Abstract
Traditionally regarded as single cell organisms, bacteria naturally and preferentially build multicellular communities that enable them to react efficiently to external stimuli in a coordinated fashion and with extremely effective outcomes. These communities are bacterial biofilms, where single cells or microcolonies are embedded in self-built Extracellular Polymeric Substance (EPS), composed of different macromolecules, e.g., polysaccharides, proteins, lipids, and extracellular DNA (eDNA). Despite being the most common form in nature and having many biotechnologically useful applications, biofilm is often regarded as a life-threatening form of bacterial infection. Since this form of bacterial life is intrinsically more resistant to antibiotic treatment and antimicrobial resistance is reaching alarming levels, we will focus our attention on how nanotechnology made new tools available to the medical community for the prevention and treatment of these infections. After a brief excursus on biofilm formation and its main characteristics, different types of nanomaterials developed to prevent or counteract these multicellular forms of bacterial infection will be described. A comparison of different classifications adopted for nanodrugs and a final discussion of challenges and future perspectives are also presented.
1. Introduction
The bell has tolled. Antimicrobial resistance is set to cause 30 million deaths by 2050 and will continue to cause a huge economic impact on health-care providers, around 100 Trillion USD$ [1]. The World Health Organization (WHO) has also recently outlined and ranked the microbes that should be highly prioritized for the development of new antibiotics [2]. These two documents highlight the extreme necessity of new therapies for microbial infection, claiming for research on the development of new antibiotics. Since the golden age of antibiotics, between 1940 and 1960, many different classes of molecules were discovered and introduced in clinical practice [3]. Right after introduction into clinics, specific antibiotic-resistant strains began to appear. From the 1970s, the development of new classes of antibiotics, i.e., completely different molecules compared to the ones already marketed, started to slow down, resulting in very few new antibacterial drugs introduced into practice. The last new antibiotic active against Gram-negative bacteria, quinolones, appeared in 1962. More recent is the availability of two new classes of drugs against Gram-positive bacteria. Lipopeptides and oxazolidinones were approved in 2000 and 2003, respectively [4]. Furthermore, the aforementioned rapid appearance of resistant strains terribly complicates the landscape. The generation of a new class of molecules requires at least a decade of work, whereas the onset of resistant isolates may take as fast as one or two years after introduction into clinics [5].
The problem of bacterial morbidity is especially prominent in implant-related or indwelling device-related infections, where bacteria stably colonize different types of implants, catheters or prosthetic valves. The colonization of these devices is mainly caused by the establishment of a biofilm, a bacterial community embedded in a self-produced Extracellular Polymeric Substance (EPS) that enables the microorganisms to settle at the site of infection and efficiently cope with various external potentially deleterious stresses [6]. This milieu provides ideal conditions for bacteria to withstand antibiotic treatment, decreasing antibiotic efficacy due to lower penetration into deeper layers of the biofilm and the metabolic state of bacterial cells that is predominantly in the stationary phase. It is known that many antibiotics require an active bacterial proliferation, i.e., in the logarithmic phase. This condition is different from genetic resistance and is better referred to as tolerance [7] and it further intricates the landscape of bacterial infections.
If examined together, these data highlight the need to complement the antimicrobial field with different preventive and therapeutic strategies that must be regarded with mounting interest. Indeed, the field of nanomedicine has emerged in the last decades as a promising option for new diagnostic and therapeutic alternatives in the clinic. Most clinical applications of nanotechnological discoveries involve non-infectious diseases, addressing mainly cancer [8]. Other fields of application include central nervous system diseases, cardiovascular and inflammatory pathologies. In addition, diagnostic and theranostic applications are also valued fields of investigation. It has to be mentioned that most of the research in this area is at a pre-clinical stage or undergoing clinical trials [8,9]. The translation from bench-to-bed occurred mainly for cancer treatment. Since 1995, when Doxil®, a liposomal-encapsulated form of doxorubicin to reduce its cardiotoxicity [10], earned the approval from the Federal Drug Administration (FDA), many other nano-drugs reached the clinics. The only example regarding infectious diseases is represented by AmBisome, a liposomal form of amphotericin B to treat fungal infections [11].
Therefore, the aim of this review is to summarize the state of the art regarding nanotechnological research in order to counteract biofilm communities. Most literature addresses bacterial infections analyzing planktonic populations, not considering that these multicellular communities mainly mediate an established and recurrent infection. To better understand biofilm biology, we will first focus on its formation and its characteristics. Then, nanothechnological advances will be illustrated analyzing both different classifications proposed in the literature and their applications towards biofilm infections. Finally, a brief excursus on the difficulties of the clinical translation process will be discussed.
2. Biofilm Formation
As previously mentioned, bacteria are traditionally considered unicellular organisms. Initial observations that led to the formulation of the hypothesis of the formation of sessile communities that are able to stick to surfaces date back to the 1970s thanks to the contributions of Marshall and Costerton [12,13,14]. Over the last forty years, progress has been made in the understanding of this form of bacterial association, understanding many different mechanisms related to biofilm biology, excellently reviewed recently [15,16].
Concerning formation, schematized in Figure 1, the first step is adherence to a surface. It can be mediated both by non-specific factors, like physico-chemical factors, as well as by specific bacterial protein components. The former mainly involves on one side the nature of the bacterial cell-wall and superficial proteins and on the other the hydrophobicity of the surface, as analyzed in different bacterial species, like Staphylococcus aureus (Gram-positive) [17] and Pseudomonas aeruginosa (Gram-negative) [18,19]. In these cases, interaction of cell wall components, surfaces proteins and cellular protrusions have been reported to play a role in the attachment to a surface. However, the contribution of single components of the bacterial surface is still debatable, especially due to the methods of analysis and their relevance in in vivo conditions [15]. Another relevant physical factor recognized to be important for biofilm formation is shear stress that allows the formation of a much stronger biofilm when it is applied during formation of the community and influences the dispersal process, increasing the chance to disseminate the infection. Many works established the influence of this physical factor on biofilm [20,21,22,23], including also natural biofilm in rivers in which a particular form of biofilm microcolonies, termed streamers, are aligned to the flow with a sessile “head” and a flowing unattached “tail” [24]. Recently, the molecular basis of mechanosensing of shear was clarified in P. aeruginosa, further underlining the relevance of this physical factor [25]. Regarding in vivo conditions, upon interaction with body fluids, any surface becomes covered with ExtraCellular Matrix (ECM) components, thus coating the surface. The case of S. aureus Cell Wall Anchored (CWA) proteins details the specific adhesion to defined ECM molecules. Fibrinogen [26,27], fibronectin [28,29] and collagen [30,31] are the ligands of specific staphylococcal proteins that have been extensively studied and recently re-classified according to their structure [32]. Even specific biofilm-associated proteins have been reported [33]. These proteins have been substantially shown to bind to their respective ligand and thus promoting specific adherence to a coated surface. Similar cases are reported for Gram-negative bacteria [34]. These specific interactions provide a more realistic base for the first step of biofilm formation in medically relevant conditions.
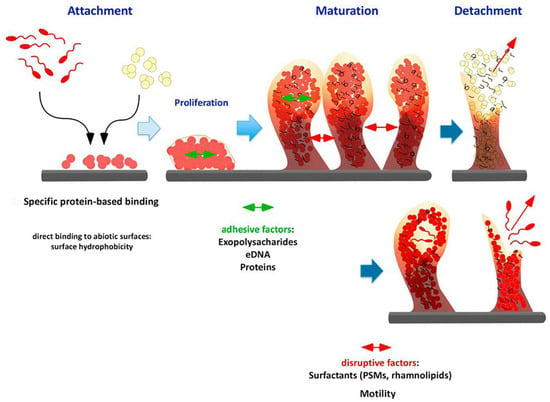
Figure 1.
