1. Introduction
Due to recent changes in legislation both at the federal and state levels, there has been a surge of interest in the growing, processing, selling, and using of products containing cannabidiol (CBD), derived from hemp flowers. Hemp is legally defined as
Cannabis sativa strains with a tetrahydrocannabinol (THC) concentration no greater than 0.3% in any part of the plant (Congress, [
1,
2]).
Cannabis sativa strains with a THC concentration greater than 0.3% in any part of the plant are considered marijuana.
Cannabis sativa contains over 100 cannabinoids, which include THC and CBD. It is well known that THC has psychoactive effects. Many have reported health benefits from marijuana, which may be associated with non-THC cannabinoids, such as CBD. The broad interest in CBD is for health benefits similar to marijuana but without the psychoactive effects of THC.
Hemp has historically been grown for fiber and seed, and due to recent changes in legislation, it is being grown for flowers. Hemp grown for flowers (floral hemp) follows a horticultural production model either in a greenhouse or bedded field compared to fiber and seed hemp, which follows an agronomic production model.
There is little published research investigating fertility requirements for floral hemp. As applied research is conducted to determine nutrient rates to maximize yield and minimize inputs, as well as to develop a target range of plant sufficiency ranges to aid in nutrient management, this study provides an invaluable basis to identify nutrient deficiency in the field and to develop sufficiency ranges where nutrient corrective action can be made before visual symptoms are expressed.
The impacts of plant nutrition on plant growth and yield, as well as plant primary and secondary metabolites, are well-established (Il’in, [
3]). In hemp fiber varieties, Bosca et al. [
4] reported higher levels of nitrogen increased plant leaf weight and decreased leaf THC content, presumably due to THC dilution. In a marijuana strain, phosphorus treatments had greater combined leaf and flower dry weight, as well as higher THC concentration, compared to the no phosphorus treatment (Coffman and Gentner, [
5,
6]). Hemp producers are seeking THC levels <0.3% and high CBD concentrations (i.e., 10–20%). Given the high energy and resource requirements for plants to produce secondary plant metabolites, such as cannabinoids (Taura et al., [
7,
8]), it would be reasonable to assume that nutritive disorders would impact the production and quality of these metabolites.
Plant tissue analysis has been used extensively for many decades to evaluate the nutritional status of a crop. Nutrient sufficiency levels (the tissue concentration at which growth or yield is not limited) have been established for most major agricultural and horticultural plants. Because cannabis has not been widely grown legally, nutrient sufficiency ranges have not been established. The development of sufficiency ranges for the essential plant nutrients would require extensive rate studies that measure both elemental concentrations in the leaf and the yield parameters of both floral biomass and cannabinoid concentrations. Where sufficiency ranges of a crop are not available, survey ranges can be used to approximate the nutritional status of the plant. Survey ranges for Cannabis sativa in greenhouse nursery production have been published by Bryson et al. [
9], and more recently, a survey of five hemp cultivars in greenhouse production, including the cultivar used in this study, by Landis et al. [
10]. These tissue values are useful for cannabis growers as they aid in fertility management. This study adds to this body of knowledge as the complete fertilizer controls can serve as an additional set of survey ranges for cannabis.
The second contribution of this study results in information about nutrient deficiency levels in the leaf tissue of cannabis. Once a plant begins showing visual symptoms of impaired growth, there is a reduction in plant health or yield is implicit. In this study, when plants began showing deficiency symptoms for each nutrient, most recently, mature leaf samples were analyzed for that nutrient. This information can be used by growers and researchers to confirm the visual diagnosis with leaf concentrations.
Additionally, no visual guides of nutrient deficiencies in cannabis supported with leaf tissue analysis and documenting a progression of symptomology have been published. Tracking the specific symptomologies of various nutritional disorders over time is important because symptomologies change in appearance and location as the deficiency progresses, making correct diagnosis challenging. Therefore, this study was conducted to provide cannabis growers and researchers with descriptions of nutrient disorders, high-quality images to track the progression of these disorders, and leaf tissue nutrient concentrations associated with documented deficiency symptomology.
