

Multi-Sensing Monitoring of the Microalgae Biomass Cultivation Systems for Biofuels and Added Value Products Synthesis—Challenges and Opportunities




Abstract
1. Introduction
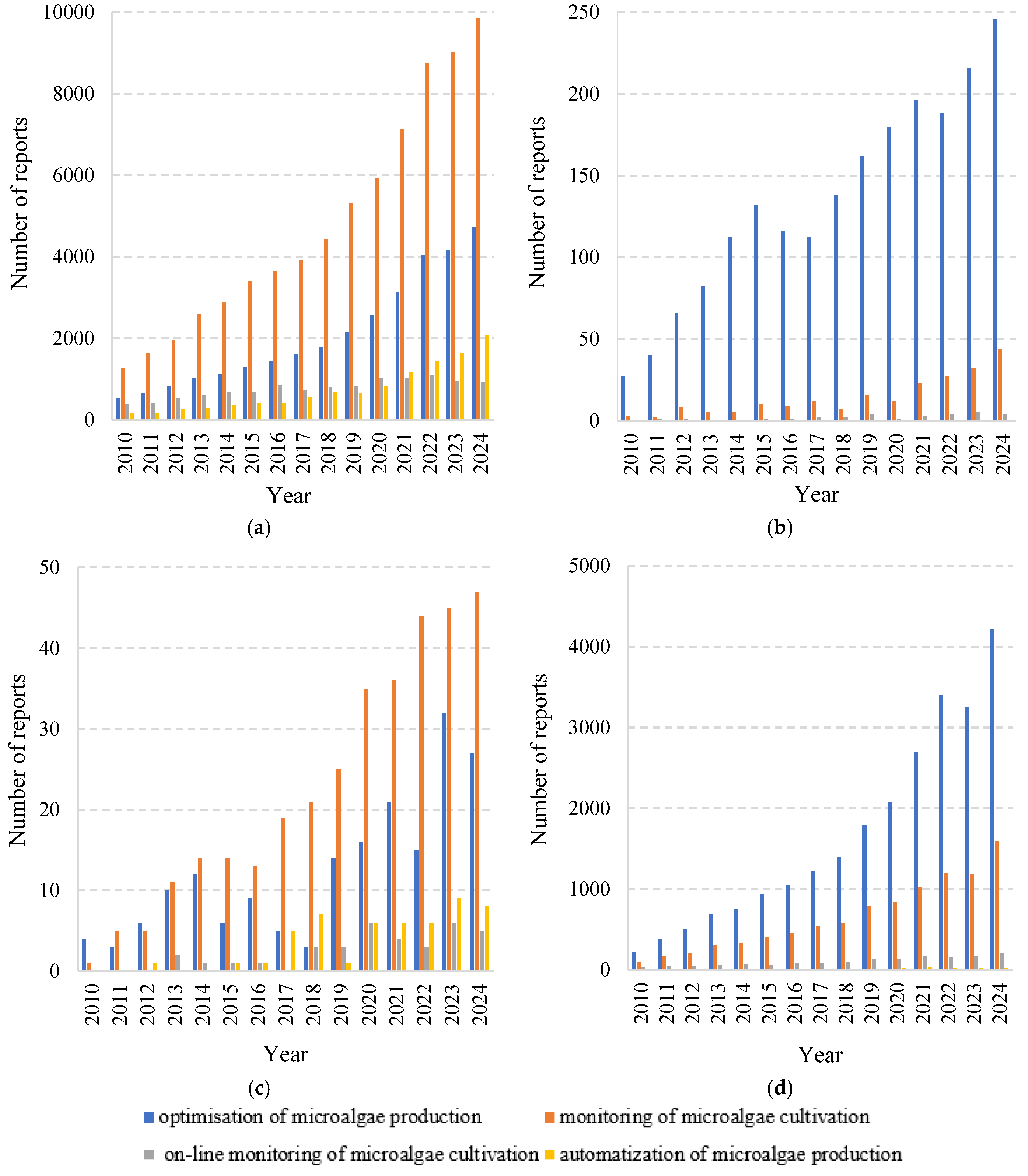
2. Bibliographical Analysis, Research Directions, and Scientific Potential
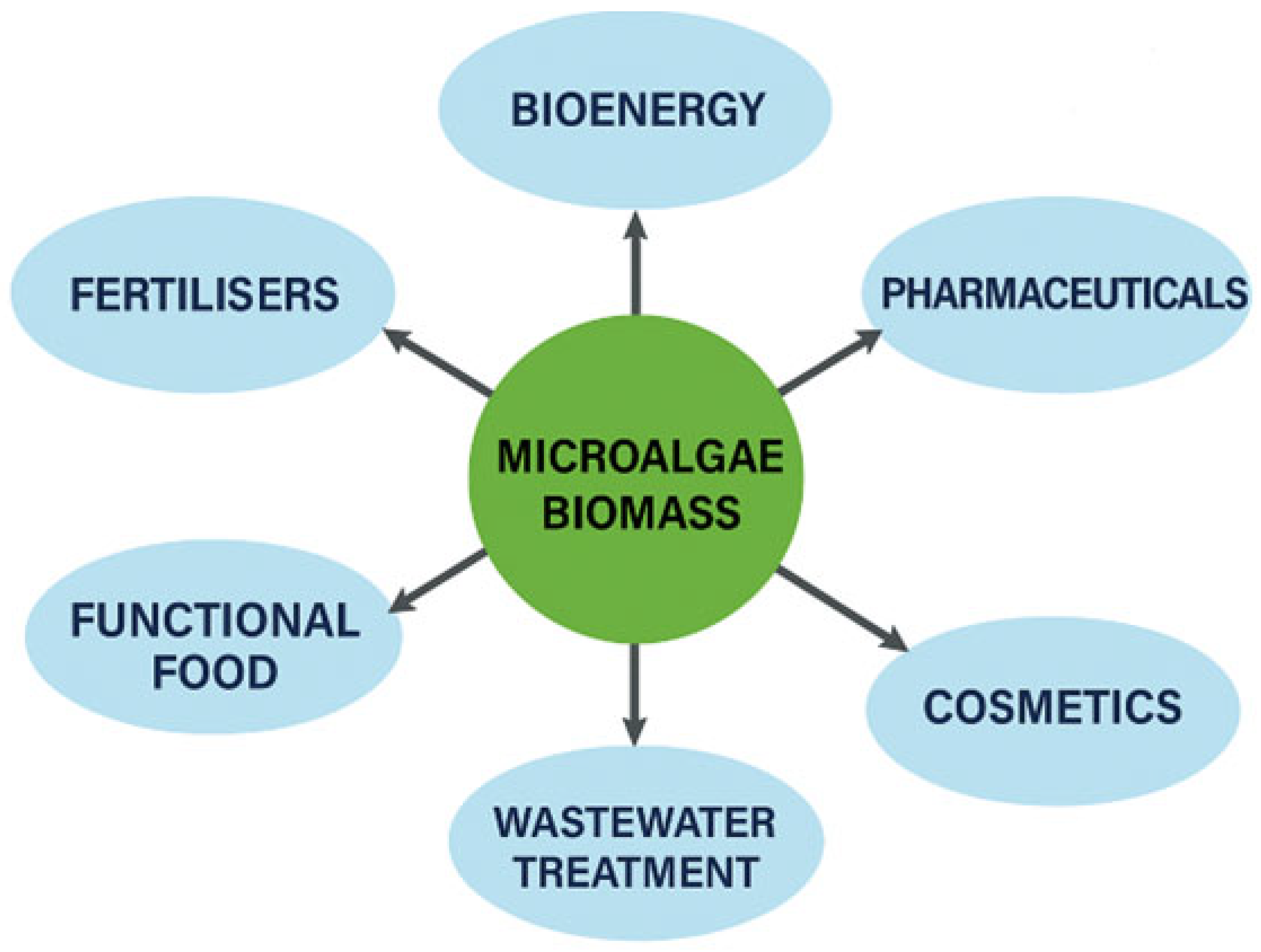
3. Application Sectors and Valuable Products from Microalgae Biomass
4. Optimisation of the Microalgae Biomass Cultivation and Valuable Products Synthesis
5. Monitored and Controlled Indicators Critical to Microalgae Cultivation
5.1. The Concentration of Nutrients and Microelements
5.2. Carbon Dioxide Concentration
5.3. pH Value
5.4. Temperature
5.5. Source and Intensity of Lighting
5.6. Oxygen Concentration
6. Methods of Microalgae Cultivation Monitoring
6.1. Light Monitoring
6.2. Culture Temperature Monitoring
6.3. Monitoring of pH Changes in the Culture Medium and CO2 Content
6.4. Monitoring of O2 Concentration in the Culture Medium and Gaseous Metabolites
6.5. Monitoring of Nutrient Content in the Culture Medium
6.6. Dynamics of Growth, Concentration, and Biomass Composition
6.7. Photosynthetic Activity
7. New Approaches to Monitoring and Optimisation of Microalgae Cultivation Systems
7.1. Two-Dimensional and Infrared Spectroscopy
7.2. Multiparameter Flow Cytometry
7.3. Metaheuristic-Based Predictions
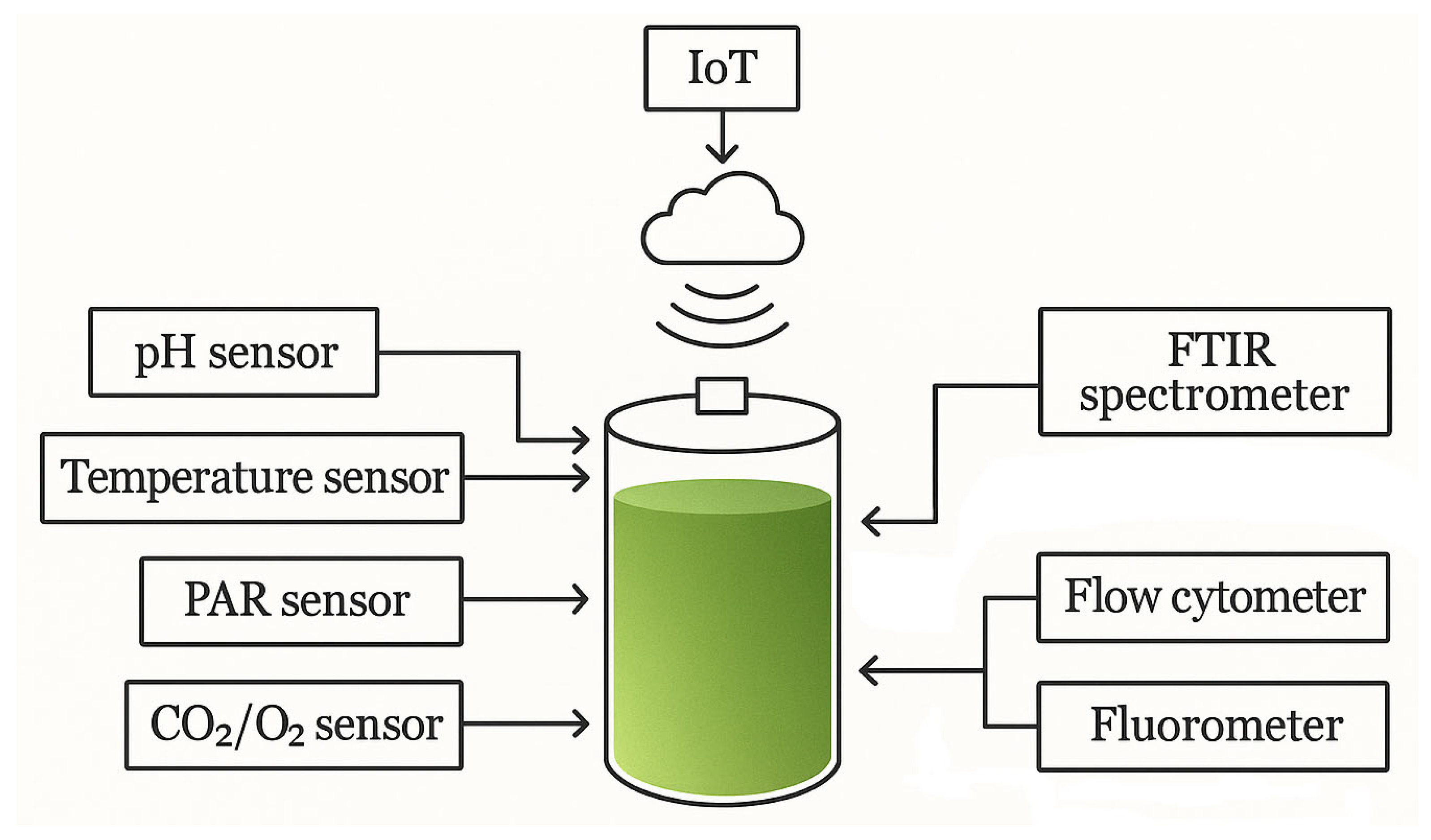
7.4. Internet of Things
7.5. RGB Sensors (Red–Green–Blue)
7.6. Smart Photobioreactors
8. Classification of Monitoring Techniques and Principles for Their Application
9. Summary and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Balamurugan, S.; Li, D.W.; Wang, X.; Li, H.Y. Unleashing the Potential of Biotechnological Strategies for the Sustainable Production of Microalgal Polysaccharides. Crit. Rev. Food Sci. Nutr. 2025, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Deamici, K.M.; Figueiredo, D.; Guerra, I.; Letras, P.; Pereira, H. Global Market and Future Trends of Microalgae-Based Products. Algal Bioreact. Sci. Eng. Technol. Upstream Process. 2025, 1, 11–25. [Google Scholar] [CrossRef]
- Yeheyo, H.A.; Ealias, A.M.; George, G.; Jagannathan, U. Bioremediation Potential of Microalgae for Sustainable Soil Treatment in India: A Comprehensive Review on Heavy Metal and Pesticide Contaminant Removal. J. Environ. Manag. 2024, 363, 121409. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, Y.; Zhang, Q. Assessing Global Carbon Sequestration and Bioenergy Potential from Microalgae Cultivation on Marginal Lands Leveraging Machine Learning. Sci. Total Environ. 2024, 948, 174462. [Google Scholar] [CrossRef]
- Samoraj, M.; Çalış, D.; Trzaska, K.; Mironiuk, M.; Chojnacka, K. Advancements in Algal Biorefineries for Sustainable Agriculture: Biofuels, High-Value Products, and Environmental Solutions. Biocatal. Agric. Biotechnol. 2024, 58, 103224. [Google Scholar] [CrossRef]
- Çelekli, A.; Özbal, B.; Bozkurt, H. Challenges in Functional Food Products with the Incorporation of Some Microalgae. Foods 2024, 13, 725. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae Cultivation Technologies as an Opportunity for Bioenergetic System Development—Advantages and Limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Musa Ardo, F.; Wei Lim, J.; Ramli, A.; Kee Lam, M.; Kiatkittipong, W.; Alaaeldin Abdelfattah, E.; Kashif Shahid, M.; Usman, A.; Wongsakulphasatch, S.; Tasnim Sahrin, N. A Review in Redressing Challenges to Produce Sustainable Hydrogen from Microalgae for Aviation Industry. Fuel 2022, 330, 125646. [Google Scholar] [CrossRef]
- Dębowski, M.; Świca, I.; Kazimierowicz, J.; Zieliński, M. Large Scale Microalgae Biofuel Technology–Development Perspectives in Light of the Barriers and Limitations. Energies 2022, 16, 81. [Google Scholar] [CrossRef]
- Dudek, M.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M.; Quattrocelli, P.; Nowicka, A. The Cultivation of Biohydrogen-Producing Tetraselmis Subcordiformis Microalgae as the Third Stage of Dairy Wastewater Aerobic Treatment System. Sustainability 2022, 14, 12085. [Google Scholar] [CrossRef]
- Falfushynska, H. Advancements and Prospects in Algal Biofuel Production: A Comprehensive Review. Phycology 2024, 4, 548–575. [Google Scholar] [CrossRef]
- Mrinal; Jaiswal, K.S.; Singh, S.; Khalid, Z.; Alam, S.N.; Sundaramurthy, S.; Shukla, N.K.; Ramaswamy, A.P.; Jaiswal, K.K.; Singh, B.; et al. Microalgal Biofuels Production and Advances in Sustainable Applications. In Microalgal Biofuels; Woodhead Publishing: Cambridge, UK, 2025; pp. 395–413. [Google Scholar] [CrossRef]
- Yap, J.K.; Sankaran, R.; Chew, K.W.; Halimatul Munawaroh, H.S.; Ho, S.H.; Rajesh Banu, J.; Show, P.L. Advancement of Green Technologies: A Comprehensive Review on the Potential Application of Microalgae Biomass. Chemosphere 2021, 281, 130886. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Patial, S.K.; Singh, S. Algae Biomass: Importance, Harvesting Techniques, Extraction Methods, and Associated Challenges. In Value Added Products from Bioalgae Based Biorefineries: Opportunities and Challenges; Springer Nature: Singapore, 2024; pp. 67–94. [Google Scholar] [CrossRef]
- Yadav, G.; Mathimani, T.; Sekar, M.; Sindhu, R.; Pugazhendhi, A. Strategic Evaluation of Limiting Factors Affecting Algal Growth—An Approach to Waste Mitigation and Carbon Dioxide Sequestration. Sci. Total Environ. 2021, 796, 149049. [Google Scholar] [CrossRef]
- Olabi, A.G.; Shehata, N.; Sayed, E.T.; Rodriguez, C.; Anyanwu, R.C.; Russell, C.; Abdelkareem, M.A. Role of Microalgae in Achieving Sustainable Development Goals and Circular Economy. Sci. Total Environ. 2023, 854, 158689. [Google Scholar] [CrossRef] [PubMed]
- Khor, W.H.; Kang, H.S.; Lim, J.W.; Iwamoto, K.; Tang, C.H.H.; Goh, P.S.; Quen, L.K.; Bin Shaharuddin, N.M.R.; Lai, N.Y.G. Microalgae Cultivation in Offshore Floating Photobioreactor: State-of-the-Art, Opportunities and Challenges. Aquac. Eng. 2022, 98, 102269. [Google Scholar] [CrossRef]
- Psachoulia, P.; Chatzidoukas, C. Illumination Policies for Stichococcus Sp. Cultures in an Optimally Operating Lab-Scale PBR toward the Directed Photosynthetic Production of Desired Products. Sustainability 2021, 13, 2489. [Google Scholar] [CrossRef]
- Porras Reyes, L.; Havlik, I.; Beutel, S. Software Sensors in the Monitoring of Microalgae Cultivations. Rev. Environ. Sci. Bio/Technol. 2024, 23, 67–92. [Google Scholar] [CrossRef]
- Havlik, I.; Beutel, S.; Scheper, T.; Reardon, K.F. On-Line Monitoring of Biological Parameters in Microalgal Bioprocesses Using Optical Methods. Energies 2022, 15, 875. [Google Scholar] [CrossRef]
- Yu, B.S.; Pyo, S.; Lee, J.; Han, K. Microalgae: A Multifaceted Catalyst for Sustainable Solutions in Renewable Energy, Food Security, and Environmental Management. Microb. Cell Factories 2024, 23, 308. [Google Scholar] [CrossRef]
- Chieti, M.G.; Petrucciani, A.; Mollo, L.; Gerotto, C.; Eusebi, A.L.; Fatone, F.; Norici, A.; González-Camejo, J. Acclimated Green Microalgae Consortium to Treat Sewage in an Alternative Urban WWTP in a Coastal Area of Central Italy. Sci. Total Environ. 2024, 945, 174056. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Vdovychenko, A.; Kazimierowicz, J. The Use of the Autotrophic Culture of Arthrospira Platensis for CO2 Fixation from Biogas Combustion. Processes 2024, 12, 396. [Google Scholar] [CrossRef]
- Emalya, N.; Munawar, E. Synergistic Removal of Organic and Nutrients from Landfill Leachate Using Photobiore-Actor-Cultivated Microalgae-Bacteria Consortium. Glob. J. Environ. Sci. Manag. 2024, 10, 683–698. [Google Scholar] [CrossRef]
- Gonçalves, R.F.; Bastos, L.P.; Nariyoshi, Y.N.; Borges, R.M.; Keller, R.; Silveira, D.D. Assessment of Selected Parameters in CO2 and CH4 Mass Transfer During Photosynthetic Biogas Upgrading Using Bubble Columns Filled with Wastewater-Derived Microalgae. Energy Fuels 2025, 39, 7314–7325. [Google Scholar] [CrossRef]
- Mallick, N.C.; Mozammil, S.; Koshta, E. Investigation on Chlorella Sp. HS2 Microalgae Growth Using CO2 from Engine Exhaust to Study Carbon Sequestration. Biofuels 2024, 15, 279–290. [Google Scholar] [CrossRef]
- Song, Y.; Xie, L.; Zhang, X.; Hu, Z.; Li, S.; Zhang, P.; Yang, X. Enhancement of Biomass, Lipid Accumulation, and Carbon Sequestration Potential in Microalgae via Cultivation with Aggregation-Induced Emission Light-Conversion Films. Chem. Eng. J. 2024, 483, 149148. [Google Scholar] [CrossRef]
- Sirohi, R.; Il Choi, H.; Sim, S.J. Microalgal Fuels: Promising Energy Reserves for the Future. Fuel 2022, 312, 122841. [Google Scholar] [CrossRef]
- Dębowski, M.; Dudek, M.; Kazimierowicz, J.; Quattrocelli, P.; Rusanowska, P.; Barczak, Ł.; Nowicka, A.; Zieliński, M. Analysis of Multi-Biofuel Production during Cultivation of the Green Microalga Tetraselmis Subscordiformis. Energies 2024, 17, 3670. [Google Scholar] [CrossRef]
- Ebhodaghe, S.O.; Imanah, O.E.; Ndibe, H. Biofuels from Microalgae Biomass: A Review of Conversion Processes and Procedures. Arab. J. Chem. 2022, 15, 103591. [Google Scholar] [CrossRef]
- Sarwer, A.; Hamed, S.M.; Osman, A.I.; Jamil, F.; Al-Muhtaseb, A.H.; Alhajeri, N.S.; Rooney, D.W. Algal Biomass Valorization for Biofuel Production and Carbon Sequestration: A Review. Environ. Chem. Lett. 2022, 20, 2797–2851. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Ali, S.S.; Ramadan, H.; El-Aswar, E.I.; Eltawab, R.; Ho, S.H.; Elsamahy, T.; Li, S.; El-Sheekh, M.M.; Schagerl, M.; et al. Microalgae-Based Wastewater Treatment: Mechanisms, Challenges, Recent Advances, and Future Prospects. Environ. Sci. Ecotechnol. 2023, 13, 100205. [Google Scholar] [CrossRef]
- Ferreira Carraro, C.d.F.; Almeida Loures, C.C.; de Castro, J.A. Microalgae Bioremediation and CO2 Fixation of Industrial Wastewater. Clean. Eng. Technol. 2022, 8, 100466. [Google Scholar] [CrossRef]
