Microbial Ecology Evaluation of an Iberian Pig Processing Plant through Implementing SCH Sensors and the Influence of the Resident Microbiota on Listeria monocytogenes


 ,
,
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Approach
2.2. Surface Sensors
2.3. Sampling Procedure and Recovery of Adhered Microorganisms
2.4. Identification Methodology
2.5. Statistical Analysis of the Data
3. Results and Discussion
3.1. Identified Species
3.2. Ecological Profiles of the Different Areas: Cluster Analysis
3.3. Unusual Identification of Mannheimia haemolytica
3.4. Influence of the Microbiota on the Presence of L. Monocytogenes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fratamico, P.M.; Annous, B.A.; Gunther, N.W. Biofilms in the Food and Beverage Industries; Elsevier: Amsterdam, The Netherlands, 2009; ISBN 978-1-84-569362-6. [Google Scholar]
- Dantas, S.T.A.; Rossi, B.F.; Bonsaglia, E.C.R.; Castilho, I.G.; Hernandes, R.T.; Fernandes, A.; Rall, V.L.M. Cross-contamination and biofilm formation by Salmonella enterica serovar Enteritidis on various cutting boards. Foodborne Pathog. Dis. 2018, 15, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Dourou, D.; Simpson, C.; Yoon, Y.; Belk, K.E.; Smith, G.C.; Nychas, G.E.; Sofos, J.N. Attachment and biofilm formation by E. coli O157:H7 at different temperatures, on various food-contact surfaces encountered in beef processing. Int. J. Food Microbiol. 2011, 149, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Grove, S.F.; Suriyanarayanan, A.; Puli, B.; Zhao, H.; Li, M.; Li, D.; Schaffner, D.W.; Lee, A. Norovirus cross-contamination during preparation of fresh produce. Int. J. Food Microbiol. 2015, 198, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.A.; Danyluk, M.D.; Harris, L.J.; Schaffner, D.W. Quantifying bacterial cross-contamination rates between fresh-cut produce and hands. J. Food Prot. 2017, 80, 213–219. [Google Scholar] [CrossRef]
- Kim, A.; Young Park, S.; Bae, S.-C.; Oh, M.-H.; Ha, S.-D. Survival of norovirus surrogate on various food-contact surfaces. Food Environ. Virol. 2014, 6, 182–188. [Google Scholar] [CrossRef]
- Sauders, B.D.; D’Amico, D.J. Listeria monocytogenes cross-contamination of cheese: Risk throughout the food supply chain. Epidemiol. Infect. 2016, 144, 2693–2697. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S. Residential bacteria on surfaces in the food industry and their implications for food safety and quality. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.J.; Kacániová, M.; Czaczyk, K.; et al. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 1–26. [Google Scholar] [CrossRef]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Carpentier, B.; Chassaing, D. Interactions in biofilms between Listeria monocytogenes and resident microorganisms from food industry premises. Int. J. Food Microbiol. 2004, 97, 111–122. [Google Scholar] [CrossRef]
