
Cytotoxic and Antioxidant Properties and Profile of Active Compounds in Kale and Lupine Sprouts Supplemented with γ-Polyglutamic Acid During Sprouting
 , , , and
, , , and
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions for Sprouts
2.2. Extracts Preparation
2.3. HPLC Determination of Phenolic Acids and Isoflavones in Sprouts
2.4. Cytotoxicity and Viability Assay
2.5. Determination of Antioxidant Activity and Total Reducing Capacity
2.6. Statistical Analysis
3. Results and Discussion
3.1. Active Compounds
3.2. Cytotoxic Activity
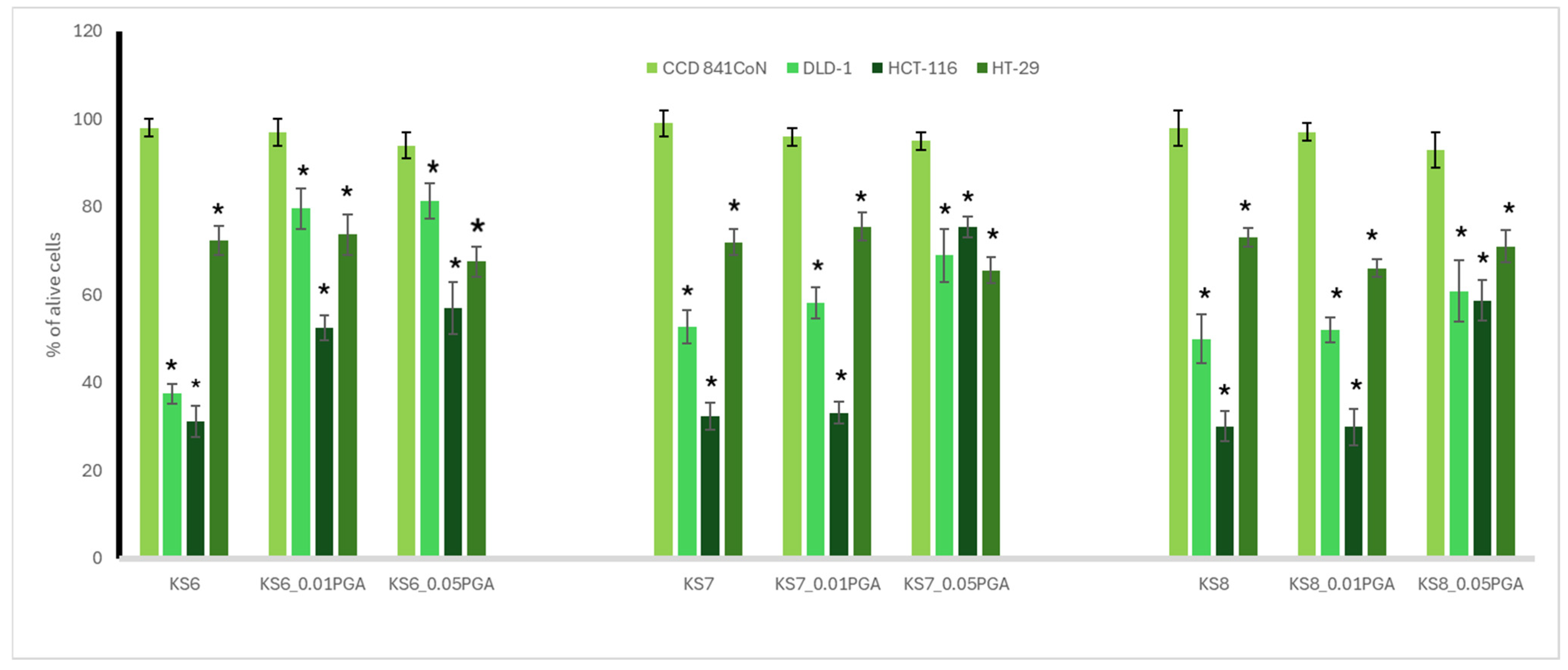
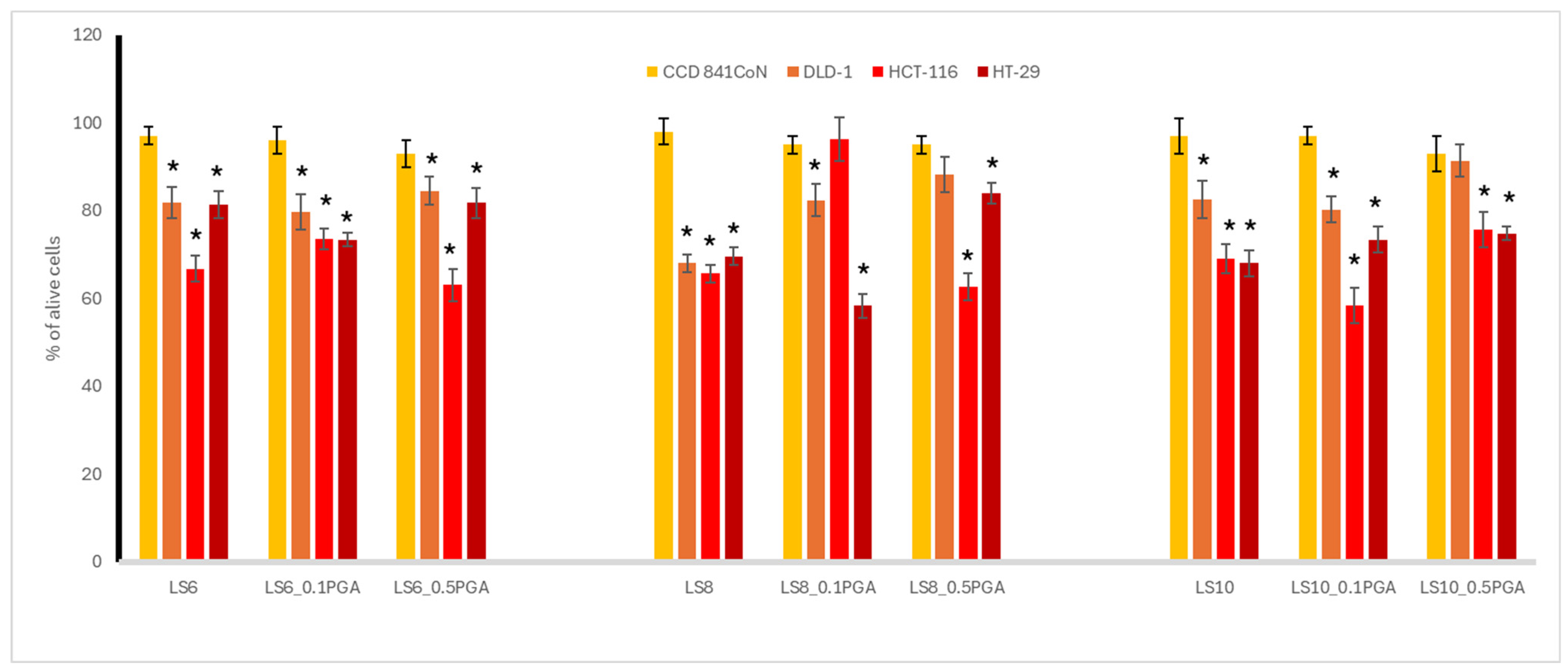
3.3. Antioxidant Potential
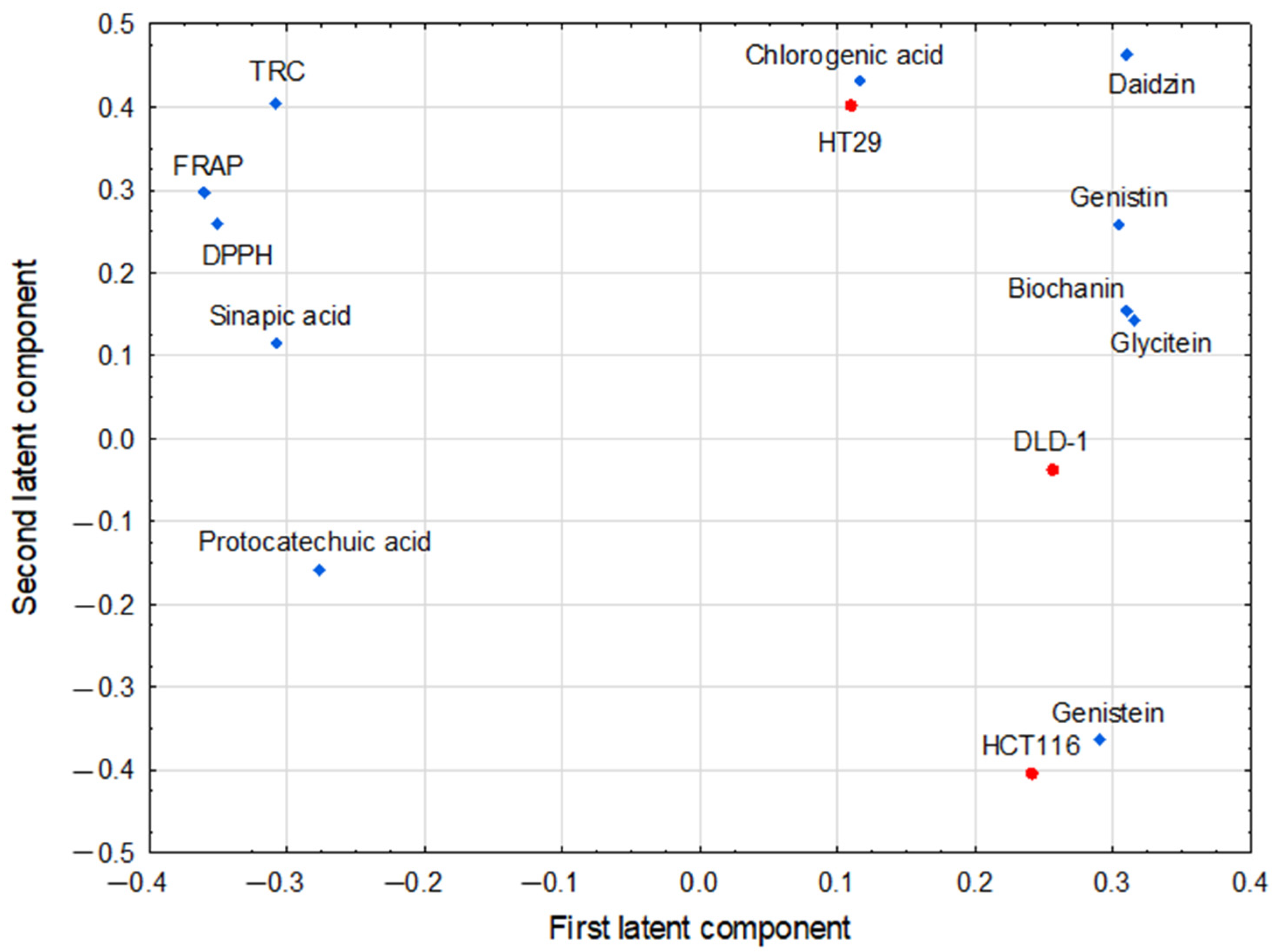
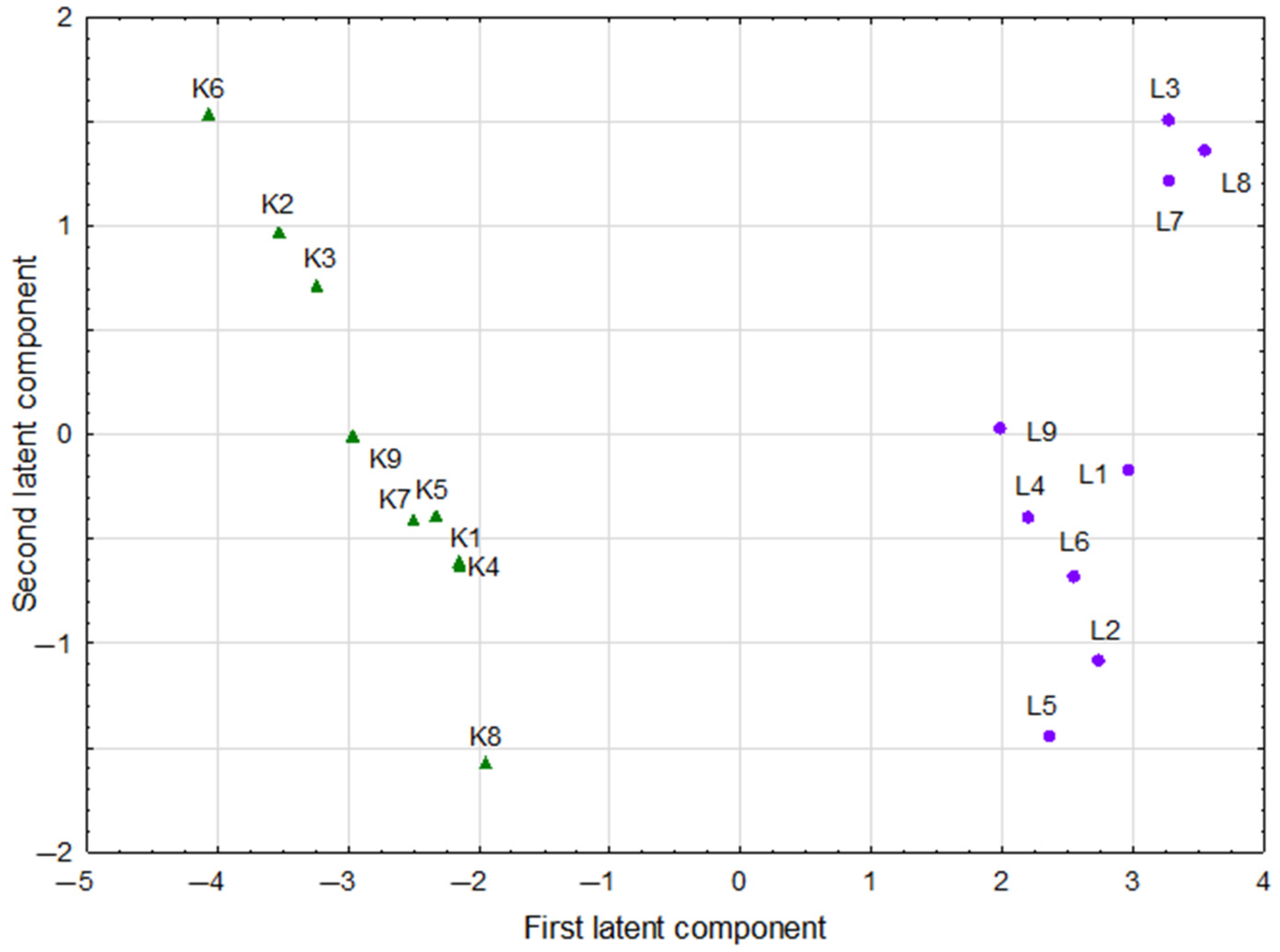
3.4. Multivariate Analysis of Studied Parameters
3.5. Limitations of This Work
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Chen, S.; Yu, B. Poly-γ-glutamic acid: Recent achievements, diverse applications and future perspectives. Trends Food Sci. Technol. 2022, 119, 1–12. [Google Scholar] [CrossRef]
- Wojtowicz, K.; Steliga, T.; Kapusta, P.; Brzeszcz, J.; Skalski, T. Evaluation of the effectiveness of the biopreparation in combination with the polymer γ-PGA for the biodegradation of petroleum contaminants in soil. Materials 2022, 15, 400. [Google Scholar] [CrossRef]
- Ogunleye, A.; Bhat, A.; Irorere, V.U.; Hill, D.; Williams, C.; Radecka, I. Poly-γ-glutamic acid: Production, properties and applications. Microbiology 2015, 161, 1–17. [Google Scholar] [CrossRef]
- Luo, Z.; Guo, Y.; Liu, J.; Qiu, H.; Zhao, M.; Zou, W.; Li, S. Microbial synthesis of poly-γ-glutamic acid: Current progress, challenges, and future perspectives. Biotechnol. Biofuels Bioprod. 2016, 9, 134. [Google Scholar] [CrossRef]
- Feng, J.; Gu, Y.; Quan, Y.; Cao, M.; Gao, W.; Zhang, W. Improved poly-γ-glutamic acid production in Bacillus amyloliquefaciens by modular pathway engineering. Metab. Eng. 2015, 32, 106–115. [Google Scholar] [CrossRef]
- Nabi, I.; Das, S. Poly-glutamic acid (PGA)—Structure, synthesis, genomic organization and its application: A review. Int. J. Pharm. Sci. Res. 2015, 6, 2258–2280. [Google Scholar]