- Sforza, E.; Kumkum, P.; Barbera, E.; Kumar, S. Bioremediation of Industrial Effluents: How a Biochar Pretreatment May Increase the Microalgal Growth in Tannery Wastewater. J. Water Process Eng. 2020, 37, 101431. [Google Scholar] [CrossRef]
- Dȩbowski, M.; Rusanowska, P.; Zieliński, M.; Dudek, M.; Romanowska-Duda, Z. Biomass Production and Nutrient Removal by Chlorella Vulgaris from Anaerobic Digestion Effluents. Energies 2018, 11, 1654. [Google Scholar] [CrossRef]
- Tawfik, A.; Eraky, M.; Alhajeri, N.S.; Osman, A.I.; Rooney, D.W. Cultivation of Microalgae on Liquid Anaerobic Digestate for Depollution, Biofuels and Cosmetics: A Review. Environ. Chem. Lett. 2022, 20, 3631–3656. [Google Scholar] [CrossRef]
- Kandimalla, P.; Vatte, P.; Bandaru, C.S.R. Phycoremediation of Automobile Exhaust Gases Using Green Microalgae. Environ. Dev. Sustain. 2021, 23, 6301–6322. [Google Scholar] [CrossRef]
- Wang, B.; Xu, Y.F.; Sun, Z.L. Mass Transfer Characteristics and Effect of Flue Gas Used in Microalgae Culture. Appl. Microbiol. Biotechnol. 2022, 106, 7013–7025. [Google Scholar] [CrossRef]
- Dahai, H.; Zhihong, Y.; Lin, Q.; Yuhong, L.; Lei, T.; Jiang, L.; Liandong, Z. The Application of Magical Microalgae in Carbon Sequestration and Emission Reduction: Removal Mechanisms and Potential Analysis. Renew. Sustain. Energy Rev. 2024, 197, 114417. [Google Scholar] [CrossRef]
- Tamil Selvan, S.; Balasubaramaian, A.; Manickam Dakshinamoorthi, B.K.; Velramar, B.; Fareed, M. Green Integrative Large Scale Treatment of Tannery Effluent, CO2 Sequestration, and Biofuel Production Using Oleaginous Green Microalga Nannochloropsis Oculata TSD05: An Ecotechnological Approach. Biocatal. Agric. Biotechnol. 2024, 61, 103370. [Google Scholar] [CrossRef]
- Hassaan, M.A.; Abdelaziz, N.I.M.; Nazir, M.A.; Jamshaid, M.; Hassouna, M.S.; El Nemr, A. Box–Behnken Modeling of Biodiesel Production from Botryococcus Braunii Microalgae. Biofuels Bioprod. Biorefining 2024, 18, 1321–1354. [Google Scholar] [CrossRef]
- Moradi, P.; Saidi, M. Biodiesel Production from Chlorella Vulgaris Microalgal-Derived Oil via Electrochemical and Thermal Processes. Fuel Process. Technol. 2022, 228, 107158. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, K.; Pan, X.; Liu, G.; Cai, Y.; Zaidi, A.A.; Zhang, K. Substrate Degradation, Biodiesel Production, and Microbial Community of Two Electro-Fermentation Systems on Treating Oleaginous Microalgae Nannochloropsis Sp. Bioresour. Technol. 2021, 329, 124932. [Google Scholar] [CrossRef]
- Acebu, P.I.G.; de Luna, M.D.G.; Chen, C.Y.; Abarca, R.R.M.; Chen, J.H.; Chang, J.S. Bioethanol Production from Chlorella Vulgaris ESP-31 Grown in Unsterilized Swine Wastewater. Bioresour. Technol. 2022, 352, 127086. [Google Scholar] [CrossRef] [PubMed]
- Ma’mun, S.; Prasetio, M.W.; Anugrah, A.R.; Ruliandi, A.P.; Pramuwardani, D. Bioethanol from Arthrospira Platensis Biomass Using a Combined Pretreatment. Chem. Eng. J. Adv. 2024, 19, 100616. [Google Scholar] [CrossRef]
- Silva, G.; Cerqueira, K.; Rodrigues, J.; Silva, K.; Coelho, D.; Souza, R. Cultivation of Microalgae Chlorella Vulgaris in Open Reactor for Bioethanol Production. Phycology 2023, 3, 325–336. [Google Scholar] [CrossRef]
- El-Moustaqim, K.; Rachiq, T.; Mabrouki, J.; Slaoui, M.; Hmouni, D. Application of Microalgae in Green Hydrogen Production: A Diverse Biomass for Multiple Applications. Environ. Sci. Eng. 2024, Part F3279, 231–240. [Google Scholar] [CrossRef]
- Ramprakash, B.; Lindblad, P.; Eaton-Rye, J.J.; Incharoensakdi, A. Current Strategies and Future Perspectives in Biological Hydrogen Production: A Review. Renew. Sustain. Energy Rev. 2022, 168, 112773. [Google Scholar] [CrossRef]
- Zhang, Y.; Hong, Y.; Wang, X. Recent Advances on Using Functional Materials to Increase the Pollutant Removal Capabilities of Microalgae and Bacteria: Especially for Their Symbiotic Systems. Curr. Pollut. Rep. 2023, 9, 272–291. [Google Scholar] [CrossRef]
- Dębowski, M.; Kazimierowicz, J.; Świca, I.; Zieliński, M. Ultrasonic Disintegration to Improve Anaerobic Digestion of Microalgae with Hard Cell Walls—Scenedesmus Sp. and Pinnularia Sp. Plants 2022, 12, 53. and Pinnularia Sp. Plants 2022, 12, 53. [Google Scholar] [CrossRef]
- Xie, T.; Herbert, C.; Zitomer, D.; Kimbell, L.; Stafford, M.; Venkiteshwaran, K. Biogas Conditioning and Digestate Recycling by Microalgae: Acclimation of Chlorella Vulgaris to H2S-Containing Biogas and High NH4-N Digestate and Effect of Biogas: Digestate Ratio. Chem. Eng. J. 2023, 453, 139788. [Google Scholar] [CrossRef]
- Torres, A.; Padrino, S.; Brito, A.; Díaz, L. Biogas Production from Anaerobic Digestion of Solid Microalgae Residues Generated on Different Processes of Microalgae-to-Biofuel Production. Biomass Convers. Biorefinery 2023, 13, 4659–4672. [Google Scholar] [CrossRef]
- Eusébio, A.; Santos, C.A.; Marques, I.P. Anaerobic Digestion of Microalga Chlorella Protothecoides and Metagenomic Analysis of Reddish-Colored Digestate. Appl. Sci. 2023, 13, 3325. [Google Scholar] [CrossRef]
- Markou, G.; Ilkiv, B.; Brulé, M.; Antonopoulos, D.; Chakalis, L.; Arapoglou, D.; Chatzipavlidis, I. Methane Production through Anaerobic Digestion of Residual Microalgal Biomass after the Extraction of Valuable Compounds. Biomass Convers. Biorefinery 2022, 12, 419–426. [Google Scholar] [CrossRef]
- Manthos, G.; Tsigkou, K.; Koutra, E.; Mingou, L.; Kornaros, M. Exploring the Impact of Microalgal Fatty Acid Content on Anaerobic Digestion and Methane Production: An Experimental and Mathematical Modeling Study. Renew. Energy 2024, 230, 120859. [Google Scholar] [CrossRef]
- Dębowski, M.; Dudek, M.; Nowicka, A.; Quattrocelli, P.; Kazimierowicz, J.; Zieliński, M. Suitability of Pre-Digested Dairy Effluent for Mixotrophic Cultivation of the Hydrogen-Producing Microalgae Tetraselmis Subcordiformis. Environ. Technol. 2024, 45, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Dudek, M.; Dębowski, M.; Nowicka, A.; Kazimierowicz, J.; Zieliński, M. The Effect of Autotrophic Cultivation of Platymonas Subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield. Resources 2022, 11, 31. [Google Scholar] [CrossRef]
- Sitthikitpanya, N.; Sittijunda, S.; Khamtib, S.; Reungsang, A. Co-Generation of Biohydrogen and Biochemicals from Co-Digestion of Chlorella Sp. Biomass Hydrolysate with Sugarcane Leaf Hydrolysate in an Integrated Circular Biorefinery Concept. Biotechnol. Biofuels 2021, 14, 197. [Google Scholar] [CrossRef]
- Raheem, A.; Abbasi, S.A.; Mangi, F.H.; Ahmed, S.; He, Q.; Ding, L.; Memon, A.A.; Zhao, M.; Yu, G. Gasification of Algal Residue for Synthesis Gas Production. Algal Res. 2021, 58, 102411. [Google Scholar] [CrossRef]
- Scopel, L.; Marcantonio, V. Gasification of Chlorella Vulgaris for Syngas Production and Energy Generation Through Gas Turbine. Energies 2024, 17, 6085. [Google Scholar] [CrossRef]
- Banihashemi, F.; Ibrahim, A.F.M.; Deng, S.; Lin, J.Y.S. Pyrolysis and Gasification Characteristics of Galdieria Sulphuraria Microalgae. Bioenergy Res. 2023, 16, 611–621. [Google Scholar] [CrossRef]
- Sharma, A.K.; Ghodke, P.; Sharma, P.K.; Manna, S.; Pugazhendhi, A.; Matsakas, L.; Patel, A. Holistic Utilization of Chlorella Pyrenoidosa Microalgae for Extraction of Renewable Fuels and Value-Added Biochar through in Situ Transesterification and Pyrolysis Reaction Process. Biomass Convers. Biorefinery 2024, 14, 5261–5274. [Google Scholar] [CrossRef]
- Makowska, M.; Dziosa, K. Influence of Different Pyrolysis Temperatures on Chemical Composition and Graphite-like Structure of Biochar Produced from Biomass of Green Microalgae Chlorella Sp. Environ. Technol. Innov. 2024, 35, 103667. [Google Scholar] [CrossRef]
- Saiyud, N.; Deethayat, T.; Asanakham, A.; Duongbia, N.; Kamopas, W.; Kiatsiriroat, T. Biochar Production from Co-Pyrolysis of Coffee Ground and Native Microalgae Consortium. Biomass Convers. Biorefinery 2024, 14, 6855–6863. [Google Scholar] [CrossRef]
- Tang, Z.; Chen, W.; Chen, Y.; Hu, J.; Yang, H.; Chen, H. Preparation of Low-Nitrogen and High-Quality Bio-Oil from Microalgae Catalytic Pyrolysis with Zeolites and Activated Carbon. J. Anal. Appl. Pyrolysis 2021, 159, 105182. [Google Scholar] [CrossRef]
- Mahfud, M.; Qadariyah, L.; Haqqyana, H.; Aswie, V. Optimization Bio-Oil Production from Chlorella Sp. through Microwave-Assisted Pyrolysis Using Response Surface Methodology. Green Energy Resour. 2024, 2, 100057. [Google Scholar] [CrossRef]
- Mustapha, S.I.; Mohammed, U.A.; Rawat, I.; Bux, F.; Isa, Y.M. Production of High-Quality Pyrolytic Bio-Oils from Nutrient-Stressed Scenedesmus Obliquus Microalgae. Fuel 2023, 332, 126299. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Wang, T.; Zhang, X.; Wang, L.; Hu, X.; Guo, Q. Co-Production of Upgraded Bio-Oils and H2-Rich Gas from Microalgae via Chemical Looping Pyrolysis. Int. J. Hydrogen Energy 2021, 46, 24942–24955. [Google Scholar] [CrossRef]
- Kong, W.; Shen, B.; Ma, J.; Kong, J.; Feng, S.; Wang, Z.; Xiong, L. Pyrolysis of Spirulina Platensis, Tetradesmus Obliquus and Chlorella Vulgaris by TG-FTIR and Py-GC/MS: Kinetic Analysis and Pyrolysis Behaviour. Energy 2022, 244, 123165. [Google Scholar] [CrossRef]
- Al-Ansari, M.M.; Al-Humaid, L.; Al-Dahmash, N.D.; Aldawsari, M. Assessing the Benefits of Chlorella Vulgaris Microalgal Biodiesel for Internal Combustion Engines: Energy and Exergy Analyses. Fuel 2023, 344, 128055. [Google Scholar] [CrossRef]
- Romadinullah, R.A.; Sukarni, S.; Zakaria, Y.; Aminullah, A.Y.; Permanasari, A.A.; Anis, S.; Johari, A. Thermal Behavior and Combustion Kinetics of Arthrospira Platensis-Activated Carbon Mixture at Mass Ratio of 10:3. In AIP Conference Proceedings; AIP Publishing: Melville, NY, USA, 2024; Volume 3124. [Google Scholar] [CrossRef]
- Dessì, F.; Mureddu, M.; Ferrara, F.; Fermoso, J.; Orsini, A.; Sanna, A.; Pettinau, A. Thermogravimetric Characterisation and Kinetic Analysis of Nannochloropsis sp. and Tetraselmis sp. Microalgae for Pyrolysis, Combustion and Oxy-Combustion. Energy 2021, 217, 119394. [Google Scholar] [CrossRef]