Biofilm formation stages. Phenol-Soluble Modulins (PMSs) (Modified with permission, from [15]).
The second step is the maturation of the biofilm where increase in biomass and the production of the Extracellular Polymeric Substance (EPS) occur to achieve a complex 3D structure. The main component of EPS is water (up to 97%) [16] followed by different biological macromolecules as polysaccharides, proteins and extracellular DNA (eDNA). Specific types of polysaccharides are produced when bacteria are embedded within biofilm: poly-N-acetyl-glucosamine (PNAG or PIA—Polysaccharide Intercellular Adhesin) for staphylococcal species mainly [35,36] and alginate together with Psl, a mannose-rich polysaccharide from gene cluster psl (polysaccharide locus) for P. aeruginosa [37]. The expression of these polysaccharides is strictly regulated by operons and regulons that are specifically activated at precise moments of biofilm growth. Another macromolecular component of EPS are proteins, whose role inside the biofilm is to promote adherence between cell and surface but also cell-cell adhesion and communication. This double role is again best exemplified by staphylococcal CWA proteins [32,38,39]. Furthermore, extracellular enzymes are present to degrade substances sorbed by the biofilm [16,40,41], in some cases reported to be within extracellular vesicles [42], and proteinaceous component derived from died cells cellular debris has also been detected [43]. These enzymes provide many insights into the “social” behavior of biofilms since the digestion of complex molecules present in the matrix allows both remodeling of the EPS and increases the availability of nutrients [16,44]. The last main component of the EPS is eDNA, which revealed itself to be a fundamental component of the biofilm, showing to be necessary upon its formation and maintenance, firstly reported in P. aeruginosa [45]. The role of eDNA is also relevant in staphylococci but not equally in all species. As an example, its role in S. aureus biofilms is fundamental [46], much less in S. epidermidis [47].
The final stage of the biofilm life is the detachment or dispersal, where small biofilm clumps or single cells are released to promote colonization of other sites. This fate of biofilm is triggered by a plethora of stimuli and finely regulated with precise gene expression patterns, extensively reviewed elsewhere [48]. Examples can be drawn from P. aeruginosa where the motility is activated and the productions of surfactants (rhamnolipids) occurs. Phenol-Soluble Modulins (PSM) are surfactants produced by staphylococci to achieve EPS disruption and consequent dispersal [15].
3. Biofilm Properties
The highly heterogeneous nature of EPS provides bacterial communities with many characteristics that influence their behavior and allows the formation of a peculiar microenvironment in which they can better survive and cooperate, as shown in Figure 2.
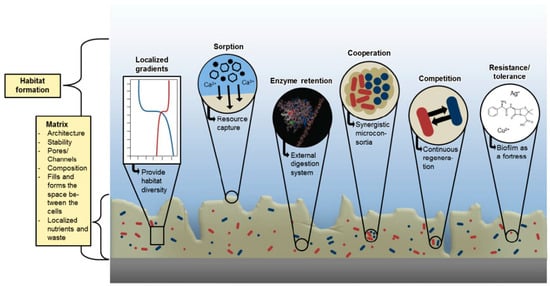
Figure 2.
Biofilm microenviromental features ( With permission, from [14]).
The presence of an extracellular matrix is first a physical barrier to external stresses. As previously said, the main component of EPS is water retained by the hydrophilic molecules of the matrix, thus allowing survival also upon desiccation. In some instances, one further layer of dehydration prevention is provided by the formation of an external skin that forms a physical barrier to water evaporation [49]. Furthermore, the “sticky” nature of the matrix allows non-selective sorption of substances, that may be nutrients, ions or also toxic compounds, as documented in environmental samples of biofilms [50,51,52]. Retained compounds can be digested by enzymes secreted by bacteria embedded into the biofilm as described above, thus rendering nutrients available for the entire community. Further nutrients are provided by died bacterial cells that can be “cannibalized” as documented in Bacillus subtilis biofilms [53]. These sorption and retention capacities allows the growth of biofilm in environments that can be highly oligotrophic, which is typical of most natural environments.
Probably the most medically relevant feature that is connected to biofilm is its intrinsic tolerance that shows to any treatment. Traditionally, planktonic bacteria become resistant to a determined antimicrobial agent thanks to genetic elements that allow them to escape the mechanism blocked by the antimicrobial. By contrast, it is known that bacterial cells within biofilm are not genetically resistant to antimicrobials but always show a high survival rate upon treatment [54]. This phenomenon can be referred to as tolerance (according to Flemming and coauthors [16]). It has been shown that silver nanoparticles (AgNPs) inhibited to a lower extent biofilm viability compared to the planktonic counterpart. Once suspended, biofilm-embedded Pseudomonas aeruginosa cells retained their sensitivity to AgNPs, comparable to the planktonic culture [55]. Thus, an explanation of this effect may be the physical barrier formed by EPS, which can chelate and modify antimicrobial molecules, owing to the presence of secreted enzymes mentioned above. Another reason of the intrinsic tolerance of bacterial cells once embedded in EPS matrix is their metabolic state. EPS causes the formation of different gradients inside the biofilm community (nutrient, oxygen, pH) [14,16] and this leads bacteria to respond differently based on the condition they find. Low nutrients and oxygen paucity may lead to a dormant state, in which these so-called persister cells are still viable but not actively proliferating, being therefore non-susceptible to most antimicrobial treatments [56,57]. This way, at least two mechanisms contribute to biofilm survival upon the presence of antimicrobials.
Furthermore, the close proximity in which bacteria are held thanks to the biofilm matrix enables a “social” behavior of the community. This phenomenon can be explained in multiple ways: the primary way of communication inside the biofilm is thought to be quorum sensing (QS). This communication system is based on chemical signals that are secreted by bacterial population. The concentration of the signal increases as the population grows, reaching the quorum that indicates that the population is conspicuous enough to trigger changes in gene expression patterns [58,59]. QS is studied through the use of batch cultures in the laboratory, allowing high concentration of bacterial cells in a relatively small volume. By contrast, the role of QS in nature is questionable since bacteria rarely reach such high concentrations in close proximity to allow diffusion and sensing of the chemical signals. Instead, within biofilm matrix there is clearly the possibility to increase the local concentration of signal molecules. Therefore, the role of QS has been extensively investigated in biofilm, also as a therapeutic target. Nevertheless, even if in some studies it is deemed relevant for biofilm infection, in other cases it is clear that bacteria that established the biofilm are QS-mutants. Joo and Otto proposed a unified vision of this apparent conflict supported by other studies: QS is typically activated in acute-phase infections whereas it is downregulated upon established chronic diseases [15].
Another intriguing form of communication reported in biofilm relays on electrical signals that can be transmitted by ferric ions and potassium ion channels present in the matrix, as it has been reported in different species (Bacillus subtilis, Geobacter sulfurreducens and Thiobacillus denitrificans) [60,61].
The restricted spatial distribution enables these communities to exert their effect on close-by cells in different manners: if nutritional sources are scarce, competition may occur between species, but many cooperative interactions have been proven [62,63,64,65]. In one instance, detossification of a toxic herbicide was achieved by a three-species biofilm (composed of Variovorax sp., Comamonas testosteroni and Hyphomicrobium sulfonivorans), whereas the cognate monospecies biofilms were not able to degrade it [66]. Another typical example of bacterial biofilm, where cooperation has been clearly described, is dental plaque, in which recruitment of different and successive species is strictly dependent on previous colonizers [67]. These examples show the many possible interactions that the biofilm environment offers and led to the utterly intriguing term of “sociomicrobiology” [68]. Further, this provides the basis for almost all biofilm-related concerns that are relevant for healthcare systems.