- Li, D.; Hou, L.; Gao, Y.; Tian, Z.; Fan, B.; Wang, F.; Li, S. Recent advances in microbial synthesis of poly-γ-glutamic acid: A review. Foods 2022, 11, 739. [Google Scholar] [CrossRef]
- Xu, Z.; Lei, P.; Feng, X.; Xu, X.; Xu, H.; Yang, H.; Tang, W. Effect of poly(γ-glutamic acid) on microbial community and nitrogen pools of soil. Acta Agr. Scand. 2013, 63, 657–668. [Google Scholar] [CrossRef]
- Xu, Z.; Wan, C.; Xu, X.; Feng, X.; Xu, H. Effect of poly (γ-glutamic acid) on wheat productivity, nitrogen use efficiency and soil microbes. J. Soil. Sci. Plant Nut. 2013, 13, 744–755. [Google Scholar] [CrossRef]
- Xu, Z.; Lei, P.; Pang, X.; Li, H.; Feng, X.; Xu, H. Exogenous application of poly-γ-glutamic acid enhances stress defense in Brassica napus L. seedlings by inducing crosstalks between Ca2+, H2O2, brassinolide, and jasmonic acid in leaves. Plant Physiol. Biochem. 2017, 118, 460–470. [Google Scholar] [CrossRef]
- Ngearnpat, N.; Chunhachart, O.; Kotabin, N.; Issakul, K. Comparative assessment of gamma-polyglutamic acid and Bacillus subtilis cells as biostimulants to improve rice growth and soil quality. J. Ecol. Eng. 2023, 24, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Skalski, T.; Zając, E.; Jędrszczyk, E.; Papaj, K.; Kohyt, J.; Góra, A.; Kasprzycka, A.; Shytum, D.; Skowera, B.; Ziernicka-Wojtaszek, A. Effects of γ-polyglutamic acid on grassland sandy soil properties and plant functional traits exposed to drought stress. Sci. Rep. 2024, 14, 3769. [Google Scholar] [CrossRef]
- Xu, Z.; Ma, J.; Lei, P.; Wang, Q.; Feng, X.; Xu, H. Poly-γ-glutamic acid induces system tolerance to drought stress by promoting abscisic acid accumulation in Brassica napus L. Sci. Rep. 2020, 10, 252. [Google Scholar] [CrossRef]
- Wang, L.; Wang, N.; Mi, D.; Luo, Y.; Guo, J. Poly-γ-glutamic acid productivity of Bacillus subtilis BsE1 has positive function in motility and biocontrol against Fusarium graminearum. J. Microbiol. 2017, 55, 554–560. [Google Scholar] [CrossRef]