- Mariani, C.; Oulahal, N.; Chamba, J.F.; Dubois-Brissonnet, F.; Notz, E.; Briandet, R. Inhibition of Listeria monocytogenes by resident biofilms present on wooden shelves used for cheese ripening. Food Control 2011, 22, 1357–1362. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S.; Heir, E. Bacteria on meat abattoir process surfaces after sanitation: Characterisation of survival properties of Listeria monocytogenes and the commensal bacterial flora. Adv. Microbiol. 2013, 3, 255–264. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Villatoro, D.; López, P.; Dávila, J.C.; Martínez-Suárez, J.V. A 3-year surveillance of the genetic diversity and persistence of Listeria monocytogenes in an iberian pig slaughterhouse and processing plant. Foodborne Pathog. Dis. 2010, 7, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; Huffel, M.-A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoët, A.S.; Martínez-Suárez, J.V.; Capita, R.; Rodríguez-Jerez, J.J. Evaluation of the microbiological contamination of food processing environments through implementing surface sensors in an iberian pork processing plant: An approach towards the control of Listeria monocytogenes. Food Control 2019, 99, 40–47. [Google Scholar] [CrossRef]
- Montañez-Izquierdo, V.; Ríos-Castillo, A.G.; Fontecha-Umaña, F.; Rodríguez-Jerez, J.J. Uso de sensores de superficie y métodos rápidos para controlar la contaminación ambiental en las industrias alimentarias: Hacia el control on line. In XI Workshop de Métodos Rápidos y Automatización en Microbiología Alimentaria; Universidad Autónoma de Barcelona: Barcelona, Spain, 2012; pp. 1–6. [Google Scholar]
- Moen, B.; Røssvoll, E.; Måge, I.; Møretrø, T.; Langsrud, S. Microbiota formed on attached stainless steel coupons correlates with the natural biofilm of the sink surface in domestic kitchens. Can. J. Microbiol. 2016, 62, 148–160. [Google Scholar] [CrossRef]
- Ripolles-Avila, C. Supervivencia de Listeria monocytogenes Sobre Superficies de Contacto con Alimentos: Un Abordaje Multidisciplinar de un Problema Complejo. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2018. [Google Scholar]
- Sharma, M.; Anand, S.K. Characterization of constitutive microflora of biofilms in dairy processing lines. Food Microbiol. 2002, 19, 627–636. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Sabiel, Y.A. Detection of microbial contamination of processed beef meat by ssing API strips and automated Vitek 2 compact system. Microbiol. Res. J. Int. 2016, 13, 1–8. [Google Scholar] [CrossRef]