- Chen, C.; Tang, T.; Shi, Q.; Zhou, Z.; Fan, J. The Potential and Challenge of Microalgae as Promising Future Food Sources. Trends Food Sci. Technol. 2022, 126, 99–112. [Google Scholar] [CrossRef]
- Loke Show, P. Global Market and Economic Analysis of Microalgae Technology: Status and Perspectives. Bioresour. Technol. 2022, 357, 127329. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Hegde, A.S.; Sharma, K.; Parmar, P.; Srivatsan, V. Microalgae as a Sustainable Source of Edible Proteins and Bioactive Peptides—Current Trends and Future Prospects. Food Res. Int. 2022, 157, 111338. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Reyes, J.A.; González-Hernández, J.C. Vitamins from Microalgae. In Handbook of Food and Feed from Microalgae; Academic Press: Cambridge, MA, USA, 2023; pp. 111–115. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Torrella, J.R.; Pagès, T.; Viscor, G.; Torres, J.L. Edible Microalgae and Their Bioactive Compounds in the Prevention and Treatment of Metabolic Alterations. Nutrients 2021, 13, 563. [Google Scholar] [CrossRef]
- Mutaf-Kılıc, T.; Oncel, S.S. Algal and Cyanobacterial Colorants. From Chlorophyll to Phycocyanin. In Microbial Colorants: Chemistry, Biosynthesis and Applications; Wiley: Hoboken, NJ, USA, 2025; pp. 81–105. [Google Scholar] [CrossRef]
- Nair, A.; Rao, A.S.; Ayesha, B.; Salu, H.A.; Ashraf, Q.; Hebbar, A.S.; Yuktha, K.; Khadyal, P.; Poulamee, S.; Veena, S.M.; et al. Phycobiliproteins. Algal and Cyanobacterial Pigments Radiating Vivid Colors. In Microbial Colorants: Chemistry, Biosynthesis and Applications; Wiley: Hoboken, NJ, USA, 2025; pp. 241–259. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.Y.; Filaire, E. Microalgae N-3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, Y.; Maltseva, K. Fatty Acids of Microalgae: Diversity and Applications. Rev. Environ. Sci. Bio/Technol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Kaamoush, M.; El-Agawany, N.; El Salhin, H.; El-Zeiny, A. Monitoring Effect of Nickel, Copper, and Zinc on Growth and Photosynthetic Pigments of Spirulina Platensis with Suitability Investigation in Idku Lake. Environ. Sci. Pollut. Res. 2022, 29, 78942–78959. [Google Scholar] [CrossRef]
- Reshma, R.; Chitra Devi, K.; Dinesh Kumar, S.; Santhanam, P.; Perumal, P.; Krishnaveni, N.; Begum, A.; Pragnya, M.; Arthikha, R.; Dhanalakshmi, B.; et al. Enhancement of Pigments Production in the Green Microalga Dunaliella Salina (PSBDU05) under Optimized Culture Condition. Bioresour. Technol. Rep. 2021, 14, 100672. [Google Scholar] [CrossRef]
- Grujić, V.J.; Todorović, B.; Ambrožič-Dolinšek, J.; Kranvogl, R.; Ciringer, T. Diversity and Content of Carotenoids and Other Pigments in the Transition from the Green to the Red Stage of Haematococcus Pluvialis Microalgae Identified by HPLC-DAD and LC-QTOF-MS. Plants 2022, 11, 1026. [Google Scholar] [CrossRef]
- Ma, W.; Liu, M.; Zhang, Z.; Xu, Y.; Huang, P.; Guo, D.; Sun, X.; Huang, H. Efficient Co-Production of EPA and DHA by Schizochytrium Sp. via Regulation of the Polyketide Synthase Pathway. Commun. Biol. 2022, 5, 1356. [Google Scholar] [CrossRef]
- Navarro López, E.; Jiménez Callejón, M.J.; Macías Sánchez, M.D.; González Moreno, P.A.; Robles Medina, A. Obtaining Eicosapentaenoic Acid-Enriched Polar Lipids from Microalga Nannochloropsis Sp. by Lipase-Catalysed Hydrolysis. Algal Res. 2023, 71, 103073. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, M.; Cao, J.; Wang, Y.; Zhang, L.; Yan, X.; Li, Y.; Xu, J. Enhancement of Docosahexaenoic Acid and Eicosapentaenoic Acid Biosynthesis in Isochrysis Galbana by Bacillus Jeotgali. Mar. Biotechnol. 2024, 26, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, R.A.; El Latif, A.A.; Elkaw, E.M.; Alotaibi, A.S.; Alenzi, A.M.; Hamza, H.A. Assessment of Antioxidant and Anticancer Activities of Microgreen Alga Chlorella Vulgaris and Its Blend with Different Vitamins. Molecules 2022, 27, 1602. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, R.A.; El-Boraey, N.G.; El Bialy, B.E.; Alrdahe, S.S.; Darwish, D.B.E. Vitamin Supplements Enhance Spirulina Platensis Biomass and Phytochemical Contents. Green Process. Synth. 2022, 11, 266–274. [Google Scholar] [CrossRef]
- Cunha, S.A.; Coscueta, E.R.; Nova, P.; Silva, J.L.; Pintado, M.M. Bioactive Hydrolysates from Chlorella Vulgaris: Optimal Process and Bioactive Properties. Molecules 2022, 27, 2505. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qi, F.; Yang, F.; Khan, Q.U.; Faizan; Fu, Q.; Li, J.; Wang, X.; Wang, X.; Wang, J.; et al. Nutritional Composition and Functional Properties of A. Platensis-Derived Peptides: A Green and Sustainable Protein-Rich Supplement. Processes 2024, 12, 2608. [Google Scholar] [CrossRef]
- Lopes, D.; Rey, F.; Gomes, A.; Duarte, L.; Pereira, J.; Pinho, M.; Melo, T.; Domingues, R. Tracing the Impact of Domestic Storage Conditions on Antioxidant Activity and Lipid Profiles in the Edible Microalgae Chlorella Vulgaris and Tetraselmis Chui. Mar. Drugs 2024, 22, 254. [Google Scholar] [CrossRef]
- Gargouch, N.; Elleuch, F.; Karkouch, I.; Tabbene, O.; Pichon, C.; Gardarin, C.; Rihouey, C.; Picton, L.; Abdelkafi, S.; Fendri, I.; et al. Potential of Exopolysaccharide from Porphyridium Marinum to Contend with Bacterial Proliferation, Biofilm Formation, and Breast Cancer. Mar. Drugs 2021, 19, 66. [Google Scholar] [CrossRef]
- Wang, T.; Li, D.; Tian, X.; Huang, G.; He, M.; Wang, C.; Kumbhar, A.N.; Woldemicael, A.G. Mitigating Salinity Stress through Interactions between Microalgae and Different Forms (Free-Living & Alginate Gel-Encapsulated) of Bacteria Isolated from Estuarine Environments. Sci. Total Environ. 2024, 926, 171909. [Google Scholar] [CrossRef]
- Sarkar, P.; Bandyopadhyay, T.K.; Gopikrishna, K.; Nath Tiwari, O.; Bhunia, B.; Muthuraj, M. Algal Carbohydrates: Sources, Biosynthetic Pathway, Production, and Applications. Bioresour. Technol. 2024, 413, 131489. [Google Scholar] [CrossRef]
- Cichoński, J.; Chrzanowski, G. Microalgae as a Source of Valuable Phenolic Compounds and Carotenoids. Molecules 2022, 27, 8852. [Google Scholar] [CrossRef]
- Haoujar, I.; Cacciola, F.; Chairi, H.; Baltemimi, A.; Essafi, A.; Abrini, J.; Senhaji, N.S. Evaluation of the Antioxidant Properties of Microalgae Naturally Isolated from Mediterranean Morocco. Egypt. J. Aquat. Biol. Fish. 2022, 26, 311. [Google Scholar] [CrossRef]
- Basha, A.N.; Akhir, F.N.M.; Othman, N.; Hara, H. Antioxidant and Anticancer Potential of Bioactive Compounds from Locally Isolated Microalgae. J. Health Qual. Life 2024, 3, 40–54. [Google Scholar] [CrossRef]
- Parveen, A.; Bhatnagar, P.; Gautam, P.; Bisht, B.; Nanda, M.; Kumar, S.; Vlaskin, M.S.; Kumar, V. Enhancing the Bio-Prospective of Microalgae by Different Light Systems and Photoperiods. Photochem. Photobiol. Sci. 2023, 22, 2687–2698. [Google Scholar] [CrossRef]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent Advancements in the Genetic Engineering of Microalgae. Algal Res. 2021, 53, 102158. [Google Scholar] [CrossRef]
- Mourya, M.; Khan, M.J.; Ahirwar, A.; Schoefs, B.; Marchand, J.; Rai, A.; Varjani, S.; Rajendran, K.; Banu, J.R.; Vinayak, V. Latest Trends and Developments in Microalgae as Potential Source for Biofuels: The Case of Diatoms. Fuel 2022, 314, 122738. [Google Scholar] [CrossRef]
- Villanova, V.; Roques, J.A.C.; Forghani, B.; Shaikh, K.M.; Undeland, I.; Spetea, C. Two-Phase Microalgae Cultivation for RAS Water Remediation and High-Value Biomass Production. Front. Plant Sci. 2023, 14, 1186537. [Google Scholar] [CrossRef]
- Ahmad, I.; Abdullah, N.; Ahmad, M.D.; Koji, I.; Yuzir, A. Harnessing Solar Radiation for Potential Algal Biomass Production. In Handbook of Algal Biofuels; Elsevier: Amsterdam, The Netherlands, 2022; pp. 421–449. [Google Scholar] [CrossRef]
- Talebi, A.F.; Tohidfar, M.; Mousavi Derazmahalleh, S.M.; Sulaiman, A.; Baharuddin, A.S.; Tabatabaei, M. Biochemical Modulation of Lipid Pathway in Microalgae Dunaliella Sp. for Biodiesel Production. Biomed Res. Int. 2015, 2015, 597198. [Google Scholar] [CrossRef]
- Rengel, R.; Smith, R.T.; Haslam, R.P.; Sayanova, O.; Vila, M.; León, R. Overexpression of Acetyl-CoA Synthetase (ACS) Enhances the Biosynthesis of Neutral Lipids and Starch in the Green Microalga Chlamydomonas Reinhardtii. Algal Res. 2018, 31, 183–193. [Google Scholar] [CrossRef]
- Tibocha-Bonilla, J.D.; Zuñiga, C.; Godoy-Silva, R.D.; Zengler, K. Advances in Metabolic Modeling of Oleaginous Microalgae Mike Himmel. Biotechnol. Biofuels 2018, 11, 241. [Google Scholar] [CrossRef]
- Daly, G.; Ghini, V.; Adessi, A.; Fondi, M.; Buchan, A.; Viti, C. Towards a Mechanistic Understanding of Microalgae–Bacteria Interactions: Integration of Metabolomic Analysis and Computational Models. FEMS Microbiol. Rev. 2022, 46, fuac020. [Google Scholar] [CrossRef]
- Maria González-González, L.; De-Bashan, L.E.; Khozin-Goldberg, I.; Zilberg, D.; Solovchenko, A. Toward the Enhancement of Microalgal Metabolite Production through Microalgae–Bacteria Consortia. Biology 2021, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Richmond, A.; Hu, Q. Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Gimpel, J.A.; Specht, E.A.; Georgianna, D.R.; Mayfield, S.P. Advances in Microalgae Engineering and Synthetic Biology Applications for Biofuel Production. Curr. Opin. Chem. Biol. 2013, 17, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal Triacylglycerols as Feedstocks for Biofuel Production: Perspectives and Advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Nerantzis, E. Microalgae for High-Value Compounds and Biofuels Production: A Review with Focus on Cultivation under Stress Conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from Microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Sharma, K.K.; Garg, S.; Li, Y.; Malekizadeh, A.; Schenk, P.M. Critical Analysis of Current Microalgae Dewatering Techniques. Biofuels 2013, 4, 397–407. [Google Scholar] [CrossRef]
