An Amalgamation of Hormone Inspired Arbitration Systems for Application in Robot Swarms



Abstract
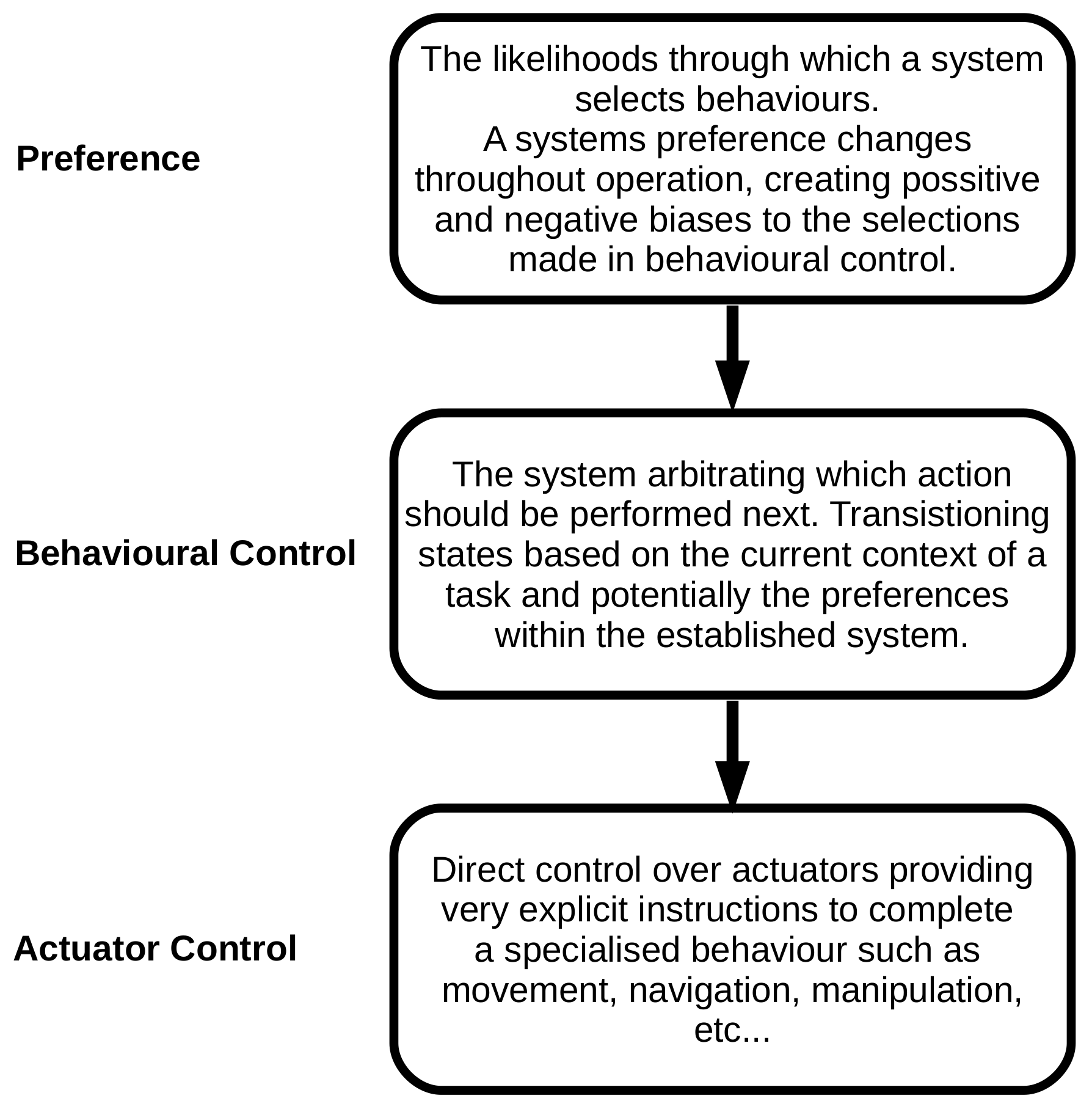
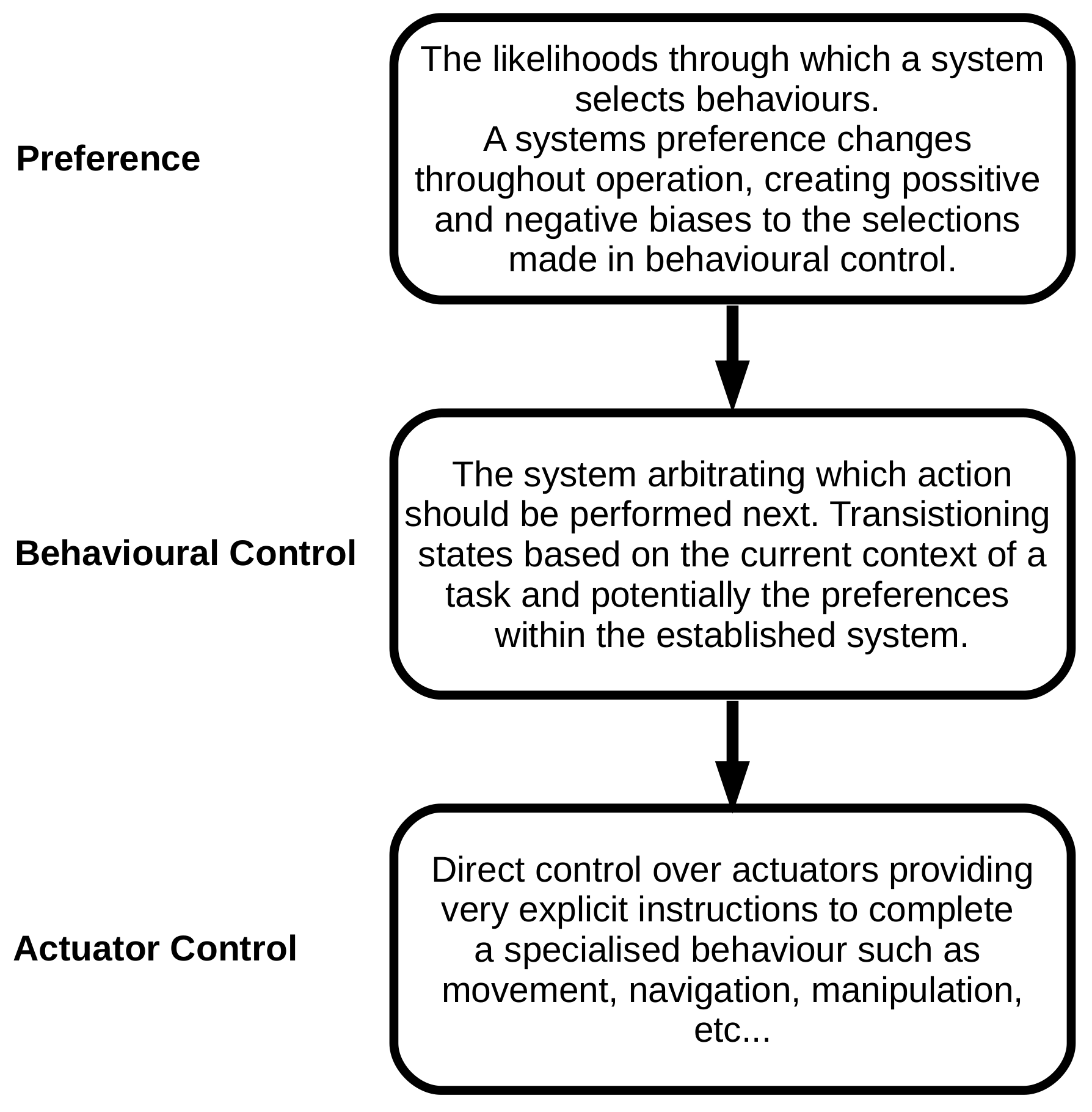
:1. Introduction
2. Background
3. HIBAS Implementation for Control of a Foraging System with Deviating Motor Speeds
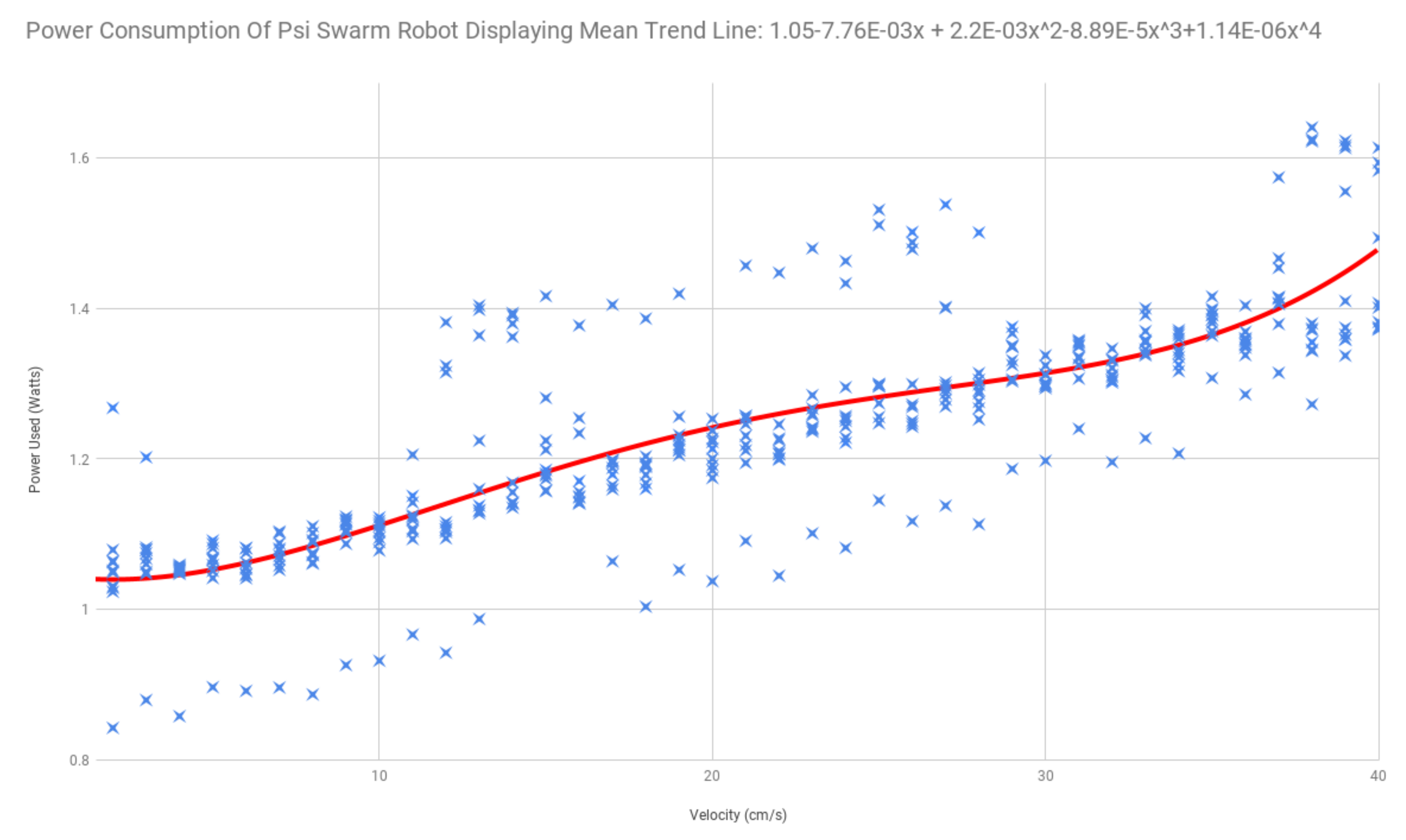
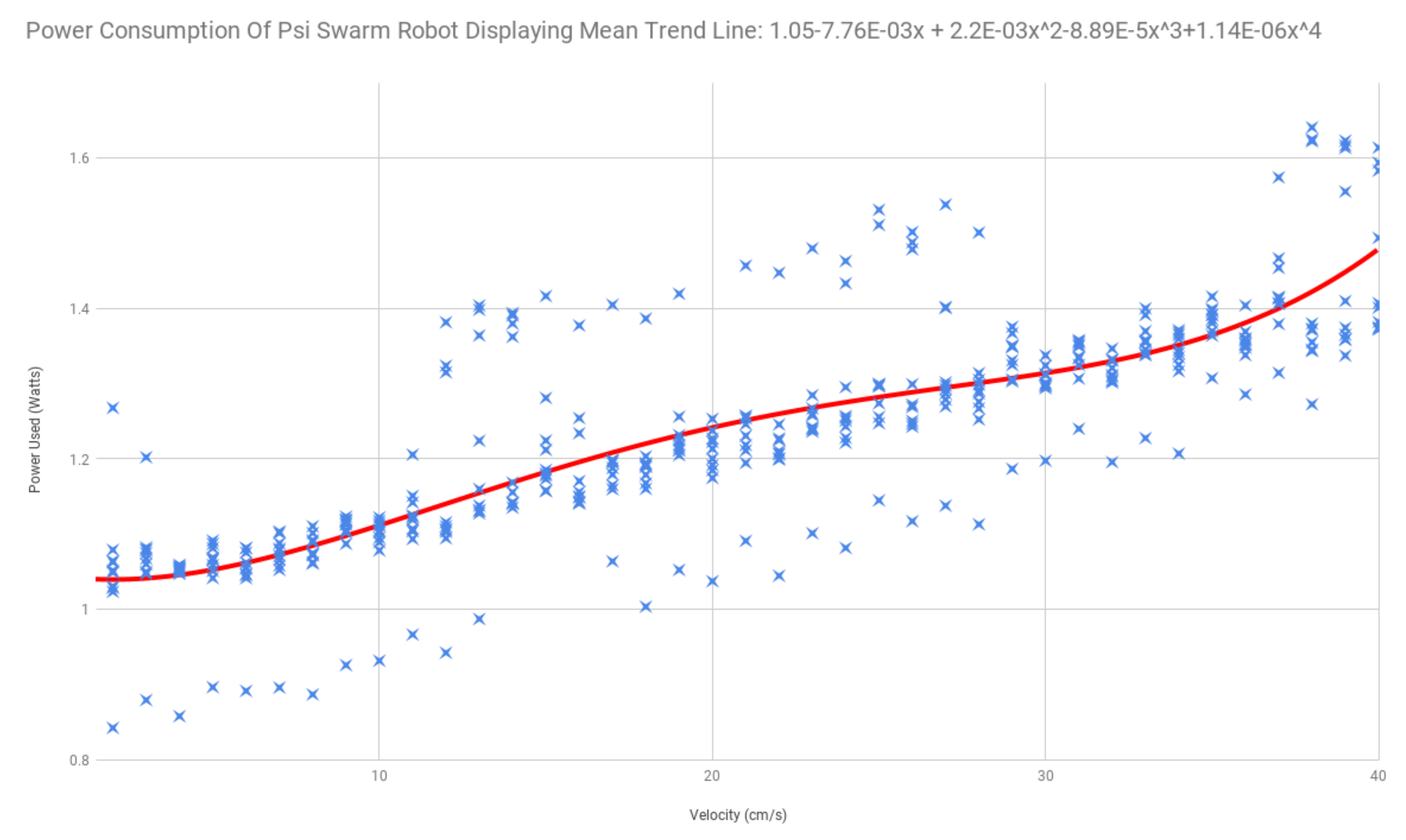
3.1. Energy Characteristics of Psi Swarm Robot Hardware
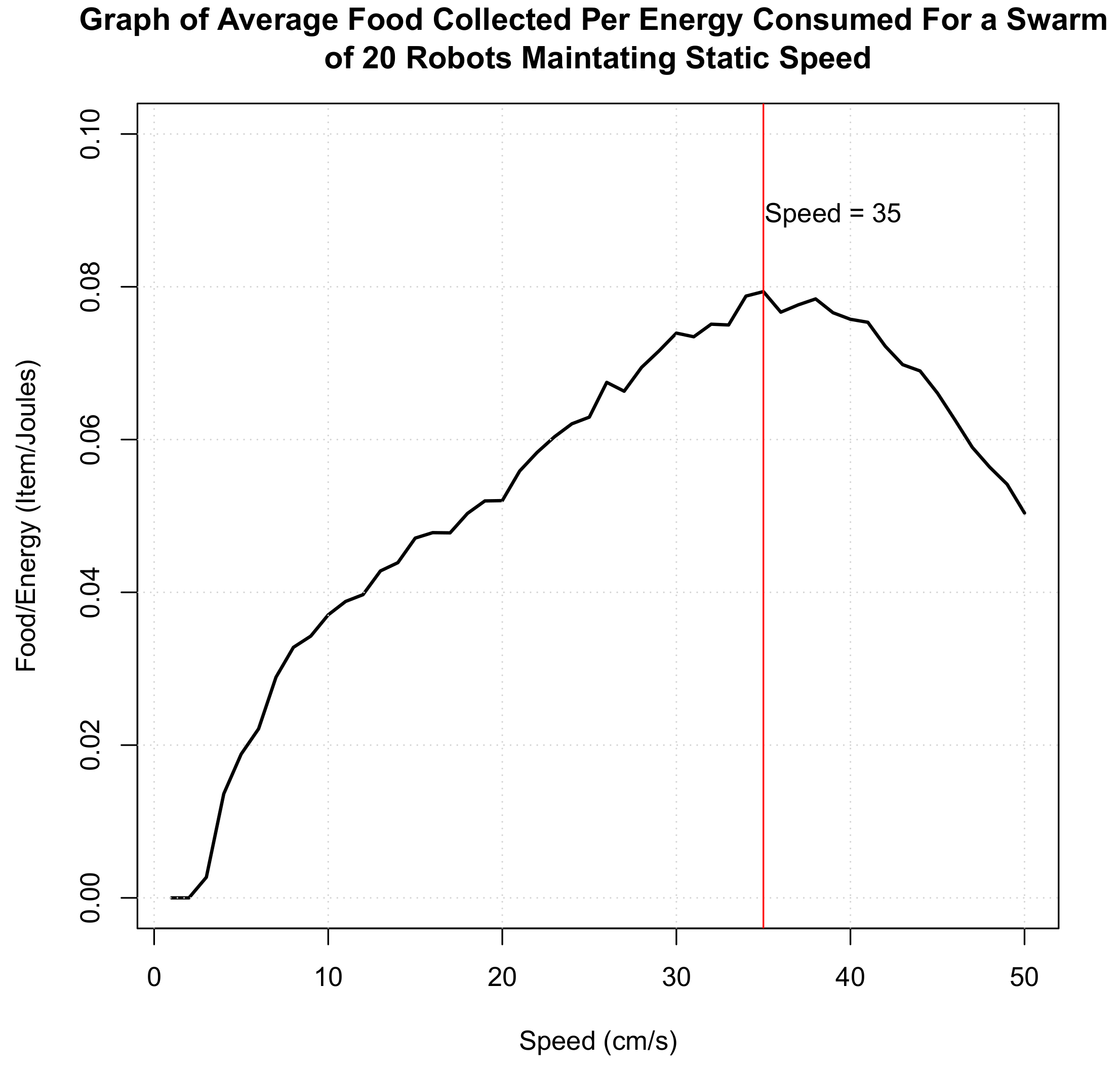
3.2. Hormone Interaction with Motor Speed
3.2.1. Demand
3.2.2. Return Hormone