- Xu, Z.; Lei, P.; Feng, X.; Xu, X.; Liang, J.; Chi, B.; Xu, H. Calcium involved in the poly(γ-glutamic acid)-mediated promotion of Chinese cabbage nitrogen metabolism. Plant Physiol. Biochem. 2014, 80, 144–152. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, J.; Chen, T.; Zhang, H.; Hou, X.; Li, J. Effects of γ-polyglutamic acid supplementation on alfalfa growth and rhizosphere soil microorganisms in sandy soil. Sci. Rep. 2024, 14, 6440. [Google Scholar] [CrossRef]
- Tanimoto, H. Food applications of poly-gamma-glutamic acid. In Amino-Acid Homopolymers Occurring in Nature; Hamano, Y., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 155–168. [Google Scholar]
- Jeon, Y.O.; Lee, J.S.; Lee, H.G. Improving solubility, stability, and cellular uptake of resveratrol by nanoencapsulation with chitosan and γ-poly (glutamic acid). Colloids Surf. B Biointerfaces 2016, 147, 224–233. [Google Scholar] [CrossRef]
- Tanimoto, H.; Fox, T.; Eagles, J.; Satoh, H.; Nozawa, H.; Okiyama, A.; Morinaga, Y.; Susan, J.; Fairweather-Tait, S.J. Acute effect of poly-glutamic acid on calcium absorption in post-menopausal women. J. Am. Coll. Nutr. 2007, 26, 645–649. [Google Scholar] [CrossRef]
- Shyu, Y.S.; Hwang, J.Y.; Hsu, C.K. Improving the rheological and thermal properties of wheat dough by the addition of c-polyglutamic acid. LWT Food Sci. Technol. 2008, 41, 982–987. [Google Scholar] [CrossRef]
- Elbanna, K.; Alsulami, F.S.; Neyaz, L.A.; Abulreesh, H.H. Poly (γ) glutamic acid: A unique microbial biopolymer with diverse commercial applicability. Front. Microbiol. 2024, 15, 1348411. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Q.; Yang, Y.M.; Lee, S.U.; Han, J.S.; Jang, K.J.; Tong, T. Applications and func-tions of γ-poly-glutamic acid and its derivatives in medicine. Curr. Pharm. Biotechnol. 2021, 22, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Xu, Z.; Liang, J.; Luo, X.; Zhang, Y.; Feng, X. Poly(γ-glutamic acid) enhanced tolerance to salt stress by promoting proline accumulation in Brassica napus L. Plant Growth Regul. 2016, 78, 233–241. [Google Scholar] [CrossRef]
