
A Simple High-Throughput Procedure for Microscale Extraction of Bioactive Compounds from the Flowers of Saint John’s Wort (Hypericum perforatum L.)

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Determination of Flower Fresh Weight
2.3. Collection and Extraction of a Bulk Flower Sample
2.4. Collection and Extraction of Single Flower Samples
2.5. LC/DAD/MS Analysis
2.6. LC/UV-Vis Analysis
2.7. Statistical Analysis
3. Results
3.1. Selection of the Stage of Flower Development for Analysis
3.2. Bulk-Flower Extraction and LC-DAD-QTOF Analysis
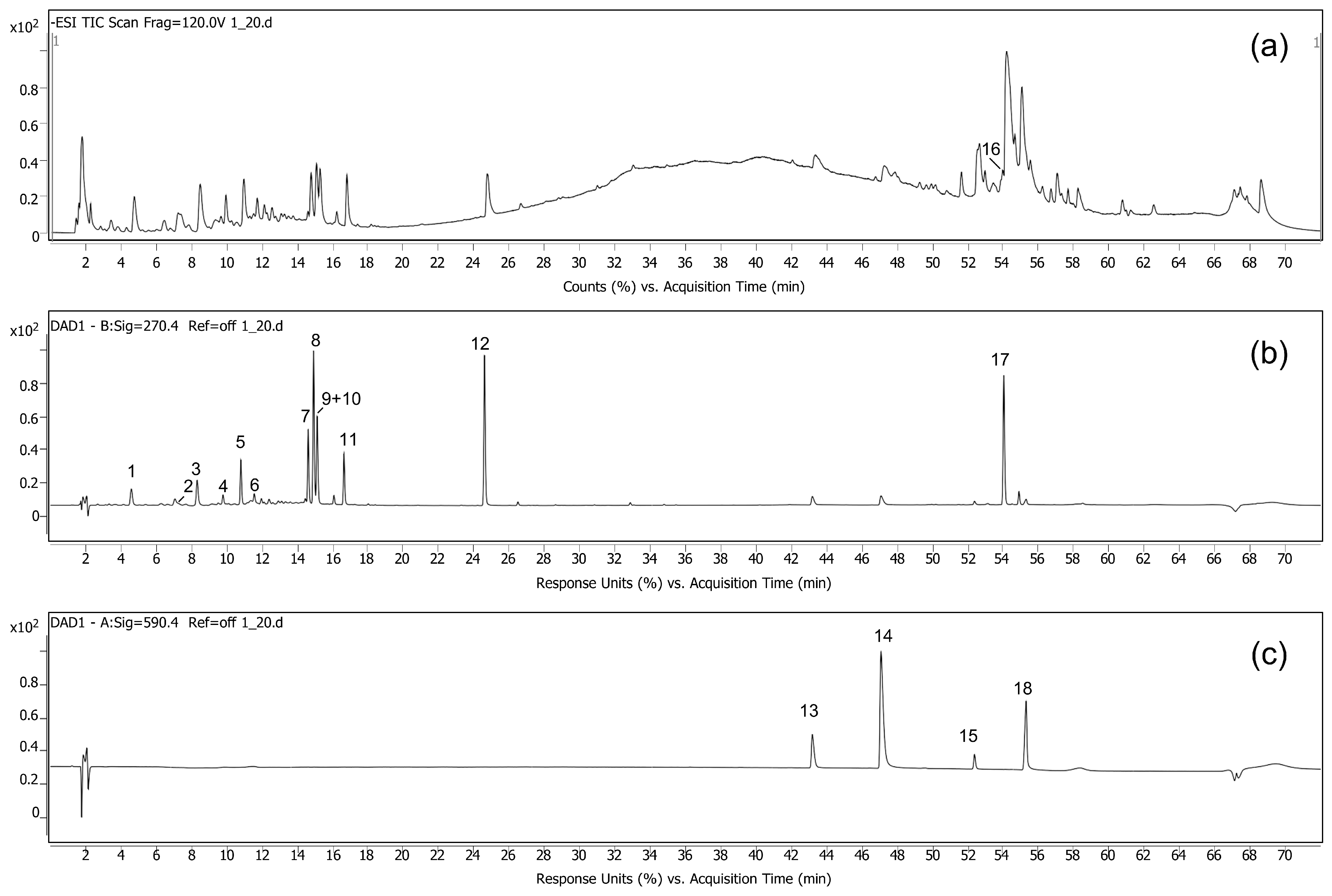
3.2.1. LC-DAD-QTOF Analysis and Identification of the Compounds
3.2.2. Testing Different Material-to-Solvent Ratios
3.2.3. Testing Different Durations of the Extraction
3.3. Single-Flower Extraction
3.4. Analysis of Plants Obtained Through Apomixis Using the “Single-Flower” Procedure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SSR | Simple sequence repeat |
| LC-MS | Liquid chromatography–mass spectrometry |
| DAD | Diode array detector |
| QTOF | Quadrupole time of flight |
| ESI | Electrospray ionization |
| TIC | Total ion current |
| RSD | Relative standard deviation |
| UV-Vis | Ultraviolet–visible |
References
- Hunt, E.J.; Lester, C.E.; Lester, E.A.; Tackett, R.L. Effect of St. John’s wort on free radical production. Life Sci. 2001, 69, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Franchi, G.G.; Nencini, C.; Collavoli, E.; Massarelli, P. Composition and antioxidant activity in vitro of different St. John’s Wort (Hypericum perforatum L.) extracts. J. Med. Plants Res. 2011, 5, 4349–4353. [Google Scholar]
- Muzykiewicz, A.; Florkowska, K.; Nowak, A.; Zielonka-Brzezicka, J.; Klimowicz, A. Antioxidant activity of St. John’s Wort extracts obtained with ultrasound-assisted extraction. Pomeranian J. Life Sci. 2019, 65, 89–93. [Google Scholar] [CrossRef]
- Avato, P.; Raffo, F.; Guglielmi, G.; Vitali, C.; Rosato, A. Extracts from St John’s wort and their antimicrobial activity. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2004, 18, 230–232. [Google Scholar] [CrossRef]
- Schempp, C.M.; Pelz, K.; Wittmer, A.; Schöpf, E.; Simon, J.C. Antibacterial activity of hyperforin from St John’s wort, against multiresistant Staphylococcus aureus and gram-positive bacteria. Lancet 1999, 353, 2129. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. St John’s wort (Hypericum perforatum L.): A review of its chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2001, 53, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.M.; Feinman, L.; Liebes, L.; Ostrow, N.; Koslowski, V.; Tobia, A.; Cabana, B.E.; Lee, D.-H.; Spritzler, J.; Prince, A.M. Pharmacokinetics, safety, and antiviral effects of hypericin, a derivative of St. John’s wort plant, in patients with chronic hepatitis C virus infection. Antimicrob. Agents Chemother. 2001, 45, 517–524. [Google Scholar] [CrossRef]
- Gulick, R.M.; McAuliffe, V.; Holden-Wiltse, J.; Crumpacker, C.; Liebes, L.; Stein, D.S.; Meehan, P.; Hussey, S.; Forcht, J.; Valentine, F.T. Phase I studies of hypericin, the active compound in St. John’s wort, as an antiretroviral agent in HIV-infected adults: AIDS clinical trials group protocols 150 and 258. Ann. Intern. Med. 1999, 130, 510–514. [Google Scholar] [CrossRef]