3.2.3. Speed Hormone
3.2.4. Parameters
3.3. Comparison Systems
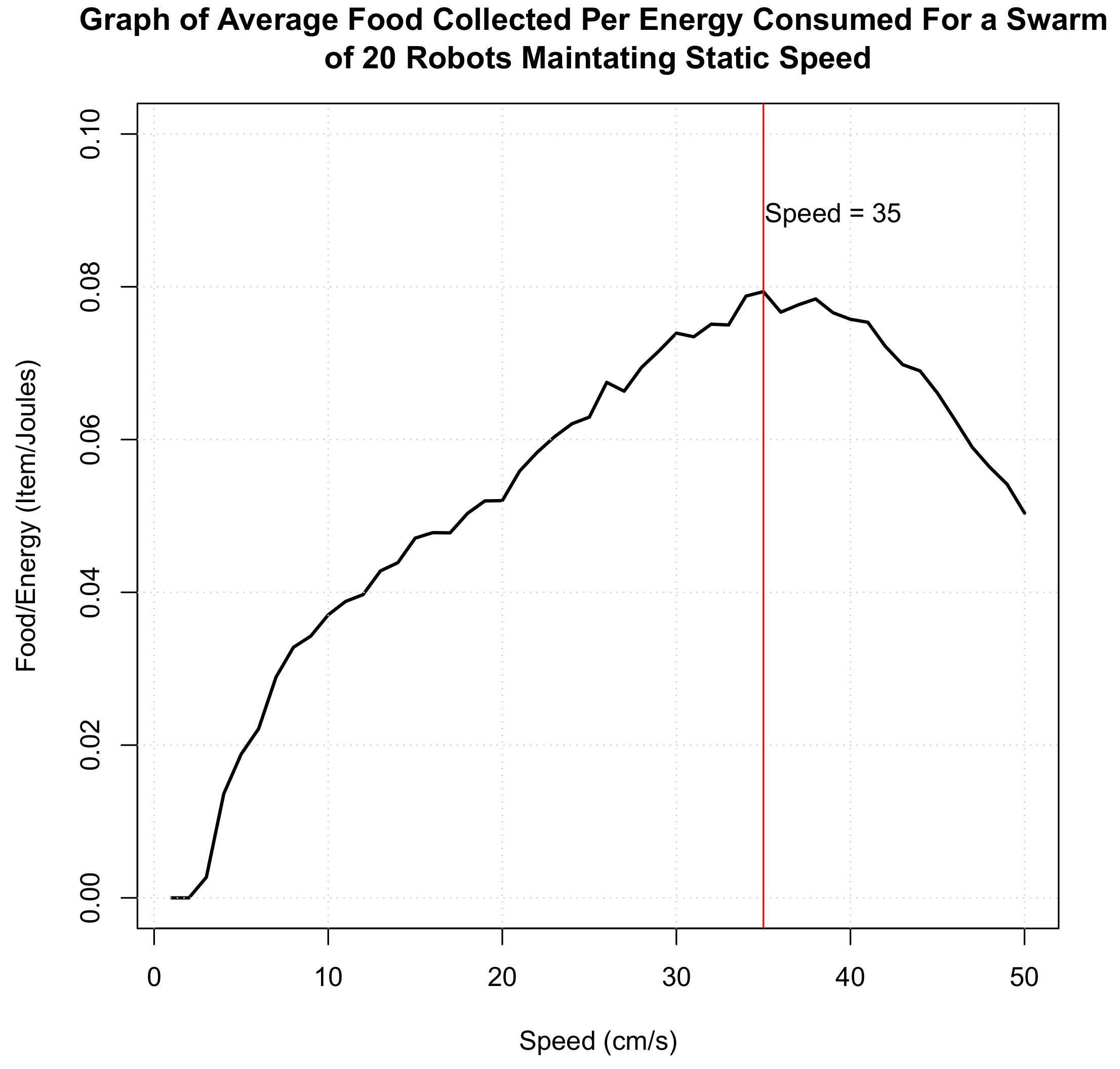
3.4. Analysis of Systems Highlighting the Need for Adaptation
3.4.1. Environments
3.4.2. Simulation
3.5. Results
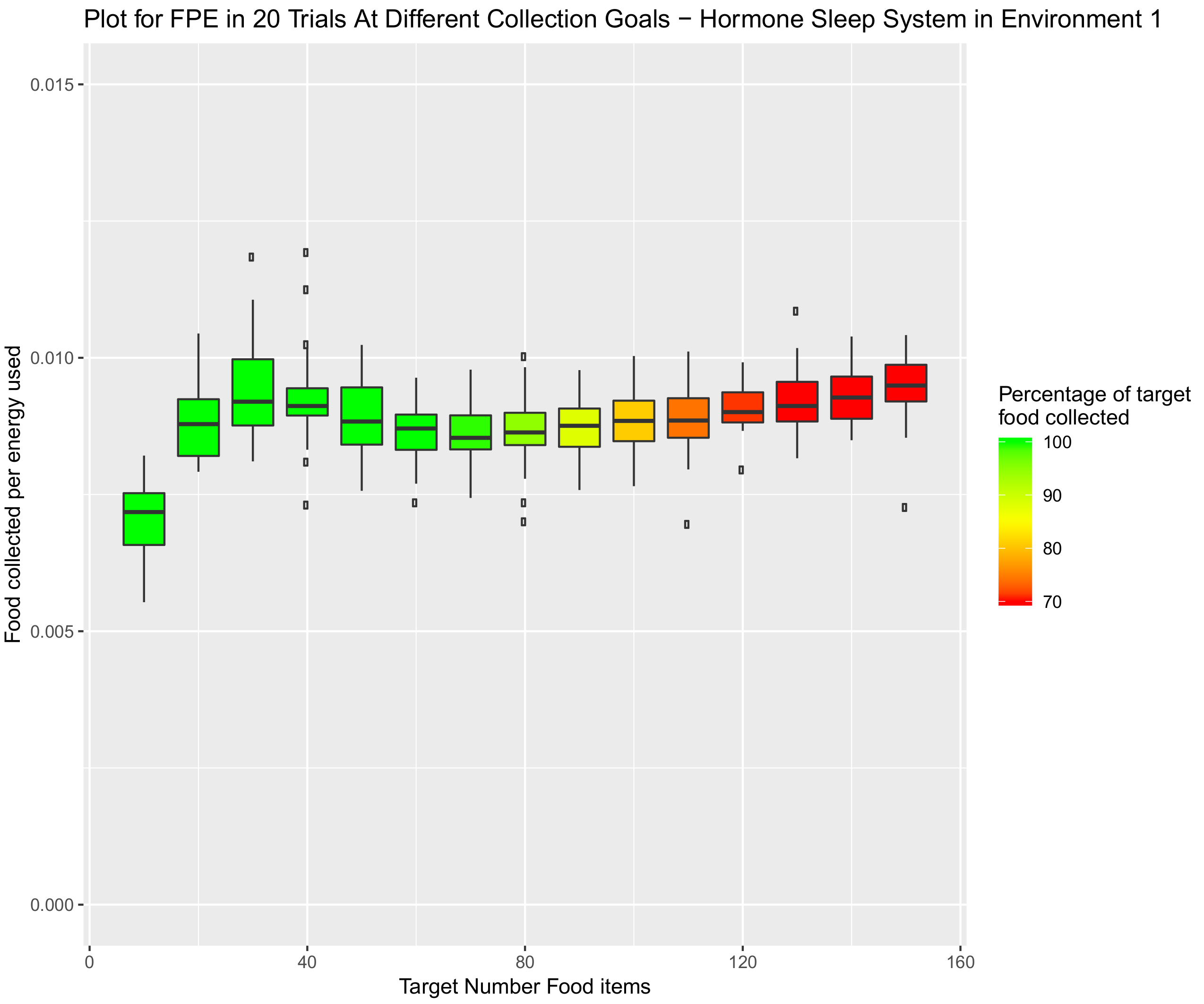
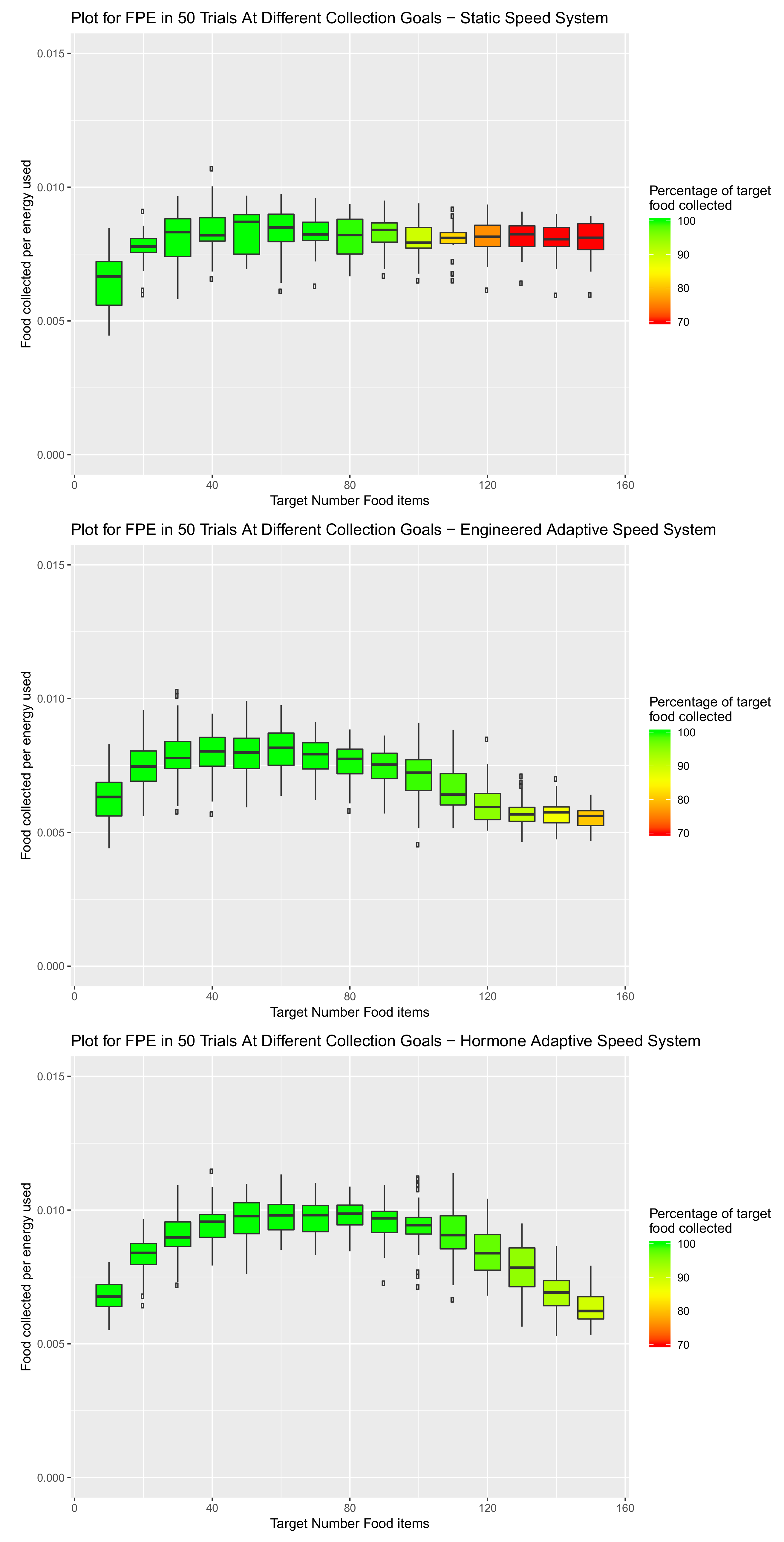
3.5.1. Environment 1—Square Open Arena
- Sensitivity:
- The hormone system is sensitive to collisions and capable of not only returning robots to the nest due to collisions, but also reducing speed due to the prolonged influence of collisions.