4. Nanotechnology as a Tool to Counteract Biofilm Infections: A Still Preclinical Issue
The relevance of biofilm in clinical settings arose in the 1980s when medical implants were found contaminated by bacteria upon analysis with electron microscopy [69,70,71].
As reported in Figure 3, the main sites of initial infections are either sites where medical manipulation occurred (prosthesis or catheter implantation) or areas where bacteria can gain easy access to bloodstream (periodontal or “gum” disease—as previously said, dental plaque is a form of biofilm–). The contamination at these sites enables access to bloodstream, that allows dispersion of single cells or biofilm clumps in the entire body, therefore reaching even deeper tissues (e.g., endocarditis). The adherence to a substratum and its localization in a specific site (with sporadic bacteremia) are among the four characteristics that Parsek and Singh proposed for biofilm-related infectious diseases [72]. The other two features are presence of a matrix-embedded biofilm upon microscopic observation and increased resistance to antibiotics despite antibiotic sensitivity of the constituent planktonic organisms. The latter is particularly relevant since it claims for alternative or complementary approaches for the reduction of the infection burden, since in many cases infection persists even after long-term aggressive antibiotic therapy, after which prosthesis or implant removal is necessary [73]. This implies higher costs in many different aspects, both from the patient side and the healthcare providers.
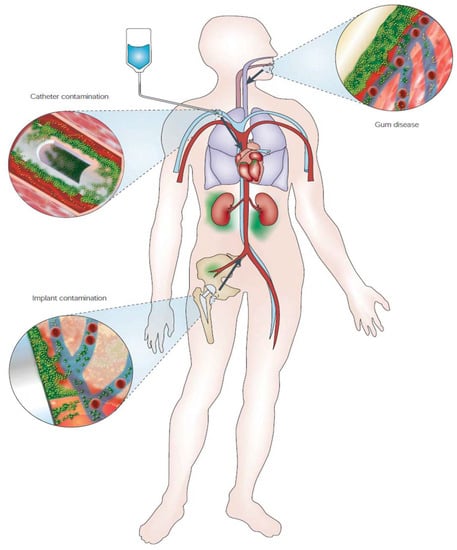
Figure 3.
Schematic representation of biofilm-associated infection disseminations, indicated by arrows. For more details, see main text. (With permission, from [24]).
The design of nanomaterials that are anti-infective by-design may help the field to improve not only medical treatments, but also environmental applications. The development of nanoparticles and nanomaterials in general is more focused on cancer and cardiovascular treatment and diagnostic purposes, in which these nanostructured materials have reached successful clinical applications (for extensive reviews, see [8,74,75]). In the field of antimicrobials, the application of nanotechnology is still at a pre-clinical or academic research level [76,77], with only some examples of wound-dressing, catheters, bone cements and cardiovascular implants containing silver as antimicrobial already available for clinical use [78]. As shown in Table 1, a simple search of the main US literature and clinical trial databases (PubMed and Clinicaltrail.gov, respectively) demonstrates that substantial effort is invested in academic research on nanoparticles in general, with minor attention to infections (166.221 vs. 2.604 hits) and biofilm (166.221 vs. 1.135 hits).

Table 1.
Academic and clinical research on metal nanoparticles in infectious diseases. Search of the databases was performed on 11th April 2018 using the mentioned search strings.
The clinical research and application are already a major barrier to most of the nanoparticle preparations, especially for infectious diseases, as reflected by the paucity of clinical trials involving nanoparticles. Clinical applications of nanosystems as antimicrobials are almost absent to the best of our knowledge. Therefore, we will focus on the preclinical research that has been conducted in this field.
5. Classification of Nanoproducts for Antimicrobial Nanotechnology
5.1. Traditional Classification
The development of nanosystems for antimicrobial application, as said, is intensively investigated at a preclinical level. A vast diversity of nanoproducts have been generated and different classifications have been used: the most common refers to the chemical composition of the nanoparticles. By contrast, the one proposed by Torchilin is based on functional aspects of the nanoformulated pharmaceutical [9]. A summary of these two different classifications is depicted in Figure 4.
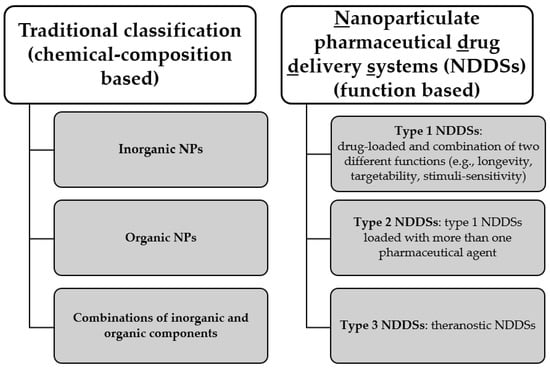
Figure 4.
Two different classifications of nanoparticles (NPs), based on their chemical composition or on their function. NDDSs classification has been proposed by Torchilin [9].
The first classification proposed divides the nanoparticles into different groups based on their chemical nature. This is by far the most common approach adopted in several reports to describe the different nanosystems generated (for extensive reviews, see [79,80,81,82,83]). Harnessing of metals or metal nanoparticles is the most frequently investigated nanotechnological application to endow materials with antibacterial and antibiofilm properties. The intrinsic nature of antimicrobials of various metal is known and enhanced by their formulation as inorganic nanoparticles, which allows a higher surface-to-volume ratio and increases controllability upon design, synthesis and tunability of their physico-chemical parameters. Mechanisms of bacterial killing are different with respect to both bacteria classes (Gram-positive or Gram-negative) and nanoparticles types and have been recently extensively reviewed [84,85,86,87]. Among the most common antibacterial mechanisms are membrane damage mediated by direct contact of nanoparticles with bacterial surface, protein chelation at Fe-S clusters or thiol residues, DNA and RNA damage and Reactive Oxygen Species (ROS) generation. A comprehensive picture of nanoparticles-mediated damages to the bacterial cell is given by Pelgrift and Friedman [84], reported in Figure 5.
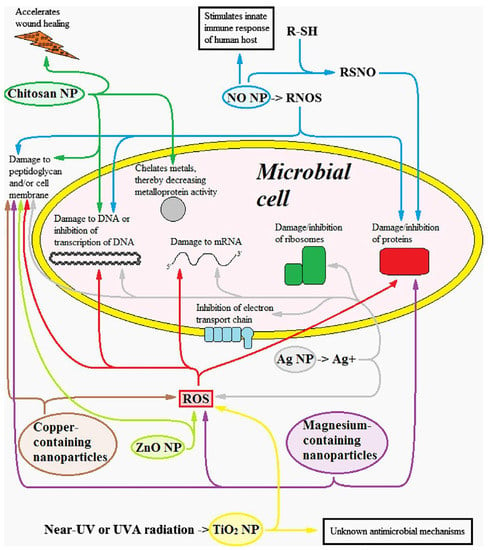
Figure 5.
Damages caused to the bacterial cell by different types of nanoparticles (NPs). Reactive Nitrogen Oxide Species (RNOS). Reactive Oxygen Species (ROS). R-SH schematizes a compound with a thiol (SH) group. RSNO stands for S-nitrosothiol. (With permission, from [84]).
According to the traditional classification, the second class of nanoparticles is the group of organic NPs, where organic molecules are part of the composition of the nanoparticles and exert an inherent antibiofilm action. In this class, lipid-based nanoparticles are also included. Examples are chitosan NPs [88] (which antibacterial effects are explained in Figure 5), functionalisation of polystyrene NPs with sulfate groups [89], nitrofuratonin conjugated to poly-l-lactic acid plastic [90], combination of perfluorinated lubricating fluids with polytetrafluoroethylene (PTFE) in membranes [91]. All these systems proved effective against biofilm.