- Linde, K.; Ramirez, G.; Mulrow, C.D.; Pauls, A.; Weidenhammer, W.; Melchart, D. St John’s wort for depression—An overview and meta-analysis of randomised clinical trials. BMJ 1996, 313, 253–258. [Google Scholar] [CrossRef]
- Bilia, A.R.; Gallori, S.; Vincieri, F.F. St. John’s wort and depression: Efficacy, safety and tolerability-an update. Life Sci. 2002, 70, 3077–3096. [Google Scholar] [CrossRef]
- Gaster, B.; Holroyd, J. St John’s wort for depression: A systematic review. Arch. Intern. Med. 2000, 160, 152–156. [Google Scholar] [CrossRef]
- Tedeschi, E.; Menegazzi, M.; Margotto, D.; Suzuki, H.; Förstermann, U.; Kleinert, H. Anti-inflammatory actions of St. John’s wort: Inhibition of human inducible nitric-oxide synthase expression by down-regulating signal transducer and activator of transcription-1α (STAT-1α) activation. J. Pharmacol. Exp. Ther. 2003, 307, 254–261. [Google Scholar] [CrossRef]
- Novelli, M.; Masiello, P.; Beffy, P.; Menegazzi, M. Protective role of St. John’s wort and its components hyperforin and hypericin against diabetes through inhibition of inflammatory signaling: Evidence from in vitro and in vivo studies. Int. J. Mol. Sci. 2020, 21, 8108. [Google Scholar] [CrossRef] [PubMed]
- Schempp, C.M.; Kirkin, V.; Simon-Haarhaus, B.; Kersten, A.; Kiss, J.; Termeer, C.C.; Gilb, B.; Kaufmann, T.; Borner, C.; Sleeman, J.P. Inhibition of tumour cell growth by hyperforin, a novel anticancer drug from St. John’s wort that acts by induction of apoptosis. Oncogene 2002, 21, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Krysko, D.V.; Vandenabeele, P.; Agostinis, P. Hypericin-based photodynamic therapy induces surface exposure of damage-associated molecular patterns like HSP70 and calreticulin. Cancer Immunol. Immunother. 2012, 61, 215–221. [Google Scholar] [CrossRef]
- Tekel’ová, D.; Repák, M.; Zemková, E.; Tóth, J. Quantitative Changes of Dianthrones, Hyperforin and Flavonoids Content in the Flower Ontogenesis of Hypericum perforatum. Planta Medica 2000, 66, 778–780. [Google Scholar] [CrossRef]
- Galeotti, N. Hypericum perforatum (St John’s wort) beyond depression: A therapeutic perspective for pain conditions. J. Ethnopharmacol. 2017, 200, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Rossi, A.; Bauer, J.; Dehm, F.; Verotta, L.; Northoff, H.; Sautebin, L.; Werz, O. Hyperforin, an anti-inflammatory constituent from St. John’s wort, inhibits microsomal prostaglandin E2 synthase-1 and suppresses prostaglandin E2 formation in vivo. Front. Pharmacol. 2011, 2, 7. [Google Scholar] [CrossRef]
- Hofrichter, J.; Krohn, M.; Schumacher, T.; Lange, C.; Feistel, B.; Walbroel, B.; Heinze, H.-J.; Crockett, S.; Sharbel, T.F.; Pahnke, J. Reduced Alzheimer’s disease pathology by St. John’s Wort treatment is independent of hyperforin and facilitated by ABCC1 and microglia activation in mice. Curr. Alzheimer Res. 2013, 10, 1057–1069. [Google Scholar] [CrossRef]
- Shabani, A.; Karapandzova, M.; Karanfilova, I.C.; Stefkov, G.; Crcarevska, M.S.; Kulevanova, S. Distribution of total phenols, flavonoids and hypericin in different plant organs of wild-growing St. John’s-wort (Hypericum perforatum L., Hypericaceae) from North Macedonia. Maced. Pharm. Bull. 2019, 65, 39–47. [Google Scholar] [CrossRef]