- Dispersion:
- Rather than consistent speeds, or speeds of specific increments, the speeds of the hormone driven robots fluctuate during the search. Thus, dispersion is a by-product of efficiency as speed will be diverse amongst the swarm. This in turn will lead to less traffic and more energy efficient item collection.
- Gradual variability:
- Speed can build over the duration of a search. This is contrary to the engineered system, which made relatively large (and potentially exaggerated) changes in speed on an individual’s return to the nest.
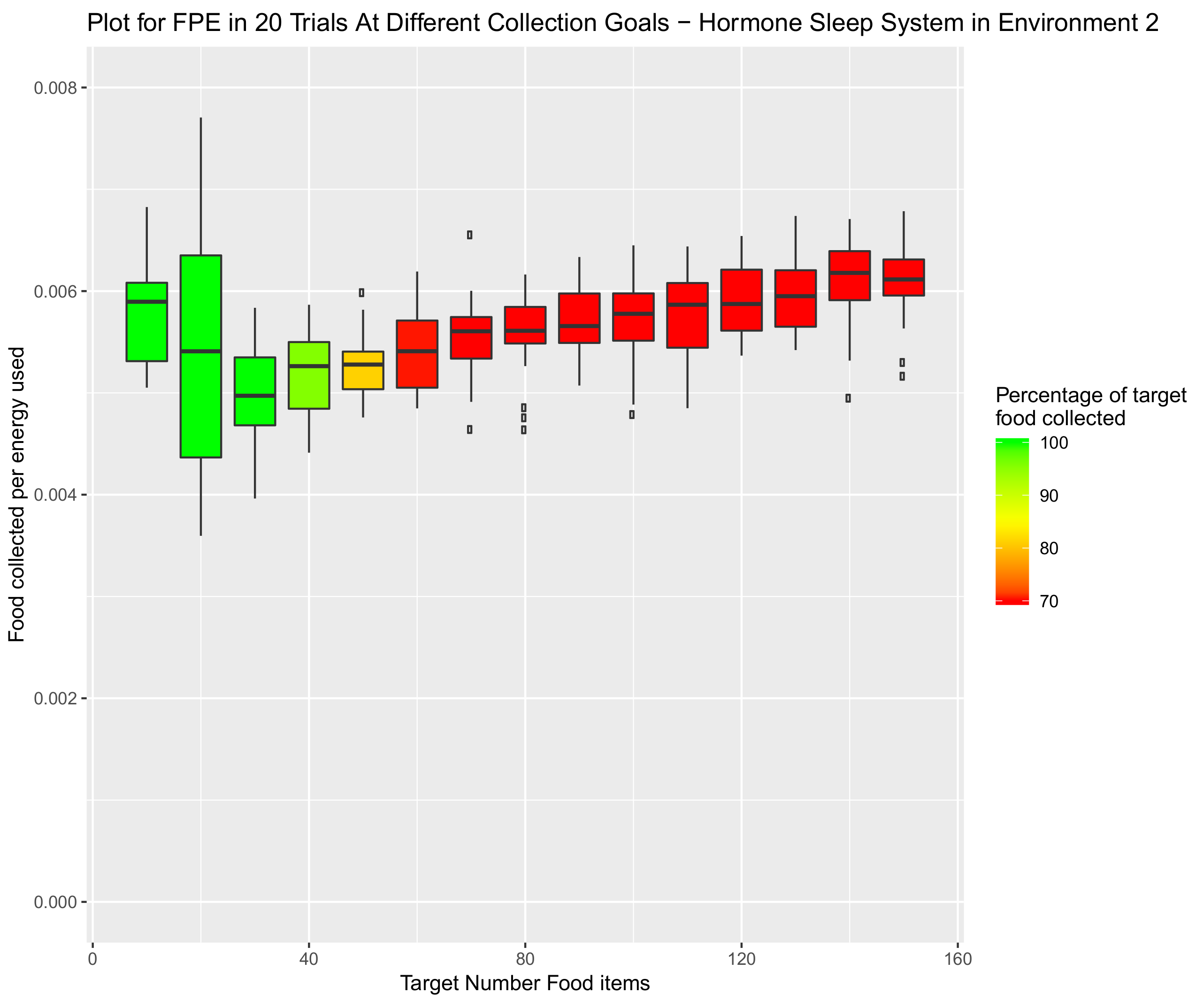
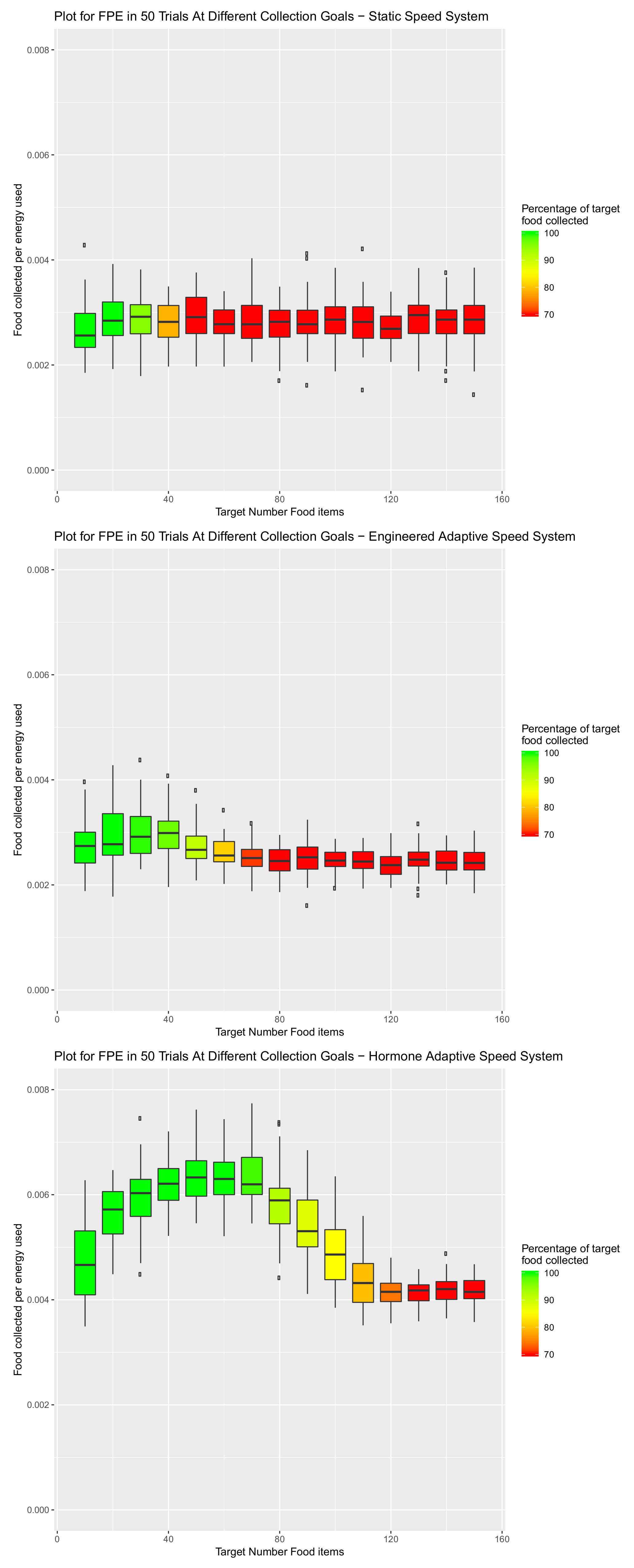
3.5.2. Environment 2—Funnelled Corridor Arena
4. Introduction of the Sleep Hormone to a Foraging Swarm
4.1. Preliminary Tests for Sleep Hormone in A Demand Lead Foraging Task
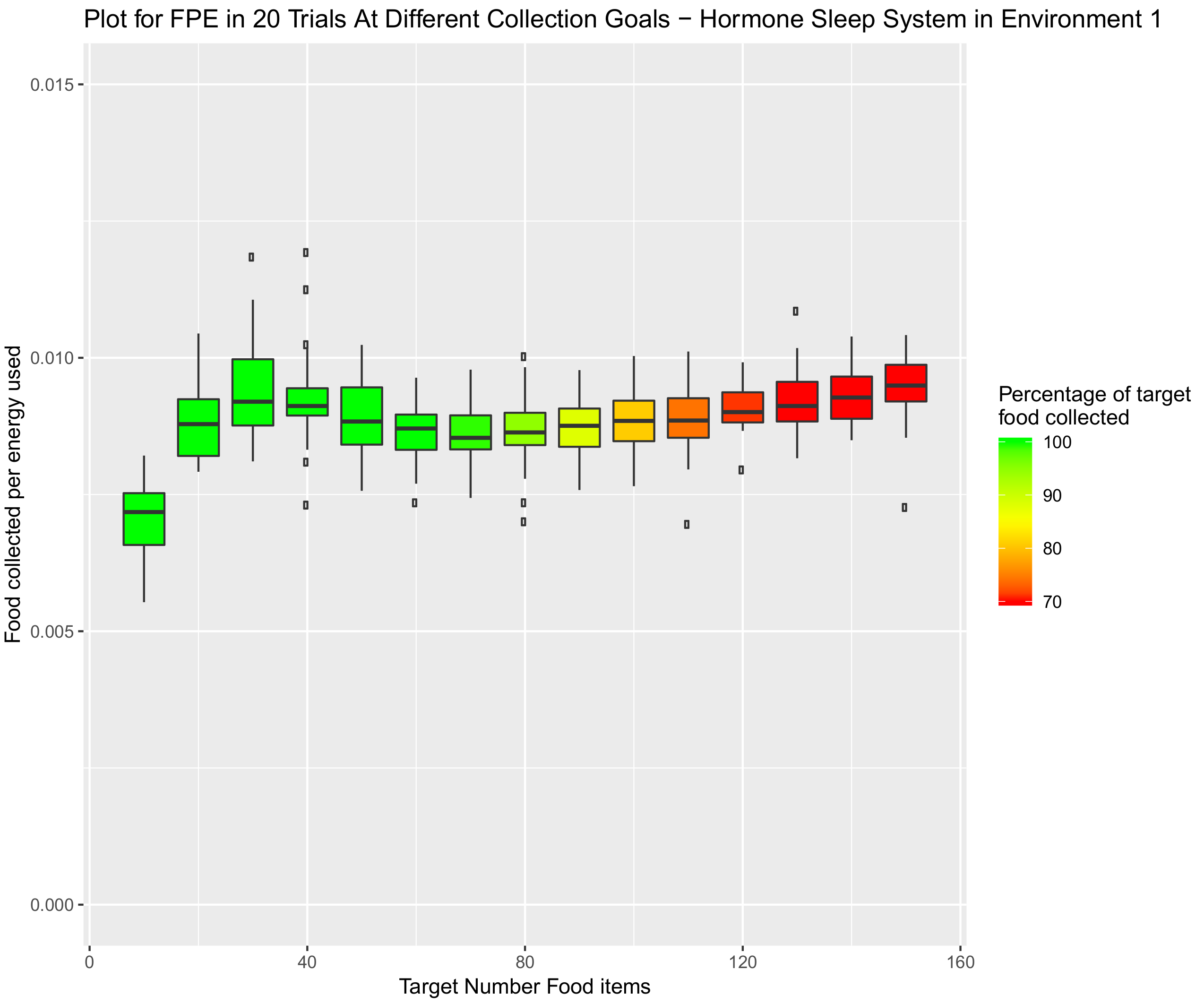
4.1.1. Environment 1—Square Open Arena
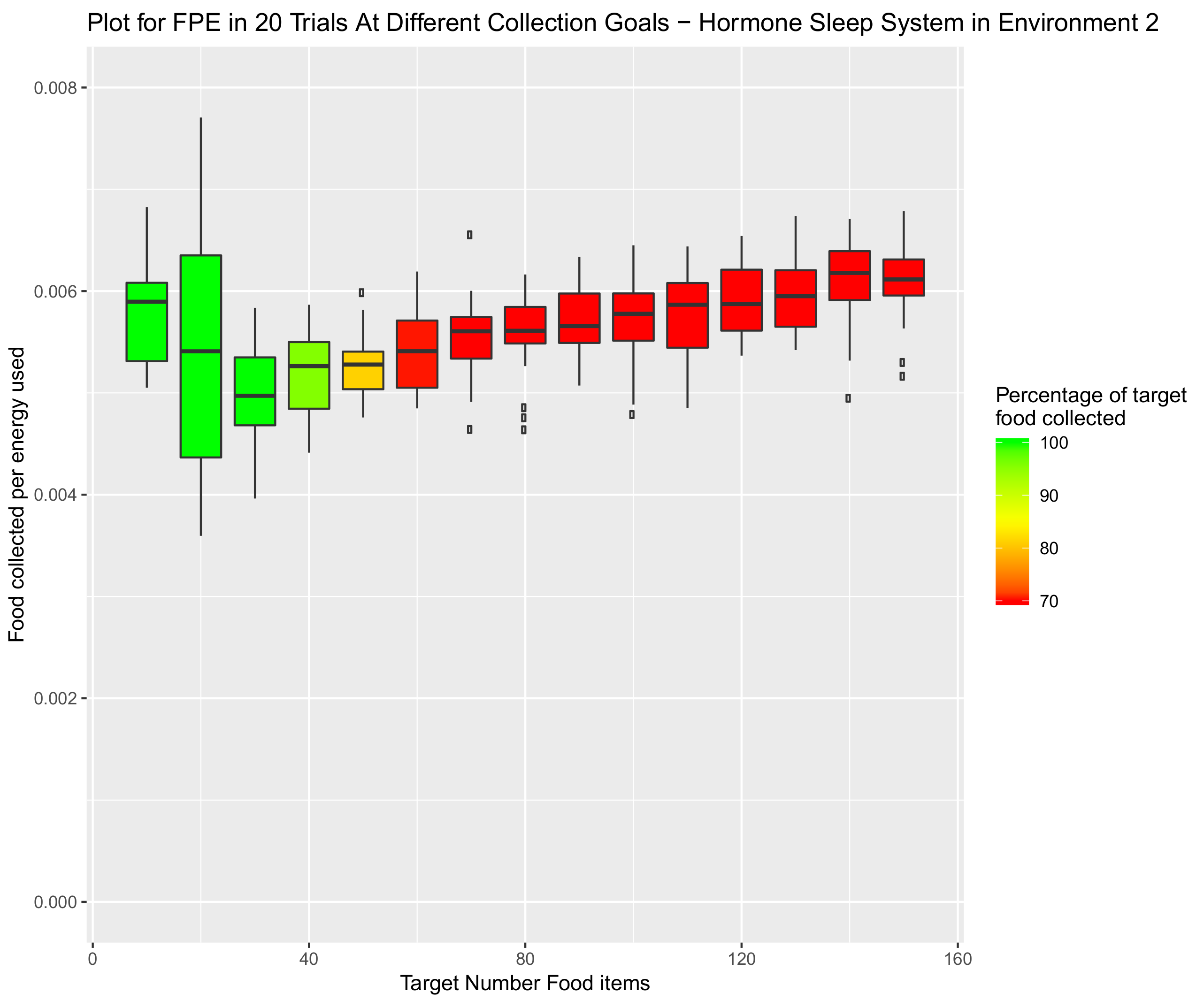
4.1.2. Environment 2—Funnelled Corridor Arena
4.2. Combining the Sleep Hormone with the Speed Deviating System
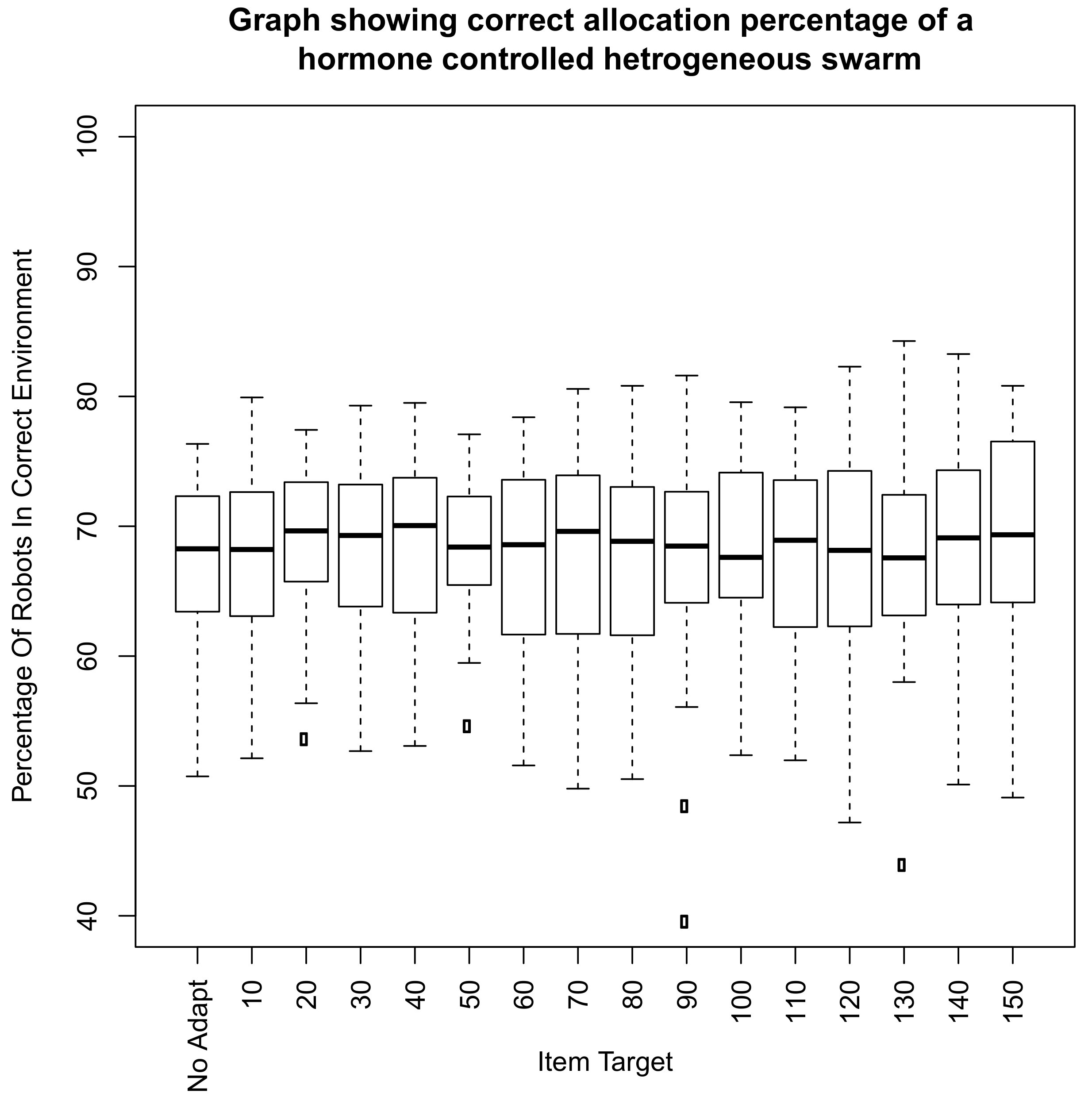
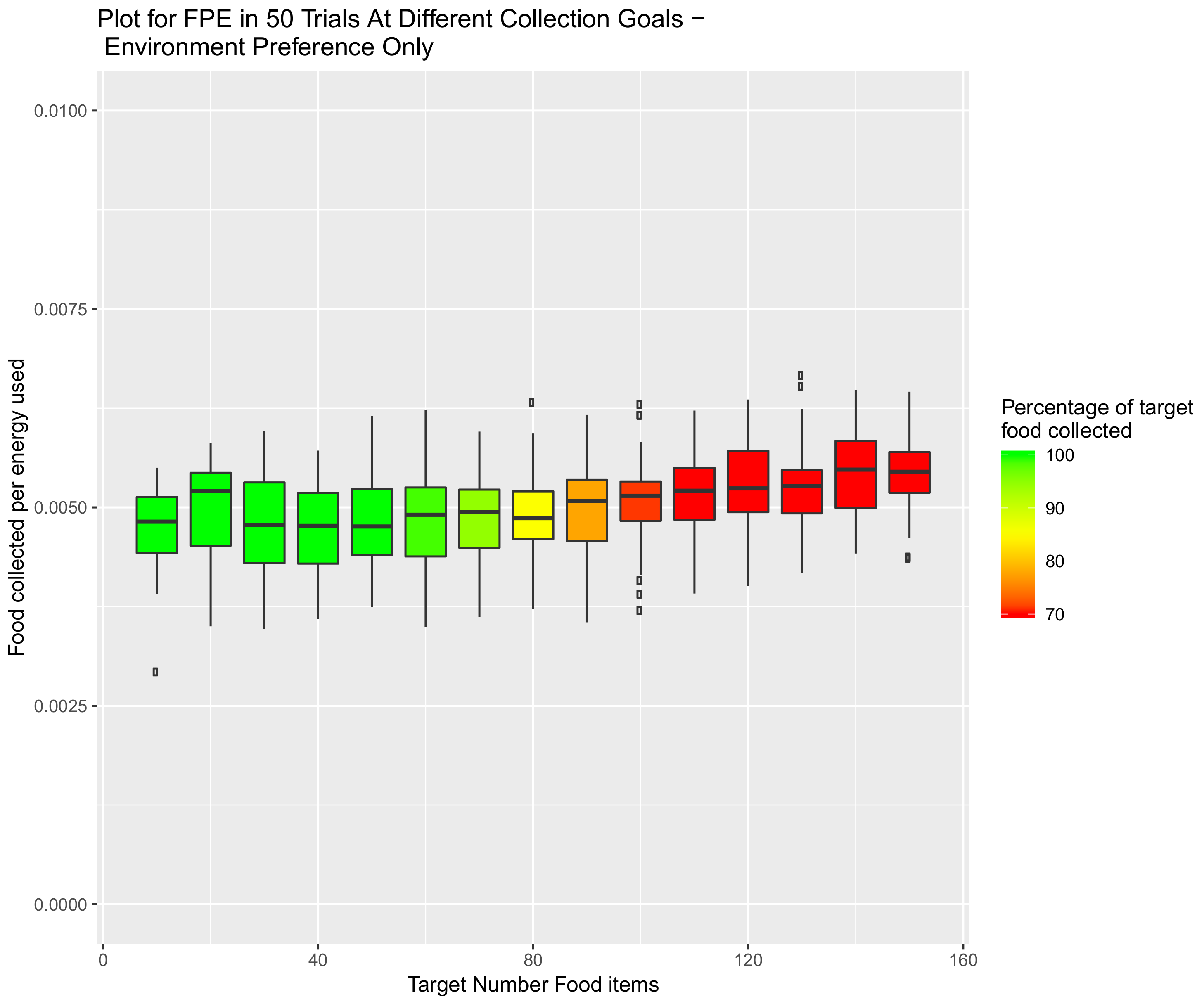
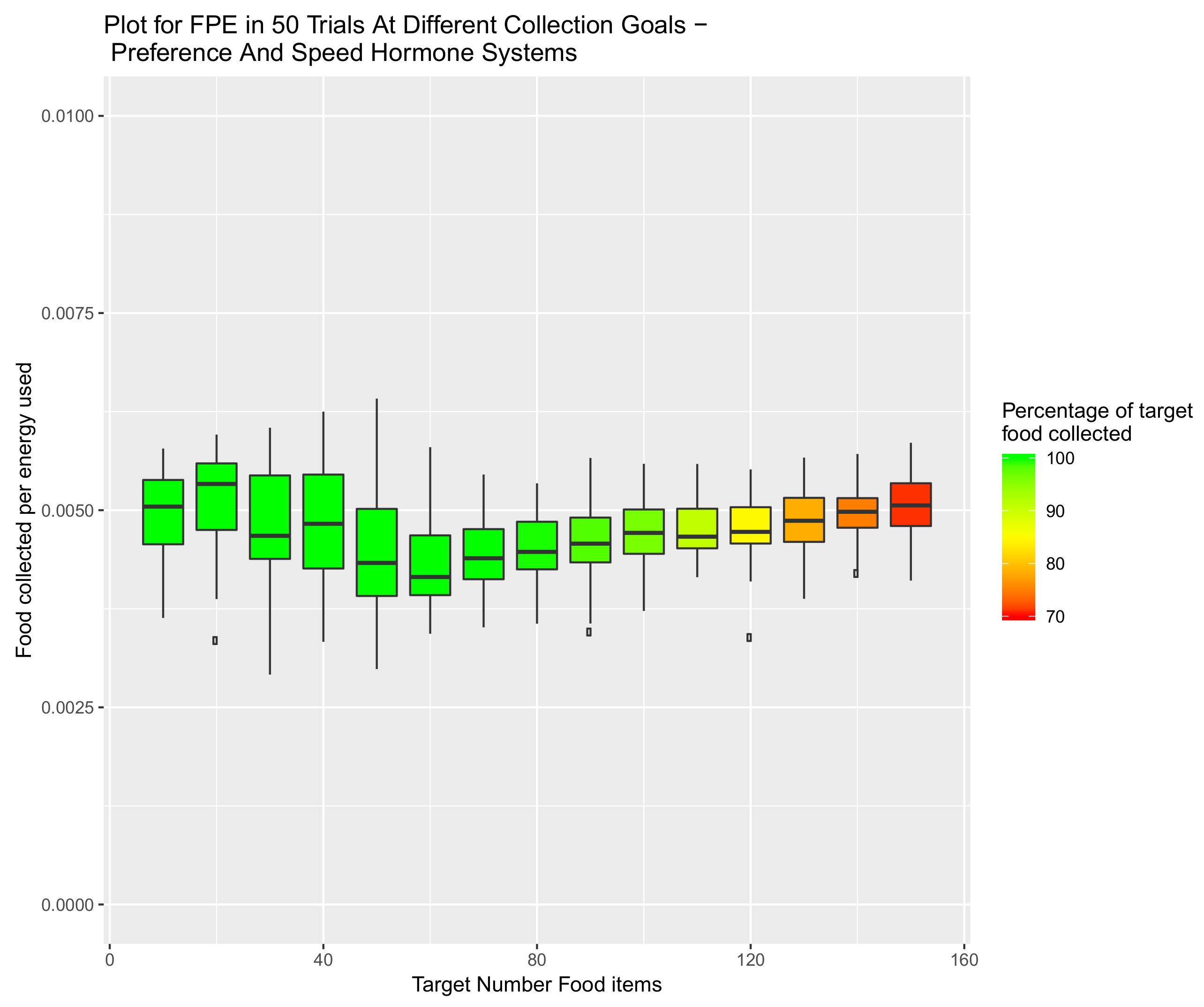
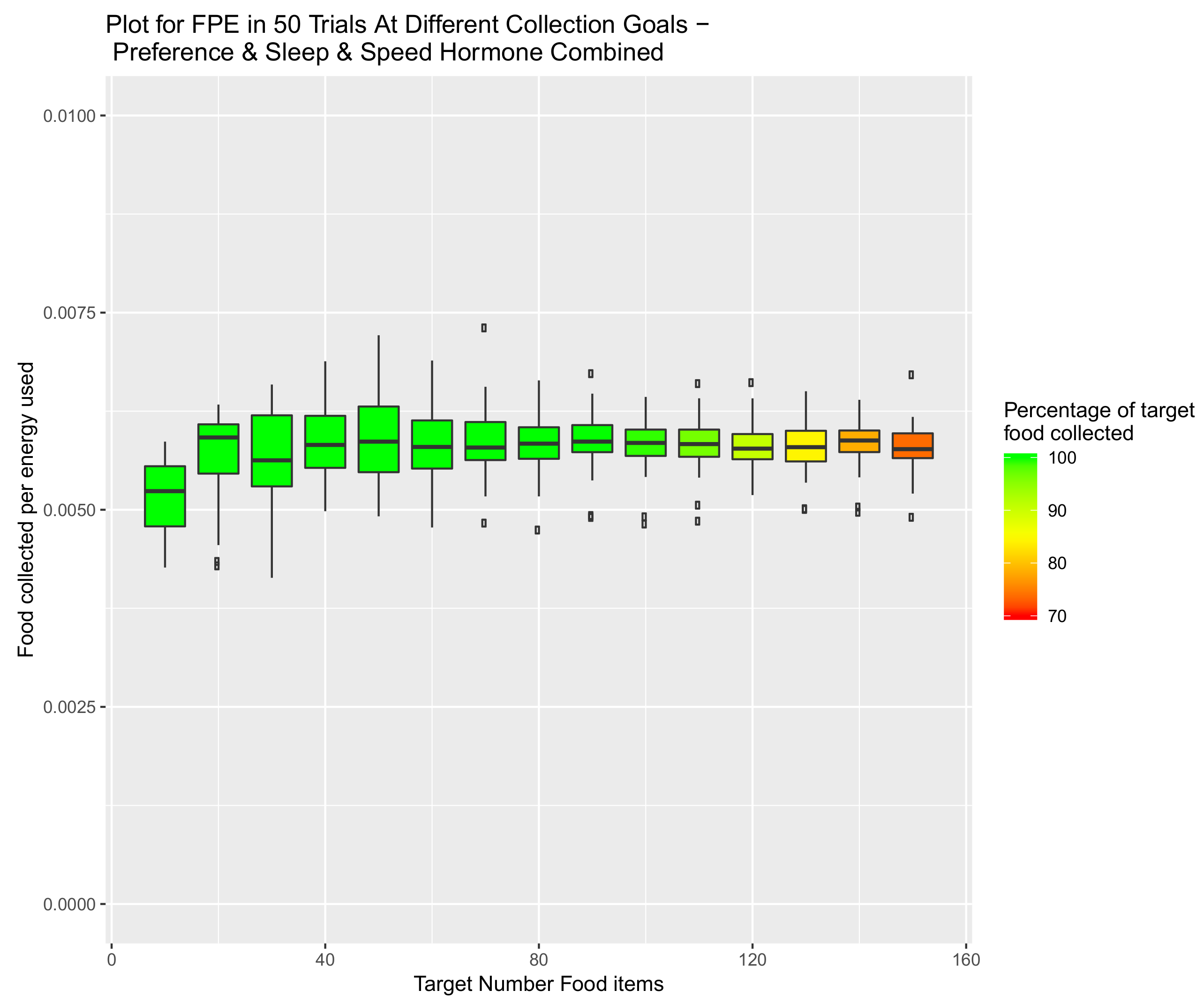
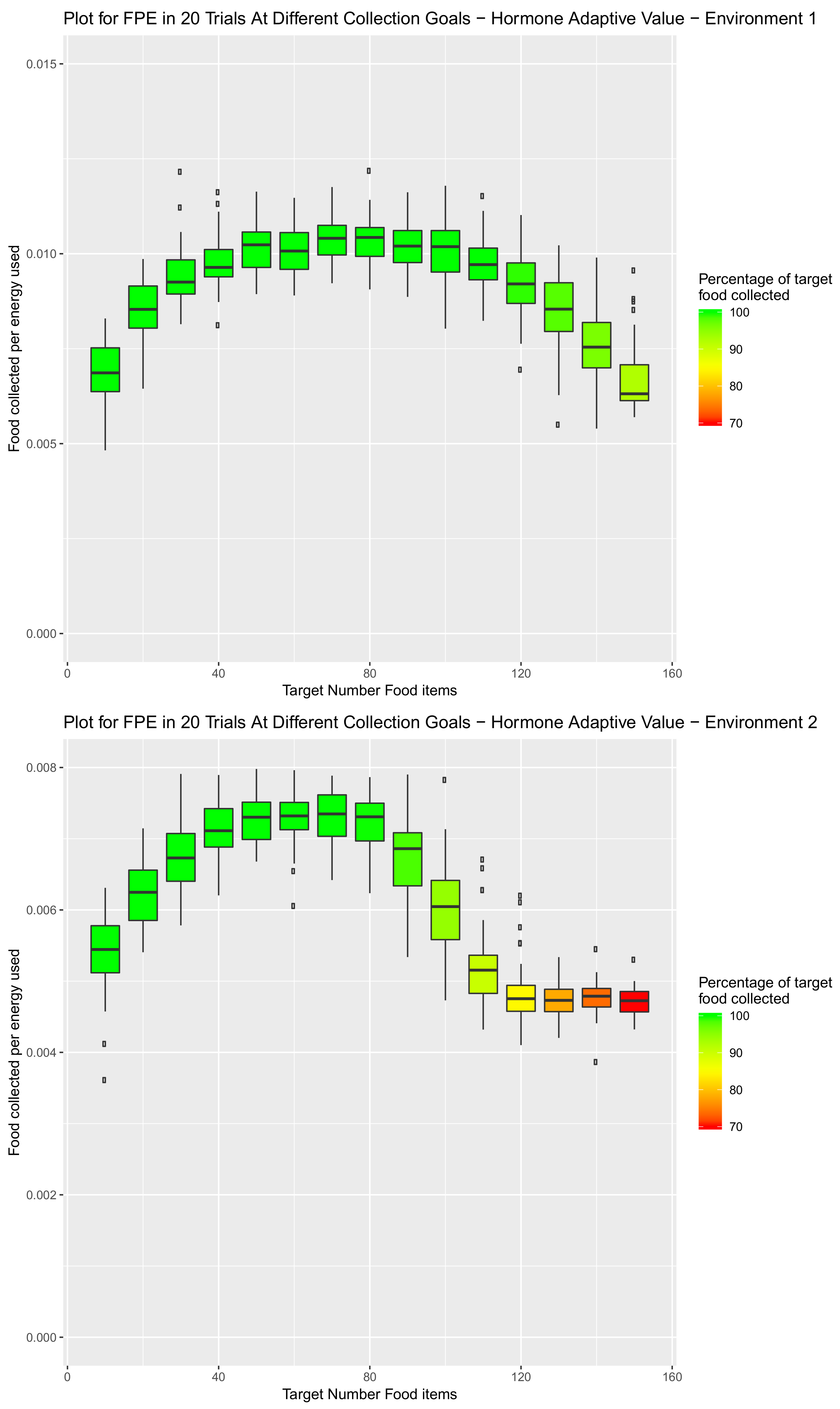
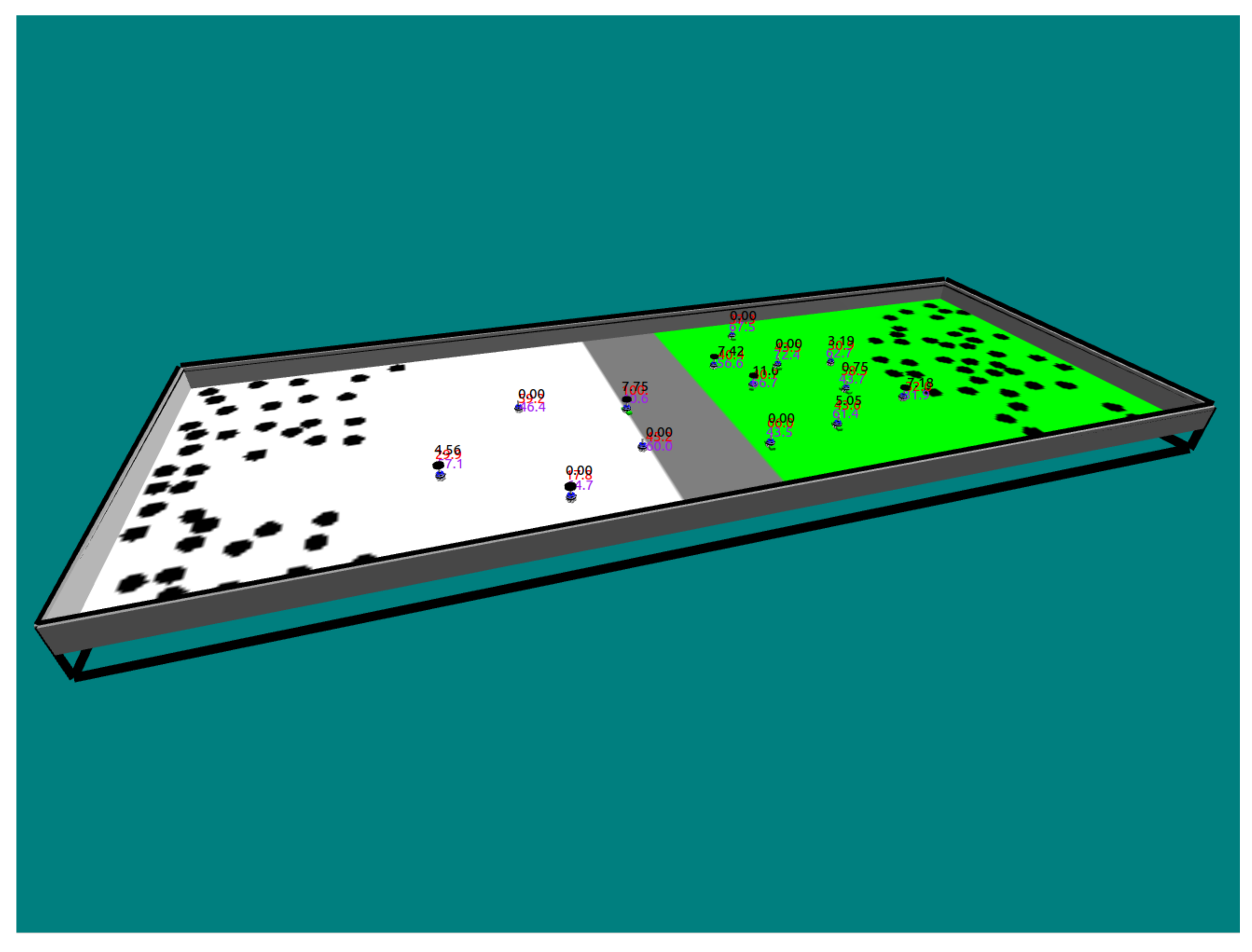
5. Introduction of Environment Selection Hormones with Sleep and Speed Regulating Hormone Systems
5.1. Environmental Setup
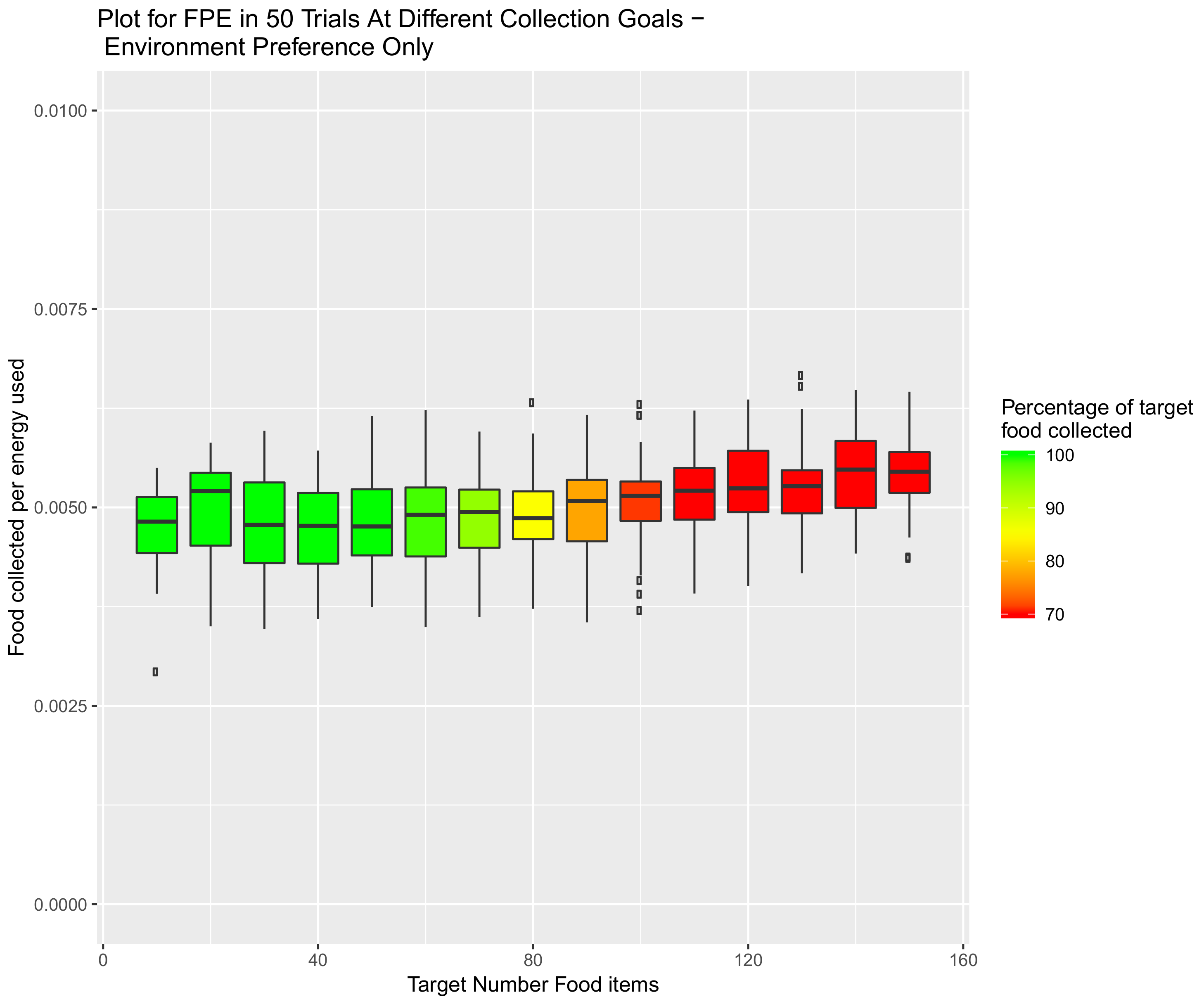
5.2. Effect of Demand on Environment Selection When the Speed Hormone Is Combined with Environmental Preference Hormone
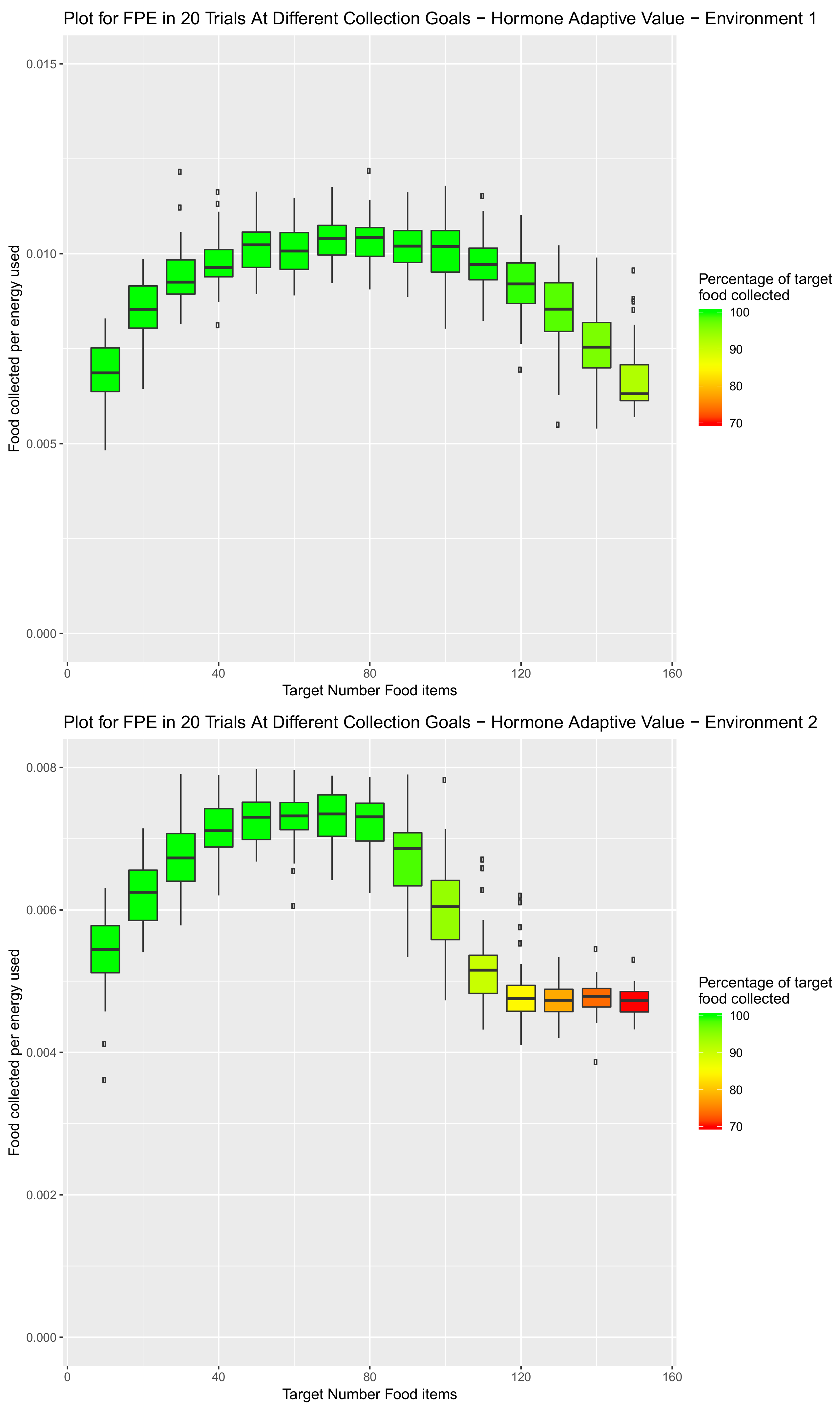
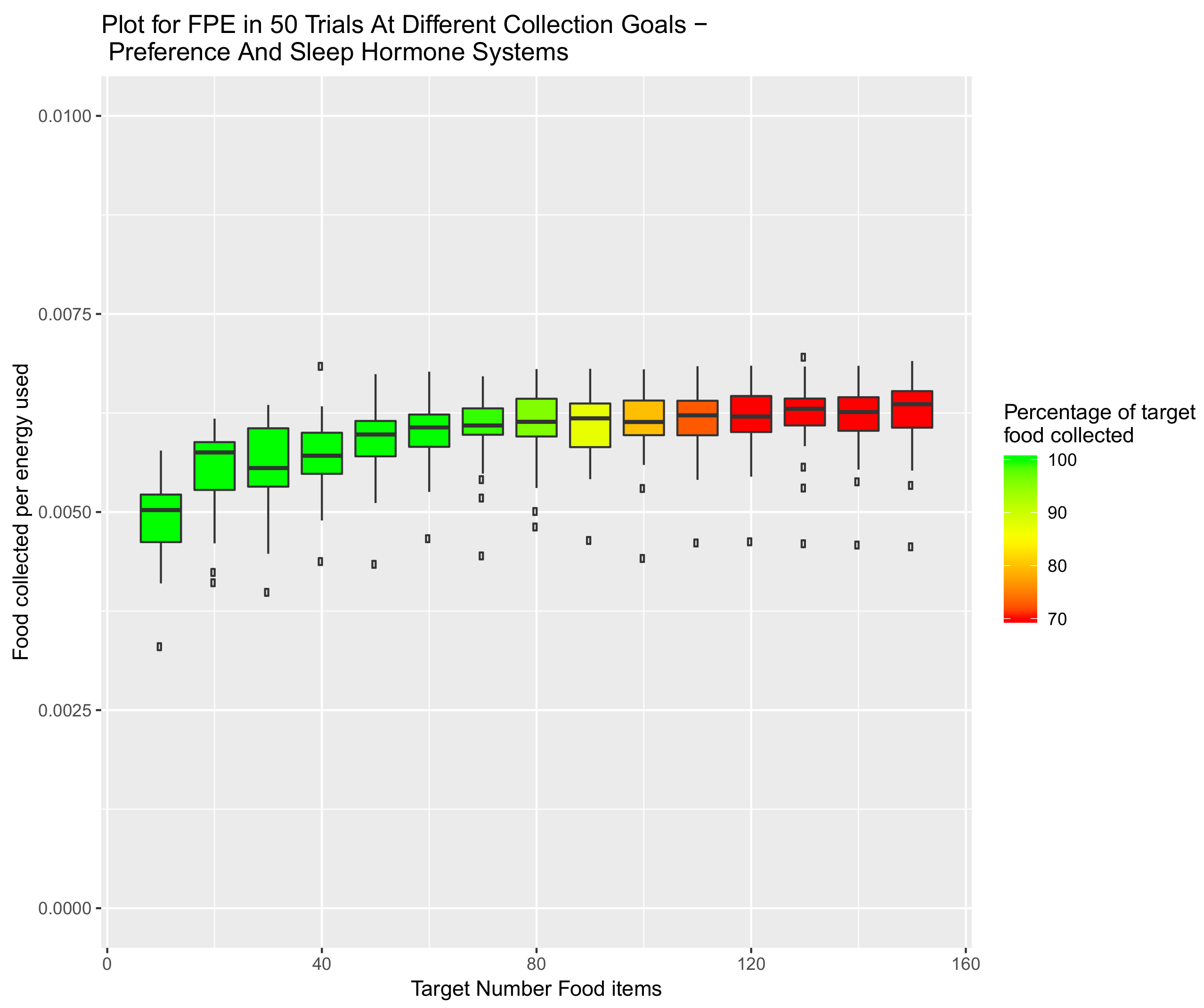
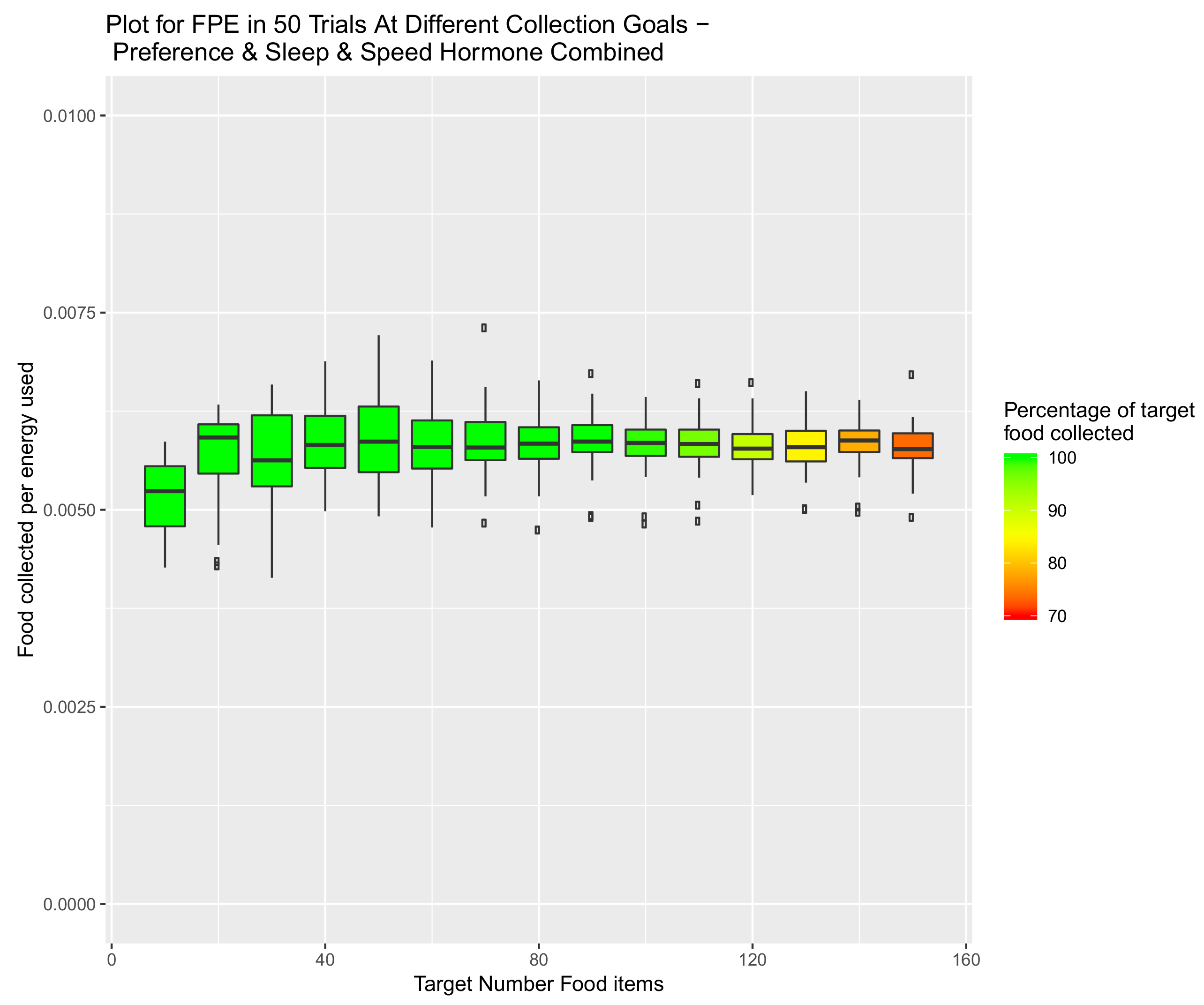
5.3. System Combining Sleep, Speed and Preference Hormones
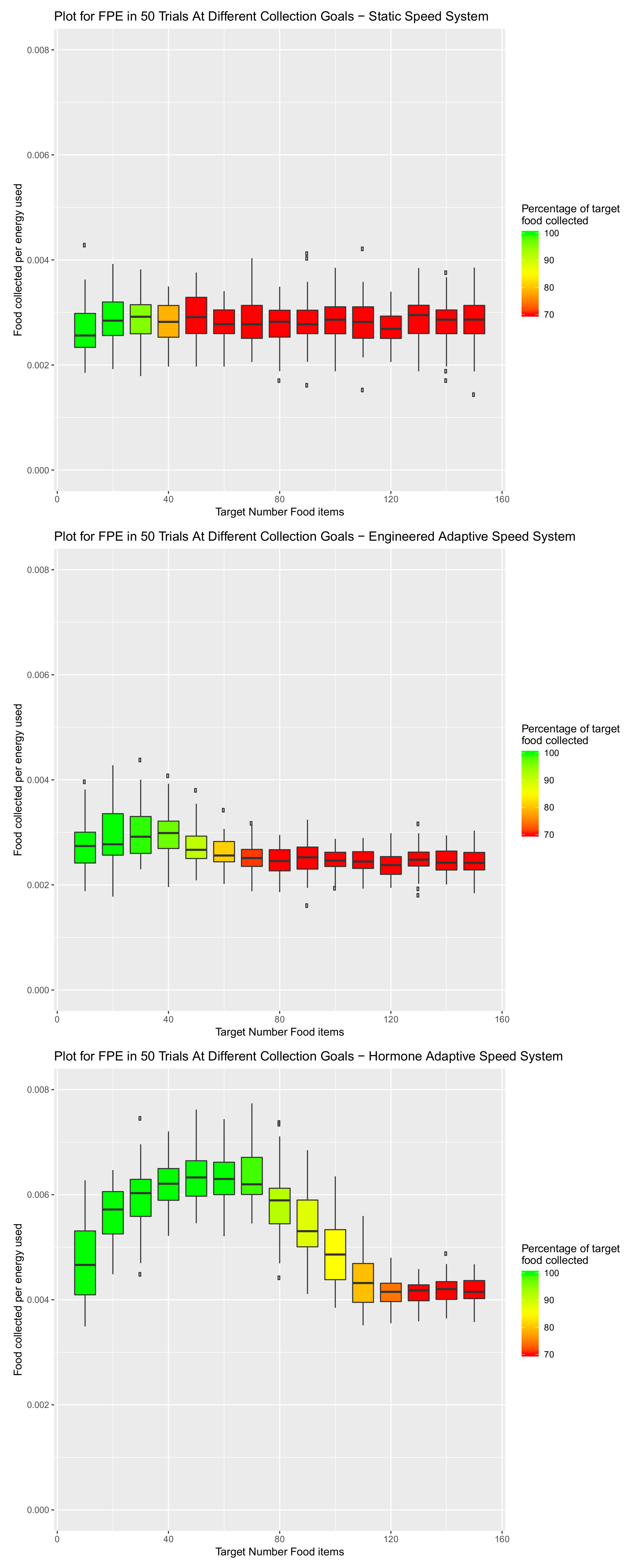
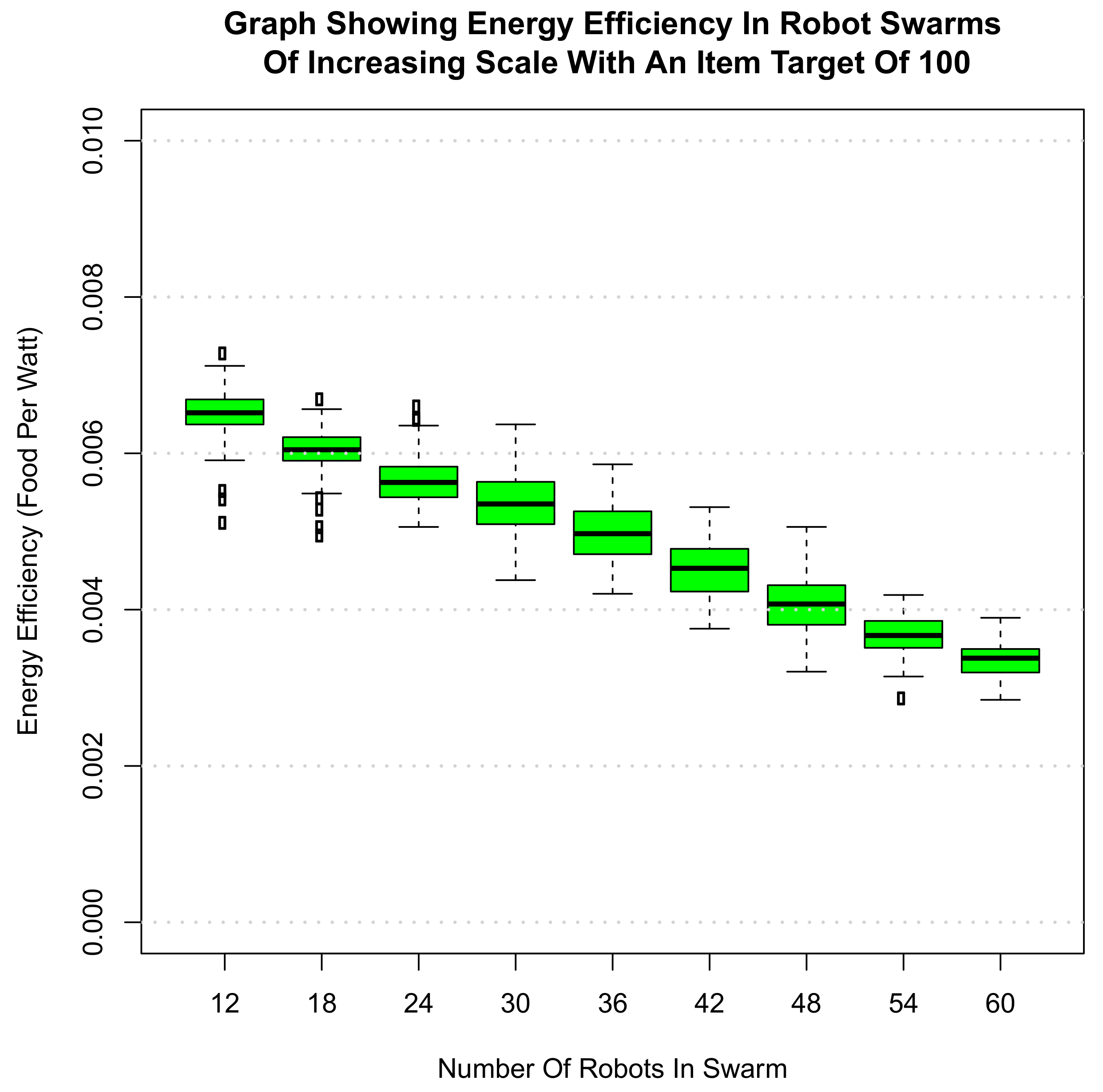
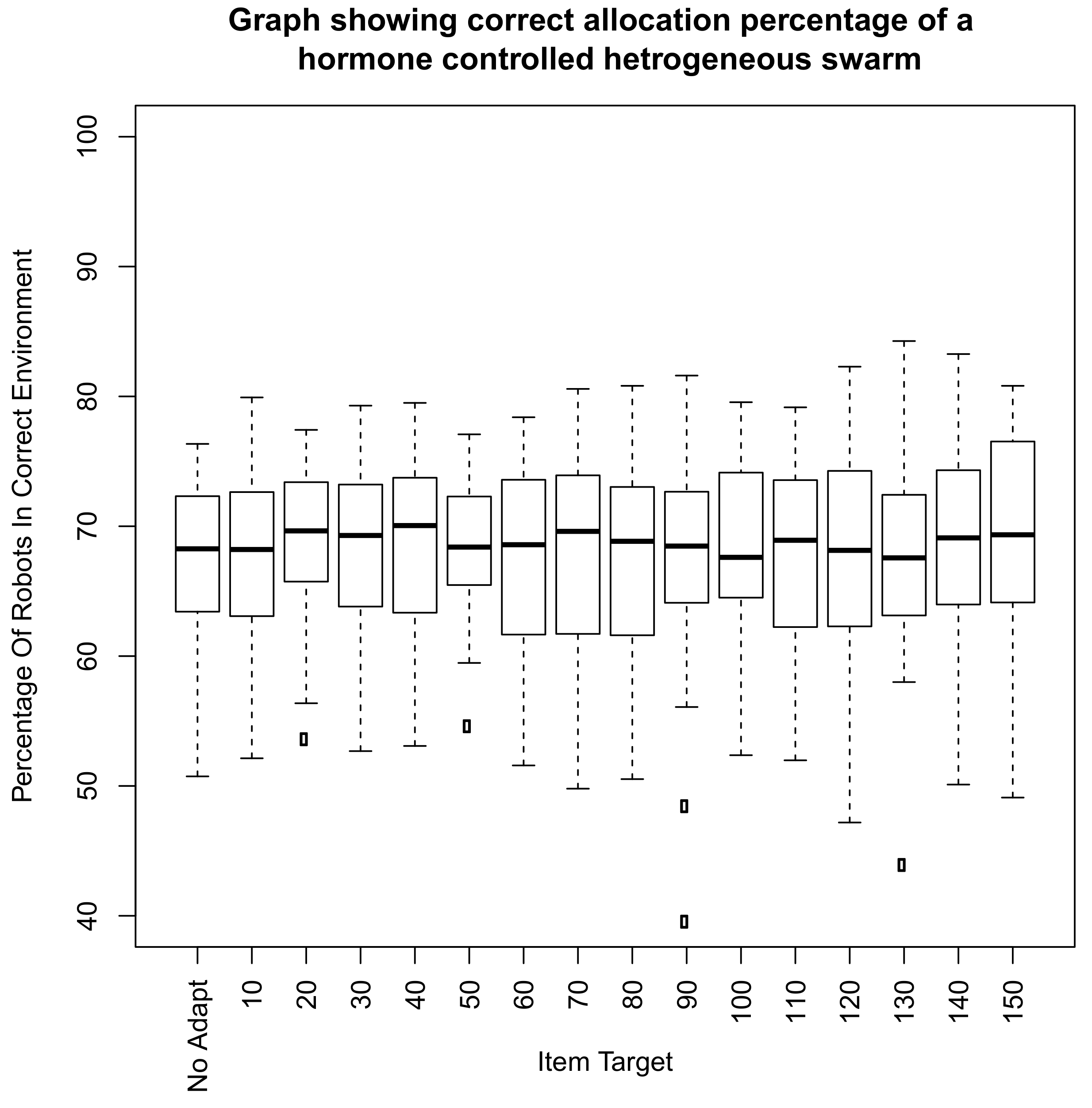