- Vonshak, A. (Ed.) Spirulina Platensis Arthrospira: Physiology, Cell-Biology And Biotechnology; CRC Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Ravishankar, G.A.; Ambati, R.R. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells 2021, 10, 393. [Google Scholar] [CrossRef]
- Huang, Y.; Lou, C.; Luo, L.; Wang, X.C. Insight into Nitrogen and Phosphorus Coupling Effects on Mixotrophic Chlorella Vulgaris Growth under Stably Controlled Nutrient Conditions. Sci. Total Environ. 2021, 752, 141747. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Z.; Xiao, Y.; Yuan, M.; Zhou, C.; Liu, G.; Fang, J.; Yang, B. Biochemical and Morphological Changes Triggered by Nitrogen Stress in the Oleaginous Microalga Chlorella Vulgaris. Microorganisms 2022, 10, 566. [Google Scholar] [CrossRef]
- Plouviez, M.; Bolot, P.; Shilton, A.; Guieysse, B. Phosphorus Uptake and Accumulation in Chlamydomonas Reinhardtii: Influence of Biomass Concentration, Phosphate Concentration, Phosphorus Depletion Time, and Light Supply. Algal Res. 2023, 71, 103085. [Google Scholar] [CrossRef]
- Medeiros, D.L.; Moreira, Í.T.A. Microalgae Biomass Production from Cultivation in Availability and Limitation of Nutrients: The Technical, Environmental and Economic Performance. J. Clean. Prod. 2022, 370, 133538. [Google Scholar] [CrossRef]
- Abreu, A.P.; Morais, R.C.; Teixeira, J.A.; Nunes, J. A Comparison between Microalgal Autotrophic Growth and Metabolite Accumulation with Heterotrophic, Mixotrophic and Photoheterotrophic Cultivation Modes. Renew. Sustain. Energy Rev. 2022, 159, 112247. [Google Scholar] [CrossRef]
- Dragone, G. Challenges and Opportunities to Increase Economic Feasibility and Sustainability of Mixotrophic Cultivation of Green Microalgae of the Genus Chlorella. Renew. Sustain. Energy Rev. 2022, 160, 112284. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, H.; Sun, Y.; Yan, B.; Chen, W.; Fan, D. The Potential Use of Microalgae for Nutrient Supply and Health Enhancement in Isolated and Confined Environments. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13418. [Google Scholar] [CrossRef]
- da Gama, R.C.N.; Assemany, P.P.; de Assis, L.R.; Oliveira, L.V.; Cecon, P.R.; Calijuri, M.L. Influence of C/N Ratio on Microalgae-Bacteria Joint Culture: Treatment Performance and Phytoplankton Dynamics in Mixed Wastewaters. Bioresour. Technol. Rep. 2023, 23, 101516. [Google Scholar] [CrossRef]
- Roy, U.K.; Wagner, J.; Radu, T. Production of Metabolites in Microalgae Under Alkali Halophilic Growth Medium Using a Dissolved Inorganic Carbon Source. Waste Biomass Valorization 2023, 14, 3339–3354. [Google Scholar] [CrossRef]
- Udayan, A.; Sirohi, R.; Sreekumar, N.; Sang, B.I.; Sim, S.J. Mass Cultivation and Harvesting of Microalgal Biomass: Current Trends and Future Perspectives. Bioresour. Technol. 2022, 344, 126406. [Google Scholar] [CrossRef]
- Filali, R.; Tian, H.; Micheils, E.; Taidi, B. Evaluation of the Growth Performance of Microalgae Based on Fine PH Changes. Austin J. Biotechnol. Bioeng. 2021, 8, 1109. [Google Scholar] [CrossRef]
- Sutherland, D.L.; Howard-Williams, C.; Turnbull, M.H.; Broady, P.A.; Craggs, R.J. The Effects of CO2 Addition along a PH Gradient on Wastewater Microalgal Photo-Physiology, Biomass Production and Nutrient Removal. Water Res. 2015, 70, 9–26. [Google Scholar] [CrossRef]
- Rajanren, J.R.; Ismail, H.M. Investigation of Chlorella Vulgaris Microalgae as a Source for Renewable Fuel. Biofuels 2017, 8, 37–47. [Google Scholar] [CrossRef]
- Saputri, D.A.E.; Prabaningtyas, S.; Aridowi, D. Effect of Temperature Differences on Cell Growth Chlorella Vulgaris Culture by Fixing Bacteria Nitrogen and Producing IAA (Indole Acetic Acid). In AIP Conference Proceedings; AIP Publishing: Melville, NY, USA, 2025; Volume 3186. [Google Scholar] [CrossRef]
- Talaei, M.; Mahdavinejad, M.; Azari, R.; Haghighi, H.M.; Atashdast, A. Thermal and Energy Performance of a User-Responsive Microalgae Bioreactive Façade for Climate Adaptability. Sustain. Energy Technol. Assess. 2022, 52, 101894. [Google Scholar] [CrossRef]
- Soares, L.T.; Vilas-Bôas, R.N.; Mendes, M.F.; de Mendonça, H.V. Assessment of Biofuel Production Yield Using Microalgae Biomass in Cattle Wastewater. Sustain. Water Resour. Manag. 2024, 10, 135. [Google Scholar] [CrossRef]
- Iakovidou, G.; Itziou, A.; Tsiotsias, A.; Lakioti, E.; Samaras, P.; Tsanaktsidis, C.; Karayannis, V. Application of Microalgae to Wastewater Bioremediation, with CO2 Biomitigation, Health Product and Biofuel Development, and Environmental Biomonitoring. Appl. Sci. 2024, 14, 6727. [Google Scholar] [CrossRef]
- Masojídek, J.; Ranglová, K.; Lakatos, G.E.; Benavides, A.M.S.; Torzillo, G. Variables Governing Photosynthesis and Growth in Microalgae Mass Cultures. Processes 2021, 9, 820. [Google Scholar] [CrossRef]
- Rasheed, R.; Schipper, K.; Gifuni, I.; Al-Jabri, H.; Barbosa, M.J.; Gonçalves, O.; Pruvost, J. Thermal Regulation of Algae Cultures in Raceway Ponds Utilizing Ground Heat: Improving Techno-Economic Feasibility and Process Sustainability of Large-Scale Algae Production in Qatar. Sustain. Energy Technol. Assess. 2023, 60, 103497. [Google Scholar] [CrossRef]
- Rodríguez-Miranda, E.; Sánchez-Zurano, A.; Guzmán, J.L.; Acién, G.; Visioli, A. A Seasonal Simulation Approach for Culture Depth Influence on the Temperature for Different Characterized Microalgae Strains. Biotechnol. J. 2022, 17, 2100489. [Google Scholar] [CrossRef]
- Kwan, P.P.; Banerjee, S.; Shariff, M.; Md. Yusoff, F. Influence of Light on Biomass and Lipid Production in Microalgae Cultivation. Aquac. Res. 2021, 52, 1337–1347. [Google Scholar] [CrossRef]
- Rehman, M.; Kesharvani, S.; Dwivedi, G.; Gidwani Suneja, K. Impact of Cultivation Conditions on Microalgae Biomass Productivity and Lipid Content. Mater. Today Proc. 2022, 56, 282–290. [Google Scholar] [CrossRef]
- Sarker, N.K.; Kaparaju, P. A Critical Review on the Status and Progress of Microalgae Cultivation in Outdoor Photobioreactors Conducted over 35 Years (1986–2021). Energies 2023, 16, 3105. [Google Scholar] [CrossRef]
- Köktürk, G.; Ünal, A.; Tokuç, A. Of an Innovative Building-Integrated Photobioreactor. In Exergetic, Energetic and Environmental Dimensions; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Dębowski, M.; Kisielewska, M.; Kazimierowicz, J.; Zieliński, M. Influence of the Light Source on the Chlorella Vulgaris Biomass Growth in the Culture Medium Supplemented with Anaerobic Digestate. Annu. Set Environ. Prot. 2020, 22, 605–621. [Google Scholar]
- De Bhowmick, G.; Sen, R.; Sarmah, A.K. Consolidated Bioprocessing of Wastewater Cocktail in an Algal Biorefinery for Enhanced Biomass, Lipid and Lutein Production Coupled with Efficient CO2 Capture: An Advanced Optimization Approach. J. Environ. Manag. 2019, 252, 109696. [Google Scholar] [CrossRef] [PubMed]
- Ao, X.; Kexin, R.; Jingmiao, Z.; Yun, H.; Xianqing, Z.; Xun, Z.; Qiang, L.; Ao, X.; Kexin, R.; Jingmiao, Z.; et al. Promotion of Carbon Fixation and Emission Reduction by Microalgae with Optical Fiber/Light Guide Tubes. COAL Sci. Technol. 2024, 52, 329–337. [Google Scholar] [CrossRef]
- Chen, Y.; Wan, W.W.; Cui, K.H.; Lau, B.P.Y.; Lee, F.W.F.; Xu, S.J.L. Feasibility and Efficiency of Microalgae Cultivation for Nutrient Recycling and Energy Recovery from Food Waste Filtrate. PLoS ONE 2025, 20, e0315801. [Google Scholar] [CrossRef]
- Contreras-Ropero, J.E.; Lidueñez-Ballesteros, V.S.; Rodríguez-Bohórquez, A.D.; García-Martínez, J.B.; Urbina-Suarez, N.A.; López-Barrera, G.L.; Barajas-Solano, A.F.; Bryan, S.J.; Zuorro, A.; Contreras-Ropero, J.E.; et al. The Effect of LEDs on Biomass and Phycobiliproteins Production in Thermotolerant Oscillatoria Sp. Appl. Sci. 2022, 12, 11664. [Google Scholar] [CrossRef]
- Serôdio, J.; Campbell, D.A. Photoinhibition: Fundamentals and Implications for Primary Productivity. In Life Below Water; Springer: Cham, Switzerland, 2021; pp. 1–13. [Google Scholar] [CrossRef]
- Gao, S.; Edmundson, S.; Huesemann, M. Oxygen Stress Mitigation for Microalgal Biomass Productivity Improvement in Outdoor Raceway Ponds. Algal Res. 2022, 68, 102901. [Google Scholar] [CrossRef]
- Liebscher, L.-J.; Höger, A.-L.; Kleinert, C.; Matthes, S.; Griehl, C.; Ecke, M. Prevention and Control of Parasitic Contamination in Industrial Microalgae Cultures. J. Appl. Phycol. 2024, 37, 1–13. [Google Scholar] [CrossRef]
- Nordio, R.; Belachqer-El Attar, S.; Clagnan, E.; Sánchez-Zurano, A.; Pichel, N.; Viviano, E.; Adani, F.; Guzmán, J.L.; Acién, G. Exploring Microbial Growth Dynamics in a Pilot-Scale Microalgae Raceway Fed with Urban Wastewater: Insights into the Effect of Operational Variables. J. Environ. Manag. 2024, 369, 122385. [Google Scholar] [CrossRef]
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Hong-ying, H.; Ke, G.; Ying-xue, S. Effects of Different Nitrogen and Phosphorus Concentrations on the Growth, Nutrient Uptake, and Lipid Accumulation of a Freshwater Microalga Scenedesmus Sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Stevens, D.; Murray, J.; Diepeveen, D.; Toohey, D.; Stevens, J.D.; Murray, D.; Diepeveen, D.; Toohey, D. Adaptalight: An Inexpensive PAR Sensor System for Daylight Harvesting in a Micro Indoor Smart Hydroponic System. Horticulturae 2022, 8, 105. [Google Scholar] [CrossRef]
- Amaral, R.; Duci, D.; Cotta, F.C.; Bacellar, F.L.; Oliveira, S.; Verret, F.; Asadi, K.; Vandamme, L.K.J.; Reis, N.M.; Bryant, L.D.; et al. Ion-Driven Communication and Acclimation Strategies in Microalgae. Chem. Eng. J. 2023, 473, 144985. [Google Scholar] [CrossRef]
- Chunzhuk, E.A.; Grigorenko, A.V.; Kiseleva, S.V.; Chernova, N.I.; Vlaskin, M.S.; Ryndin, K.G.; Butyrin, A.V.; Ambaryan, G.N.; Dudoladov, A.O. Effects of Light Intensity on the Growth and Biochemical Composition in Various Microalgae Grown at High CO2 Concentrations. Plants 2023, 12, 3876. [Google Scholar] [CrossRef]