- Umek, A.; Kreft, S.; Kartnig, T.; Heydel, B. Quantitative phytochemical analyses of six Hypericum species growing in Slovenia. Planta Medica 1999, 65, 388–390. [Google Scholar] [CrossRef]
- Çirak, C.; Radusiene, J.; Janulis, V.; Ivanauskas, L. Pseudohypericin and hyperforin in Hypericum perforatum from Northern Turkey: Variation among populations, plant parts and phenological stages. J. Integr. Plant Biol. 2008, 50, 575–580. [Google Scholar] [CrossRef]
- Cirak, C.; Radusiěnë, J.; Karabük, B.; Janulis, V.; Ivanauskas, L. Variation of bioactive compounds in Hypericum perforatum growing in Turkey during its phenological cycle. J. Integr. Plant Biol. 2007, 49, 615–620. [Google Scholar] [CrossRef]
- Çirak, C.; Radušienė, J.; Karabük, B.S.; Janulis, V. Variation of bioactive substances and morphological traits in Hypericum perforatum populations from Northern Turkey. Biochem. Syst. Ecol. 2007, 35, 403–409. [Google Scholar] [CrossRef]
- Klemow, K.M.; Bilbow, E.; Grasso, D.; Jones, K.; McDermott, J.; Pape, E. Medical attributes of St. John’s wort (Hypericum perforatum). Oxidative Stress Dis. 2004, 14, 757–780. [Google Scholar]
- Zagorcheva, T.; Rusanov, K.; Rusanova, M.; Aneva, I.; Stancheva, I.; Atanassov, I. Genetic and flower volatile diversity in two natural populations of Hyssopus officinalis L. in Bulgaria. Biotechnol. Biotechnol. Equip. 2020, 34, 1265–1272. [Google Scholar] [CrossRef]
- Rusanov, K.E.; Kovacheva, N.M.; Atanassov, I.I. Comparative GC/MS analysis of rose flower and distilled oil volatiles of the oil-bearing rose Rosa damascena. Biotechnol. Biotechnol. Equip. 2011, 25, 2210–2216. [Google Scholar] [CrossRef]
- Zagorcheva, T.; Stanev, S.; Rusanov, K.; Atanassov, I. Comparative GC/MS analysis of lavender (Lavandula angustifolia Mill.) inflorescence and essential oil volatiles. Agric. Sci. Technol. 2013, 5, 459–462. [Google Scholar]
- Zagorcheva, T.; Rusanov, K.; Stanev, S.; Atanassov, I. A simple procedure for comparative GC-MS analysis of lavender (Lavandula angustifolia Mill.) flower volatile composition. IOSR J. Pharm. Biol. Sci. 2016, 11, 9–14. [Google Scholar] [CrossRef]
- Alekseeva, M.; Zagorcheva, T.; Rusanova, M.; Rusanov, K.; Atanassov, I. Genetic and flower volatile diversity in natural populations of Origanum vulgare subsp. hirtum (Link) Ietsw. in Bulgaria: Toward the development of a core collection. Front. Plant Sci. 2021, 12, 679063. [Google Scholar] [CrossRef]
- Bagdonaitė, E.; Mártonfi, P.; Repčák, M.; Labokas, J. Variation in concentrations of major bioactive compounds in Hypericum perforatum L. from Lithuania. Ind. Crops Prod. 2012, 35, 302–308. [Google Scholar] [CrossRef]
- Koperdáková, J.; Košuth, J.; Čellárová, E. Variation in the content of hypericins in four generations of seed progeny of Hypericum perforatum somaclones. J. Plant Res. 2007, 120, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Xenophontos, M.; Stavropoulos, E.; Avramakis, E.; Navakoudis, E.; Dörnemann, D.; Kotzabasis, K. Influence of the developmental stage on the (proto)-hypericin and (proto) pseudohypericin levels of Hypericum plants from Crete. Planta Medica 2007, 73, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Smelcerovic, A.; Zuehlke, S.; Hussain, M.A.; Ahmad, S.M.; Ziebach, T.; Qazi, G.N.; Spiteller, M. Phenolic constituents and genetic profile of Hypericum perforatum L. from India. Biochem. Syst. Ecol. 2008, 36, 201–206. [Google Scholar] [CrossRef]