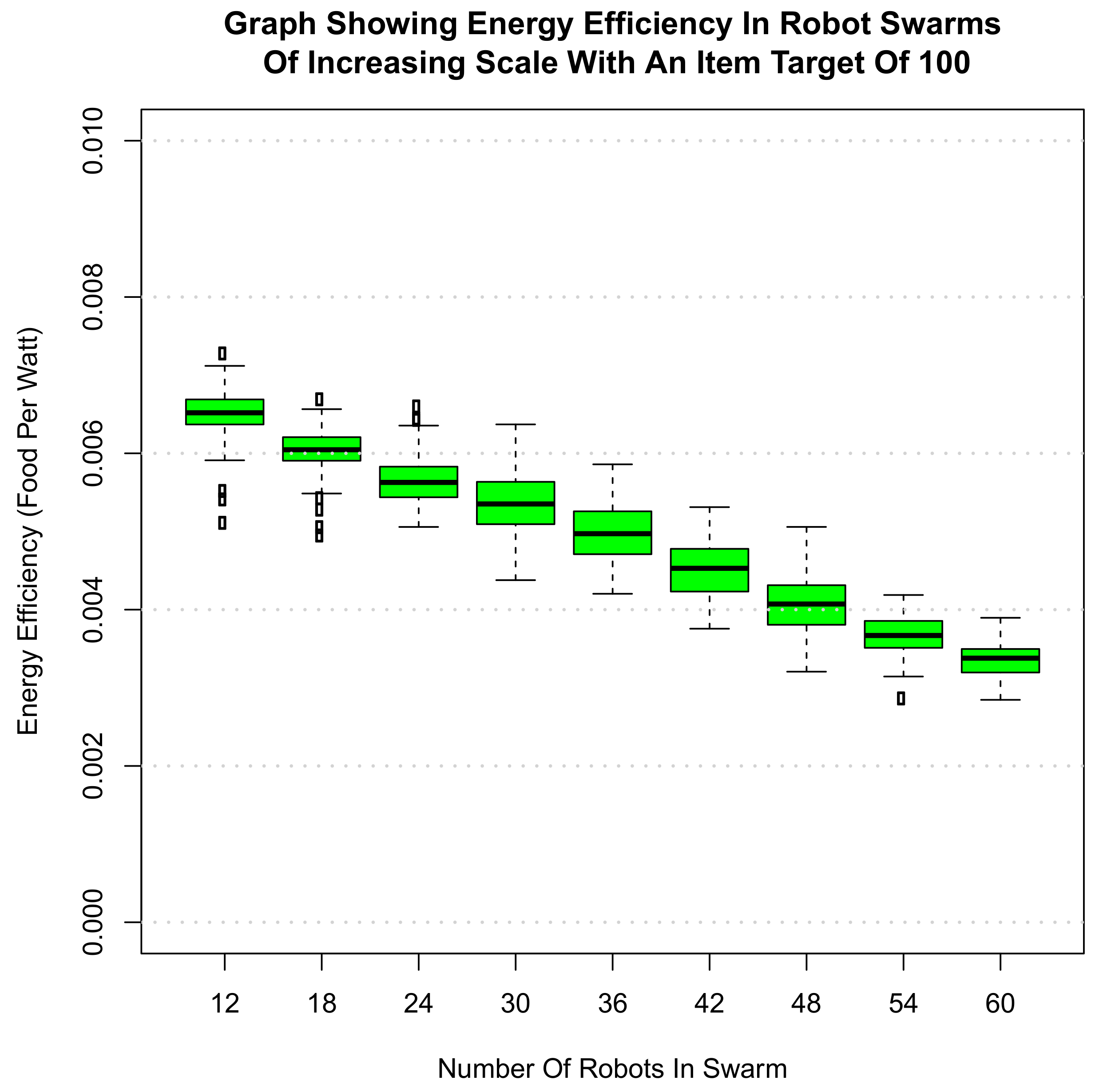
6. Scalability of Final Amalgamated Hormone System
7. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Jaycox, E.R. Behavioral changes in worker honey bees (Apis mellifera L.) after injection with synthetic juvenile hormone (Hymenoptera: Apidae). J. Kansas Entomol. Soc. 1976, 46, 165–170. [Google Scholar]
- Adkins-Regan, E. Hormones and Animal Social Behavior; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Baird, T.A. Lizards and other reptiles as model systems for the study of contest behaviour. In Animal Contests; Cambridge University Press: Cambridge, UK, 2013; pp. 258–286. [Google Scholar]
- Kuyucu, T.; Tanev, I.; Shimohara, K. Hormone-inspired behaviour switching for the control of collective robotic organisms. Robotics 2013, 2, 165–184. [Google Scholar] [CrossRef]
- Wilson, J.; Timmis, J.; Tyrrell, A. A Hormone Arbitration System for Energy Efficient Foraging in Robot Swarms. In Annual Conference Towards Autonomous Robotic Systems; Springer: Berlin, Germany, 2018; pp. 305–316. [Google Scholar]
- Shen, W.; Will, P.; Galstyan, A.; Chuong, C. Hormone-inspired self-organization and distributed control of robotic swarms. Auton. Robots 2004, 17, 93–105. [Google Scholar] [CrossRef]
- Wilson, J.; Timmis, J.; Tyrrell, A. A Hormone-Inspired Arbitration System For Self Identifying Abilities Amongst A Heterogeneous Robot Swarm. In Proceedings of the 2018 IEEE Symposium Series on Computational Intelligence (SSCI), Bangalore, India, 18–21 November 2018; pp. 843–850. [Google Scholar]
- Stradner, J.; Hamann, H.; Schmickl, T.; Crailsheim, K. Analysis and implementation of an artificial homeostatic hormone system: A first case study in robotic hardware. In Proceedings of the 2009 IEEE/RSJ International Conference on Intelligent Robots and Systems, St. Louis, MO, USA, 10–15 October 2009; pp. 595–600. [Google Scholar]