- Quan, J.; Zheng, W.; Tan, J.; Li, Z.; Wu, M.; Hong, S.B. Glutamic acid and poly-γ-glutamic acid enhanced the heat resistance of Chinese Cabbage (Brassica rapa L. ssp. pekinensis) by improving carotenoid biosynthesis, photosynthesis, and ROS signaling. Int. J. Mol. Sci. 2022, 23, 11671. [Google Scholar]
- Kapusta-Duch, J.; Kusznierewicz, B. Young shoots of white and red headed cabbages like novel sources of glucosinolates as well as antioxidative substances. Antioxidants 2021, 10, 1277. [Google Scholar] [CrossRef]
- Baenas, N.; Moreno, D.A.; García-Viguera, C. Selecting sprouts of Brassicaceae for optimum phytochemical composition. J. Agric. Food Chem. 2012, 60, 11409–11420. [Google Scholar] [CrossRef]
- Le, T.N.; Chiu, C.H.; Hsieh, P.C. Bioactive compounds and bioactivities of Brassica oleracea L. Var. Italica sprouts and microgreens: An updated overview from a nutraceutical perspective. Plants 2020, 9, 946. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. Acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of glucosinolates and their breakdown products: Impact of processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef]
- Ortega-Hernández, E.; Antunes-Ricardo, M.; Jacobo-Velázquez, D.A. Improving the health-benefits of kales (Brassica oleracea L. var. acephala DC) through the application of controlled abiotic stresses: A review. Plants 2021, 10, 2629. [Google Scholar] [CrossRef]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Technol. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]
- Bryant, L.; Rangan, A.; Grafenauer, S. Lupins and health outcomes: A systematic literature review. Nutrients 2022, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Estivi, L.; Brandolini, A.; Gasparini, A.; Hidalgo, A. Lupin as a source of bioactive antioxidant compounds for food products. Molecules 2023, 28, 7529. [Google Scholar] [CrossRef] [PubMed]
- Villarino, C.B.J.; Jayasena, V.; Coorey, R.; Chakrabarti-Bell, S.; Johnson, S.K. Nutritional, health, and technological functionality of lupin flour addition to bread and other baked products: Benefits and challenges. Crit. Rev. Food Sci. Nutr. 2015, 56, 835–857. [Google Scholar] [CrossRef] [PubMed]