- Rögner, M. Photosynthesis: Biotechnological Applications with Microalgae; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2021; pp. 1–310. [Google Scholar] [CrossRef]
- Larochelle, J.; Klueppel, J.; McCormick, R.; Biegert, K.; Comella, L.M. A Low-Power Optical Sensor With Dynamically Adjustable Field of View for Photosynthetically Active Radiation (PAR) Measurement. IEEE Sens. J. 2024, 24, 7711–7728. [Google Scholar] [CrossRef]
- Muñoz, I.L.; Bernard, O. Modeling the Influence of Temperature, Light Intensity and Oxygen Concentration on Microalgal Growth Rate. Processes 2021, 9, 496. [Google Scholar] [CrossRef]
- Saccardo, A.; Bezzo, F.; Sforza, E. Microalgae Growth in Ultra-Thin Steady-State Continuous Photobioreactors: Assessing Self-Shading Effects. Front. Bioeng. Biotechnol. 2022, 10, 977429. [Google Scholar] [CrossRef]
- Ifrim, G.A.; Titica, M.; Deppe, S.; Frahm, B.; Barbu, M.; Caraman, S. Multivariable Control Strategy for the Photosynthetic Cultures of Microalgae. In Proceedings of the 2019 23rd International Conference on System Theory, Control and Computing (ICSTCC), Sinaia, Romania, 9–11 October 2019; pp. 218–223. [Google Scholar] [CrossRef]
- Wolf, J.; Stephens, E.; Steinbusch, S.; Yarnold, J.; Ross, I.L.; Steinweg, C.; Doebbe, A.; Krolovitsch, C.; Müller, S.; Jakob, G.; et al. Multifactorial Comparison of Photobioreactor Geometries in Parallel Microalgae Cultivations. Algal Res. 2016, 15, 187–201. [Google Scholar] [CrossRef]
- Jakubowski, P.; Gryc, I. PL243889B1 System for Measuring Intensity Parameters of Optical Radiation. 2020. Available online: https://worldwide.espacenet.com/patent/search/family/076547986/publication/PL243889B1?q=PL243889B1 (accessed on 12 April 2025).
- Festo GR. PhotoBionicCell. Available online: https://www.festo.com/gr/en/e/about-festo/research-and-development/bionic-learning-network/future-concepts-for-biologisation/photobioniccell-id_1330847/ (accessed on 27 April 2025).
- Nwoba, E.G.; Parlevliet, D.A.; Laird, D.W.; Alameh, K.; Moheimani, N.R. Light Management Technologies for Increasing Algal Photobioreactor Efficiency. Algal Res. 2019, 39, 101433. [Google Scholar] [CrossRef]
- Rossi, S.; Carecci, D.; Ficara, E. Thermal Response Analysis and Compilation of Cardinal Temperatures for 424 Strains of Microalgae, Cyanobacteria, Diatoms and Other Species. Sci. Total Environ. 2023, 873, 162275. [Google Scholar] [CrossRef]
- Legrand, J.; Artu, A.; Pruvost, J. A Review on Photobioreactor Design and Modelling for Microalgae Production. React. Chem. Eng. 2021, 6, 1134–1151. [Google Scholar] [CrossRef]
- Agrawal, R. Fermentor. In Textbook of Industrial Microbiology; Springer: Singapore, 2024; pp. 97–150. [Google Scholar] [CrossRef]
- Paladino, O.; Neviani, M. Scale-up of Photo-Bioreactors for Microalgae Cultivation by π-Theorem. Biochem. Eng. J. 2020, 153, 107398. [Google Scholar] [CrossRef]
- Ng, H.; Tan, C.H.; Nomanbhay, S.; Show, P.L. Sustainability and Development of Microalgae 4.0. In Microalgae for Environmental Biotechnology; CRC Press: Boca Raton, FL, USA, 2022; pp. 397–420. [Google Scholar] [CrossRef]
- Gabrielyan, D.A.; Sinetova, M.A.; Gabrielyan, A.K.; Bobrovnikova, L.A.; Bedbenov, V.S.; Starikov, A.Y.; Zorina, A.A.; Gabel, B.V.; Los, D.A. Laboratory System for Intensive Cultivation of Microalgae and Cyanobacteria. Russ. J. Plant Physiol. 2023, 70, 20. [Google Scholar] [CrossRef]
- Carneiro, M.; Cicchi, B.; Maia, I.B.; Pereira, H.; Zittelli, G.C.; Varela, J.; Malcata, F.X.; Torzillo, G. Effect of Temperature on Growth, Photosynthesis and Biochemical Composition of Nannochloropsis Oceanica, Grown Outdoors in Tubular Photobioreactors. Algal Res. 2020, 49, 101923. [Google Scholar] [CrossRef]
- Shekh, A.; Sharma, A.; Schenk, P.M.; Kumar, G.; Mudliar, S. Microalgae Cultivation: Photobioreactors, CO2 Utilization, and Value-Added Products of Industrial Importance. J. Chem. Technol. Biotechnol. 2022, 97, 1064–1085. [Google Scholar] [CrossRef]
- Hu, Z.; Wu, Y.; Wang, X.; Yu, Z.; Mao, W.; Cheng, C.; Che, G.; Zhao, L.; Li, T.; Yang, W.; et al. Flue Gas Waste Heat Affects Algal Liquid Temperature for Microalgal Production in Column Photobioreactors. Heat Transf. 2024, 53, 2173–2190. [Google Scholar] [CrossRef]
- Meier, L.; Vilchez, C.; Cuaresma, M.; Torres-Aravena, Á.; Jeison, D. Effect of PH Change on the Microalgae-Based Biogas Upgrading Process. Appl. Sci. 2022, 12, 12194. [Google Scholar] [CrossRef]
- Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13061. [Google Scholar] [CrossRef]
- Gao, K. Approaches and Involved Principles to Control PH/PCO2 Stability in Algal Cultures. J. Appl. Phycol. 2021, 33, 3497–3505. [Google Scholar] [CrossRef]
- Savio, S.; Di Natale, C.; Paolesse, R.; Lvova, L.; Congestri, R. Keeping Track of Phaeodactylum Tricornutum (Bacillariophyta) Culture Contamination by Potentiometric E-Tongue. Sensors 2021, 21, 4052. [Google Scholar] [CrossRef]
- Kim, G.Y.; Heo, J.; Kim, K.; Chung, J.; Han, J.I. Electrochemical PH Control and Carbon Supply for Microalgae Cultivation. Chem. Eng. J. 2021, 426, 131796. [Google Scholar] [CrossRef]
- Sandaruwan, H.H.P.B.; Manatunga, D.C.; N. Liyanage, R.; Costha, N.P.; Dassanayake, R.S.; Wijesinghe, R.E.; Zhou, Y.; Liu, Y. Next-Generation Methods for Precise PH Detection in Ocular Chemical Burns: A Review of Recent Analytical Advancements. Anal. Methods 2025, 17, 408–431. [Google Scholar] [CrossRef]
- Caparroz, M.; Guzmán, J.L.; Berenguel, M.; Acién, F.G. A Novel Data-Driven Model for Prediction and Adaptive Control of PH in Raceway Reactor for Microalgae Cultivation. N. Biotechnol. 2024, 82, 1–13. [Google Scholar] [CrossRef]
- Idam, E.I.; Qadeer, A.; Oli, I.C.; Fagorite, V.I.; Baxter, T.; Grischek, T.; Nwobi, N.O.; Onyeagoro, R.U.; Adama, I.O.; Abass, G.F. Optimizing Wastewater Treatment: Algae-Mediated Calcite Formation and Carbon Sequestration through Bicarbonate Control. Algal Res. 2024, 82, 103631. [Google Scholar] [CrossRef]
- Cogne, G.; Ballestas, F.F.; Titica, M.; Legrand, J. A Tool for On-Line Monitoring Microalgal Bioprocesses Based on Gas Balance Analysis. Biotechnol. Bioeng. 2025, 122, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Severo, I.A.; Deprá, M.C.; Barin, J.S.; Wagner, R.; de Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. Bio-Combustion of Petroleum Coke: The Process Integration with Photobioreactors. Chem. Eng. Sci. 2018, 177, 422–430. [Google Scholar] [CrossRef]
- Yang, Y.W.; Li, M.J.; Hung, T.C. The Study on Coupled CO2 Fixation and Power Generation in Microalgae-Microbial Fuel Cells Embedded with Oxygen-Consuming Biofilms. Fuel 2024, 367, 131410. [Google Scholar] [CrossRef]
- Ansari, A.; Verma, N.K.; Saravanan, M.; Verma, N. Selective Conversion of Aqueous CO2 to Formate Using the Wastewater-Isolated Microalage in a Packed-Bed Photobioreactor Under Flow Condition. Available at SSRN 2025. [Google Scholar] [CrossRef]
- Du, N.; Gholami, P.; Kline, D.I.; DuPont, C.L.; Dickson, A.G.; Mendola, D.; Martz, T.; Allen, A.E.; Greg Mitchell, B. Simultaneous Quantum Yield Measurements of Carbon Uptake and Oxygen Evolution in Microalgal Cultures. PLoS ONE 2018, 13, e0199125. [Google Scholar] [CrossRef]
- Hossain, F.M.; Nurun Nabi, M.; Brown, R.J. Investigation of Diesel Engine Performance and Exhaust Emissions of Microalgae Fuel Components in a Turbocharged Diesel Engine. Energy Convers. Manag. 2019, 186, 220–228. [Google Scholar] [CrossRef]
- Dębowski, M.; Krzemieniewski, M.; Zieliński, M.; Kazimierowicz, J. Immobilized Microalgae-Based Photobioreactor for CO2 Capture (IMC-CO2PBR): Efficiency Estimation, Technological Parameters, and Prototype Concept. Atmosphere 2021, 12, 1031. [Google Scholar] [CrossRef]
- Borodin, V.B. Specifics of the Clark-Type O2-Electrodes Application in Biological Studies. Russ. J. Plant Physiol. 2021, 68, 774–782. [Google Scholar] [CrossRef]
- Fallahi Chegeni, N.; Ijadi Maghsoodi, P.; Habibi, M.; Zare-Behtash, H.; Majles Ara, M.H.; Heydari, E. Hybrid Dissolved-Oxygen and Temperature Sensing: A Nanophotonic Probe for Real-Time Monitoring of Chlorella Algae. Sensors 2021, 21, 6553. [Google Scholar] [CrossRef] [PubMed]
- Liao, R.; Li, X.; Yan, M.; Ma, H.; Leung, P.T.Y. Polarimetric Learning: A Siamese Approach to Learning Distance Metrics of Algal Mueller Matrix Images. Appl. Opt. 2018, 57, 3829–3837. [Google Scholar] [CrossRef]
- Rehman, R.; Kazmi, S.F.; Irshad, M.; Bilal, M.; Hafeez, F.; Ahmed, J.; Shaheedi, S.; Nazir, R. Microalgae-Assisted Treatment of Wastewater Originating from Varied Sources, Particularly in the Context of Heavy Metals and Antibiotic-Resistant Bacteria. Water 2024, 16, 3305. [Google Scholar] [CrossRef]
- Abiusi, F.; Wijffels, R.H.; Janssen, M. Oxygen Balanced Mixotrophy under Day-Night Cycles. ACS Sustain. Chem. Eng. 2020, 8, 11682–11691. [Google Scholar] [CrossRef]
- Song, X.; Kong, F.; Liu, B.F.; Song, Q.; Ren, N.Q.; Ren, H.Y. Antioxidants Alleviated Low-Temperature Stress in Microalgae by Modulating Reactive Oxygen Species to Improve Lipid Production and Antioxidant Defense. Bioresour. Technol. 2024, 413, 131451. [Google Scholar] [CrossRef]
- Skok, A.; Bazel, Y.; Vishnikin, A. New Analytical Methods for the Determination of Sulfur Species with Microextraction Techniques: A Review. J. Sulfur Chem. 2022, 43, 443–471. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Alves, M.; Leitão, F.; Tacão, M.; Henriques, I.; Castro, P.M.L.; Amorim, C.L. Bioremediation of Coastal Aquaculture Effluents Spiked with Florfenicol Using Microalgae-Based Granular Sludge—A Promising Solution for Recirculating Aquaculture Systems. Water Res. 2023, 233, 119733. [Google Scholar] [CrossRef]