- Kernbach, S.; Meister, E.; Schlachter, F.; Jebens, K.; Szymanski, M.; Liedke, J.; Laneri, D.; Winkler, L.; Schmickl, T.; Thenius, R.; et al. Symbiotic robot organisms: REPLICATOR and SYMBRION projects. In Proceedings of the 8th workshop on performance metrics for intelligent systems, Gaithersburg, MD, USA, 19–21 August 2008; pp. 62–69. [Google Scholar]
- Jin, Y.; Guo, H.; Meng, Y. Robustness analysis and failure recovery of a bio-inspired self-organizing multi-robot system. In Proceedings of the 2009 Third IEEE International Conference on Self-Adaptive and Self-Organizing Systems, San Francisco, CA, USA, 14–18 September 2009; pp. 154–164. [Google Scholar]
- Palmieri, N.; Yang, X.S.; De Rango, F.; Marano, S. Comparison of bio-inspired algorithms applied to the coordination of mobile robots considering the energy consumption. Neural Comput. Appl. 2017, 31, 263–286. [Google Scholar] [CrossRef]
- Liu, W.; Winfield, A.F.T.; Sa, J.; Chen, J.; Dou, L. Towards energy optimization: Emergent task allocation in a swarm of foraging robots. Adapt. Behav. 2007, 15, 289–305. [Google Scholar] [CrossRef]
- De Martinisl, V.; Gallo, M.; D’Acierno, L. Estimating the benefits of energy-efficient train driving strategies: A model calibration with real data. Urban Transp. XIX 2013, 130, 201–211. [Google Scholar]
- Miyatake, M.; Ko, H. Optimization of train speed profile for minimum energy consumption. IEEJ Trans. Electr. Electron. Eng. 2010, 5, 263–269. [Google Scholar] [CrossRef]
- Lee, J.H.; Ahn, C.W. Improving energy efficiency in cooperative foraging swarm robots using behavioral model. In Proceedings of the 2011 IEEE Sixth International Conference on Bio-Inspired Computing: Theories and Applications, Penang, Malaysia, 27–29 September 2011; pp. 39–44. [Google Scholar]