The third class, which comprises all types of combinations between inorganic and organic components, is by far one of the most intensively investigated to endow the nanosystem with different antimicrobial activities, such as improved delivery of antibiotics or metals, increased stability and accumulation at the infection site, as well as increased cytocompatibility. Gallium-releasing Polyether urethane (PEU)-polyethylene glycol (PEG) blends proved effective in preventing P. aeruginosa infection upon implantation of the doped blend. The same blend loaded with Zinc showed much less efficacy [92]. Apart from metals, also antibiotics have been loaded in liposomes and their effectiveness has been proved [93], in some instances also in cystic fibrosis animal models [94].
5.2. Function-Based Classification
A more function-oriented classification has been recently proposed by Torchilin [9]. The three proposed groups distinguish between different types of Nanoparticulate pharmaceutical Drug Delivery Systems (NDDSs), as the author defines them. This perspective highlights the target of the NDDSs and their multiple functions. The majority of the applications of these systems are not for infectious diseases. However, some examples are present. Types 1 and 2 are drug-loaded NDDSs with at least two different functions (e.g., longevity, targetability, stimuli-sensitivity or cell penetration), the latter differing from the former, since it encapsulates more than one drug. Further examples of different multicomponent drug-delivery systems, apart from the ones cited above as combinations of organic and inorganic components that are part of this class if regarded from the point of view of functional classification, are nanoparticles releasing nitric oxide (NO) [95], three-component NPs (silver, chitosan and iodine) [96] and gentamicin-loaded silica hydrogel NPs [97]. The complexity of these nanosystems successfully targeted bacterial pathogens, but the analysis of their effect on biofilm is lacking. The combinations of multiple factors able to tackle bacterial survival results in the substantial merging of some classes belonging to different classification systems. In fact, nanoproducts included in the class in which inorganic and organic components are combined can fit either in type 1 or type 2 NDDSs, based on the number of different components that the nanosystem carries.
The third group encompasses theranostic NDDSs that conjugate diagnostic and therapeutic purposes. This last group has never been applied to biofilm infection, therefore this group will not be further described.
The two classifications presented here (composition-based and function-based) are therefore intended to better focus on the nanoproducts characteristics and possible applications and are not necessarily in contrast. Indeed, some points of contact are present.
6. Antibiofilm Application of Nanotechnology
Many strategies have been devised and proposed to tackle biofilm infections that can be classified in: (i) surface modification, to prevent bacterial adhesion and proliferation; (ii) nanoparticles or small molecules that can passively or actively target the biofilm; (iii) physical removal of the slime with different methods, as presented in Figure 6.
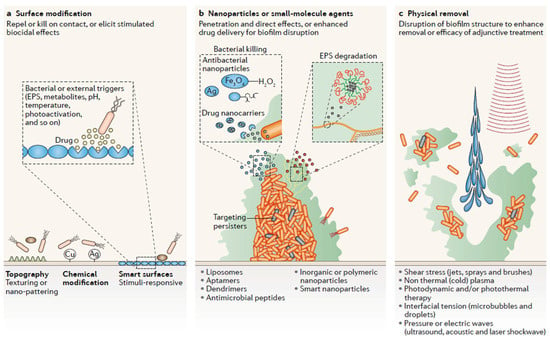
Figure 6.
Anti- biofilm strategies based on nanotechnological applications: (a) Modification of the surface , either topography or linkage with metal or stimuli-sensitive nanoparticles; (b) Direct contact between various types of nanoparticles and biofilm; (c) Physical methods to disperse biofilm coupled to nanotechnological improvement (With permission, from [7]).
6.1. Surface Modification
To reduce biofilm formation from its initial formation stages, the generation of coated surfaces that reduce or impede bacterial adhesion is intensively investigated, as summarized in Figure 6a. Nanotopography has proved relevant to promote bacterial killing, even though the mechanistic base is still unclear [98,99,100,101]. As previously said, impregnation or cross-linking of surfaces with metal-based nanoparticles (e.g., silver, gold, zinc and their oxides) is often investigated and provides successful biofilm reduction [102,103,104,105,106,107,108]. The use of “smart surfaces” with peculiar designs, that may change their behavior upon precise trigger, is of increasing interest in recent years [7,109,110,111].
6.2. Nanoparticles or Small-Molecule Agents
Direct effect of different types of nanoparticles on biofilm is the main investigation field, where many different approaches can be envisaged (Figure 6b). Simple and composite metallic nanoparticles are known to have effect both on planktonic and biofilm-embedded bacteria [76,112,113,114,115,116]. The attention to further aspects besides antimicrobial efficacy is crucial for further development and translation to clinics. The use of “green chemistry” to produce nanoparticles without polluting and toxic reagents is crucial to improve cytocompatibility and lower toxicity of the final product. For example, AgNP were produced using pectin from citrus peel acting as both reductant and stabilizer of the nanoparticle preparation. This strategy yielded an extremely stable product that was also fully cytocompatible and able to promote wound healing in vitro, besides showing high antibacterial and antibiofilm effects [117]. The design of NDDSs in recent years has increased [9], ranging from polymeric or organic nanoparticles, that might be conjugated to targeting molecules, antimicrobial compounds or enzymes in order to degrade biofilm matrix. The delivery of antibiotics is investigated to reduce side effects and increase local concentration of the active molecule inside the biofilm [118]. Furthermore, also the conjugation with enzymes to increase biofilm penetration has been explored, as in the case of poly(lactic-co-glycolic acid) PLGA nanocarriers loaded with DNase I and ciprofloxacin [119], demonstrating increased efficacy and good biofilm inhibition, since DNA is an essential component of EPS. The use of antimicrobial peptides conjugated to nanoparticles has also been explored, since these peptides are smaller and therefore more stable and protease-resistant than entire enzymes; this approach was efficient in vitro but no reports are available in in vivo models [120,121]. Targeting of persisters cells has been mainly achieved with free compounds [7,57,122,123]. Examples of persister cells causing latent infection upon dormant survival in macrophages is well-known and described for different pathogens [57,124,125]. Specific targeting of bacteria [126] or of macrophage-engulfed bacteria [127] was attained with multifunctional antibiotic-loaded nanoparticles, demonstrating the principles to tackle bacteria embedded inside immune cells. However, to the best of our knowledge, there is no report of the combinatorial use of nanomaterials in conjugation to antimicrobials able to tackle persister cells in biofilm. The evaluation of the synergistic effects provided by nanotechnology and antimicrobials will probably help to tackle persisters, mainly responsible for relapsing infections.
6.3. Physical Factors
Combination of physical removal of biofilm with other treatments is regarded with increasing attention since the sole mechanic disruption may lead to dispersal of small residues, therefore increasing the risk of infection [7,73]. Photothermal activity has been studied in conjunction with gold nanoparticles, owing to their intrinsic physico-chemical properties, resulting in significant biofilm reduction of both Gram-negative and Gram-positive bacteria [128,129]. Photodynamic treatment of biofilm in combination with organic compounds is another strategy that has been tested and yielded good results on staphylococcal biofilms [39,130,131,132].
Therefore, a wide range of nanotechnological applications are now available to tackle biofilm infection. Many of them showed promising features at laboratory level. Research effort should be focused on translating these principles into clinical application to reduce the well-known medical burden of biofilm communities.