- Feligini, M.; Panelli, S.; Buffoni, J.N.; Bonacina, C.; Andrighetto, C.; Lombardi, A. Identification of microbiota present on the surface of Taleggio cheese using PCR-DGGE and RAPD-PCR. J. Food Sci. 2012, 77, 609–615. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.E. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef]
- Bagge-Ravn, D.; Ng, Y.; Hjelm, M.; Christiansen, J.N.; Johansen, C.; Gram, L. The microbial ecology of processing equipment in different fish industries—Analysis of the microflora during processing and following cleaning and disinfection. Int. J. Food Microbiol. 2003, 87, 239–250. [Google Scholar] [CrossRef]
- Dzieciol, M.; Schornsteiner, E.; Muhterem-Uyar, M.; Stessl, B.; Wagner, M.; Schmitz-Esser, S. Bacterial diversity of floor drain biofilms and drain waters in a Listeria monocytogenes contaminated food processing environment. Int. J. Food Microbiol. 2016, 223, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Maes, S.; Heyndrickx, M.; Vackier, T.; Steenackers, H.; Verplaetse, A.; De Reu, K. Identification and spoilage potential of the remaining dominant microbiota on food contact surfaces after cleaning and disinfection in different food industries. J. Food Prot. 2019, 82, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Mertz, A.W.; Kyung, O.; Bryan, C.A.O.; Morawicki, R.; Sirsat, S.A.; Neal, J.A.; Crandall, P.G.; Ricke, S.C. Microbial ecology of meat slicers as determined by denaturing gradient gel electrophoresis. Food Control 2014, 42, 242–247. [Google Scholar] [CrossRef]
- Stellato, G.; La Storia, A.; De Filippis, F.; Borriello, G.; Villani, F.; Ercolini, D. Overlap of spoilage-associated microbiota between meat and the meat processing environment in small-scale and large-scale retail distributions. Appl. Environ. Microbiol. 2016, 82, 4045–4054. [Google Scholar] [CrossRef] [PubMed]
- Marouani-Gadri, N.; Augier, G.; Carpentier, B. Characterization of bacterial strains isolated from a beef-processing plant following cleaning and disinfection-influence of isolated strains on biofilm formation by Sakaï and EDL 933 E. coli O157:H7. Int. J. Food Microbiol. 2009, 133, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Hoodt, S.K.; Zottola, E.A. Isolation and identification of adherent gram-negative microorganisms from four meat-processing facilities. J. Food Prot. 1997, 60, 1135–1138. [Google Scholar] [CrossRef]
- Schön, K.; Schornsteiner, E.; Wagner, M.; Müller, M.; Schmitz-Esser, S.; Dzieciol, M. Microbial communities in dairy processing environment floor-drains are dominated by product-associated bacteria and yeasts. Food Control 2016, 70, 210–215. [Google Scholar] [CrossRef]