- Kaplan, M.; Köprü, S.; Say, R.; Karaman, K.; Yılmaz, M.M. Characterization of in vitro bioactive performance of Hypericum perforatum using response surface methodology. Sigma J. Eng. Nat. Sci. 2021, 39, 392–403. [Google Scholar] [CrossRef]
- Frommenwiler, D.A.; Reich, E.; Sudberg, S.; Sharaf, M.H.; Bzhelyansky, A.; Lucas, B. St. John’s wort versus counterfeit St. John’s wort: An HPTLC study. J. AOAC Int. 2016, 99, 1204–1212. [Google Scholar] [CrossRef]
- Huck-Pezzei, V.; Bittner, L.; Pallua, J.; Sonderegger, H.; Abel, G.; Popp, M.; Bonn, G.; Huck, C. A chromatographic and spectroscopic analytical platform for the characterization of St John’s wort extract adulterations. Anal. Methods 2013, 5, 616–628. [Google Scholar] [CrossRef]
- Wuthold, K.; Roos, G.; Simmen, U.; Kovar, K.-A. Analytical study of extracts of St John’s wort (Hypericum perforatum), evaluation of HPTLC plates by multivariate data analysis. JPC-J. Planar Chromatogr.-Mod. TLC 2003, 16, 15–18. [Google Scholar] [CrossRef]
- Kitanov, G.M. Hypericin and pseudohypericin in some Hypericum species. Biochem. Syst. Ecol. 2001, 29, 171–178. [Google Scholar] [CrossRef]
- Maleš, Ž.; Plazibat, M.; Vundać, V.; Žuntar, I.; Pilepić, K. Thin-layer chromatographic analysis of flavonoids, phenolic acids, and amino acids in some Croatian Hypericum taxa. JPC-J. Planar Chromatogr.-Mod. TLC 2004, 17, 280–285. [Google Scholar] [CrossRef]
- Mulinacci, N.; Bardazzi, C.; Romani, A.; Pinelli, P.; Vincieri, F.F.; Costantini, A. HPLC-DAD and TLC-Densitometry for quantification of hypericin in Hypericum perforatum L. Extracts. Chromatographia 1999, 49, 197–201. [Google Scholar] [CrossRef]
- Tolonen, A.; Hohtola, A.; Jalonen, J. Fast high-performance liquid chromatographic analysis of naphthodianthrones and phloroglucinols from Hypericum perforatum extracts. Phytochem. Anal. 2003, 14, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Ang, C.Y.; Cui, Y.; Chang, H.C.; Luo, W.; Heinze, T.M.; Lin, L.J.; Mattia, A. Determination of St. John’s wort components in dietary supplements and functional foods by liquid chromatography. J. AOAC Int. 2002, 85, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Agapouda, A.; Booker, A.; Kiss, T.; Hohmann, J.; Heinrich, M.; Csupor, D. Quality control of Hypericum perforatum L. analytical challenges and recent progress. J. Pharm. Pharmacol. 2019, 71, 15–37. [Google Scholar] [CrossRef]
- Brolis, M.; Gabetta, B.; Fuzzati, N.; Pace, R.; Panzeri, F.; Peterlongo, F. Identification by high-performance liquid chromatography–diode array detection–mass spectrometry and quantification by high-performance liquid chromatography–UV absorbance detection of active constituents of Hypericum perforatum. J. Chromatogr. A 1998, 825, 9–16. [Google Scholar] [CrossRef]
- Mauri, P.; Pietta, P. High performance liquid chromatography/electrospray mass spectrometry of Hypericum perforatum extracts. Rapid Commun. Mass. Spectrom. 2000, 14, 95–99. [Google Scholar] [CrossRef]
- Chandrasekera, D.H.; Welham, K.J.; Ashton, D.; Middleton, R.; Heinrich, M. Quantitative analysis of the major constituents of St John’s wort with HPLC-ESI-MS. J. Pharm. Pharmacol. 2005, 57, 1645–1652. [Google Scholar] [CrossRef]
