent fine structure in the frequency-following response: The byproduct of multiple generators. Hear. Res. 2017, 348, 1–15. [Google Scholar] [CrossRef]
- Tognoli, E.; Kelso, J.A.S. Brain coordination dynamics: True and false faces of phase synchrony and metastability. Prog. Neurobiol. 2009, 87, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Sauvé, S.A.; Bolt, E.L.W.; Nozaradan, S.; Zendel, B.R. Aging effects on neural processing of rhythm and meter. Front. Aging Neurosci. 2022, 14, 848608. [Google Scholar] [CrossRef]
- Alain, C.; Zendel, B.R.; Hutka, S.; Bidelman, G.M. Turning down the noise: The benefit of musical training on the aging auditory brain. Hear. Res. 2014, 308, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Bones, O.; Plack, C.J. Losing the Music: Aging Affects the Perception and Subcortical Neural Representation of Musical Harmony. J. Neurosci. 2015, 35, 4071–4080. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.J.; Herrmann, B.; Kunke, D.; Obleser, J. Aging affects the balance of neural entrainment and top-down neural modulation in the listening brain. Nat. Commun. 2017, 8, 15801. [Google Scholar] [CrossRef]
- Zendel, B.R.; Alain, C. Enhanced attention-dependent activity in the auditory cortex of older musicians. Neurobiol. Aging 2014, 35, 55–63. [Google Scholar] [CrossRef]
- Goodman, M.S.; Kumar, S.; Zomorrodi, R.; Ghazala, Z.; Cheam, A.S.M.; Barr, M.S.; Daskalakis, Z.J.; Blumberger, D.M.; Fischer, C.; Flint, A.; et al. Theta-Gamma Coupling and Working Memory in Alzheimer’s Dementia and Mild Cognitive Impairment. Front. Aging Neurosci. 2018, 10, 101. [Google Scholar] [CrossRef]
- Güntekin, B.; Başar, E. Review of evoked and event-related delta responses in the human brain. Int. J. Psychophysiol. 2016, 103, 43–52. [Google Scholar] [CrossRef]
- Hata, M.; Kazui, H.; Tanaka, T.; Ishii, R.; Canuet, L.; Pascual-Marqui, R.D.; Aoki, Y.; Ikeda, S.; Kanemoto, H.; Yoshiyama, K.; et al. Functional connectivity assessed by resting state EEG correlates with cognitive decline of Alzheimer’s disease–An eLORETA study. Clin. Neurophysiol. 2016, 127, 1269–1278. [Google Scholar] [CrossRef]
- Koenig, T.; Prichep, L.; Dierks, T.; Hubl, D.; Wahlund, L.O.; John, E.R.; Jelic, V. Decreased EEG synchronization in Alzheimer’s disease and mild cognitive impairment. Neurobiol. Aging 2005, 26, 165–171. [Google Scholar] [CrossRef]
- Reinhart, R.M.G.; Nguyen, J.A. Working memory revived in older adults by synchronizing rhythmic brain circuits. Nat. Neurosci. 2019, 22, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Iaccarino, H.F.; Singer, A.C.; Martorell, A.J.; Rudenko, A.; Gao, F.; Gillingham, T.Z.; Mathys, H.; Seo, J.; Kritskiy, O.; Abdurrob, F.; et al. Gamma frequency entrainment attenuates amyloid load and modifies microglia. Nature 2016, 540, 230–235. [Google Scholar] [CrossRef]
- Martorell, A.J.; Paulson, A.L.; Suk, H.-J.; Abdurrob, F.; Drummond, G.T.; Guan, W.; Young, J.Z.; Kim, D.N.-W.; Kritskiy, O.; Barker, S.J.; et al. Multi-sensory Gamma Stimulation Ameliorates Alzheimer’s-Associated Pathology and Improves Cognition. Cell 2019, 177, 256–271.e22. [Google Scholar] [CrossRef]
- Ashley, R. Do[n’t] Change a Hair for Me: The Art of Jazz Rubato. Music Percept. 2002, 19, 311–332. [Google Scholar] [CrossRef]
- Chapin, H.; Jantzen, K.; Scott Kelso, J.A.; Steinberg, F.; Large, E. Dynamic Emotional and Neural Responses to Music Depend on Performance Expression and Listener Experience. PLoS ONE 2010, 5, e13812. [Google Scholar] [CrossRef] [PubMed]
- Istók, E.; Friberg, A.; Huotilainen, M.; Tervaniemi, M. Expressive Timing Facilitates the Neural Processing of Phrase Boundaries in Music: Evidence from Event-Related Potentials. PLoS ONE 2013, 8, e55150. [Google Scholar] [CrossRef]
- Kim, J.C.; Large, E.W. Signal Processing in Periodically Forced Gradient Frequency Neural Networks. Front. Comput. Neurosci. 2015, 9, 152. [Google Scholar] [CrossRef]
- Kim, J.C.; Large, E.W. Mode locking in periodically forced gradient frequency neural networks. Phys. Rev. E 2019, 99, 022421. [Google Scholar] [CrossRef]
- Kim, J.C.; Large, E.W. Multifrequency Hebbian plasticity in coupled neural oscillators. Biol. Cybern. 2021, 115, 43–57. [Google Scholar] [CrossRef]