- Bañón, S.; Martínez, A.; López, A.M. Maduración de chorizo y salchichón de Chato Murciano con diferentes cultivos iniciadores (bacterias ácido-lácticas y estafilococos). An. Vet. Murcia 2011, 27, 101–118. [Google Scholar] [CrossRef]
- Hultman, J.; Rahkila, R.; Ali, J.; Rousu, J.; Johanna, K. Meat processing plant microbiome and contamination patterns of cold-tolerant bacteria causing food safety and spoilage risks in the manufacture of vacuum-packaged cooked sausages. Appl. Environ. Microbiol. 2015, 81, 7088–7097. [Google Scholar] [CrossRef]
- Padilla-Frausto, J.J.; Cepeda-Marquez, L.G.; Salgado, L.M.; Iturriaga, M.H.; Arvizu-Medrano, S.M. Detection and genotyping of Leuconostoc spp. in a sausage processing plant. J. Food Prot. 2015, 78, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Gounadaki, A.S.; Skandamis, P.N.; Drosinos, E.H.; Nychas, G.E. Microbial ecology of food contact surfaces and products of small-scale facilities producing traditional sausages. Food Microbiol. 2008, 25, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, I.; Ammor, S.; Laguet, A.; Labayle, S.; Castanet, V.; Dufour, E.; Talon, R. Microbial ecology of a small-scale facility producing traditional dry sausage. Food Control 2006, 17, 446–453. [Google Scholar] [CrossRef]
- Talon, R.; Leroy, S.; Lebert, I. Microbial ecosystems of traditional fermented meat products: The importance of indigenous starters. Meat Sci. 2007, 77, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Adler, R. Pseudomonas. Salem Press Encyclopedia of Health. Available online: http://mendeley.csuc.cat/fitxers/3b01d5716b505758e1d15ab62b7d7601 (accessed on 29 October 2019).
- Silva, O.F. Aeromonas spp. Rev. Chil. Infectol. 2011, 28, 157–158. [Google Scholar] [CrossRef]
- Cheney, R. Corynebacterium. Salem Press Encyclopedia of Health. Available online: http://mendeley.csuc.cat/fitxers/b50c6dd66f3a8698da8be0571a312e98 (accessed on 29 October 2019).
- Mühlhauser, M. Leifsonia aquatica. Rev. Chil. Infectol. 2016, 33, 313–314. [Google Scholar] [CrossRef]
- Kte’pi, B. Bacillus (bacteria). Salem Press Encyclopedia of Health. Available online: http://mendeley.csuc.cat/fitxers/315e4f3c33439a6298ababd24553641d (accessed on 29 October 2019).
- Stamou, A.; Pavlopoulos, C.; Roumeliotis, S.; Samoladas, E.; Xatzokos, I.; Kontopoulou, K. Nonunion humerous fracture infection caused by Rhizobium radiobacter in a 24-year-old healthy patient: A rare case report. Case Rep. Infect. Dis. 2018, 2018, 1–4. [Google Scholar] [CrossRef]
- Stiles, M.E.; Ng, L.K. Enterobacteriaceae associated with meats and meat handling. Appl. Environ. Microbiol. 1981, 41, 867–872. [Google Scholar]