- Keller, J.H.; Karas, M.; Müller, W.E.; Volmer, D.A.; Eckert, G.P.; Tawab, M.A.; Blume, H.H.; Dingermann, T.; Schubert-Zsilavecz, M. Determination of hyperforin in mouse brain by high-performance liquid chromatography/tandem mass spectrometry. Anal. Chem. 2003, 75, 6084–6088. [Google Scholar] [CrossRef]
- Matzk, F.; Meister, A.; Brutovská, R.; Schubert, I. Reconstruction of reproductive diversity in Hypericum perforatum L. opens novel strategies to manage apomixis. Plant J. 2001, 26, 275–282. [Google Scholar] [CrossRef]
- Velkoska-Markovska, L.; Jankulovska, M.S.; Petanovska-Ilievska, B.; Hristovski, K. Development and validation of RRLC–UV method for determination of chlorogenic acid in green coffee. Acta Chromatogr. 2020, 32, 34–38. [Google Scholar] [CrossRef]
- Sun, L.; Wei, H.; Zhang, F.; Gao, S.; Zeng, Q.; Lu, W.; Chen, W.; Chai, Y. Qualitative analysis and quality control of Traditional Chinese Medicine preparation Tanreqing injection by LC-TOF/MS and HPLC-DAD-ELSD. Anal. Methods 2013, 5, 6431–6440. [Google Scholar] [CrossRef]
- Wang, L.; Yamashita, Y.; Saito, A.; Ashida, H. An analysis method for flavan-3-ols using high performance liquid chromatography coupled with a fluorescence detector. J. Food Drug Anal. 2017, 25, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Wessjohann, L.A. Metabolome classification of commercial Hypericum perforatum (St. John’s Wort) preparations via UPLC-qTOF-MS and chemometrics. Planta Medica 2012, 78, 488–496. [Google Scholar] [CrossRef]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L. HMDB 5.0: The human metabolome database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Cioffi, E.; Comune, L.; Piccolella, S.; Buono, M.; Pacifico, S. Quercetin 3-O-glucuronide from Aglianico vine leaves: A selective sustainable recovery and accumulation monitoring. Foods 2023, 12, 2646. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2025 update. Nucleic Acids Res. 2024, 53, D1516–D1525. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Tusevski, O.; Krstikj, M.; Petreska Stanoeva, J.; Stefova, M.; Gadzovska Simic, S. Phenolic compounds composition of Hypericum perforatum L. wild-growing plants from the Republic of Macedonia. Agric. Conspec. Sci. 2019, 84, 67–75. [Google Scholar]
- Heinrich, M.; Daniels, R.; Stintzing, F.; Kammerer, D. Comprehensive phytochemical characterization of St. John’s wort (Hypericum perforatum L.) oil macerates obtained by different extraction protocols via analytical tools applicable in routine control. Die Pharm.-Int. J. Pharm. Sci. 2017, 72, 131–138. [Google Scholar] [CrossRef]
- Lyles, J.T.; Kim, A.; Nelson, K.; Bullard-Roberts, A.L.; Hajdari, A.; Mustafa, B.; Quave, C.L. The chemical and antibacterial evaluation of St. John’s Wort oil macerates used in Kosovar traditional medicine. Front. Microbiol. 2017, 8, 1639. [Google Scholar] [CrossRef] [PubMed]
- Bruni, R.; Sacchetti, G. Factors affecting polyphenol biosynthesis in wild and field grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682–725. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, P.; Altschmied, L.; Ravindran, B.M.; Rutten, T.; D’Auria, J.C. The biochemical and genetic basis for the biosynthesis of bioactive compounds in Hypericum perforatum L., one of the largest medicinal crops in Europe. Genes 2020, 11, 1210. [Google Scholar] [CrossRef]