2. Materials and Methods
Cuttings were taken from a hemp Cannabis sativa ‘T1’on 3 July 2018 (Ryes Greenhouses: Sanford, NC, USA) and stuck into 72-cell plug trays filled with a substrate mix of 80:20 (v:v) Canadian sphagnum peat moss (Conrad Fafard, Agawam, MA, USA) and horticultural coarse perlite (Perlite Vermiculite Packaging Industries, Inc., North Bloomfield, OH, USA) amended with dolomitic lime at 8.875 kg/m3 (Rockydale Agricultural, Roanoke, VA, USA) and wetting agent (Aquatrols, Cherry Hill, NJ, USA) at 600.3 g/m3. After three weeks of rooting, plugs were transplanted (27 July 2018) into 15.24-cm diameter (1.76 L) plastic pots filled with acid-washed silica-sand (Millersville #2 (0.8 to 1.2 mm diameter) from Southern Products and Silica Co., Hoffman, NC, USA).
The experiment was conducted in a glass greenhouse in Raleigh, NC, USA (35°N latitude), with 24°C/20°C (D/N) temperature setpoints. Plugs were transplanted, and nutrient treatments started immediately in the automated, recirculating irrigation system made from 10.2-cm diameter PVC pipe (Charlotte Plastics, Charlotte, NC, USA), fit with 12.7-cm diameter openings to hold the pots. Control plants were grown with a complete modified Hoagland’s all-nitrate solution consisting of 15 mM nitrate-nitrogen (NO
3−), 1 mM phosphate-phosphorus (H
2PO
4−), 6 mM potassium (K
+), 5 mM calcium (Ca
2+), 2 mM magnesium (Mg
2+), and 2 mM sulfate-sulfur (SO
42−) (Hoagland and Arnon, [
11]); plus 72 µM iron (Fe
2+), 18 µM manganese (Mn
2+), 3 µM copper (Cu
2+), 3 µM zinc (Zn
2+), 45 µM boron (BO
33−), and 0.1 µM molybdenum (MoO