- Galanty, A.; Zagrodzki, P.; Miret, M.; Paśko, P. Chickpea and lupin sprouts, stimulated by different led lights, as novel examples of isoflavones-rich functional food, and their impact on breast and prostate cells. Molecules 2022, 27, 9030. [Google Scholar] [CrossRef]
- Mahaboob Ali, A.A.; Momin, B.; Ghogare, P. Isolation of a novel poly-γ-glutamic acid-producing Bacillus licheniformis A14 strain and optimization of fermentation conditions for high-level production. Prep. Biochem. Biotech. 2020, 50, 445–452. [Google Scholar] [CrossRef]
- Yao, J.; Jing, J.; Xu, H.; Liang, J.; Wu, Q.; Feng, X.; Ouyang, P. Investigation on enzymatic degradation of γ-polyglutamic acid from Bacillus subtilis NX-J. Mol. Catal. B Enzym. 2009, 56, 158–164. [Google Scholar] [CrossRef]
- Galanty, A.; Juncewicz, P.; Podolak, I.; Grabowska, K.; Służały, P.; Paśko, P. Comparative analysis of polyphenolic profile and chemopreventive potential of hemp sprouts, leaves, and flowers of the Sofia variety. Plants 2024, 13, 2023. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pasko, P.; Gdula-Argasinska, J.; Podporska-Carroll, J.; Quilty, B.; Wietecha-Posluszny, R.; Tyszka-Czochara, M.; Zagrodzki, P. Influence of selenium supplementation on fatty acids profile and biological activity of four edible amaranth sprouts as new kind of functional food. J. Food Sci. Technol. 2015, 52, 4724–4736. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Paśko, P.; Galanty, A.; Zagrodzki, P.; Żmudzki, P.; Bieniek, U.; Prochownik, E. Varied effect of fortification of kale sprouts with novel organic selenium compounds on the synthesis of sulphur and phenolic compounds in relations to cytotoxic, antioxidant and anti-inflammatory activity. Microchem. J. 2022, 179, 107509. [Google Scholar] [CrossRef]
- Zagrodzki, P.; Paśko, P.; Galanty, A.; Tyszka-Czochara, M.; Wietecha-Posłuszny, R.; Salvans Rubio, P.; Bartoń, H.; Prochownik, E.; Muszyńska, B.; Sułkowska-Ziaja, K.; et al. Does selenium fortification of kale and kohlrabi sprouts change significantly their biochemical and cytotoxic properties. J. Trace Elem. Med. Biol. 2020, 59, 126466. [Google Scholar] [CrossRef] [PubMed]