- Filippini, R.; Piovan, A.; Borsarini, A.; Caniato, R. Study of dynamic accumulation of secondary metabolites in three subspecies of Hypericum perforatum. Fitoterapia 2010, 81, 115–119. [Google Scholar] [CrossRef]
- Kladar, N.; Mrđanović, J.; Anačkov, G.; Šolajić, S.; Gavarić, N.; Srđenović, B.; Božin, B. Hypericum perforatum: Synthesis of active principles during flowering and fruitification—Novel aspects of biological potential. Evid.-Based Complement. Altern. Med. 2017, 2017, 2865610. [Google Scholar] [CrossRef]
- Ayan, A.K.; Çirak, C. Hypericin and Pseudohypericin contents in some Hypericum. species growing in Turkey. Pharm. Biol. 2008, 46, 288–291. [Google Scholar] [CrossRef]
- Couceiro, M.; Afreen, F.; Zobayed, S.; Kozai, T. Variation in concentrations of major bioactive compounds of St. John’s wort: Effects of harvesting time, temperature and germplasm. Plant Sci. 2006, 170, 128–134. [Google Scholar] [CrossRef]
- Baugh, S.F. Simultaneous Determination of Protopseudohypericin, Pseudohypericin, Protohypericin, and Hypericin Without Light Exposure. J. AOAC Int. 2005, 88, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Poutaraud, A.; Di Gregorio, F.; Tin, V.C.; Girardin, P. Effect of light on hypericins contents in fresh flowering top parts and in an extract of St. John’s Wort (Hypericum perforatum). Planta Medica 2001, 67, 254–259. [Google Scholar] [CrossRef]
- Kopleman, S.H.; NguyenPho, A.; Zito, W.S.; Muller, F.X.; Augsburger, L.L. Selected physical and chemical properties of commercial Hypericum perforatum extracts relevant for formulated product quality and performance. AAPS PharmSci 2001, 3, 26. [Google Scholar] [CrossRef]
- Rusanov, K.; Vassileva, P.; Rusanova, M.; Atanassov, I. Identification of QTL controlling the ratio of linalool to linalyl acetate in the flowers of Lavandula angustifolia Mill var. Hemus. Biotechnol. Biotechnol. Equip. 2023, 37, 2288929. [Google Scholar] [CrossRef]
- Georgieva, P.; Rusanov, K.; Rusanova, M.; Kitanova, M.; Atanassov, I. Construction of Simple Sequence Repeat-Based Genetic Linkage Map and Identification of QTLs for Accumulation of Floral Volatiles in Lavender (Lavandula angustifolia Mill.). Int. J. Mol. Sci. 2025, 26, 3705. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Flower Stage | Plant 3_29 | Plant 90_44 | Plant 139_28 | Across Genotypes | ||||
|---|---|---|---|---|---|---|---|---|
| Mean FW | RSD | Mean FW | RSD | Mean FW | RSD | Mean FW | RSD | |
| 1st | 15.14 | 4.83 | 23.24 | 12.05 | 19.04 | 3.86 | 19.14 ± 4.05 a | 21.16 |
| 2nd | 21.79 | 4.66 | 41.92 | 14.58 | 34.16 | 7.93 | 32.62 ± 10.15 a | 31.13 |
| 3rd | 47.36 | 2.36 | 53.63 | 1.24 | 52.58 | 6.02 | 51.19 ± 3.36 b | 6.57 |
| 4th | 62.27 | 5.96 | 66.91 | 13.90 | 65.23 | 4.30 | 64.80 ± 2.35 b | 3.63 |
| 5th | 43.48 | 4.32 | 48.92 | 2.43 | 60.08 | 4.31 | 50.83 ± 8.46 b | 16.65 |
| Compound | tR min | Identity | Trivial Name | UV λ Max nm | [M−H]− m/z | Δppm | MS2 Fragments | Reference |
|---|---|---|---|---|---|---|---|---|
| 1 * | 5.1 | caffeoylquinic acid | chlorogenic acid | 218, 300, 325 | 353.0885 | 1.96 | 191.06, 179.03, 135.05 | [50,51] |
| 2 | 7.3 | procyanidin B1 | - | 279 | 577.13583 | 1.17 | 425.09, 407.08, 289.07, 125.02 | [52,53] |
| 3 * | 8.8 | catechin | - | 278 | 289.0721 | 1.16 | 245.08, 205.05, 125.02 | [54] |
| 4 * | 10.2 | procyanidin B2 | - | 280 | 577.1357 | 0.94 | 425.09, 407.08, 289.07, 125.02 | [52,53] |