- Silva, F.; Pabla Martínez, O.; Pabla, T.M. Complejo Enterobacter cloacae. Rev. Chil. Infectol. 2018, 35, 297–298. [Google Scholar] [CrossRef]
- Hara, C.M.O.; Rhoden, D.L.; Miller, J.M. Reevaluation of the API 20E identification system versus conventional biochemicals for identification of members of the family Enterobacteriaceae: A new look at an old product. J. Clin. Microbiol. 1992, 30, 123–125. [Google Scholar]
- Jaramillo-Arango, C.J.; Trigo Tavera, F.J.; Suárez-Güemes, F. Mannheimiosis bovina: Etiología, prevención y control. Vet. Mex. 2009, 40, 293–314. [Google Scholar]
- Angen, Ø.; Ahrens, P.; Bisgaard, M. Phenotypic and genotypic characterization of Mannheimia (Pasteurella) haemolytica-like strains isolated from diseased animals in Denmark. Vet. Microbiol. 2002, 84, 103–114. [Google Scholar] [CrossRef]
- Puga, C.H.; Dahdouh, E.; SanJose, C.; Orgaz, B. Listeria monocytogenes colonizes Pseudomonas fluorescens biofilms and induces matrix over-production. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jessen, B.; Lammert, L. Biofilm and disinfection in meat processing plants. Int. Biodeterior. Biodegrad. 2003, 51, 265–269. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Cervantes-Huaman, B.H.; Hascoët, A.S.; Yuste, J.; Rodríguez-Jerez, J.J. Quantification of mature Listeria monocytogenes biofilm cells formed by an in vitro model: A comparison of different methods. Int. J. Food Microbiol. 2019, 289, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Ripolles-Avila, C.; Hascoët, A.S.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
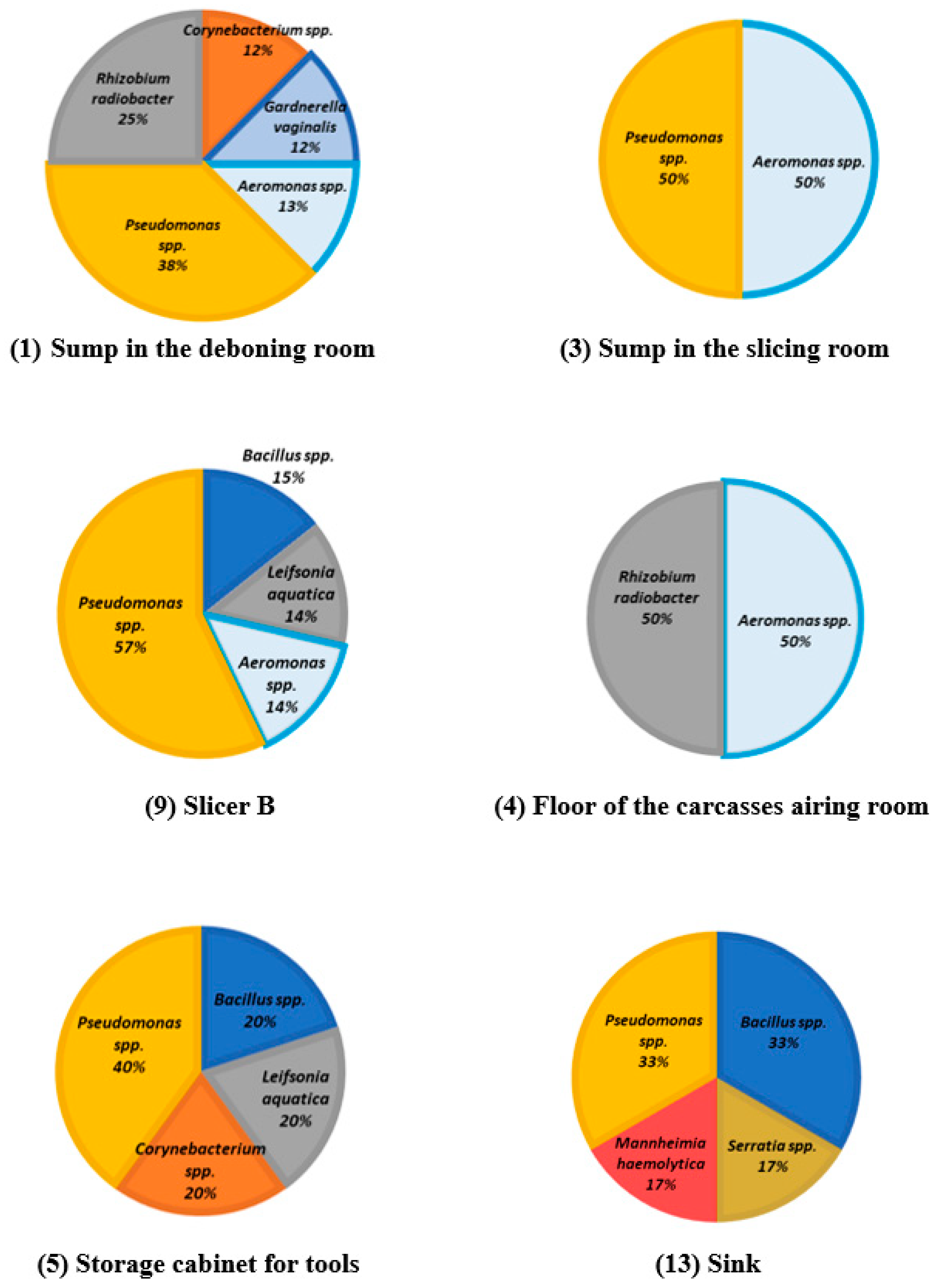
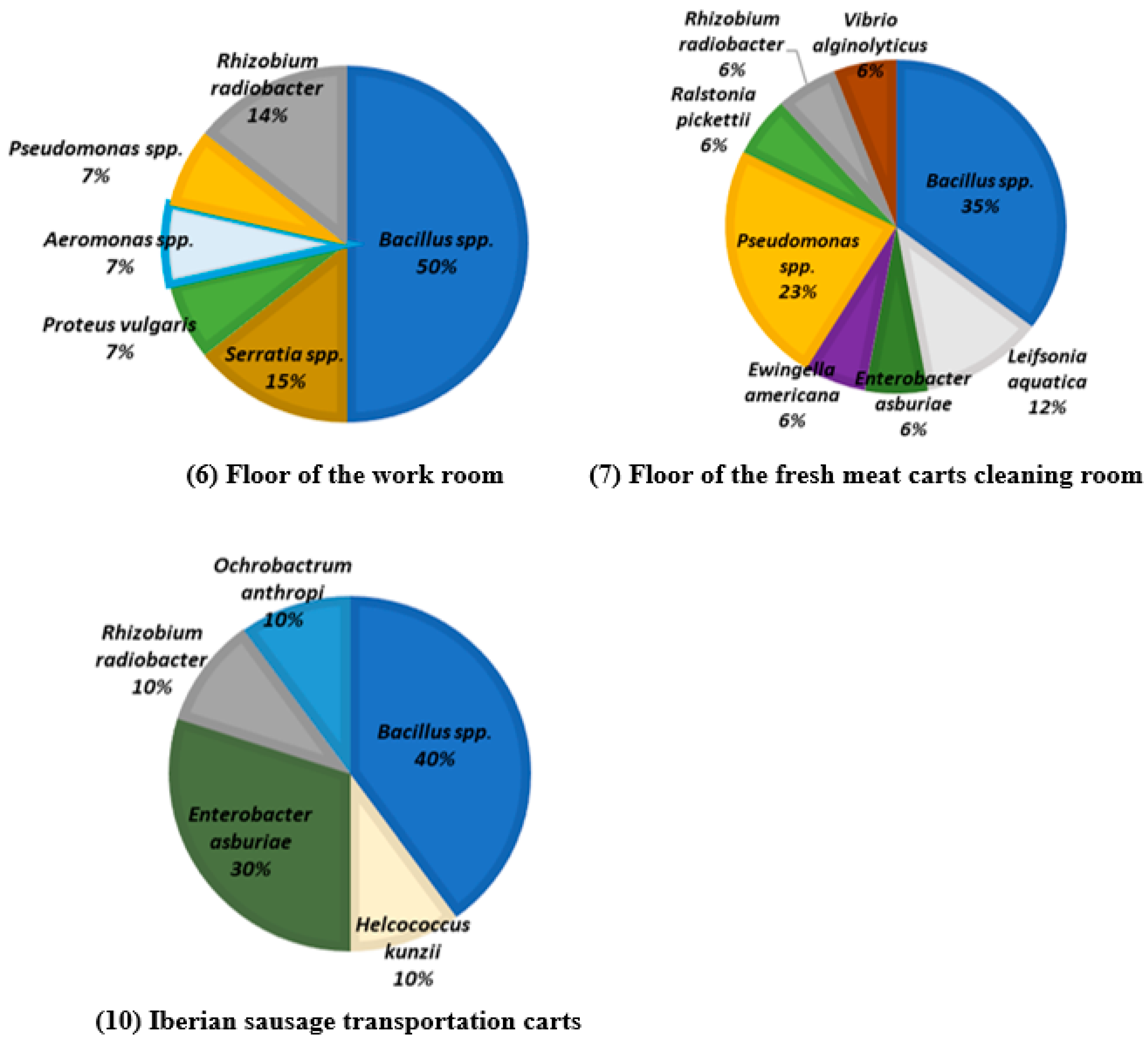
| Processing Plant | ID | Surface |
|---|---|---|
| A | 1 | Sump in the deboning room |
| 2 | Slicer A | |
| 3 | Sump in the slicing room | |
| B | 4 | Floor of the carcasses airing room |
| 5 | Storage cabinet for tools | |
| 6 | Floor of the work room | |
| 7 | Floor of the fresh meat carts cleaning room | |
| 8 | Floor of the cured meat carts cleaning room | |
| 9 | Slicer B | |
| 10 | Iberian sausage transportation carts | |
| 11 | Slide of vacuum machine | |
| 12 | Floor of the heat treatment room | |
| 13 | Sink |
| Identified Species | % vs Isolated Bacteria | % vs Isolated Microorganisms |
|---|---|---|
| Molds and yeasts | - | 29.03 |
| Bacillus spp.1 | 28.18 | 20.00 |
| Pseudomonas spp.2 | 21.82 | 15.48 |
| Mannheimia haemolytica | 9.09 | 6.45 |
| Rhizobium radiobacter | 7.27 | 5.16 |
| Staphylococcus spp. | 6.36 | 4.52 |
| Aeromonas spp. | 5.45 | 3.87 |
| Leifsonia aquatica | 3.64 | 2.58 |
| Serratia spp. | 3.64 | 2,58 |
| Enterobacter asburiae | 3.64 | 2.58 |
| Proteus vulgaris | 1.82 | 1.29 |