- Lambert, A.J.; Weyde, T.; Armstrong, N. Adaptive Frequency neural networks for dynamic pulse and metre perception. In Proceedings of the 17th ISMIR Conference, New York, NY, USA, 7–11 August 2016. [Google Scholar]
- Large, E.W.; Almonte, F.V.; Velasco, M.J. A canonical model for gradient frequency neural networks. Phys. Nonlinear Phenom. 2010, 239, 905–911. [Google Scholar] [CrossRef]
- Velasco, M.; Large, E. Pulse Detection in Syncopated Rhythms using Neural Oscillators. In Proceedings of the 12th International Society for Music Information Retrieval Conference, Miami, FL, USA, 24–28 October 2011; Volume 1, pp. 3–4. [Google Scholar]
- Tichko, P.; Kim, J.C.; Large, E.W. Bouncing the network: A dynamical systems model of auditory–vestibular interactions underlying infants’ perception of musical rhythm. Dev. Sci. 2021, 24, e13103. [Google Scholar] [CrossRef] [PubMed]
- Tichko, P.; Large, E.W. Modeling infants’ perceptual narrowing to musical rhythms: Neural oscillation and Hebbian plasticity. Ann. N. Y. Acad. Sci. 2019, 1453, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, T.; Chew, E. Detecting Low Frequency Oscillations in Cardiovascular Signals Using Gradient Frequency Neural Networks. In Proceedings of the 2019 Computing in Cardiology Conference, Singapore, 30 December 2019. [Google Scholar]
- Kim, J.C. A Dynamical Model of Pitch Memory Provides an Improved Basis for Implied Harmony Estimation. Front. Psychol. 2017, 8, 666. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bosch der Nederlanden, C.M.; Joanisse, M.F.; Grahn, J.A. Music as a scaffold for listening to speech: Better neural phase-locking to song than speech. NeuroImage 2020, 214, 116767. [Google Scholar] [CrossRef]
- Fiveash, A.; Schön, D.; Canette, L.-H.; Morillon, B.; Bedoin, N.; Tillmann, B. A stimulus-brain coupling analysis of regular and irregular rhythms in adults with dyslexia and controls. Brain Cogn. 2020, 140, 105531. [Google Scholar] [CrossRef]
- Mas-Herrero, E.; Marco-Pallares, J.; Lorenzo-Seva, U.; Zatorre, R.J.; Rodriguez-Fornells, A. Individual Differences in Music Reward Experiences. Music Percept. 2013, 31, 118–138. [Google Scholar] [CrossRef]
- Müllensiefen, D.; Gingras, B.; Musil, J.; Stewart, L. The Musicality of Non-Musicians: An Index for Assessing Musical Sophistication in the General Population. PLoS ONE 2014, 9, e89642. [Google Scholar] [CrossRef]
- Peretz, I.; Champod, A.S.; Hyde, K. Varieties of musical disorders: The Montreal Battery of Evaluation of Amusia. Ann. N. Y. Acad. Sci. 2003, 999, 58–75. [Google Scholar] [CrossRef]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in behavior made easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Mullen, T.R.; Kothe, C.A.E.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.-P.; Cauwenberghs, G. Real-time neuroimaging and cognitive monitoring using wearable dry EEG. IEEE Trans. Biomed. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Hsu, S.-H.; Pion-Tonachini, L.; Jung, T.-P. Evaluation of Artifact Subspace Reconstruction for Automatic Artifact Components Removal in Multi-Channel EEG Recordings. IEEE Trans. Biomed. Eng. 2020, 67, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage 2019, 198, 181–197. [Google Scholar] [CrossRef]
- Zilany, M.S.A.; Bruce, I.C. Modeling auditory-nerve responses for high sound pressure levels in the normal and impaired auditory periphery. J. Acoust. Soc. Am. 2006, 120, 1446–1466. [Google Scholar] [CrossRef] [PubMed]
- Bello, J.P.; Duxbury, C.; Davies, M.; Sandler, M. On the Use of Phase and Energy for Musical Onset Detection in the Complex Domain. IEEE Signal Process. Lett. 2004, 11, 553–556. [Google Scholar] [CrossRef]
- Cohen, M.X. Analyzing Neural Time Series Data: Theory and Practice; The MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Lalor, E.C.; Foxe, J.J. Neural responses to uninterrupted natural speech can be extracted with precise temporal resolution. Eur. J. Neurosci. 2010, 31, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Crosse, M.J.; Di Liberto, G.M.; Bednar, A.; Lalor, E.C. The Multivariate Temporal Response Function (mTRF) Toolbox: A MATLAB Toolbox for Relating Neural Signals to Continuous Stimuli. Front. Hum. Neurosci. 2016, 10, 604. [Google Scholar] [CrossRef]
- Canty, A.; Ripley, B.D. Boot: Bootstrap R (S-Plus) Functions. 2021. Available online: https://cran.r-project.org/web/packages/boot/index.html (accessed on 1 May 2022).