- Luang-In, V.; Saengha, W.; Buranrat, B.; Chantiratikul, A.; Ma, N.L. Cytotoxicity of selenium-enriched Chinese kale (Brassica oleracea var. alboglabra L.) seedlings against Caco-2, MCF-7 and HepG2 cancer cells. Pharmacogn. J. 2020, 12, 674–681. [Google Scholar] [CrossRef]
- EL-Nahas, F.G.; M El Gezery, H.M. Effect of kale, lupine and its mixture sprouts on the biochemical parameters of hepatotoxicity rats with CCl4. Med. Environ. Sci. 2023, 18, 1957–1973. [Google Scholar] [CrossRef]
- Kestwal, R.M.; Lin, C.J.; Bagal-Kestwal, D.; Chiang, B.H. Glucosinolates fortification of cruciferous sprouts by sulphur supplementation during cultivation to enhance anti-cancer activity. Food Chem. 2011, 126, 1164–1171. [Google Scholar] [CrossRef]
- Frias, J.; Gulewicz, P.; Martínez-Villaluenga, C.; Pilarski, R.; Blazquez, E.; Jiménez, B.; Gulewicz, K.; Vidal-Valverde, C. Influence of germination with different selenium solutions on nutritional value and cytotoxicity of lupin seeds. J. Agric. Food Chem. 2009, 57, 1319–1325. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.; Liu, S.; Al-Mheiri, A.; Abushelaibi, A. Cytotoxicity, antihypertensive, antidiabetic and antioxidant activities of solid-state fermented lupin, quinoa and wheat by Bifidobacterium species: In-vitro investigations. LWT 2018, 95, 295–302. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Groups of Sprouts | Biochanin (µg/100 g dw) | Daidzin (µg/100 g dw) | Genistein (µg/100 g dw) | Genistin (mg/100 g dw) | Glycitein (µg/100 g dw) | Chlorogenic Acid (µg/100 g dw) | Protocatechuic Acid (µg/100 g dw) | Sinapic Acid (µg/100 g dw) |
|---|---|---|---|---|---|---|---|---|
| L1 | 43 ± 8 | 264 ± 4 a | 238 ± 8 a | 1.82 ± 0.02 ad | 271 ± 3 d | 679 ± 12 a | ND | ND |
| L2 | 43 ± 7 | 151 ± 14 a | 513 ± 3 a | 1.22 ± 0.01 ae | 216 ± 34 a | 803 ± 68 | ND | ND |
| L3 | 89 ± 19 | 203 ± 13 | 396 ± 26 | 1.38 ± 0.02 | 359 ± 13 a | 1465 ± 11 am | ND | ND |
| L4 | 37 ± 12 a | 140 ± 17 c | 224 ± 6 bd | 2.26 ± 0.03 b | 124 ± 6 bd | 755 ± 6 e,f | ND | ND |