- González-Camejo, J.; Aparicio, S.; Pachés, M.; Borrás, L.; Seco, A. Comprehensive Assessment of the Microalgae-Nitrifying Bacteria Competition in Microalgae-Based Wastewater Treatment Systems: Relevant Factors, Evaluation Methods and Control Strategies. Algal Res. 2022, 61, 102563. [Google Scholar] [CrossRef]
- Chiellini, C.; Guglielminetti, L.; Pistelli, L.; Ciurli, A. Screening of Trace Metal Elements for Pollution Tolerance of Freshwater and Marine Microalgal Strains: Overview and Perspectives. Algal Res. 2020, 45, 101751. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, J.; Yin, Z.; Liu, Z.; Chen, J.; Xu, J.; Gao, Q.; Liu, J. A Method for Quantifying the Contribution of Algal Sources to CODMn in Water Bodies Based on Ecological Chemometrics and Its Potential Applications. J. Environ. Chem. Eng. 2024, 12, 111943. [Google Scholar] [CrossRef]
- Kumar, D.; Sahoo, D. Assessment of Physiochemical Parameters and Bioremediation of Complex Contaminated Yamuna River, India: An Algal-Based Approach. Water. Air. Soil Pollut. 2024, 235, 90. [Google Scholar] [CrossRef]
- Supraja, K.V.; Behera, B.; Balasubramanian, P. Efficacy of Microalgal Extracts as Biostimulants through Seed Treatment and Foliar Spray for Tomato Cultivation. Ind. Crops Prod. 2020, 151, 112453. [Google Scholar] [CrossRef]
- Maroneze, M.M.; Martinez, A. Minerals and Trace Elements in Microalgal Biomass. In Handbook of Food and Feed from Microalgae; Academic Press: Cambridge, MA, USA, 2023; pp. 103–109. [Google Scholar] [CrossRef]
- Gorbunova, S.Y.; Avsiyan, A.L. Real-Time Automation and Monitoring of the Batch Growth of Microalga Tetraselmis Viridis and Cyanobacterium Limnospira Platensis. Syst. Microbiol. Biomanufacturing 2025, 5, 795–804. [Google Scholar] [CrossRef]
- Solovchenko, A.; Lukyanov, A.; Vasilieva, S.; Lobakova, E. Chlorophyll Fluorescence as a Valuable Multitool for Microalgal Biotechnology. Biophys. Rev. 2022, 14, 973–983. [Google Scholar] [CrossRef]
- Van Wychen, S.; Rowland, S.M.; Lesco, K.C.; Shanta, P.V.; Dong, T.; Laurens, L.M.L. Advanced Mass Balance Characterization and Fractionation of Algal Biomass Composition. J. Appl. Phycol. 2021, 33, 2695–2708. [Google Scholar] [CrossRef]
- Paula, S.F.A.; Chagas, B.M.E.; Pereira, M.; Rangel, A.H.N.; Sassi, C.F.C.; Borba, L.H.F.; Santos, E.S.; Asevedo, E.A.; Câmara, F.R.A.; Araújo, R.M. Pyrolysis-GCMS of Spirulina Platensis: Evaluation of Biomasses Cultivated under Autotrophic and Mixotrophic Conditions. PLoS ONE 2022, 17, e0276317. [Google Scholar] [CrossRef]
- Michalak, I.; Baśladyńska, S.; Mularczyk, M.; Marycz, K. Investigation on the Potential Sorbents—Aluminosilicate, Microalga and Grass Hay as Feed Additives. Environ. Technol. Innov. 2021, 24, 101816. [Google Scholar] [CrossRef]
- Ni, Z.; Tan, L.; Wang, J.; Chen, Y.; Zhang, N.; Meng, F.; Wang, J. Toxic Effects of Pristine and Aged Polystyrene and Their Leachate on Marine Microalgae Skeletonema Costatum. Sci. Total Environ. 2023, 857, 159614. [Google Scholar] [CrossRef]
- Williamson, C.J.; Anesio, A.M.B.; Benning, L.G.; Tranter, M. New Method Provides First Evidence of Fine-Scale in Situ Heterogeneity in Glacier Algal Photophysiology. Eur. J. Phycol. 2025, 60, 35–42. [Google Scholar] [CrossRef]
- Stablein, M.J.; Baracho, D.H.; Watson, J.T.; Silva, J.C.; Zhang, Y.; Lombardi, A.T. Microalgal Photosynthetic Inhibition and Mixotrophic Growth in Post Hydrothermal Liquefaction Wastewater (PHW). Algal Res. 2021, 60, 102548. [Google Scholar] [CrossRef]
- des Aulnois, M.G.; Roux, P.; Caruana, A.; Réveillon, D.; Briand, E.; Hervé, F.; Savar, V.; Bormans, M.; Amzil, Z. Physiological and Metabolic Responses of Freshwater and Brackish-Water Strains of Microcystis Aeruginosa Acclimated to a Salinity Gradient: Insight into Salt Tolerance. Appl. Environ. Microbiol. 2019, 85, e01614-19. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, L.; Pang, T.; Liu, J. Comparative Transcriptome Profiling of Kappaphycus Alvarezii (Rhodophyta, Gigartinales) in Response to Two Extreme Temperature Treatments: An RNA-Seq-Based Resource for Photosynthesis Research. Eur. J. Phycol. 2019, 54, 162–174. [Google Scholar] [CrossRef]
- Hupp, J.; McCoy, J.I.E.; Millgan, A.J.; Peers, G. Simultaneously Measuring Carbon Uptake Capacity and Chlorophyll a Fluorescence Dynamics in Algae. Algal Res. 2021, 58, 102399. [Google Scholar] [CrossRef]
- Derksen, G.C.H.; Blommaert, L.; Bastiaens, L.; Hasşerbetçi, C.; Fremouw, R.; van Groenigen, J.; Twijnstra, R.H.; Timmermans, K.R. ATR-FTIR Spectroscopy Combined with Multivariate Analysis as a Rapid Tool to Infer the Biochemical Composition of Ulva Laetevirens (Chlorophyta). Front. Mar. Sci. 2023, 10, 1154461. [Google Scholar] [CrossRef]
- Ferro, L.; Gojkovic, Z.; Gorzsás, A.; Funk, C. Statistical Methods for Rapid Quantification of Proteins, Lipids, and Carbohydrates in Nordic Microalgal Species Using ATR–FTIR Spectroscopy. Molecules 2019, 24, 3237. [Google Scholar] [CrossRef]
- Dammak, M.; Ben Hlima, H.; Elleuch, F.; Pichon, C.; Michaud, P.; Fendri, I.; Abdelkafi, S. Flow Cytometry Assay to Evaluate Lipid Production by the Marine Microalga Tetraselmis Sp. Using a Two Stage Process. Renew. Energy 2021, 177, 280–289. [Google Scholar] [CrossRef]
- Satpati, G.G.; Pal, R. Rapid Detection of Neutral Lipid in Green Microalgae by Flow Cytometry in Combination with Nile Red Staining—An Improved Technique. Ann. Microbiol. 2015, 65, 937–949. [Google Scholar] [CrossRef]
- Havlik, I.; Reardon, K.F.; Ünal, M.; Lindner, P.; Prediger, A.; Babitzky, A.; Beutel, S.; Scheper, T. Monitoring of Microalgal Cultivations with On-Line, Flow-through Microscopy. Algal Res. 2013, 2, 253–257. [Google Scholar] [CrossRef]
- Connan, S. Spectrophotometric Assays of Major Compounds Extracted from Algae. Methods Mol. Biol. 2015, 1308, 75–101. [Google Scholar] [CrossRef]
- Yeh, Y.C.; Ebbing, T.; Frick, K.; Schmid-Staiger, U.; Haasdonk, B.; Tovar, G.E.M. Improving Determination of Pigment Contents in Microalgae Suspension with Absorption Spectroscopy: Light Scattering Effect and Bouguer–Lambert–Beer Law. Mar. Drugs 2023, 21, 619. [Google Scholar] [CrossRef] [PubMed]
- Salgueiro, J.L.; Pérez, L.; Sanchez, Á.; Cancela, Á.; Míguez, C. Microalgal Biomass Quantification from the Non-Invasive Technique of Image Processing through Red–Green–Blue (RGB) Analysis. J. Appl. Phycol. 2022, 34, 871–881. [Google Scholar] [CrossRef]
- Sarrafzadeh, M.H.; La, H.J.; Lee, J.Y.; Cho, D.H.; Shin, S.Y.; Kim, W.J.; Oh, H.M. Microalgae Biomass Quantification by Digital Image Processing and RGB Color Analysis. J. Appl. Phycol. 2015, 27, 205–209. [Google Scholar] [CrossRef]
- White, S.; Anandraj, A.; Bux, F. PAM Fluorometry as a Tool to Assess Microalgal Nutrient Stress and Monitor Cellular Neutral Lipids. Bioresour. Technol. 2011, 102, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Thiviyanathan, V.A.; Ker, P.J.; Amin, E.P.P.; Tang, S.G.H.; Yee, W.; Jamaludin, M.Z. Quantifying Microalgae Growth by the Optical Detection of Glucose in the NIR Waveband. Molecules 2023, 28, 1318. [Google Scholar] [CrossRef] [PubMed]
- Koczoń, P.; Hołaj-Krzak, J.T.; Palani, B.K.; Bolewski, T.; Dąbrowski, J.; Bartyzel, B.J.; Gruczyńska-Sękowska, E. The Analytical Possibilities of FT-IR Spectroscopy Powered by Vibrating Molecules. Int. J. Mol. Sci. 2023, 24, 1013. [Google Scholar] [CrossRef]
- Esther Elizabeth Grace, C.; Briget Mary, M.; Vaidyanathan, S.; Srisudha, S. Response to Nutrient Variation on Lipid Productivity in Green Microalgae Captured Using Second Derivative FTIR and Raman Spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 270, 120830. [Google Scholar] [CrossRef]
- Zamanileha, E.F.; Mitantsoa, J.T.; Vaonalamihanta, P.F.; Razafintsalama, A.R.; Andrianony, F.A.; Ravelonandro, P.H.; Zamanileha, E.F.; Mitantsoa, J.T.; Vaonalamihanta, P.F.; Razafintsalama, A.R.; et al. Various Approaches to Fourier-Transform Infrared Spectroscopy (FTIR) for Bioanalytical and Biotechnological Applications in Marine Algae. In Recent Advances in Infrared Spectroscopy and Its Applications in Biotechnology; IntechOpen: London, UK, 2024. [Google Scholar] [CrossRef]
- Samokhvalov, A.; McCombs, S. In Situ Time-Dependent Attenuated Total Reflection Fourier Transform Infrared (ATR FT-IR) Spectroscopy of a Powdered Specimen in a Controlled Atmosphere: Monitoring Sorption and Desorption of Water Vapor. Appl. Spectrosc. 2023, 77, 308–319. [Google Scholar] [CrossRef]
- Havlik, I.; Lindner, P.; Scheper, T.; Reardon, K.F. On-Line Monitoring of Large Cultivations of Microalgae and Cyanobacteria. Trends Biotechnol. 2013, 31, 406–414. [Google Scholar] [CrossRef]
- Liu, J.Y.; Zeng, L.H.; Ren, Z.H. Recent Application of Spectroscopy for the Detection of Microalgae Life Information: A Review. Appl. Spectrosc. Rev. 2020, 55, 26–59. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Karpiński, T.M. Live-Cell Systems in Real-Time Biomonitoring of Water Pollution: Practical Considerations and Future Perspectives. Sensors 2021, 21, 7028. [Google Scholar] [CrossRef] [PubMed]
- Harshkova, D.; Zielińska, E.; Aksmann, A. Optimization of a Microplate Reader Method for the Analysis of Changes in Mitochondrial Membrane Potential in Chlamydomonas Reinhardtii Cells Using the Fluorochrome JC-1. J. Appl. Phycol. 2019, 31, 3691–3697. [Google Scholar] [CrossRef]
- Howell, J.; Omwenga, S.; Jimenez, M.; Hammarton, T.C. Analysis of the Leishmania Mexicana Promastigote Cell Cycle Using Imaging Flow Cytometry Provides New Insights into Cell Cycle Flexibility and Events of Short Duration. PLoS ONE 2024, 19, e0311367. [Google Scholar] [CrossRef] [PubMed]
- Ozdalgic, B.; Ustun, M.; Dabbagh, S.R.; Haznedaroglu, B.Z.; Kiraz, A.; Tasoglu, S. Microfluidics for Microalgal Biotechnology. Biotechnol. Bioeng. 2021, 118, 1716–1734. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I. Photosynthesis: New Approaches to the Molecular, Cellular, and Organismal Levels; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 1–386. [Google Scholar] [CrossRef]