- Davison, A.C.; Hinkley, D.V. Bootstrap Methods and Their Applications; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Hall, J.W. Handbook of Auditory Evoked Responses; Allyn and Bacon: Boston, MA, USA, 1992. [Google Scholar]
- Riha, C.; Güntensperger, D.; Kleinjung, T.; Meyer, M. Accounting for Heterogeneity: Mixed-Effects Models in Resting-State EEG Data in a Sample of Tinnitus Sufferers. Brain Topogr. 2020, 33, 413–424. [Google Scholar] [CrossRef]
- DeBruine, L.M.; Barr, D.J. Understanding mixed effects models through data simulation. Adv. Methods Pract. Psychol. Sci. 2021, 4, 22. [Google Scholar] [CrossRef]
- Heckerman, D.; Gurdasani, D.; Kadie, C.; Pomilla, C.; Carstensen, T.; Martin, H.; Ekoru, K.; Nsubuga, R.N.; Ssenyomo, G.; Kamali, A.; et al. Linear mixed model for heritability estimation that explicitly addresses environmental variation. Proc. Natl. Acad. Sci. USA 2016, 113, 7377–7382. [Google Scholar] [CrossRef]
- Jaeger, B.C.; Edwards, L.J.; Das, K.; Sen, P.K. An R2 statistic for fixed effects in the generalized linear mixed model. J. Appl. Stat. 2017, 44, 1086–1105. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 May 2022).
- Cramer, A.O.J.; van Ravenzwaaij, D.; Matzke, D.; Steingroever, H.; Wetzels, R.; Grasman, R.P.P.P.; Waldorp, L.J.; Wagenmakers, E.-J. Hidden multiplicity in exploratory multiway ANOVA: Prevalence and remedies. Psychon. Bull. Rev. 2016, 23, 640–647. [Google Scholar] [CrossRef]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Belfi, A.M.; Moreno, G.L.; Gugliano, M.; Neill, C. Musical reward across the lifespan. Aging Ment. Health. 2021, 26, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Hove, M.J.; Marie, C.; Bruce, I.C.; Trainor, L.J. Superior time perception for lower musical pitch explains why bass-ranged instruments lay down musical rhythms. Proc. Natl. Acad. Sci. USA 2014, 111, 10383–10388. [Google Scholar] [CrossRef]
- Weineck, K.; Wen, O.X.; Henry, M.J. Neural synchronization is strongest to the spectral flux of slow music and depends on familiarity and beat salience. eLife 2022, 11, e75515. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrode Cluster | Electrodes |
|---|---|
| Left frontal (LF) | AF7, AF3, F7, F5, F3, F1 |
| Right frontal (RF) | AF4, AF8, F2, F4, F6, F8 |
| Left central (LC) | FT7, FC5, FC3, T7, C5, C3 |
| Midline central (MC) | FC1, FCz, FC2, C1, Cz, C2 |
| Right central (RC) | FC4, FC6, FC8, C4, C6, T8 |
| Left parietal (LP) | TP7, CP5, CP3, P7, P5, P3 |
| Midline parietal (MP) | CP1, CPZ, CP2, P1, Pz, P2 |
| Right parietal (RP) | CP4, CP6, CP8, P4, P6, P8 |
| Occipital (O) | PO3, POZ, PO4, O1, OZ, O2 |
| All Participants | Younger Adults | Older Adults | ||
|---|---|---|---|---|
| Age | Mean | 45.38 | 19.81 | 70.94 |
| SD | 26.66 | 1.60 | 8.48 | |
| BMRQ | Mean | 75.88 | 80.75 | 71.00 |
| SD | 13.85 | 10.75 | 15.17 | |
| Gold-MSI | Mean | 184.19 | 203.69 | 164.69 |
| SD | 46.06 | 36.79 | 47.12 | |
| MBEA | Mean | 23.09 | 23.94 | 22.25 |
| SD | 3.74 | 3.73 | 3.68 |
| (a) | ||||
| Pulse Level: Mixed Model ANOVA Table (Type-III Tests, S-method) | ||||