- Pang, B.; Zhang, C.; Song, Y.; Wang, H. Self-organized task allocation in swarm robotics foraging based on dynamical response threshold approach. In Proceedings of the 2017 IEEE 18th International Conference on Advanced Robotics (ICAR), Hong Kong, China, 10–12 July 2017; pp. 256–261. [Google Scholar]
- Hilder, J.; Horsfield, A.; Millard, A.G.; Timmis, J. The Psi Swarm: A Low-Cost Robotics Platform and Its Use in an Education Setting. In Conference Towards Autonomous Robotic Systems; Springer: Berlin, Germany, 2016; pp. 158–164. [Google Scholar]
- Bonani, M.; Longchamp, V.; Magnenat, S.; Rétornaz, P.; Burnier, D.; Roulet, G.; Vaussard, F.; Bleuler, H.; Mondada, F. The marXbot, a miniature mobile robot opening new perspectives for the collective-robotic research. In Proceedings of the 2010 IEEE/RSJ International Conference on Intelligent Robots and Systems, Taipei, Taiwan, 18–22 October 2010; pp. 4187–4193. [Google Scholar]
- Keysight Technologies. N6700 Modular Power System Family; Keysight Technologies: Santa Rosa, CA, USA, 2016. [Google Scholar]
- Buchanan, E.; Pomfret, A.; Timmis, J. Dynamic Task Partitioning for Foraging Robot Swarms. Prodeedings of the International Conference on Swarm Intelligence, Bali, Indonesia, 25–30 June 2016; Springer: Berlin, Germany, 2016; pp. 113–124. [Google Scholar]
- Pinciroli, C.; Trianni, V.; O’Grady, R.; Pini, G.; Brutschy, A.; Brambilla, M.; Mathews, N.; Ferrante, E.; Di Caro, G.; Ducatelle, F.; et al. ARGoS: A Modular, Parallel, Multi-Engine Simulator for Multi-Robot Systems. Swarm Intell. 2012, 6, 271–295. [Google Scholar] [CrossRef]
- Robinson, S. Simulation: The Practice of Model Development and Use; Wiley: Chichester, UK, 2004. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0.9977 | 5 | 0.999 | 9 | 0.01 |
| System Type | Engineered vs. Static | Hormone vs. Static | Hormone vs. Engineered |
|---|---|---|---|