42−). Nutrient deficiencies began at transplant and were induced by withholding a single nutrient from this solution. Boron (B) and manganese (Mn) toxicities were induced by increasing the concentration tenfold higher than the complete nutrient formula. Reagent grade chemicals and deionized (DI) water (18 MΩ) were used to formulate treatment solutions. The plants were drip-irrigated with a sump-pump (model 1A, Little Giant Pump Co., Oklahoma City, OK, USA) system as needed between 6:00 and 18:00 hours. Irrigation solution drained from the pot and was captured for reuse. Nutrient solutions were replaced weekly. The experiment was terminated 9 weeks after treatments began.
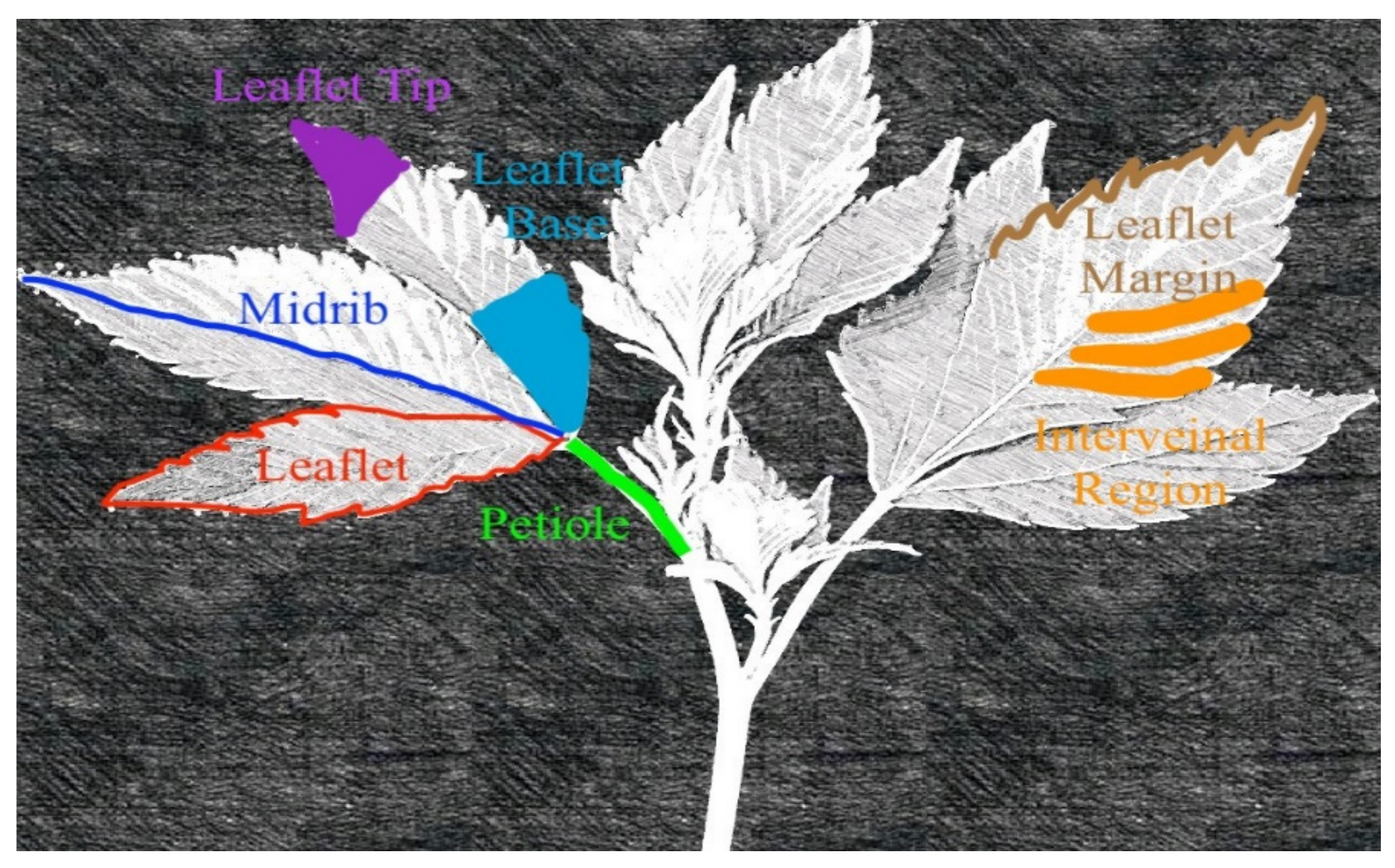
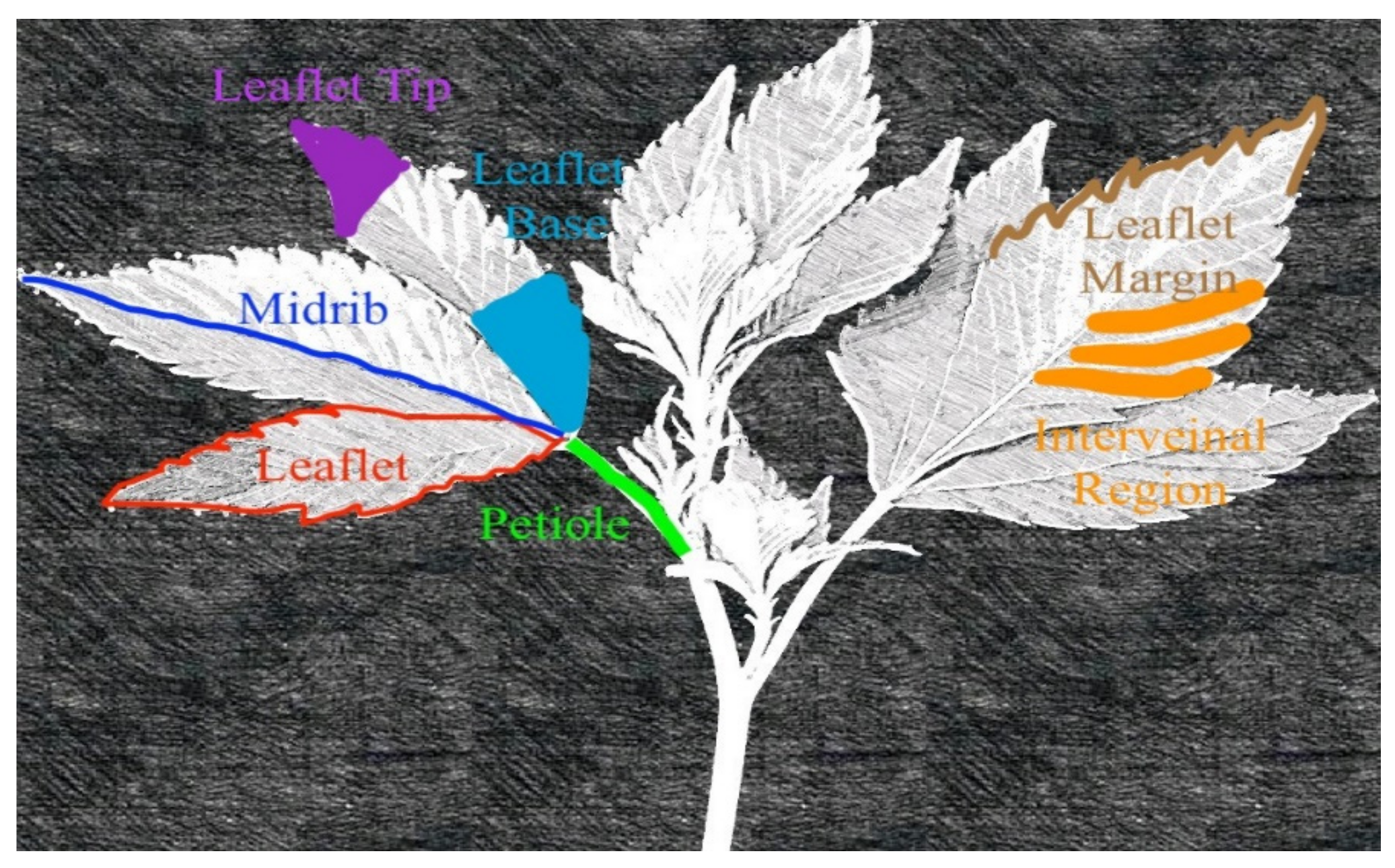
Plants were tracked daily, and deficiency symptoms were photographed at the initial, intermediate, and advanced stages of symptomology. Plant anatomy terminology used to describe deficiency symptoms is given in
Figure 1. Upon initial symptom development, four plants were selected for sampling. The remaining treatments, which were not symptomatic, were grown until visual symptoms appeared and then were harvested. At the onset of initial visual symptomology, whole plants (n=4) were destructively harvested, and most recent mature leaves (MRML) were subsampled and rinsed with DI water, washed in a solution of 0.5 M HCl, and again rinsed with DI water. Leaf tissue (MRML) was taken below the meristematic stem regions (apical meristem, axillary meristems, tertiary meristematic regions, etc.), and only recently matured leaves were sampled. Leaf maturity and morphology were determined based on observational leaf maturity and from indices hybridized from Heslop–Harrison and Heslop–Harrison [
12] and Mediavilla et al. [
13] and from established standard leaf tissue harvesting protocols (Bryson et al., [