| Effect | DF | F | p-Value | Semi-Partial R2 |
| Rhythmic_Level | 1, 29.00 | 35.17 *** | <0.001 | 0.144 |
| Age_Group | 1, 29.00 | 0.13 | 0.717 | 0.005 |
| Rhythmic_Level:Age_Group | 1, 29.00 | 0.2111 | 0.649 | 0.002 |
| (b) | ||||
| Pulse Level: Mixed Model ANOVA Table (Type-III Tests, S-method) | ||||
| Effect | DF | F | p-Value | Holm p-Value |
| Electrode_Group | 8, 57.13 | 2.58 | 0.018 | 0.070 |
| Age_Group | 1, 29.00 | 0.40 | 0.532 | 1.00 |
| Rhythmic_Level | 1, 29.00 | 8.97 * | 0.006 | 0.029 |
| Electrode_Group:Age_Group | 8, 57.13 | 0.49 | 0.860 | 1.00 |
| Electrode_Group:Rhythmic_Level | 8, 3006.57 | 5.53 *** | <0.001 | <0.001 |
| Age_Group:Rhythmic_Level | 1, 29.00 | 0.30 | 0.590 | 1.00 |
| Electrode_Group:Age_Group:Rhythmic_Level | 8, 3006.57 | 7.53 *** | <0.001 | <0.001 |
| Sub-Harmonic Level: Mixed Model ANOVA Table (Type-III Tests, S-method) | ||||
|---|---|---|---|---|
| Effect | DF | F | p-Value | Holm p-Value |
| Electrode_Group | 8, 62.24 | 3.22 * | 0.004 | 0.020 |
| Age_Group | 1, 29.00 | 0.16 | 0.693 | 1.00 |
| Rhythmic_Level | 1, 29.00 | 10.33 * | 0.003 | 0.019 |
| Electrode_Group:Age_Group | 8, 62.24 | 0.60 | 0.773 | 1.00 |
| Electrode_Group:Rhythmic_Level | 8, 3013.38 | 8.23 *** | <0.001 | <0.001 |
| Age_Group:Rhythmic_Level | 1, 29.00 | 0.01 | 0.934 | 1.00 |
| Electrode_Group:Age_Group:Rhythmic_Level | 8, 3013.38 | 1.74 | 0.085 | 0.341 |
| Harmonic Level: Mixed Model ANOVA Table (Type-III Tests, S-method) | ||||
|---|---|---|---|---|
| Effect | DF | F | p-Value | Holm p-Value |
| Electrode_Group | 8, 69.39 | 0.29 | 0.966 | 1.00 |
| Age_Group | 1, 29.00 | 0.01 | 0.929 | 1.00 |
| Rhythmic_Level | 1, 29.00 | 11.33 * | 0.002 | 0.01 |
| Electrode_Group:Age_Group | 8, 69.39 | 1.02 | 0.432 | 1.00 |
| Electrode_Group:Rhythmic_Level | 8, 3014.81 | 7.43 *** | < 0.001 | <0.001 |
| Age_Group:Rhythmic_Level | 1, 29.00 | 0.68 | 0.415 | 1.00 |
| Electrode_Group:Age_Group:Rhythmic_Level | 8, 3014.81 | 4.74 *** | < 0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tichko, P.; Page, N.; Kim, J.C.; Large, E.W.; Loui, P. Neural Entrainment to Musical Pulse in Naturalistic Music Is Preserved in Aging: Implications for Music-Based Interventions. Brain Sci. 2022, 12, 1676. https://doi.org/10.3390/brainsci12121676
Tichko P, Page N, Kim JC, Large EW, Loui P. Neural Entrainment to Musical Pulse in Naturalistic Music Is Preserved in Aging: Implications for Music-Based Interventions. Brain Sciences. 2022; 12(12):1676. https://doi.org/10.3390/brainsci12121676
Chicago/Turabian StyleTichko, Parker, Nicole Page, Ji Chul Kim, Edward W. Large, and Psyche Loui. 2022. "Neural Entrainment to Musical Pulse in Naturalistic Music Is Preserved in Aging: Implications for Music-Based Interventions" Brain Sciences 12, no. 12: 1676. https://doi.org/10.3390/brainsci12121676
APA StyleTichko, P., Page, N., Kim, J. C., Large, E. W., & Loui, P. (2022). Neural Entrainment to Musical Pulse in Naturalistic Music Is Preserved in Aging: Implications for Music-Based Interventions. Brain Sciences, 12(12), 1676. https://doi.org/10.3390/brainsci12121676