| 5 * | 11.2 | epicatechin | - | 279 | 289.0720 | 0.82 | 245.08, 205.05, 125.02 | [54] |
| 6 * | 11.9 | procyanidin C1 | - | 280 | 865.1987 | 0.19 | 695.14, 577.13, 407.08, 287.06, 125.02 | [55] |
| 7 * | 14.8 | quercetin-3-O-rutinoside | rutin | 257, 358 | 609.1472 | 1.79 | 300.03, 271.02, 255.03, 179.00, 151.00 | [54] |
| 8 * | 15.2 | quercetin-3-O-galactoside | hyperoside | 255, 355 | 463.0892 | 2.15 | 300.03, 271.02, 255.03, 179.00, 151.00 | [54] |
| 9 ** | 15.4 | quercetin-3-O-glucoronide | querciturone | 255, 355 | 477.0683 | 1.75 | 301.03, 179.00, 151.00 | [56] |
| 10 ** | 15.5 | quercetin-3-O-glucoside | isoquercitrin | 255, 355 | 463.0889 | 1.5 | 300.03, 271.03, 255.03 | [57,58] |
| 11 * | 16.9 | quercetin-3-O-rhamnoside | quercitrin | 255, 345 | 447.0941 | 1.81 | 300.03, 271.02, 255.03, 179.00, 151.00 | [58] |
| 12 * | 24.9 | I3, II8-biapigenin | amentoflavone | 268, 330 | 537.0834 | 1.26 | 443.04, 385.07, 151.00 | [53] |
| 13 * | 43.0 | protopseudohypericin | - | - | 521.0875 | −0.59 | 477.10, 423.09 | [59] |
| 14 * | 47.1 | pseudohypericin | - | 592 | 519.0722 | 0.08 | 487.05, 475.08, 503.04, 449.07, 421.07 | [59] |
| 15 * | 52.1 | protohypericin | - | - | 505.0939 | 1.99 | 461.10, 407.09 | [60] |
| 16 | 54.0 | furohyperforin | - | - | 551.3733 | −1.63 | 411.25, 482.30, 329.18 | [61] |
| 17 * | 54.7 | hyperforin | - | 223, 272 | 535.3789 | −0.72 | 466.11, 398.24, 383.22, 315.16 | [61] |
| 18 * | 55.54 | hypericin | - | 592 | 503.0773 | 0.12 | 459.09, 487.04, 443.07, 405.08 | [61] |
| 3_29 | 90_44 | 139_28 | Mean RSD (%) per Compound | ||||
|---|---|---|---|---|---|---|---|
| Compound | Mean (%) | RSD | Mean (%) | RSD | Mean (%) | RSD | |
| 1 | 3.18 ± 0.40 b | 12.70 | 1.82 ± 0.2 a | 11.03 | 2.08 ± 0.08 a | 3.71 | 9.15 ± 4.78 |
| 3 | 4.15 ± 0.28 a | 6.83 | 1.45 ± 0.08 b | 5.53 | 2.81 ± 0.19 c | 6.86 | 6.41 ± 0.76 |
| 4 | 1.16 ± 0.0.14 a | 12.10 | 1.42 ± 0.04 b | 3.15 | 1.17 ± 0.06 a | 4.95 | 6.73 ± 4.73 |
| 5 | 3.88 ± 0.35 a | 3.09 | 5.34 ± 0.3 b | 5.69 | 4.51 ± 0.23 c | 5.01 | 4.60 ± 1.35 |
| 6 | 1.04 ± 0.03 a | 3.13 | 0.82 ± 0.01 b | 0.90 | 0.93 ± 0.05 c | 5.75 | 3.26 ± 2.43 |
| 7 | 13.37 ± 0.90 a | 6.73 | 0.08 ± 0.07 b | 7.83 | 4.78 ± 0.38 c | 7.99 | 7.52 ± 0.69 |
| 8 | 13.24 ± 0.75 a | 5.67 | 21.14 ± 1.73 b | 8.25 | 14.54 ± 0.85 a | 5.82 | 6.58 ± 1.45 |
| 9+10 | 8.26 ± 0.91 a | 11.03 | 14.80 ± 0.68 b | 4.62 | 10.52 ± 0.85 c | 8.05 | 7.90 ± 3.21 |
| 11 | 5.41 ± 0.58 a | 10.69 | 5.90 ± 0.44 a | 7.62 | 5.55 ± 0.65 a | 11.72 | 10.01 ± 2.13 |
| 12 | 11.26 ± 1.74 b | 15.45 | 7.78± 0.67 a | 8.62 | 9.68 ± 1.48 ab | 15.27 | 13.11 ± 3.89 |
| 13 | 0.97 ± 0.10 a | 10.37 | 0.56 ± 0.13 a | 22.97 | 0.86 ± 0.3 a | 34.92 | 22.75 ± 12.28 |
| 14 | 6.60 ± 0.71 a | 10.81 | 8.17 ± 0.89 a | 10.98 | 7.38 ± 0.66 a | 8.94 | 10.24 ± 1.13 |
| 15 | 0.41 ± 0.05 ab | 11.34 | 0.55 ±0.15 b | 27.06 | 0.29 ± 0.1 a | 36.51 | 24.97 ± 12.71 |
| 17 | 23.38 ± 3.04 a | 12.99 | 21.94 ± 2.94 a | 13.53 | 30.93 ± 2.42 b | 7.83 | 11.45 ± 3.15 |
| 18 | 3.68 ± 0.50 a | 13.65 | 8.21 ± 0.94 b | 11.53 | 3.98 ± 0.25 a | 6.17 | 10.45 ± 3.86 |
| Mean RSD per plant | 9.77 ± 3.79 | 9.95 ± 7.01 | 11.30 ± 10.33 | 10.34 ± 6.08 | |||
| 13+14 * | 7.58 ± 0.81 a | 10.74 | 8.74 ± 0.86 a | 9.80 | 7.38 ± 0.66 a | 8.94 | 9.66 |
| 15+18 * | 4.10 ± 0.55 a | 13.35 | 8.76 ± 0.92 b | 10.52 | 8.24 ± 0.53 b | 6.38 | 9.33 |