7. Challenges and Future Perspectives
The use of nanotechonology in clinics is much more advanced in other fields compared to infectious diseases (e.g., cancer and cardiovascular pathologies, diagnostics) [8,74]. The advances of these fields may provide fruitful insights for antimicrobial nanotechnology development, not only for the pre-clinical design and analysis but also for the problems that may arise during clinical trials. The intense academic research in the field of antimicrobial nanotechnologies should be carefully designed towards a future possible clinical application. Therefore, deep characterization of the nanosystem needs to be provided early in the development of the product, pointing out to the necessity of a strict and strong collaboration between many different expertises (chemistry, biology, pharmacological, engineering, and clinical) during the development of the nanodrug. The interest in specific targeting and accumulation of nanoparticles at the site of disease is one of the most interesting aspects. It is deeply investigated in cancer-related applications of nanoparticles. Nevertheless, passive targeting through the Enhanced Permeability and Retention (EPR) effect and active targeting thanks to specific and selective mechanisms are far from a complete understanding since in vivo complexity poses major challenges to these two concepts [8,9,133]: different mammalian model organisms may have slightly different dynamics in different districts of the body, especially at pathological sites. This would cause different responses and therefore invalidate results obtained in animal models and hinder the translation of these results to humans. Furthermore, studies on how these nanoparticles interact with entire organisms are not often closely regarded, since the interactions of the nanosystem with blood and tissue components may easily change nanoparticles surface properties, which are critical for their properties [134,135]. Further, the long-term interaction of nanoparticles with the human body may have unwanted effects that are currently unknown, as no studies in this field are available. Particular attention should be addressed to the distribution and possible accumulation in specific organs. Clearance of the nanoparticles should also be investigated since it may require longer time compared to antimicrobials currently in use [136].
The necessity of large-scale production for clinical testing and application underlines the relevance of batch-to-batch reproducibility of preparation. As stated before, especially for infectious diseases, nanotechnologial approaches are mostly investigated at a preclinical level, where batch-to-batch variability is not attentively investigated. This represents a downside of the academic research. In view of a possible clinical translation, this aspect is crucial, thus it must be considered. In parallel, complex and articulate synthesis procedures might hinder large-scale synthesis. Therefore, development of complex multifunctional NDDSs might encounter more difficulties if clinical application is pursued. Another aspect that should be considered pertains to the stability of the generated nanoproduct. A precise analysis of the different storage conditions will provide a relevant piece of information for their clinical translation.
All these aspects are far from being comprehensively considered from a regulatory point of view for nanoproducts. New regulations specific to this new class of drugs are needed because of the peculiarities of these products [76,137,138].
The market of nanomedicines is set to reach $400 billion by 2019, showing a high interest in this field [137]. Together with the increasing problem of antibiotic resistance and the major impact of biofilm infections, antimicrobial nanodrugs hold a great potential for future developments and their impact on the healthcare systems will be considerable. The lessons learned from other fields will allow a more reliable design and development of these products, allowing patients and clinicians to have more therapeutic approaches to tackle infectious diseases in the near future.
Author Contributions
All authors participated in writing and/or reviewing the manuscript.
Acknowledgments
The authors gratefully acknowledge Ranju Prasad Mandal for the critical reading of the manuscript and helpful discussions. The authors also thank Marta Bordoni for carefully proofreading the manuscript. L.V. would also like to thank COST Action iPROMEDAI TD1305, “Improved Protection of Medical Devices Against Infection (IPROMEDAI)” (2013–2018) (http://www.cost.eu/COST_Actions/tdp/TD1305).
Conflicts of Interest
The authors declare no conflict of interest.
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; The Review on Antimicrobial Resistance; AMR Review: London, UK, 2016. [Google Scholar]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Arias, C.A.; Murray, B.E. A new antibiotic and the evolution of resistance. N. Engl. J. Med. 2015, 372, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The who priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Torchilin, V.P. Multifunctional, stimuli-sensitive nanoparticulate systems for drug delivery. Nat. Rev. Drug Discov. 2014, 13, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Barenholz, Y. Doxil®—The first FDA-approved nano-drug: Lessons learned. J. Control. Release 2012, 160, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Aversa, F.; Busca, A.; Candoni, A.; Cesaro, S.; Girmenia, C.; Luppi, M.; Nosari, A.M.; Pagano, L.; Romani, L.; Rossi, G.; et al. Liposomal amphotericin B (AmBisome®) at beginning of its third decade of clinical use. J. Chemother. 2017, 29, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.C. Interfaces in Microbial Ecology; Harvard University Press: Cambridge, MA, USA, 1976; pp. 44–47. [Google Scholar]
- Costerton, J.W.; Geesey, G.G.; Cheng, K.J. How bacteria stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C. EPS—Then and now. Microorganisms 2016, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Otto, M. Molecular basis of in vivo biofilm formation by bacterial pathogens. Chem. Biol. 2012, 19, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Cramton, S.E.; Götz, F.; Peschel, A. Key role of teichoic acid net charge in Staphylococcus aureus colonization of artificial surfaces. Infect. Immun. 2001, 69, 3423–3426. [Google Scholar] [CrossRef] [PubMed]
- Vallet, I.; Olson, J.W.; Lory, S.; Lazdunski, A.; Filloux, A. The chaperone/usher pathways of Pseudomonas aeruginosa: Identification of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc. Natl. Acad. Sci. USA 2001, 98, 6911–6916. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Characklis, W.G.; Marshall, K.C. Biofilms; John Wiley & Sons, Inc.: New York, NY, USA, 1990. [Google Scholar]
- Purevdorj, B.; Costerton, J.W.; Stoodley, P. Influence of hydrodynamics and cell signaling on the structure and behavior of Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2002, 68, 4457–4464. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Cargo, R.; Rupp, C.J.; Wilson, S.; Klapper, I. Biofilm material properties as related to shear-induced deformation and detachment phenomena. J. Ind. Microbiol. Biotechnol. 2002, 29, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.W.; He, Y.; Ren, Y.; Zerdoum, A.; Libera, M.R.; Sharma, P.K.; van Winkelhoff, A.J.; Neut, D.; Stoodley, P.; van der Mei, H.C.; et al. Viscoelasticity of biofilms and their recalcitrance to mechanical and chemical challenges. FEMS Microbiol. Rev. 2015, 39, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Rodesney, C.A.; Roman, B.; Dhamani, N.; Cooley, B.J.; Katira, P.; Touhami, A.; Gordon, V.D. Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-gmp signal initiating biofilm development. Proc. Natl. Acad. Sci. USA 2017, 114, 5906–5911. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, D.; Francois, P.; Vaudaux, P.; Foster, T.J. Identification of the ligand-binding domain of the surface-located fibrinogen receptor (clumping factor) of Staphylococcus aureus. Mol. Microbiol. 1995, 16, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Ní Eidhin, D.; Perkins, S.; Francois, P.; Vaudaux, P.; Höök, M.; Foster, T.J. Clumping factor B (ClfB), a new surface-located fibrinogen-binding adhesin of Staphylococcus aureus. Mol. Microbiol. 1998, 30, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Fröman, G.; Switalski, L.M.; Speziale, P.; Höök, M. Isolation and characterization of a fibronectin receptor from Staphylococcus aureus. J. Biol. Chem. 1987, 262, 6564–6571. [Google Scholar] [PubMed]