| Item Target Number | |||
| 10 | 0.8550 | 0.0330 | 0.0053 |
| 20 | 0.1648 | p < 0.0001 | p < 0.0001 |
| 30 | 0.1800 | p < 0.0001 | p < 0.0001 |
| 40 | 0.2626 | p < 0.0001 | p < 0.0001 |
| 50 | 0.0906 | p < 0.0001 | p < 0.0001 |
| 60 | 0.8227 | p < 0.0001 | p < 0.0001 |
| 70 | 0.0068 | p < 0.0001 | p < 0.0001 |
| 80 | 0.0262 | p < 0.0001 | p < 0.0001 |
| 90 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 100 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 110 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 120 | p < 0.0001 | 0.0199 | p < 0.0001 |
| 130 | p < 0.0001 | 0.3984 | p < 0.0001 |
| 140 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 150 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| System Type | Engineered vs. Static | Hormone vs. Static | Hormone vs. Engineered |
|---|---|---|---|
| Item Target Number | |||
| 10 | 0.2482 | p < 0.0001 | p < 0.0001 |
| 20 | 0.6918 | p < 0.0001 | p < 0.0001 |
| 30 | 0.3432 | p < 0.0001 | p < 0.0001 |
| 40 | 0.1010 | p < 0.0001 | p < 0.0001 |
| 50 | 0.0817 | p < 0.0001 | p < 0.0001 |
| 60 | 0.0020 | p < 0.0001 | p < 0.0001 |
| 70 | 0.0002 | p < 0.0001 | p < 0.0001 |
| 80 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 90 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 100 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 110 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 120 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 130 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 140 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 150 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 0.01 | 0.999 | 0.01 | 0.06 | 0.0.015 | 0.999 | 10 |
| Item Target Number | Hormone Speed vs. Hormone Combination | Hormone Sleep vs. Hormone Combination | ||
|---|---|---|---|---|
| (Environment 1) | (Environment 2) | (Environment 1) | (Environment 2) | |