9]). The remaining plant material after MRML was harvested and was placed in a separate container for aerial tissue biomass determination. Leaf tissues and their respective remaining biomass were dried at 70 °C for 24 hours. Dried leaf tissue was ground in a mill (Thomas Wiley
® Mini-Mill; Thomas Scientific, Swedesboro, NJ, USA) with a 20-mesh (1 mm) screen and analyzed for nutrient concentrations by the North Carolina Department of Agriculture and Consumer Services (NCDA&CS) Agronomic Division (NCDA&CS, [
14]). Total plant dry weight (DW) was calculated by adding the oven-dry weight of the leaf tissue to the oven-dry weight of the remaining plant biomass (
Table 1). Details about experimental setup, fertilizers, and design can be found in Barnes et al. [
15]. Data were analyzed with SAS version 9.4 (SAS Institute, Cary, NC, USA) and subjected to analysis of variance (ANOVA) using PROC ANOVA and (GLM) PROC GLM. Where
F-tests indicated evidence of significant differences among means, LSD (Least Significant Difference) (
P ≤ 0.05) was used.
4. Discussion
While some anecdotal deficiency symptoms for cannabis are available in lay publications, scientifically rigorous symptomology, particularly symptomological progression, is very limited or non-existent in the literature. In this study, most documented symptoms of deficiencies were consistent with descriptions from current literature for other plant species (Bryson et al., [
9]; Barnes, [
16]; Barker and Pilbeam, [
17]; Gibson et al., [
18]) with some exceptions. Concentrations of N, P, K, Ca, Mg, and S in the leaves when deficiency symptomology first appeared were 1.62, 0.09, 0.41, 0.39, 0.12, and 0.11%, respectively. Concentrations of B, Cu, Fe, Mn, Mo, and Zn in the leaves when deficiency symptomology first appeared were 2.46, 1.41, 60.1, 7.56, 0.06, and 10.7 mg·kg
−1, respectively.