- Jönsson, K.; Signäs, C.; Müller, H.P.; Lindberg, M. Two different genes encode fibronectin binding proteins in Staphylococcus aureus. The complete nucleotide sequence and characterization of the second gene. Eur. J. Biochem. 1991, 202, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Patti, J.M.; Boles, J.O.; Höök, M. Identification and biochemical characterization of the ligand binding domain of the collagen adhesin from Staphylococcus aureus. Biochemistry 1993, 32, 11428–11435. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Xu, Y.; Liang, X.; Keene, D.R.; Höök, A.; Gurusiddappa, S.; Höök, M.; Narayana, S.V. A ‘collagen hug’ model for Staphylococcus aureus cna binding to collagen. EMBO J. 2005, 24, 4224–4236. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penadés, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, M.M.; Chapman, M.R. Curli biogenesis and function. Annu. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of staphylococcus epidermidis is a linear beta-1,6-linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 1996, 178, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Götz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [PubMed]
- Friedman, L.; Kolter, R. Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J. Bacteriol. 2004, 186, 4457–4465. [Google Scholar] [CrossRef] [PubMed]
- Speziale, P.; Pietrocola, G.; Foster, T.J.; Geoghegan, J.A. Protein-based biofilm matrices in staphylococci. Front. Cell. Infect. Microbiol. 2014, 4, 171. [Google Scholar] [CrossRef] [PubMed]
- Di Poto, A.; Sbarra, M.S.; Provenza, G.; Visai, L.; Speziale, P. The effect of photodynamic treatment combined with antibiotic action or host defence mechanisms on Staphylococcus aureus biofilms. Biomaterials 2009, 30, 3158–3166. [Google Scholar] [CrossRef] [PubMed]
- Tielen, P.; Kuhn, H.; Rosenau, F.; Jaeger, K.E.; Flemming, H.C.; Wingender, J. Interaction between extracellular lipase lipa and the polysaccharide alginate of Pseudomonas aeruginosa. BMC Microbiol. 2013, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Schooling, S.R.; Beveridge, T.J. Membrane vesicles: An overlooked component of the matrices of biofilms. J. Bacteriol. 2006, 188, 5945–5957. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identification of proteins associated with the Pseudomonas aeruginosa biofilm extracellular matrix. J. Proteome Res. 2012, 11, 4906–4915. [Google Scholar] [CrossRef] [PubMed]
- Zobell, C.E. The effect of solid surfaces upon bacterial activity. J. Bacteriol. 1943, 46, 39–56. [Google Scholar] [PubMed]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular dna required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef] [PubMed]
- Dengler, V.; Foulston, L.; DeFrancesco, A.S.; Losick, R. An electrostatic net model for the role of extracellular dna in biofilm formation by Staphylococcus aureus. J. Bacteriol. 2015, 197, 3779–3787. [Google Scholar] [CrossRef] [PubMed]
- Izano, E.A.; Amarante, M.A.; Kher, W.B.; Kaplan, J.B. Differential roles of poly-N-acetylglucosamine surface polysaccharide and extracellular dna in Staphylococcus aureus and staphylococcus epidermidis biofilms. Appl. Environ. Microbiol. 2008, 74, 470–476. [Google Scholar] [CrossRef] [PubMed]
- McDougald, D.; Rice, S.A.; Barraud, N.; Steinberg, P.D.; Kjelleberg, S. Should we stay or should we go: Mechanisms and ecological consequences for biofilm dispersal. Nat. Rev. Microbiol. 2011, 10, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C. The perfect slime. Colloids Surf. B Biointerfaces 2011, 86, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Dobor, J.; Varga, M.; Záray, G. Biofilm controlled sorption of selected acidic drugs on river sediments characterized by different organic carbon content. Chemosphere 2012, 87, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Writer, J.H.; Barber, L.B.; Ryan, J.N.; Bradley, P.M. Biodegradation and attenuation of steroidal hormones and alkylphenols by stream biofilms and sediments. Environ. Sci. Technol. 2011, 45, 4370–4376. [Google Scholar] [CrossRef] [PubMed]
- Writer, J.H.; Ryan, J.N.; Barber, L.B. Role of biofilms in sorptive removal of steroidal hormones and 4-nonylphenol compounds from streams. Environ. Sci. Technol. 2011, 45, 7275–7283. [Google Scholar] [CrossRef] [PubMed]
- López, D.; Vlamakis, H.; Losick, R.; Kolter, R. Cannibalism enhances biofilm development in bacillus subtilis. Mol. Microbiol. 2009, 74, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Königs, A.M.; Flemming, H.C.; Wingender, J. Nanosilver induces a non-culturable but metabolically active state in Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 395. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Allison, D.G.; Gilbert, P. Resistance of bacterial biofilms to antibiotics: A growth-rate related effect? J. Antimicrob. Chemother. 1988, 22, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, D.; Losick, R. How and why bacteria talk to each other. Cell 1993, 73, 873–885. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef] [PubMed]
- Prindle, A.; Liu, J.; Asally, M.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Ion channels enable electrical communication in bacterial communities. Nature 2015, 527, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 2014, 8, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.K.; Rainey, P.B.; Haagensen, J.A.; Molin, S. Evolution of species interactions in a biofilm community. Nature 2007, 445, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Bell, T. Competition, not cooperation, dominates interactions among culturable microbial species. Curr. Biol. 2012, 22, 1845–1850. [Google Scholar] [CrossRef] [PubMed]
- Breugelmans, P.; Barken, K.B.; Tolker-Nielsen, T.; Hofkens, J.; Dejonghe, W.; Springael, D. Architecture and spatial organization in a triple-species bacterial biofilm synergistically degrading the phenylurea herbicide linuron. FEMS Microbiol. Ecol. 2008, 64, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Peters, G.; Locci, R.; Pulverer, G. Microbial colonization of prosthetic devices. II. Scanning electron microscopy of naturally infected intravenous catheters. Zent. Bakteriol. Mikrobiol. Hyg. B 1981, 173, 293–299. [Google Scholar]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [PubMed]
- Marrie, T.J.; Nelligan, J.; Costerton, J.W. A scanning and transmission electron microscopic study of an infected endocardial pacemaker lead. Circulation 1982, 66, 1339–1341. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. A review of clinical translation of inorganic nanoparticles. AAPS J. 2015, 17, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Weissig, V.; Pettinger, T.K.; Murdock, N. Nanopharmaceuticals (part 1): Products on the market. Int. J. Nanomed. 2014, 9, 4357–4373. [Google Scholar] [CrossRef] [PubMed]