| Compound | Apomictic Line Hp93 | Three Independent Genotypes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MEAN | RSD | MIN | MAX | MIN/MEAN | MAX/MEAN | MEAN | RSD | MIN | MAX | MIN/MEAN | MAX/MEAN | |
| 1 | 2.06 | 8.01 | 1.79 | 2.34 | 86.67 | 113.55 | 2.36 | 30.49 | 1.82 | 3.18 | 77.22 | 134.64 |
| 3 | 1.38 | 8.08 | 1.19 | 1.64 | 86.81 | 119.04 | 2.80 | 48.07 | 1.45 | 4.15 | 51.77 | 147.91 |
| 4 | 1.77 | 8.80 | 1.48 | 2.20 | 83.71 | 124.71 | 1.25 | 11.77 | 1.16 | 1.42 | 92.63 | 113.57 |
| 5 | 5.00 | 9.13 | 4.55 | 6.17 | 90.95 | 123.49 | 4.58 | 16.02 | 3.88 | 5.34 | 84.83 | 116.75 |
| 6 | 1.53 | 11.83 | 1.27 | 1.98 | 82.78 | 129.55 | 0.93 | 12.19 | 0.82 | 1.04 | 87.82 | 112.20 |
| 7 | 0.12 | 15.33 | 0.09 | 0.16 | 80.06 | 136.35 | 6.08 | 110.85 | 0.08 | 13.37 | 1.33 | 219.95 |
| 8 | 15.48 | 8.60 | 13.80 | 18.33 | 89.15 | 118.39 | 16.31 | 26.00 | 13.24 | 21.14 | 81.18 | 129.66 |
| 9+10 | 16.04 | 7.84 | 13.89 | 17.98 | 86.61 | 112.10 | 11.20 | 29.67 | 8.26 | 14.80 | 73.82 | 132.23 |
| 11 | 6.34 | 12.63 | 5.34 | 8.29 | 84.25 | 130.71 | 5.62 | 4.51 | 5.41 | 5.90 | 96.23 | 105.00 |
| 12 | 11.40 | 27.52 | 6.59 | 16.32 | 57.82 | 143.13 | 9.57 | 18.19 | 7.78 | 11.26 | 81.29 | 117.62 |
| 13 | 0.38 | 37.29 | 0.21 | 0.63 | 55.22 | 165.87 | 0.80 | 26.82 | 0.56 | 0.97 | 70.10 | 126.06 |
| 14 | 4.54 | 10.72 | 3.76 | 5.98 | 82.88 | 131.54 | 7.39 | 10.63 | 6.60 | 8.17 | 89.40 | 109.85 |
| 15 | 0.50 | 45.01 | 0.26 | 0.86 | 51.68 | 172.70 | 0.42 | 31.60 | 0.29 | 0.55 | 68.59 | 131.42 |
| 17 | 27.39 | 7.68 | 21.04 | 29.76 | 76.81 | 108.65 | 25.42 | 18.99 | 21.94 | 30.93 | 86.32 | 121.68 |
| 18 | 6.15 | 13.11 | 5.22 | 8.22 | 84.82 | 133.59 | 5.29 | 47.94 | 3.68 | 8.21 | 69.59 | 155.60 |
| 13+14 * | 4.92 | 11.74 | 4.08 | 6.55 | 82.79 | 133.09 | 8.19 | 7.09 | 7.58 | 8.74 | 92.58 | 106.71 |
| 15+18 * | 6.65 | 14.11 | 5.49 | 8.98 | 82.61 | 135.13 | 5.71 | 46.40 | 4.10 | 8.76 | 71.78 | 153.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusanova, M.; Rusanov, K.; Alekova, M.; Georgieva, L.; Georgieva, P.; Zagorcheva, T.; Atanassov, I. A Simple High-Throughput Procedure for Microscale Extraction of Bioactive Compounds from the Flowers of Saint John’s Wort (Hypericum perforatum L.). Appl. Sci. 2025, 15, 7334. https://doi.org/10.3390/app15137334
Rusanova M, Rusanov K, Alekova M, Georgieva L, Georgieva P, Zagorcheva T, Atanassov I. A Simple High-Throughput Procedure for Microscale Extraction of Bioactive Compounds from the Flowers of Saint John’s Wort (Hypericum perforatum L.). Applied Sciences. 2025; 15(13):7334. https://doi.org/10.3390/app15137334
Chicago/Turabian StyleRusanova, Mila, Krasimir Rusanov, Marina Alekova, Liliya Georgieva, Pavlina Georgieva, Tzvetelina Zagorcheva, and Ivan Atanassov. 2025. "A Simple High-Throughput Procedure for Microscale Extraction of Bioactive Compounds from the Flowers of Saint John’s Wort (Hypericum perforatum L.)" Applied Sciences 15, no. 13: 7334. https://doi.org/10.3390/app15137334
APA StyleRusanova, M., Rusanov, K., Alekova, M., Georgieva, L., Georgieva, P., Zagorcheva, T., & Atanassov, I. (2025). A Simple High-Throughput Procedure for Microscale Extraction of Bioactive Compounds from the Flowers of Saint John’s Wort (Hypericum perforatum L.). Applied Sciences, 15(13), 7334. https://doi.org/10.3390/app15137334