| L5 | 34 ± 6 b | 108 ± 6 d | 512 ± 6 b | 1.46 ± 0.02 | 165 ± 12 e | 664 ± 5 bi,j | ND | ND |
| L6 | 49 ± 7 | 151 ± 65 | 411 ± 7 | 0.84 ± 0.02 b | 256 ± 7 b | 870 ± 20 bm | ND | ND |
| L7 | 58 ± 4 a | 302 ± 4 bc | 272 ± 18 cd | 2.93 ± 0.02 cd | 259 ± 13 c | 937 ± 13 cg,h | ND | ND |
| L8 | 79 ± 6 b | 270 ± 6 d | 422 ± 11 c | 2.33 ± 0.02 e | 299 ± 11 e | 1216 ± 6 ci,k,l | ND | ND |
| L9 | 68 ± 12 | 136 ± 12 b | 374 ± 59 | 0.85 ± 0.12 c | 354 ± 6 c | 1058 ± 16 | ND | ND |
| K1 | ND | ND | ND | ND | ND | 685 ± 12 | 204 ± 21 ad | 745 ± 21 ad |
| K2 | ND | ND | ND | ND | ND | 786 ± 30 n | 154 ± 8 ae | 1403 ± 29 ae |
| K3 | ND | ND | ND | ND | ND | 771 ± 15 k | 155 ± 8 | 1207 ± 15 |
| K4 | ND | ND | ND | ND | ND | 522 ± 45 e,g | 115 ± 8 | 937 ± 8 |
| K5 | ND | ND | ND | ND | ND | 545 ± 18 | 120 ± 9 b | 916 ± 9 b |
| K6 | ND | ND | ND | ND | ND | 889 ± 40 j | 197 ± 12 bf | 1458 ± 12 bf |
| K7 | ND | ND | ND | ND | ND | 510 ± 28 f,h | 131 ± 13 cd | 1080 ± 23 cd |
| K8 | ND | ND | ND | ND | ND | 395 ± 6 dn | 235 ± 13 ce | 757 ± 17 ce |
| K9 | ND | ND | ND | ND | ND | 797 ± 132 dl | 277 ± 18 f | 1050 ± 16 f |
| Tested Group of Sprouts | DPPH (μM TEAC/g dw) | FRAP (μM/Fe2+/g dw) | TRC (Polyphenols, µg/g dw) |
|---|---|---|---|
| L1 | 6.5 ± 0.2 ah,i | 14.5 ± 0.3 a,b | 7.8 ± 0.1 ae,f,g |
| L2 | 6.6 ± 0.6 bk | 14.3 ± 0.4 d,e | 8.5 ± 0.3 k,l,m |
| L3 | 12.4 ± 0.2 a,b | 25.2 ± 0.9 | 14 ± 0.6 ar |
| L4 | 7.0 ± 0.3 j | 17.9 ± 0.9 c | 9.8 ± 0.5 h,i |
| L5 | 6.5 ± 0.5 l,m | 14.7 ± 0.6 f | 9.4 ± 0.4 n,o |
| L6 | 6.6 ± 0.8 | 14.9 ± 0.7 | 9.9 ± 0.2 r |
| L7 | 10.2 ± 0.2 | 25.0 ± 0.5 | 12 ± 0.7 j |
| L8 | 11.4 ± 0.3 | 25.2 ± 0.3 | 12.9 ± 0.8 p |
| L9 | 10.6 ± 0.1 | 19.3 ± 0.5 | 11.7 ± 0.2 |
| K1 | 38.0 ± 0.5 a,b | 102.2 ± 0.7 a | 13.5 ± 0.3 be |
| K2 | 61.1 ± 0.2 a,c | 151.8 ± 0.6 | 20.5 ± 1.1 bs |
| K3 | 54.3 ± 0.3 b,cl | 157.7 ± 0.8 d | 19.1 ± 1.4 k,n |
| K4 | 39.8 ± 0.5 dh | 97.2 ± 3.0 | 14.6 ± 1.0 cf,h |
| K5 | 43.2 ± 0.2 e | 105.3 ± 1.9 | 15.7 ± 0.8 g,i,j |
| K6 | 63.6 ± 0.4 d,ek,m | 181.8 ± 2.1 e,f | 23.3 ± 0.9 cl,o,p |
| K7 | 44.2 ± 0.6 fi,j | 108.4 ± 1.2 b,c | 15.7 ± 0.8 |