- Natan, M.; Banin, E. From nano to micro: Using nanotechnology to combat microorganisms and their multidrug resistance. FEMS Microbiol. Rev. 2017, 41, 302–322. [Google Scholar] [CrossRef] [PubMed]
- Weissig, V.; Guzman-Villanueva, D. Nanopharmaceuticals (part 2): Products in the pipeline. Int. J. Nanomed. 2015, 10, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Li, Q.; Wang, M.; Ouyang, J.; Li, X.; Xing, M.M. Nanosilver particles in medical applications: Synthesis, performance, and toxicity. Int. J. Nanomed. 2014, 9, 2399–2407. [Google Scholar]
- Neethirajan, S.; Clond, M.A.; Vogt, A. Medical biofilms--nanotechnology approaches. J. Biomed. Nanotechnol. 2014, 10, 2806–2827. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Low, W.L.; Gupta, A.; Amin, M.C.; Radecka, I.; Britland, S.T.; Raj, P.; Kenward, K.M. Strategies for antimicrobial drug delivery to biofilm. Curr. Pharm. Des. 2015, 21, 43–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H. Nanoparticle-based therapies for wound biofilm infection: Opportunities and challenges. IEEE Trans. Nanobiosci. 2016, 15, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M.; Lee, J. Recent nanotechnology approaches for prevention and treatment of biofilm-associated infections on medical devices. Biomed. Res. Int. 2016, 2016, 1851242. [Google Scholar] [CrossRef] [PubMed]
- Hemeg, H.A. Nanomaterials for alternative antibacterial therapy. Int. J. Nanomed. 2017, 12, 8211–8225. [Google Scholar] [CrossRef] [PubMed]
- Pelgrift, R.Y.; Friedman, A.J. Nanotechnology as a therapeutic tool to combat microbial resistance. Adv. Drug Deliv. Rev. 2013, 65, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Mijnendonckx, K.; Leys, N.; Mahillon, J.; Silver, S.; Van Houdt, R. Antimicrobial silver: Uses, toxicity and potential for resistance. Biometals 2013, 26, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Neoh, K.G.; Kang, E.T.; Wang, W. Antibacterial and mechanical properties of bone cement impregnated with chitosan nanoparticles. Biomaterials 2006, 27, 2440–2449. [Google Scholar] [CrossRef] [PubMed]
- Nevius, B.A.; Chen, Y.P.; Ferry, J.L.; Decho, A.W. Surface-functionalization effects on uptake of fluorescent polystyrene nanoparticles by model biofilms. Ecotoxicology 2012, 21, 2205–2213. [Google Scholar] [CrossRef] [PubMed]
- Sandler, N.; Salmela, I.; Fallarero, A.; Rosling, A.; Khajeheian, M.; Kolakovic, R.; Genina, N.; Nyman, J.; Vuorela, P. Towards fabrication of 3d printed medical devices to prevent biofilm formation. Int. J. Pharm. 2014, 459, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Epstein, A.K.; Wong, T.S.; Belisle, R.A.; Boggs, E.M.; Aizenberg, J. Liquid-infused structured surfaces with exceptional anti-biofouling performance. Proc. Natl. Acad. Sci. USA 2012, 109, 13182–13187. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Darmawan, E.T.; Zhang, M.; Zhang, L.; Bryers, J.D. Development of a poly(ether urethane) system for the controlled release of two novel anti-biofilm agents based on gallium or zinc and its efficacy to prevent bacterial biofilm formation. J. Control. Release 2013, 172, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M.; Halwani, M.; Omri, A.; Suntres, Z.E. Antimicrobial effectiveness of liposomal polymyxin B against resistant Gram-negative bacterial strains. Int. J. Pharm. 2008, 355, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Alhariri, M.; Omri, A. Efficacy of liposomal bismuth-ethanedithiol-loaded tobramycin after intratracheal administration in rats with pulmonary Pseudomonas aeruginosa infection. Antimicrob. Agents Chemother. 2013, 57, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Mihu, M.R.; Sandkovsky, U.; Han, G.; Friedman, J.M.; Nosanchuk, J.D.; Martinez, L.R. The use of nitric oxide releasing nanoparticles as a treatment against acinetobacter baumannii in wound infections. Virulence 2010, 1, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Mallick, S.; Paul, A.; Chattopadhyay, A.; Ghosh, S.S. Heightened reactive oxygen species generation in the antimicrobial activity of a three component iodinated chitosan-silver nanoparticle composite. Langmuir 2010, 26, 5901–5908. [Google Scholar] [CrossRef] [PubMed]
- Seleem, M.N.; Munusamy, P.; Ranjan, A.; Alqublan, H.; Pickrell, G.; Sriranganathan, N. Silica-antibiotic hybrid nanoparticles for targeting intracellular pathogens. Antimicrob. Agents Chemother. 2009, 53, 4270–4274. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J.F.; Aldred, N.; Callow, M.E.; Finlay, J.A.; Callow, J.A.; Clare, A.S.; Brennan, A.B. Species-specific engineered antifouling topographies: Correlations between the settlement of algal zoospores and barnacle cyprids. Biofouling 2007, 23, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Puckett, S.D.; Taylor, E.; Raimondo, T.; Webster, T.J. The relationship between the nanostructure of titanium surfaces and bacterial attachment. Biomaterials 2010, 31, 706–713. [Google Scholar] [CrossRef] [PubMed]
- May, R.M.; Magin, C.M.; Mann, E.E.; Drinker, M.C.; Fraser, J.C.; Siedlecki, C.A.; Brennan, A.B.; Reddy, S.T. An engineered micropattern to reduce bacterial colonization, platelet adhesion and fibrin sheath formation for improved biocompatibility of central venous catheters. Clin. Transl. Med. 2015, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, A.; Sen, P.; Su, B.; Briscoe, W.H. Natural and bioinspired nanostructured bactericidal surfaces. Adv. Colloid Interface Sci. 2017, 248, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Taglietti, A.; Arciola, C.R.; D’Agostino, A.; Dacarro, G.; Montanaro, L.; Campoccia, D.; Cucca, L.; Vercellino, M.; Poggi, A.; Pallavicini, P.; et al. Antibiofilm activity of a monolayer of silver nanoparticles anchored to an amino-silanized glass surface. Biomaterials 2014, 35, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Cochis, A.; Azzimonti, B.; Della Valle, C.; De Giglio, E.; Bloise, N.; Visai, L.; Cometa, S.; Rimondini, L.; Chiesa, R. The effect of silver or gallium doped titanium against the multidrug resistant acinetobacter baumannii. Biomaterials 2016, 80, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Cochis, A.; Azzimonti, B.; Sorrentino, R.; Della Valle, C.; De Giglio, E.; Bloise, N.; Visai, L.; Bruni, G.; Cometa, S.; Pezzoli, D.; et al. Data in support of gallium (Ga3+) antibacterial activities to counteract E. coli and S. epidermidis biofilm formation onto pro-osteointegrative titanium surfaces. Data Brief. 2016, 6, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Giordano, C.; Saino, E.; Rimondini, L.; Pedeferri, M.P.; Visai, L.; Cigada, A.; Chiesa, R. Electrochemically induced anatase inhibits bacterial colonization on titanium Grade 2 and Ti6Al4V alloy for dental and orthopedic devices. Colloids Surf. B Biointerfaces 2011, 88, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Petrini, P.; Arciola, C.R.; Pezzali, I.; Bozzini, S.; Montanaro, L.; Tanzi, M.C.; Speziale, P.; Visai, L. Antibacterial activity of zinc modified titanium oxide surface. Int. J. Artif. Organs 2006, 29, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Swartjes, J.J.; Sharma, P.K.; van Kooten, T.G.; van der Mei, H.C.; Mahmoudi, M.; Busscher, H.J.; Rochford, E.T. Current developments in antimicrobial surface coatings for biomedical applications. Curr. Med. Chem. 2015, 22, 2116–2129. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Ray, B.; Ranjit, K.T.; Manna, A.C. Antibacterial activity of zno nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol. Lett. 2008, 279, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Claes, B.; Boudewijns, T.; Muchez, L.; Hooyberghs, G.; Van der Eycken, E.V.; Vanderleyden, J.; Steenackers, H.P.; De Vos, D.E. Smart metal-organic framework coatings: Triggered antibiofilm compound release. ACS Appl. Mater. Interfaces 2017, 9, 4440–4449. [Google Scholar] [CrossRef] [PubMed]
