


Valorisation of Waste Oils Through Oleaginous Yarrowia lipolytica Yeast: Insights into Lipid Stability and Nutritive Properties of Lipid-Rich Biomass

 ,
,  ,
,  , , and
, , and
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Yeast Cultures and Biomass Treatments
2.3. Water Composition Analysis
2.4. Determination of Lipid Content in Yeast Cells and Content of Residual Oil in Medium
2.5. Protein Analysis by the Kjeldahl Method
2.6. Determination of the Microbial Kinetics
2.7. Determination of Fatty Acid Composition
2.8. Oxidative Stability Analysis of Microbial Oil
2.9. Determination of the Number of Bacteria and Yeast
2.10. Statistical Analysis
3. Results
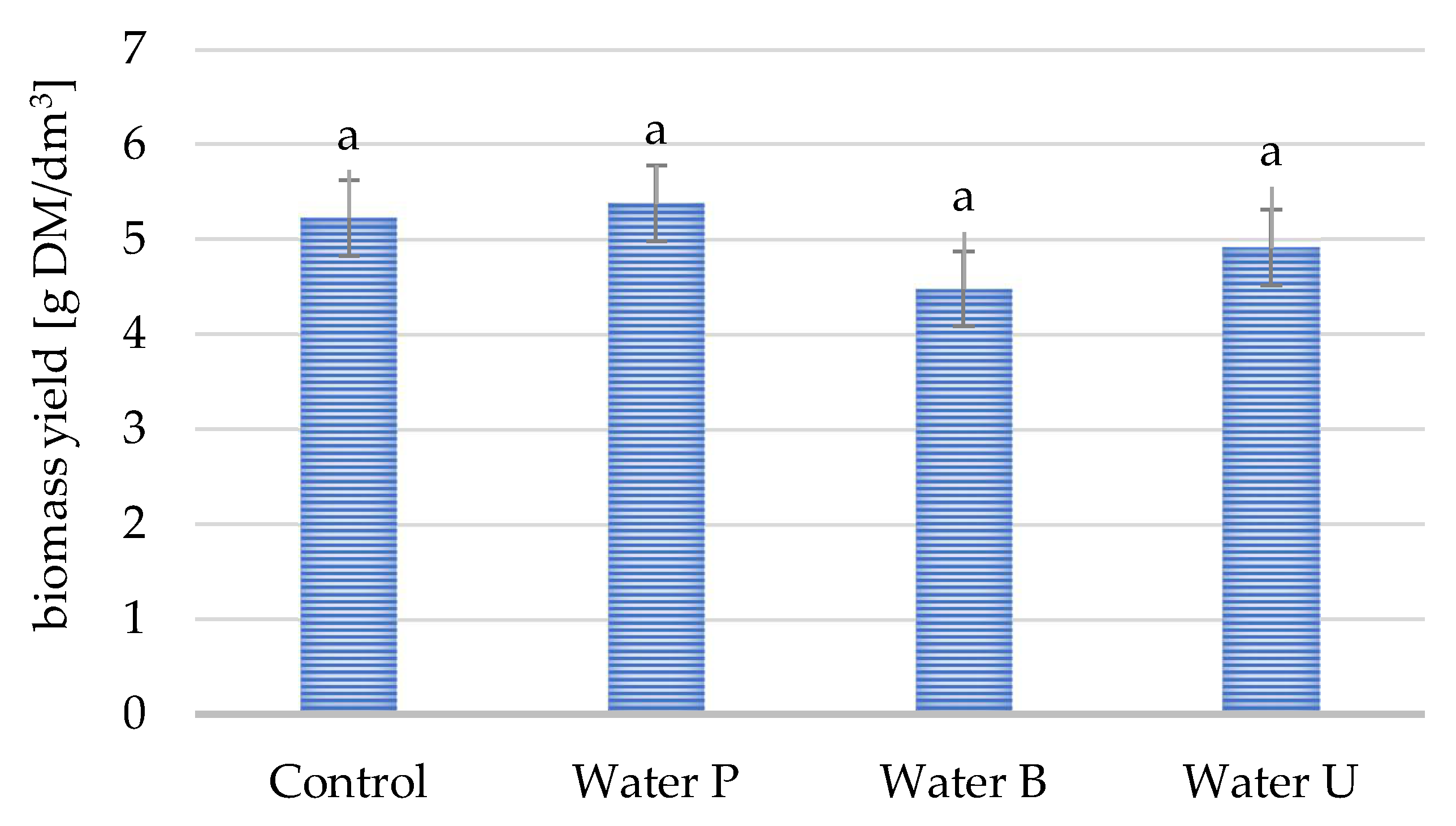
3.1. Impact of Tap Water Quality on Yeast Biomass Yield and Lipid Content
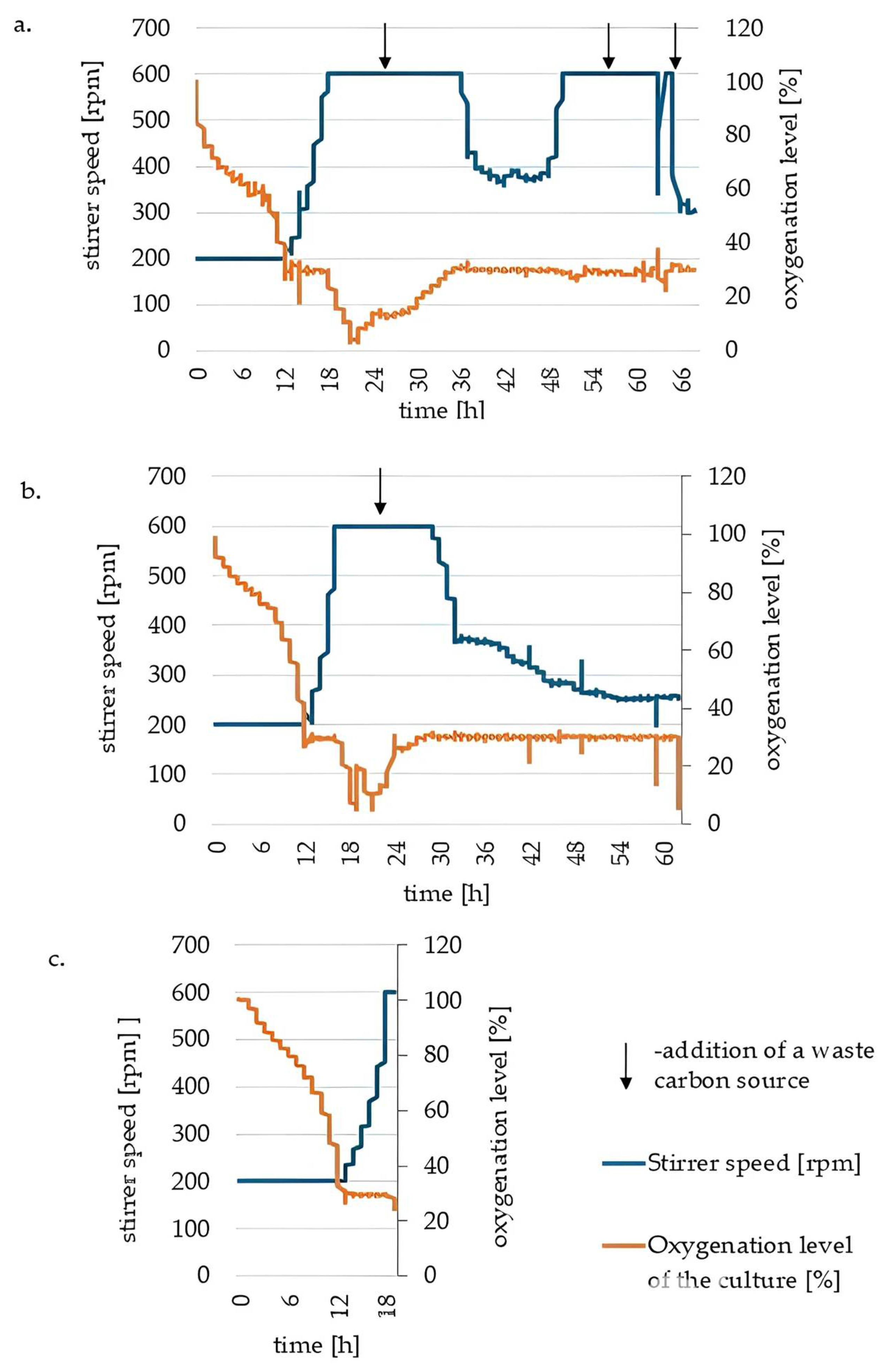
3.2. Biomass Characteristics of the Oleaginous Yeast Y. lipolytica
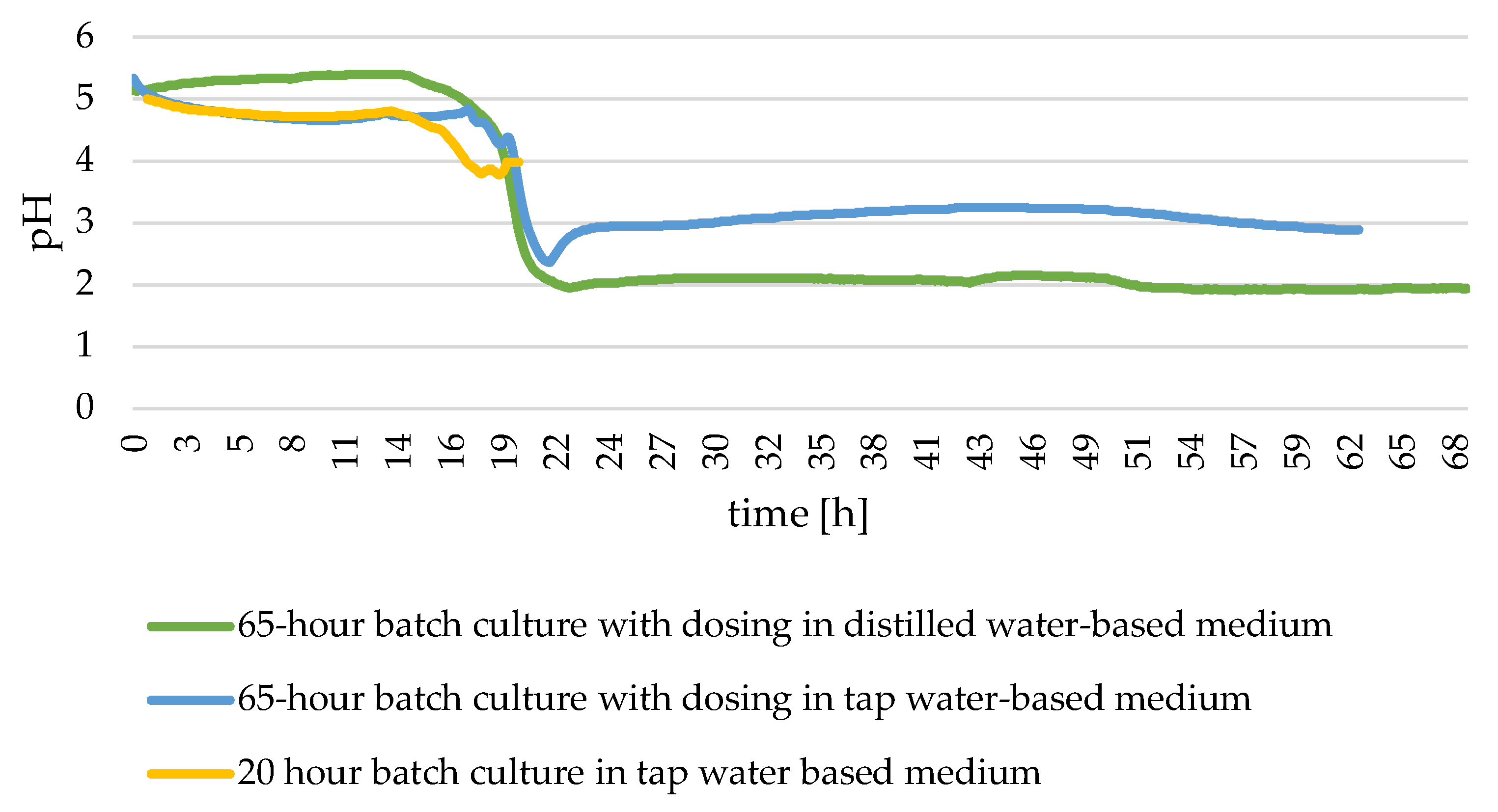
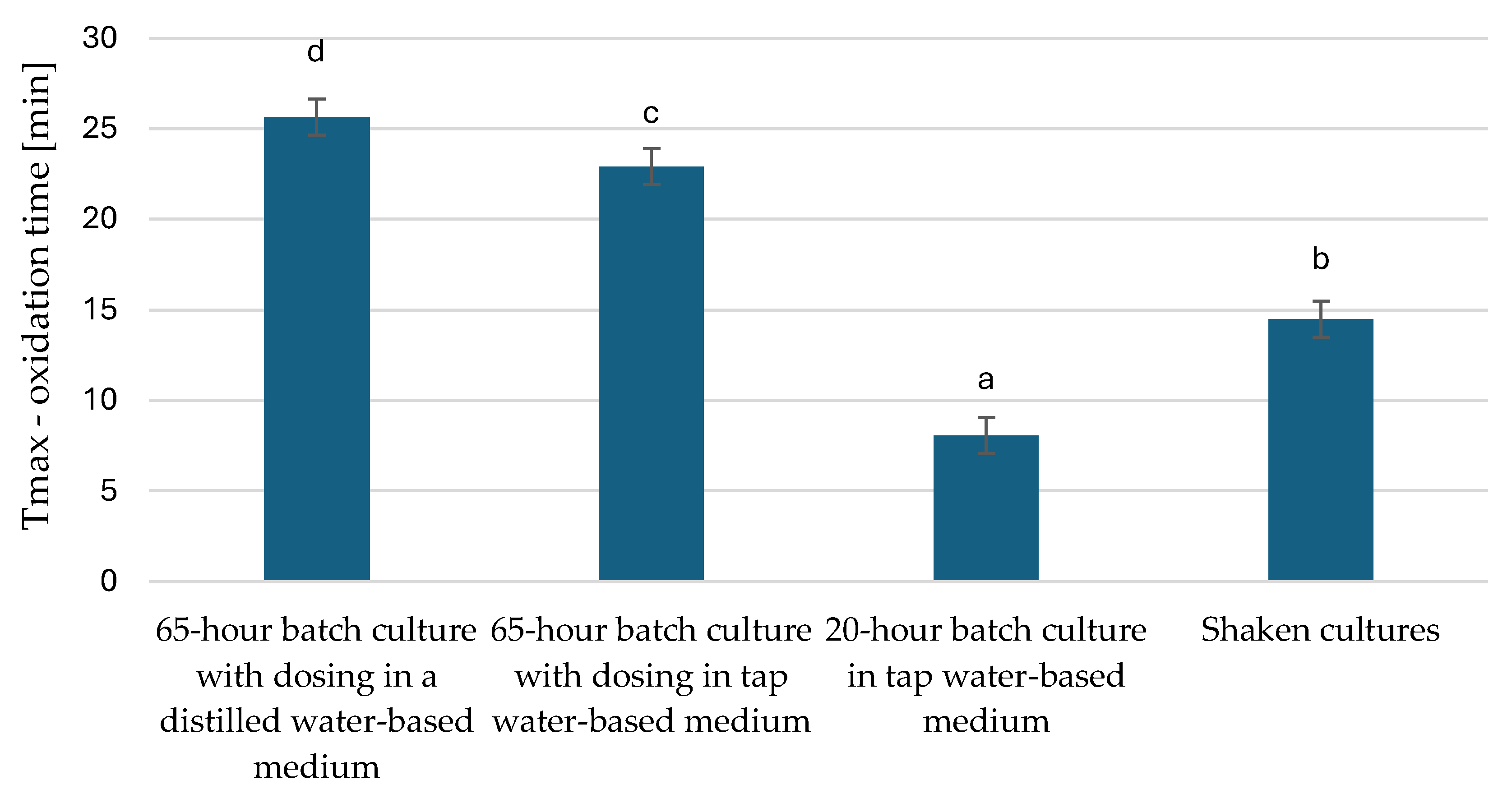
3.3. Characteristics of Microbial Oil Extracted from Y. lipolytica Yeast Cells