| 10 | 0.9680 | 0.0047 | 0.2315 | 0.019 |
| 20 | 0.8830 | 0.0040 | 0.1653 | 0.0056 |
| 30 | 0.5290 | p < 0.0001 | 0.5831 | p < 0.0001 |
| 40 | 0.6017 | p < 0.0001 | 0.0810 | p < 0.0001 |
| 50 | 0.5290 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 60 | 0.0809 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 70 | 0.0283 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 80 | 0.0047 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 90 | 0.0675 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 100 | 0.0024 | p < 0.0001 | p < 0.0001 | 0.0430 |
| 110 | 0.0910 | p < 0.0001 | 0.0024 | 0.0002 |
| 120 | 0.0763 | p < 0.0001 | 0.9042 | p < 0.0001 |
| 130 | 0.1081 | p < 0.0001 | 0.0211 | p < 0.0001 |
| 140 | 0.0227 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| 150 | 0.2648 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| Terrain Type | Wood Suited Wheels | Grass Suited Wheels |
|---|---|---|
| Grass | 0.6 | 0.7 |
| Wood | 1 | 0.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, J.; Timmis, J.; Tyrrell, A. An Amalgamation of Hormone Inspired Arbitration Systems for Application in Robot Swarms. Appl. Sci. 2019, 9, 3524. https://doi.org/10.3390/app9173524
Wilson J, Timmis J, Tyrrell A. An Amalgamation of Hormone Inspired Arbitration Systems for Application in Robot Swarms. Applied Sciences. 2019; 9(17):3524. https://doi.org/10.3390/app9173524
Chicago/Turabian StyleWilson, James, Jon Timmis, and Andy Tyrrell. 2019. "An Amalgamation of Hormone Inspired Arbitration Systems for Application in Robot Swarms" Applied Sciences 9, no. 17: 3524. https://doi.org/10.3390/app9173524
APA StyleWilson, J., Timmis, J., & Tyrrell, A. (2019). An Amalgamation of Hormone Inspired Arbitration Systems for Application in Robot Swarms. Applied Sciences, 9(17), 3524. https://doi.org/10.3390/app9173524