| Corynebacterium spp. | 1.82 | 1.29 |
| Helcococcus kunzii | 0.91 | 0.65 |
| Aerococcus urinae | 0.91 | 0.65 |
| Gardnerella vaginalis | 0.91 | 0.65 |
| Stenotrophomonas maltophilia | 0.91 | 0.65 |
| Ewingella americana | 0.91 | 0.65 |
| Ralstonia pickettii | 0.91 | 0.65 |
| Vibrio spp. | 0.91 | 0.65 |
| Ochrobactrum anthropi | 0.91 | 0.65 |
| Total | 100.00 | 100.00 |
| Gram stain | % | |
| Gram positive | 42.73 | - |
| Gram negative | 57.27 | - |
| Total | 100.00 | - |
| Identified Species | Total | % |
|---|---|---|
| Leuconostoc mesenteroides ssp. cremoris | 23.0 | 51.1 |
| Lactobacillus delbrueckii ssp. delbrueckii | 8.0 | 17.8 |
| Lactococcus lactis ssp. lactis | 6.0 | 13.3 |
| Leuconostoc mesenteroides ssp. mesenteroides | 4.0 | 8.9 |
| Lactobacillus acidophilus | 1.0 | 2.2 |
| Pediococcus damniosus | 1.0 | 2.2 |
| Non-identified | 2.0 | 4.4 |
| Total | 45.0 | 100.0 |
| Identified Species | Total | % |
|---|---|---|
| Candida zeylanoides | 27.0 | 61.4 |
| Cryptococcus uniguttulatus | 3.0 | 6.8 |
| Candida krusei | 2.0 | 4.5 |
| Candida ciferri | 2.0 | 4.5 |
| Candida spp. | 2.0 | 4.5 |
| Candida pelliculosa | 1.0 | 2.3 |
| Candida magnoliae | 1.0 | 2.3 |
| Cryptococcus terreus | 1.0 | 2.3 |
| Non-identified | 5.0 | 11.4 |
| Total | 44.0 | 100.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hascoët, A.-S.; Ripolles-Avila, C.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Microbial Ecology Evaluation of an Iberian Pig Processing Plant through Implementing SCH Sensors and the Influence of the Resident Microbiota on Listeria monocytogenes. Appl. Sci. 2019, 9, 4611. https://doi.org/10.3390/app9214611
Hascoët A-S, Ripolles-Avila C, Guerrero-Navarro AE, Rodríguez-Jerez JJ. Microbial Ecology Evaluation of an Iberian Pig Processing Plant through Implementing SCH Sensors and the Influence of the Resident Microbiota on Listeria monocytogenes. Applied Sciences. 2019; 9(21):4611. https://doi.org/10.3390/app9214611
Chicago/Turabian StyleHascoët, Anne-Sophie, Carolina Ripolles-Avila, Alfons Eduard Guerrero-Navarro, and José Juan Rodríguez-Jerez. 2019. "Microbial Ecology Evaluation of an Iberian Pig Processing Plant through Implementing SCH Sensors and the Influence of the Resident Microbiota on Listeria monocytogenes" Applied Sciences 9, no. 21: 4611. https://doi.org/10.3390/app9214611
APA StyleHascoët, A.-S., Ripolles-Avila, C., Guerrero-Navarro, A. E., & Rodríguez-Jerez, J. J. (2019). Microbial Ecology Evaluation of an Iberian Pig Processing Plant through Implementing SCH Sensors and the Influence of the Resident Microbiota on Listeria monocytogenes. Applied Sciences, 9(21), 4611. https://doi.org/10.3390/app9214611