3.4. Evaluation of Microbial Oil and Biomass Safety
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety Assessment of an Oleaginous Yeast with a Great Industrial Potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, E.; Charreau, H.; Cerrutti, P.; Foresti, M.L. Yarrowia lipolytica: A Model Yeast for Citric Acid Production. FEMS Yeast Res. 2017, 17, fox084. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, J.F.-W.; Tsang, C.-C.; Wang, H.; Guo, D.; Pan, Y.; Xiao, Y.; Yue, N.; Chen, J.H.-K.; Lau, S.K.-P.; et al. Clinical Characteristics, Laboratory Identification, and in Vitro Antifungal Susceptibility of Yarrowia (Candida) lipolytica Isolates Causing Fungemia: A Multicenter, Prospective Surveillance Study. J. Clin. Microbiol. 2015, 53, 3639–3645. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.-I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Yarrowia lipolytica Yeast Biomass as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, e05594. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and Its Multiple Applications in the Biotechnological Industry. Sci. World J. 2014, 2014, 476207. [Google Scholar] [CrossRef]
- Jach, M.E.; Masłyk, M.; Juda, M.; Sajnaga, E.; Malm, A. Vitamin B12-Enriched Yarrowia lipolytica Biomass Obtained from Biofuel Waste. Waste Biomass Valorization 2020, 11, 1711–1716. [Google Scholar] [CrossRef]
- Jach, M.E.; Serefko, A.; Ziaja, M.; Kieliszek, M. Yeast Protein as an Easily Accessible Food Source. Metabolites 2022, 12, 63. [Google Scholar] [CrossRef]
- Jach, M.E.; Malm, A. Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans. Molecules 2022, 27, 2300. [Google Scholar] [CrossRef]
- Fabiszewska, A.; Misiukiewicz-Stępień, P.; Paplińska-Goryca, M.; Zieniuk, B.; Białecka-Florjańczyk, E. An Insight into Storage Lipid Synthesis by Yarrowia lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism. Biomolecules 2019, 9, 685. [Google Scholar] [CrossRef]
- Implementing Regulation—EU—2024/2044—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg_impl/2024/2044/oj/eng (accessed on 9 June 2025).