- Taborda, T.; Moniz, P.; Reis, A.; da Silva, T.L. Evaluating Low-Cost Substrates for Crypthecodinium Cohnii Lipids and DHA Production, by Flow Cytometry. J. Appl. Phycol. 2021, 33, 263–274. [Google Scholar] [CrossRef]
- Shan Ahamed, T.; Brindhadevi, K.; Krishnan, R.; Phuong, T.N.; Ali Alharbi, S.; Chinnathambi, A.; Mathimani, T. Invivo Detection of Triacylglycerols through Nile Red Staining and Quantification of Fatty Acids in Hyper Lipid Producer Nannochloropsis sp. Cultured under Adequate Nitrogen and Deficient Nitrogen Condition. Fuel 2022, 322, 124179. [Google Scholar] [CrossRef]
- Du, X.; Chen, X.; Gao, C.; Wang, J.; Huo, X.; Chen, J. Recent Developments (After 2020) in Flow Cytometry Worldwide and Within China. Biosensors 2025, 15, 156. [Google Scholar] [CrossRef] [PubMed]
- Broger, T.; Odermatt, R.P.; Huber, P.; Sonnleitner, B. Real-Time on-Line Flow Cytometry for Bioprocess Monitoring. J. Biotechnol. 2011, 154, 240–247. [Google Scholar] [CrossRef]
- Tamayol, A.; Arab-Tehrany, E.; Desobry, S.; Imamoglu, E. Artificial Intelligence and/or Machine Learning Algorithms in Microalgae Bioprocesses. Bioeng. 2024, 11, 1143. [Google Scholar] [CrossRef]
- Oruganti, R.K.; Biji, A.P.; Lanuyanger, T.; Show, P.L.; Sriariyanun, M.; Upadhyayula, V.K.K.; Gadhamshetty, V.; Bhattacharyya, D. Artificial Intelligence and Machine Learning Tools for High-Performance Microalgal Wastewater Treatment and Algal Biorefinery: A Critical Review. Sci. Total Environ. 2023, 876, 162797. [Google Scholar] [CrossRef]
- Bilal, A.; Imran, A.; Baig, T.I.; Liu, X.; Abouel Nasr, E.; Long, H. Breast Cancer Diagnosis Using Support Vector Machine Optimized by Improved Quantum Inspired Grey Wolf Optimization. Sci. Rep. 2024, 14, 10714. [Google Scholar] [CrossRef] [PubMed]
- Mafat, I.H.; Palla, S.; Surya, D.V. Machine Learning and Artificial Intelligence for Algal Cultivation, Harvesting Techniques, Wastewater Treatment, Nutrient Recovery, and Biofuel Production and Optimization. In Value Added Products From Bioalgae Based Biorefineries: Opportunities and Challenges; Springer: Singapore, 2024; pp. 463–487. [Google Scholar] [CrossRef]
- Lim, H.R.; Khoo, K.S.; Chia, W.Y.; Chew, K.W.; Ho, S.H.; Show, P.L. Smart Microalgae Farming with Internet-of-Things for Sustainable Agriculture. Biotechnol. Adv. 2022, 57, 107931. [Google Scholar] [CrossRef]
- Abdullah, M.; Malik, H.A.; Ali, A.; Boopathy, R.; Vo, P.H.N.; Danaee, S.; Ralph, P.; Malik, S. AI-Driven Algae Biorefineries: A New Era for Sustainable Bioeconomy. Curr. Pollut. Reports 2025, 11, 1–24. [Google Scholar] [CrossRef]
- Maraveas, C.; Bartzanas, T. Application of Internet of Things (IoT) for Optimized Greenhouse Environments. AgriEngineering 2021, 3, 954–970. [Google Scholar] [CrossRef]
- Borah, D.; Eldiehy, K.S.H.; Hatiboruah, D.; Mandal, M.; Deka, D. An Integrated Approach for Simultaneous Monitoring and Data Acquisition on the Culture of Green Microalga Chlorella Homosphaera Using Different LED Illumination. Bioenergy Res. 2023, 16, 601–610. [Google Scholar] [CrossRef]
- Strachan, S.; Chakraborty, M.; Sallam, M.; Bhuiyan, S.A.; Ford, R.; Nguyen, N.T. Maximising Affordability of Real-Time Colorimetric LAMP Assays. Micromachines 2023, 14, 2101. [Google Scholar] [CrossRef] [PubMed]
- Mohagheghi, A.; Moallem, M. Measuring Photosynthetic Photon Flux Density in the Blue and Red Spectrum for Horticultural Lighting Using Machine Learning Methods. IEEE Trans. Instrum. Meas. 2024, 73, 2501410. [Google Scholar] [CrossRef]
- Behzadipour, F.; Ghasemi Nezhad Raeini, M.; Abdanan Mehdizadeh, S.; Taki, M.; Khalil Moghadam, B.; Zare Bavani, M.R.; Lloret, J. A Smart IoT-Based Irrigation System Design Using AI and Prediction Model. Neural Comput. Appl. 2023, 35, 24843–24857. [Google Scholar] [CrossRef]
- Anggraini, L.E.K.; Kumi, J.A.; Karolina, L.B.; Akansah, E.; Sulyman, H.A.; Mendonça, I.; Aritsugi, M.; Elikem, E.; Senoo, K.; Anggraini, L.; et al. IoT Solutions with Artificial Intelligence Technologies for Precision Agriculture: Definitions, Applications, Challenges, and Opportunities. Electronics 2020, 13, 1894. [Google Scholar] [CrossRef]
- Gincley, B.; Khan, F.; Hartnett, E.; Fisher, A.; Pinto, A.J. Introducing ARTiMiS: A Low-Cost Flow Imaging Microscope for Phytoplankton Monitoring in Engineered and Natural Environments. bioRxiv 2024. [Google Scholar] [CrossRef]
- D’Adamo, S.; Schiano di Visconte, G.; Lowe, G.; Szaub-Newton, J.; Beacham, T.; Landels, A.; Allen, M.J.; Spicer, A.; Matthijs, M. Engineering the Unicellular Alga Phaeodactylum Tricornutum for High-Value Plant Triterpenoid Production. Plant Biotechnol. J. 2019, 17, 75–87. [Google Scholar] [CrossRef]
- Faulkner, M.; Andrews, F.; Scrutton, N. Improving Productivity of Citramalate from CO2 by Synechocystis sp. PCC 6803 through Design of Experiment. Biotechnol. Biofuels Bioprod. 2024, 17, 143. [Google Scholar] [CrossRef] [PubMed]
- Karlický, V.; Kmecová Materová, Z.; Kurasová, I.; Nezval, J.; Štroch, M.; Garab, G.; Špunda, V. Accumulation of Geranylgeranylated Chlorophylls in the Pigment-Protein Complexes of Arabidopsis Thaliana Acclimated to Green Light: Effects on the Organization of Light-Harvesting Complex II and Photosystem II Functions. Photosynth. Res. 2021, 149, 233–252. [Google Scholar] [CrossRef]
- Bradley, T.; Rajaeifar, M.A.; Kenny, A.; Hainsworth, C.; del Pino, V.; del Valle Inclán, Y.; Povoa, I.; Mendonça, P.; Brown, L.; Smallbone, A.; et al. Life Cycle Assessment of Microalgae-Derived Biodiesel. Int. J. Life Cycle Assess. 2023, 28, 590–609. [Google Scholar] [CrossRef]
- Carnovale, G.; Rosa, F.; Shapaval, V.; Dzurendova, S.; Kohler, A.; Wicklund, T.; Horn, S.J.; Barbosa, M.J.; Skjånes, K. Starch Rich Chlorella Vulgaris: High-Throughput Screening and Up-Scale for Tailored Biomass Production. Appl. Sci. 2021, 11, 9025. [Google Scholar] [CrossRef]
- Uguz, S.; Sahin, Y.S.; Kumar, P.; Yang, X.; Anderson, G. Real-Time Algal Monitoring Using Novel Machine Learning Approaches. Big Data Cogn. Comput. 2025, 9, 153. [Google Scholar] [CrossRef]
- Szymoniak, S.; Kuczy’nski, Ł.; Kuczy’nski, K. Overview of Modern Technologies for Acquiring and Analysing Acoustic Information Based on AI and IoT. Appl. Sci. 2025, 15, 6690. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Application Area | Process | Effect | Species | References |
|---|---|---|---|---|
| Environmental Engineering and Protection | Wastewater treatment | N, P, COD, micropollutant removal | Chlorella vulgaris, Chlorella protothecoides, Scenedesmus obliquus, Botryococcus braunii | [32,33,34] |
| Leachate and digestate treatment | Chlorella sp., Chlorella vulgaris, Anabaena sp., Scenedesmus sp. | [24,35,36] | ||
| Flue gas treatment | NOx, SOx, fly ash removal | Chlorella vulgaris, Scenedesmus dimorphus, Spirulina platensis | [23,37,38] | |
| CO2 biosequestration | CO2 removal | Nannochloropsis oculata, Chlorella pyrenoidosa, Scenedesmus obliquus, Phaeodactylum tricornutum, and Chlamydomonas reinhardtii | [27,39,40] | |
| Bioenergy | Transesterification | Biodiesel | Nannochloropsis sp., Chlorella vulgaris, Botryococcus braunii | [41,42,43] |
| Alcoholic fermentation | Bioethanol | Chlorella vulgaris, Spirulina platensis | [44,45,46] | |
| Photodissociation | Biohydrogen | Chlamydomonas reinhardtii, Scenedesmus obliquus | [47,48,49] | |
| Anaerobic digestion | Biogas | Chlorella vulgaris, Chlorella sp., Nannochloropsis sp. Scenedesmus sp., Pinnularia sp. | [50,51,52] | |
| Biomethane | Chlorella vulgaris, Chlorella protothecoides, | [53,54,55] | ||
| Biohydrogen | Chlorella sp., Tetraselmis subcordiformis | [56,57,58] | ||
| Gasification | Syngas | Chlorella vulgaris, Galdieria sulphuraria | [59,60,61] | |
| Pyrolysis | Biochar | Chlorella sp., Chlorella pyrenoidosa, Spirogyra sp., Cladophora sp., Microspora, Rhizoclonium | [62,63,64] | |
| Oil | Nannochloropsis sp., Arthrospira platensis, Chlorella sp., Scenedesmus obliquus | [65,66,67] | ||
| Pyrolysis gas | Chlorella vulgaris, Arthrospira platensis, Tetradesmus obliquus, Nannochloropsis sp. | [68,69] | ||
| Combustion | Heat, Electricity | Chlorella vulgaris, Arthrospira platensis, Nannochloropsis sp., Tetraselmis sp. | [70,71,72] |
| Application Area | Process | Final Product (Outcome) | Species | References |
|---|---|---|---|---|
| Pharmacy, Cosmetology, Functional Foods, Dietary Supplements, Nutraceuticals, Biomaterials | Pigment extraction (water, alcohol) | Pigments (phycocyanin, chlorophylls, lutein, astaxanthin) | Spirulina platensis, Dunaliella salina, Haematococcus pluvialis | [82,83,84] |
| Lipid extraction | DHA, EPA (omega-3 fatty acids) | Schizochytrium sp., Nannochloropsis sp., Isochrysis galbana | [85,86,87] | |
| Vitamin extraction | Vitamins A, B, C | Chlorella vulgaris, Spirulina platensis | [88,89] | |
| Protein and peptide extraction | Bioactive proteins and amino acids | Chlorella vulgaris, Arthrospira platensis, Tetraselmis chui | [90,91,92] | |
| Polysaccharide and EPS extraction | Carrageenan, alginates, agar, EPS (exopolysaccharides) | Porphyridium cruentum, Chlorella sp., Phaeodactylum tricornutum | [93,94,95] | |
| Secondary metabolite extraction | Sterols, tocopherols, flavonoids, antioxidants | Tetraselmis suecica, Phaeodactylum tricornutum, Scenedesmus sp. | [96,97,98] |
| Optimisation Approach | Species | Achieved Outcome | References |
|---|---|---|---|
| Monitoring and maintenance of proper incubation conditions | Chlorella vulgaris, Nannochloropsis sp., Scenedesmus obliquus | Increased growth and lipid accumulation efficiency through optimisation of pH (6.5–8.5), temperature (20–30 °C), light intensity, and aeration | [109,110] |
| Genetic engineering | Chlamydomonas reinhardtii, Nannochloropsis gaditana, Phaeodactylum tricornutum | Enhanced lipid synthesis via metabolic pathway manipulation (e.g., ACCase overexpression), increased protein content in biomass, improved resistance to environmental stress | [111,112] |