- Pallavicini, P.; Donà, A.; Taglietti, A.; Minzioni, P.; Patrini, M.; Dacarro, G.; Chirico, G.; Sironi, L.; Bloise, N.; Visai, L.; et al. Self-assembled monolayers of gold nanostars: A convenient tool for near-ir photothermal biofilm eradication. Chem. Commun. 2014, 50, 1969–1971. [Google Scholar] [CrossRef] [PubMed]
- Armentano, I.; Arciola, C.R.; Fortunati, E.; Ferrari, D.; Mattioli, S.; Amoroso, C.F.; Rizzo, J.; Kenny, J.M.; Imbriani, M.; Visai, L. The interaction of bacteria with engineered nanostructured polymeric materials: A review. Sci. World J. 2014, 2014, 410423. [Google Scholar] [CrossRef] [PubMed]
- Bari, A.; Bloise, N.; Fiorilli, S.; Novajra, G.; Vallet-Regí, M.; Bruni, G.; Torres-Pardo, A.; González-Calbet, J.M.; Visai, L.; Vitale-Brovarone, C. Copper-containing mesoporous bioactive glass nanoparticles as multifunctional agent for bone regeneration. Acta Biomater. 2017, 55, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Ahmed, A.; Prasad, K.N.; Khanduja, S.; Singh, S.K.; Srivastava, J.K.; Gajbhiye, N.S. Antibiofilm and membrane-damaging potential of cuprous oxide nanoparticles against Staphylococcus aureus with reduced susceptibility to vancomycin. Antimicrob. Agents Chemother. 2015, 59, 6882–6890. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Ju, E.; Ren, J.; Qu, X. Bifunctionalized mesoporous silica-supported gold nanoparticles: Intrinsic oxidase and peroxidase catalytic activities for antibacterial applications. Adv. Mater. 2015, 27, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.H.; Ke, W.J.; Hsieh, C.T.; Lin, K.S.; Tzou, D.Y.; Chiang, C.L. Zno nanoparticles affect bacillus subtilis cell growth and biofilm formation. PLoS ONE 2015, 10, e0128457. [Google Scholar] [CrossRef] [PubMed]
- Pallavicini, P.; Arciola, C.R.; Bertoglio, F.; Curtosi, S.; Dacarro, G.; D’Agostino, A.; Ferrari, F.; Merli, D.; Milanese, C.; Rossi, S.; et al. Silver nanoparticles synthesized and coated with pectin: An ideal compromise for anti-bacterial and anti-biofilm action combined with wound-healing properties. J. Colloid Interface Sci. 2017, 498, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Fazly Bazzaz, B.S.; Khameneh, B.; Zarei, H.; Golmohammadzadeh, S. Antibacterial efficacy of rifampin loaded solid lipid nanoparticles against staphylococcus epidermidis biofilm. Microb. Pathog. 2016, 93, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Baelo, A.; Levato, R.; Julián, E.; Crespo, A.; Astola, J.; Gavaldà, J.; Engel, E.; Mateos-Timoneda, M.A.; Torrents, E. Disassembling bacterial extracellular matrix with dnase-coated nanoparticles to enhance antibiotic delivery in biofilm infections. J. Control. Release 2015, 209, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Kalia, P.; Jain, A.; Radha Krishnan, R.; Demuth, D.R.; Steinbach-Rankins, J.M. Peptide-modified nanoparticles inhibit formation of porphyromonas gingivalis biofilms with streptococcus gordonii. Int. J. Nanomed. 2017, 12, 4553–4562. [Google Scholar] [CrossRef] [PubMed]
- Niemirowicz, K.; Surel, U.; Wilczewska, A.Z.; Mystkowska, J.; Piktel, E.; Gu, X.; Namiot, Z.; Kułakowska, A.; Savage, P.B.; Bucki, R. Bactericidal activity and biocompatibility of ceragenin-coated magnetic nanoparticles. J. Nanobiotechnol. 2015, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Zhu, W.; Hendricks, G.L.; Van Tyne, D.; Steele, A.D.; Keohane, C.E.; Fricke, N.; Conery, A.L.; Shen, S.; Pan, W.; et al. A new class of synthetic retinoid antibiotics effective against bacterial persisters. Nature 2018, 556, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P.; Nakayasu, E.S.; Fleck, L.E.; LaFleur, M.D.; Isabella, V.M.; Coleman, K.; Leonard, S.N.; Smith, R.D.; Adkins, J.N.; Lewis, K. Activated clpp kills persisters and eradicates a chronic biofilm infection. Nature 2013, 503, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells: Molecular mechanisms related to antibiotic tolerance. Handb. Exp. Pharmacol. 2012, 121–133. [Google Scholar]
- Helaine, S.; Cheverton, A.M.; Watson, K.G.; Faure, L.M.; Matthews, S.A.; Holden, D.W. Internalization of salmonella by macrophages induces formation of nonreplicating persisters. Science 2014, 343, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Li, L.; Yu, F.; Wang, H. Vancomycin-modified mesoporous silica nanoparticles for selective recognition and killing of pathogenic gram-positive bacteria over macrophage-like cells. ACS Appl. Mater. Interfaces 2013, 5, 10874–10881. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.H.; Li, Y.J.; Bao, Y.; Yang, X.Z.; Hu, B.; Wang, J. Bacteria-responsive multifunctional nanogel for targeted antibiotic delivery. Adv. Mater. 2012, 24, 6175–6180. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Singh, A.K.; Senapati, D.; Neely, A.; Yu, H.; Ray, P.C. Rapid colorimetric identification and targeted photothermal lysis of salmonella bacteria by using bioconjugated oval-shaped gold nanoparticles. Chemistry 2010, 16, 5600–5606. [Google Scholar] [CrossRef] [PubMed]
- Khantamat, O.; Li, C.H.; Yu, F.; Jamison, A.C.; Shih, W.C.; Cai, C.; Lee, T.R. Gold nanoshell-decorated silicone surfaces for the near-infrared (NIR) photothermal destruction of the pathogenic bacterium E. faecalis. ACS Appl. Mater. Interfaces 2015, 7, 3981–3993. [Google Scholar] [CrossRef] [PubMed]
- Sbarra, M.S.; Di Poto, A.; Arciola, C.R.; Saino, E.; Sharma, M.; Bragheri, F.; Cristiani, I.; Speziale, P.; Visai, L. Photodynamic action of merocyanine 540 on staphylococcus epidermidis biofilms. Int. J. Artif. Organs 2008, 31, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Saino, E.; Sbarra, M.S.; Arciola, C.R.; Scavone, M.; Bloise, N.; Nikolov, P.; Ricchelli, F.; Visai, L. Photodynamic action of tri-meso (N-methyl-pyridyl), meso (N-tetradecyl-pyridyl) porphine on staphylococcus epidermidis biofilms grown on Ti6Al4V alloy. Int. J. Artif. Organs 2010, 33, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Visai, L.; Bragheri, F.; Cristiani, I.; Gupta, P.K.; Speziale, P. Toluidine blue-mediated photodynamic effects on staphylococcal biofilms. Antimicrob. Agents Chemother. 2008, 52, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, U.; Maeda, H.; Jain, R.K.; Sevick-Muraca, E.M.; Zamboni, W.; Farokhzad, O.C.; Barry, S.T.; Gabizon, A.; Grodzinski, P.; Blakey, D.C. Challenges and key considerations of the enhanced permeability and retention effect for nanomedicine drug delivery in oncology. Cancer Res. 2013, 73, 2412–2417. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, N.; Leroux, J.C. The journey of a drug-carrier in the body: An anatomo-physiological perspective. J. Control. Release 2012, 161, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, N.; Grenier, P.; Mahmoudi, M.; Lima, E.M.; Appel, E.A.; Dormont, F.; Lim, J.M.; Karnik, R.; Langer, R.; Farokhzad, O.C. Mechanistic understanding of in vivo protein corona formation on polymeric nanoparticles and impact on pharmacokinetics. Nat. Commun. 2017, 8, 777. [Google Scholar] [CrossRef] [PubMed]
- Beyth, N.; Houri-Haddad, Y.; Domb, A.; Khan, W.; Hazan, R. Alternative antimicrobial approach: Nano-antimicrobial materials. Evid. Based Complement. Altern. Med. 2015, 2015, 246012. [Google Scholar] [CrossRef] [PubMed]
- Ragelle, H.; Danhier, F.; Préat, V.; Langer, R.; Anderson, D.G. Nanoparticle-based drug delivery systems: A commercial and regulatory outlook as the field matures. Expert Opin. Drug Deliv. 2017, 14, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Hafner, A.; Lovrić, J.; Lakoš, G.P.; Pepić, I. Nanotherapeutics in the EU: An overview on current state and future directions. Int. J. Nanomed. 2014, 9, 1005–1023. [Google Scholar]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).