Plants grown without Mo did not exhibit leaf symptomology nor less dry matter production despite leaf tissue values being 96% lower than the complete controls. While visual symptoms of Cu deficiency displayed the interveinal chlorosis in younger leaves documented in other plants, the Cu-deficient cannabis also showed an odd wilting pattern (
Figure 4). This wilting in Cu-deficient hemp plants is mentioned in fiber hemp (Van der Werf, [
19]). Copper is important in cell wall metabolism (Yruela, [
20]). The condition which Van der Werf [
19] referred to as “’gummi’-hemp” may indicate that Cu is needed in greater quantities in hemp than in other species, especially when grown for fiber. While these symptoms of wilting and lodging were in fiber hemp, which has a very vigorous vertical growth habit, the same process could be occurring in floral hemp cultivars.
Additionally, B-deficient treated plants displayed similar wilting tendencies. Upon further inspection of the roots, it was shown that the terminal growing tip had died and turned necrotic, and there were many axillary roots due to the loss of apical dominance. Boron is important in cell wall development and elongation of plant cell walls, especially in the radicle and other root meristematic regions (Hu and Brown, [
21]; Whittington, [
22]). The wilting seen could have been due to a lack of root mass due to an underdeveloped root system. Given all plants were irrigated for the same amount of time, it is feasible to assume that the wilting was due to a lack of water uptake because root growth was impaired.
Nutrients varied in how rapidly deficiency symptoms were visually apparent. In treatments where N, P, Ca, Mg, S, Mn, and B were withheld, deficiency symptoms were observed within 6 to 7 weeks. Conversely, where K, Zn, Cu, Fe were withheld, visual deficiencies only occurred after 9 weeks and, as previously noted, not at all in Mo. The use of visual symptomology as a diagnostic tool may be more useful for the nutrients that demonstrate symptoms sooner rather than later. However, for field and greenhouse floral hemp production, visual symptoms may express early enough in the season where rescue nutrient applications can be made. Additionally, routine MRML tissue sampling during the season will identify if nutrients are getting near the concentrations that show symptoms, thus allowing a correcting nutrient application before symptoms are expressed.
Some nutrient deficiencies had a significant suppressive effect on yield, as measured by whole plant dry weight, while withholding other nutrients did not affect total biomass. Deficiencies of N, K, B, and Cu produced significantly less crop biomass by 50, 27, 28, and 45%, respectively, as compared to the control. While all other nutrient deficiencies had less biomass, none suppressed yield at a significance level of P > 0.05. Above-ground biomass may not be the appropriate metric for testing critical levels in these nutrients, and other factors, such as secondary metabolite production, maybe a better indicator of the minimum nutrient concentrations for optimum production.
The development of true cannabis sufficiency ranges for each essential element requires individual rate studies coupled with measurements of both dry matter production and cannabinoid yield. Until this extensive work can be completed, survey ranges are useful tools for estimating adequate nutrient levels. In this study, the control samples served as survey ranges, which can be compared to other published survey ranges for cannabis (Bryson et al., [
9]; Landis et al., [
10]). Some elements in the control samples were above (K and S) or below (Cu and Mn) the reported values from Bryson et al. [
9]. Other control values found in this study were within or above the listed values for Cannabis sativa ‘T1’ except for Mn values, which were below the listed values from Landis et al. [
10]. While the ranges reported by Bryson et al. [
9] are useful guides for estimating healthy nutrient levels in hemp tissue, it is important to remember that they are the survey ranges from plants grown in a production nursery of an unknown replicate number and unknown hemp cultivar and may not accurately predict optimum nutrient levels in all production systems in all cultivars. Recently, Landis et al. [
10] found that significant differences in nutrient concentrations occurred among CBD cultivars, suggesting that broader target nutrient ranges may be appropriate for cannabis.
Boron toxicity was first observed at an accumulated foliar leaf tissue concentration of 671.75 mg·kg
−1, and the symptoms (marginal chlorosis and necrosis in the older leaves) were consistent with B toxicity in other plants. Plants vary in their sensitivity to boron. Toxicity symptoms occur in strawberry at foliar concentrations as low as 120 mg·kg
−1 (Haydon, [
23]), suggesting that cannabis may be more tolerant of excess B than some crops.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}