| Substrate stress (nutrient limitation) | Scenedesmus obliquus, Dunaliella salina, Chlorella vulgaris | Induced lipid accumulation (nitrogen limitation), enhanced carotenoid production (salt stress, high light intensity), increased starch production (phosphorus limitation) | [112,113] |
| Design and equipment of PBRs | Spirulina platensis, Chlorella vulgaris, Haematococcus pluvialis | Improved mixing, intensified gas exchange, greater light absorption surface; increased astaxanthin production in airlift and tubular PBRs | [110,114] |
| Mathematical modelling and cultivation system dynamics | Nannochloropsis sp., Chlorella vulgaris, Tetraselmis suecica | Prediction of optimal cultivation conditions, reduced operational costs, improved growth efficiency through modelling of light access, CO2 concentration, and nutrient availability | [110,115] |
| Artificial intelligence (AI) and control systems | Scenedesmus obliquus, Chlorella vulgaris, Dunaliella salina | Automated control of cultivation parameters (pH, temperature, light, CO2) using neural networks and optimisation algorithms; adaptive strategies for regulation of growth conditions | [110] |
| Consortia with other microorganisms (bacteria, fungi, cyanobacteria) | Chlorella vulgaris + Azospirillum sp., Scenedesmus sp. + fungi Trichoderma sp., Spirulina platensis + probiotic bacteria | Improved bioavailability of nutrients, increased synthesis of bioactive compounds, more efficient CO2 removal, enhanced use of organic waste for biomass production | [109,113] |
| Modification of light spectrum | Haematococcus pluvialis, Dunaliella salina, Chlorella vulgaris | Use of LEDs (red and blue spectrum) for photosynthesis optimisation, increased production of astaxanthin, beta-carotene, and lipids | [110,113] |
| Bioremediation and utilisation of industrial waste | Chlorella vulgaris, Scenedesmus obliquus, Nannochloropsis sp. | Efficient nitrogen and phosphorus assimilation from wastewater, utilisation of flue gas as a CO2 source, improved cost-effectiveness of cultivation | [113,114] |
| Cultivation under extreme conditions | Dunaliella salina, Galdieria sulphuraria, Cyanidium caldarium | Cultivation under extreme salinity and high temperature, enhanced carotenoid accumulation, increased biomass production under reduced competition conditions | [110,116] |
| Parameter | Value | Microalgae Species | References |
|---|---|---|---|
| Nitrogen (N) | 75–300 mg/L (NO3−, NH4+, mocznik) | Chlorella vulgaris | [109,151] |
| 50–500 mg/L (NO3−) | Nannochloropsis sp. | [152] | |
| 100–250 mg/L (NH4+, NO3−) | Scenedesmus obliquus | [153] | |
| 100–400 mg/L (NO3−, mocznik) | Spirulina platensis | [116] | |
| Phosphorus (P) | 5–50 mg/L (PO43−) | Chlorella vulgaris | [151] |
| 1–20 mg/L (PO43−) | Nannochloropsis sp. | [152] | |
| 2–30 mg/L (PO43−) | Scenedesmus obliquus | [153] | |
| 5–25 mg/L (PO43−) | Spirulina platensis | [116] | |
| Microelements | Fe: 0.1–5 mg/L, Zn: 0.01–0.5 mg/L, Mn, Cu, Mo, B | Most species | [112,151] |
| Light | 50–300 µmol m−2 s−1 | Chlorella vulgaris, Scenedesmus obliquus | [109,151] |
| 100–500 µmol m−2 s−1 | Dunaliella salina, Nannochloropsis sp. | [151,152] | |
| 30–150 µmol m−2 s−1 | Spirulina platensis | [116] | |
| pH | 6.5–8.5 | Chlorella vulgaris | [109,151] |
| 7.5–8.5 | Nannochloropsis sp. | [152] | |
| 6.5–9.0 | Scenedesmus obliquus | [153] | |
| 9.0–10.5 | Spirulina platensis | [116] | |
| Temperature | 20–30 °C | Chlorella vulgaris, Scenedesmus obliquus | [109] |
| 18–28 °C | Nannochloropsis sp. | [152] | |
| 25–38 °C | Spirulina platensis | [116] | |
| 25–40 °C | Dunaliella salina | [151] | |
| CO2 concentration | 0.03–2% | Chlorella vulgaris, Nannochloropsis sp. | [112,151] |
| 0.03–5% | Scenedesmus obliquus | [112,153] | |
| 0.03–1% | Spirulina platensis | [116] | |
| O2 concentration | <200% saturation | most species | [112,151] |
| Monitored Variable | Sensor Type | Acceptable Range | Out-of-Range Consequence | Control Options | Literature |
|---|---|---|---|---|---|
| Photon flux density | Quantum sensor, PAR meter | 10–250 µmol/m2/s (optimal) 0–2000 µmol/m2/s (actual) | Low: slow growth High: photoinhibition | PBR design, mixing | [109,110] |
| Temperature | Thermoelement (Pt-100) | 15–35 °C | Low: slow growth High: culture death | Cooling/heating systems | [110,114] |
| pH | pH electrode, optical sensor | 7–10 | Growth rate decrease | CO2 injection | [109,110] |
| pO2 (liquid) | DO electrode, optical sensor | <15–25 mg/L | High: growth rate decrease | Aeration | [110,112] |
| O2 (gas) | Paramagnetic analyser | Depends on mixing | High: growth rate decrease | - | [112,115] |
| pCO2 (liquid) | pCO2 electrode, IR analyser | >0.1 kPa | Low: growth rate decreases below 0.1 kPa | CO2 injection | [112,114] |
| CO2 (gas) | IR analyser | >0.15% | Low: growth rate decreases | CO2 feed | [110,112] |
| Inorganic nutrients | UV spectroscopy, ion-selective | Varies with nutrient | Low: growth limitation; lipid or starch accumulation | Nutrient addition | [109,113] |
| Mixing | None | Re <6000–10,000 | Low: CO2 limitation/O2 inhibition High: mechanical damage to cells | Gas addition rate; agitation intensity | [110,115] |
| Monitoring Method | Monitored Variable (Concentration) | Sensor Type | Off-Line/On-Line | Refs. |
|---|---|---|---|---|
| IR radiation (MIR, NIR, FTIR) | Lipid Protein Carbohydrate content | ATR flow system Fibre-optic probe | Off-line and on-line | [216,217] |
| FC | Lipid content Cell size | Flow cytometer | Off-line and on-line | [218,219] |
| ISM | Cell number concentration Cell mass concentration Cell morphology Population composition (contamination) | Microscope + CCD | On-line | [220] |
| Absorbance spectrum | Pigments Fatty acids | Spectrophotometer | Off-line | [221,222] |
| OD, turbidity | Cell mass concentration | OD sensor Turbidity sensor | Off-line and on-line | [19,222] |
| Colour analysis | Cell mass concentration | CCD camera | Off-line | [223,224] |
| Fluorometry | Photosynthetic efficiency Quantum yield Lipids Pigments | Pulse amplitude modulated fluorometer | Off-line and on-line | [225] |
| Criterion | Category | Examples of Tools/Technologies |
|---|---|---|
| Type of monitored parameters | Physicochemical | pH, temperature, CO2, O2 sensors; light intensity; turbidity sensors |
| Biological | Flow cytometry, fluorometry, in situ microscopy, chlorophyll analysis | |
| Optical | RGB imaging, UV-VIS, FTIR/NIR spectroscopy, image-based diagnostics | |
| Multiparameter integrated systems | IoT platforms, sensor arrays, software-based sensors | |
| Technology readiness level | TRL 1–3: early-stage research | Advanced spectroscopy, online flow cytometry, AI-based diagnostics |
| TRL 4–6: prototyping and pilot testing | Hybrid AI-IoT platforms, semi-automated monitoring systems | |
| TRL 7–9: mature and deployable technologies | Basic sensors for pH, temperature, light, turbidity | |
| Cost and accessibility | Low-cost | RGB cameras, basic environmental sensors (pH, temperature, light) |
| Mid-cost | Fluorometers, UV-VIS, PAR sensors | |
| High-cost | Flow cytometers, FTIR/NIR spectroscopy, AI-integrated platforms |
| Framework Layer | Functional Description | Key Technologies/Methods | Operational Objective |
|---|---|---|---|
| Real-Time Monitoring | Continuous tracking of environmental and cultural parameters | pH, temperature, CO2/O2 sensors, turbidity, RGB, NIR sensors | Early detection of process deviations; stability maintenance |
| AI-Based Optimisation | Dynamic adjustment of cultivation conditions using real-time and historical data | Machine learning algorithms, predictive models, metaheuristics | Maximisation of growth rate and metabolite synthesis |
| Data Integration and Analysis | Cross-sensor data integration and visualisation for process insight | IoT systems, cloud analytics, digital dashboards, process control software | Centralised control and long-term decision support |
| Focus Area | Recommended Future Actions |
|---|---|
| Standardisation and benchmarking | Develop unified protocols to evaluate and compare the performance of various monitoring systems under real conditions |
| Sensor integration and interoperability | Design modular, compatible monitoring platforms for multiparameter acquisition |
| AI and adaptive prediction models | Develop predictive tools tailored to strain-specific responses and environmental fluctuations |
| Cost-efficiency and scalability | Perform techno-economic assessments to guide industrial implementation and scaling decisions |
| Open-access data management | Establish open databases and IoT-based platforms for data sharing, visualisation, and collaborative optimisation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Multi-Sensing Monitoring of the Microalgae Biomass Cultivation Systems for Biofuels and Added Value Products Synthesis—Challenges and Opportunities. Appl. Sci. 2025, 15, 7324. https://doi.org/10.3390/app15137324
Dębowski M, Kazimierowicz J, Zieliński M. Multi-Sensing Monitoring of the Microalgae Biomass Cultivation Systems for Biofuels and Added Value Products Synthesis—Challenges and Opportunities. Applied Sciences. 2025; 15(13):7324. https://doi.org/10.3390/app15137324
Chicago/Turabian StyleDębowski, Marcin, Joanna Kazimierowicz, and Marcin Zieliński. 2025. "Multi-Sensing Monitoring of the Microalgae Biomass Cultivation Systems for Biofuels and Added Value Products Synthesis—Challenges and Opportunities" Applied Sciences 15, no. 13: 7324. https://doi.org/10.3390/app15137324
APA StyleDębowski, M., Kazimierowicz, J., & Zieliński, M. (2025). Multi-Sensing Monitoring of the Microalgae Biomass Cultivation Systems for Biofuels and Added Value Products Synthesis—Challenges and Opportunities. Applied Sciences, 15(13), 7324. https://doi.org/10.3390/app15137324