| K8 | 31.5 ± 0.9 f,g | 74.8 ± 0.4 | 11.4 ± 0.4 ds |
| K9 | 48.4 ± 0.1 g | 117.6 ± 0.1 | 16.5 ± 0.3 dm |
| Pairs of Correlated Parameters | Correlation Coefficients | |
|---|---|---|
| Chlorogenic acid | HT29 | 1.000 |
| Biochanin | Glycitein | 0.999 |
| DPPH | FRAP | 0.999 |
| Genistein | HCT116 | 0.990 |
| FRAP | TRC | 0.974 |
| Biochanin | Genistin | 0.970 |
| Daidzin | Genistin | 0.962 |
| DPPH | Sinapic acid | 0.962 |
| Genistin | Glycitein | 0.961 |
| DPPH | TRC | 0.960 |
| Daidzin | HT29 | 0.949 |
| Daidzin | Chlorogenic acid | 0.948 |
| FRAP | Sinapic acid | 0.946 |
| Biochanin | Daidzin | 0.868 |
| Daidzin | Glycitein | 0.850 |
| TRC | Sinapic acid | 0.847 |
| Glycitein | Sinapic acid | −0.706 |
| TRC | HCT116 | −0.994 |
| TRC | Genistein | −1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galanty, A.; Kłos, P.; Prochownik, E.; Paśko, P.; Skalski, T.; Podsiadły, R.; Zagrodzki, P. Cytotoxic and Antioxidant Properties and Profile of Active Compounds in Kale and Lupine Sprouts Supplemented with γ-Polyglutamic Acid During Sprouting. Appl. Sci. 2025, 15, 2813. https://doi.org/10.3390/app15052813
Galanty A, Kłos P, Prochownik E, Paśko P, Skalski T, Podsiadły R, Zagrodzki P. Cytotoxic and Antioxidant Properties and Profile of Active Compounds in Kale and Lupine Sprouts Supplemented with γ-Polyglutamic Acid During Sprouting. Applied Sciences. 2025; 15(5):2813. https://doi.org/10.3390/app15052813
Chicago/Turabian StyleGalanty, Agnieszka, Paulina Kłos, Ewelina Prochownik, Paweł Paśko, Tomasz Skalski, Robert Podsiadły, and Paweł Zagrodzki. 2025. "Cytotoxic and Antioxidant Properties and Profile of Active Compounds in Kale and Lupine Sprouts Supplemented with γ-Polyglutamic Acid During Sprouting" Applied Sciences 15, no. 5: 2813. https://doi.org/10.3390/app15052813
APA StyleGalanty, A., Kłos, P., Prochownik, E., Paśko, P., Skalski, T., Podsiadły, R., & Zagrodzki, P. (2025). Cytotoxic and Antioxidant Properties and Profile of Active Compounds in Kale and Lupine Sprouts Supplemented with γ-Polyglutamic Acid During Sprouting. Applied Sciences, 15(5), 2813. https://doi.org/10.3390/app15052813