- Finco, A.M.d.O.; Mamani, L.D.G.; Carvalho, J.C.d.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Soccol, C.R. Technological Trends and Market Perspectives for Production of Microbial Oils Rich in Omega-3. Crit. Rev. Biotechnol. 2017, 37, 656–671. [Google Scholar] [CrossRef]
- El Kantar, S.; Khelfa, A.; Vorobiev, E.; Koubaa, M. Strategies for Increasing Lipid Accumulation and Recovery from Y. lipolytica: A Review. OCL 2021, 28, 51. [Google Scholar] [CrossRef]
- Abeln, F.; Chuck, C.J. The History, State of the Art and Future Prospects for Oleaginous Yeast Research. Microb. Cell Fact. 2021, 20, 221. [Google Scholar] [CrossRef]
- Al-Obeidi, W.D.M.; Al-Rawi, D.F.; Ali, L.H. Production of Single-Cell Oil from a Local Isolate Bacillus subtilis Using Palm Fronds. Int. J. Biomater. 2023, 2023, 8882842. [Google Scholar] [CrossRef]
- Bellou, S.; Triantaphyllidou, I.-E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial Oils as Food Additives: Recent Approaches for Improving Microbial Oil Production and Its Polyunsaturated Fatty Acid Content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, B.; Teixeira, J.C.; Dragone, G.; Teixeira, J.A. Oleaginous Yeasts for Sustainable Lipid Production-from Biodiesel to Surf Boards, a Wide Range of “Green” Applications. Appl. Microbiol. Biotechnol. 2019, 103, 3651–3667. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, L.A.; Chuck, C.J.; Donnelly, J.; Bannister, C.D.; Scott, R.J. Optimizing the Lipid Profile, to Produce Either a Palm Oil or Biodiesel Substitute, by Manipulation of the Culture Conditions for Rhodotorula glutinis. Biofuels 2014, 5, 33–43. [Google Scholar] [CrossRef]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of Volatile Fatty Acids into Lipids by the Oleaginous Yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and Techno-Economic Evaluation of Microbial Oil Production as a Renewable Resource for Biodiesel and Oleochemical Production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Wierzchowska, K.; Roszko, M.; Derewiaka, D.; Szulc, K.; Zieniuk, B.; Nowak, D.; Fabiszewska, A. Yeast Lipids as a Sustainable Source of Nutrients in Dairy Products Analogs. Food Biosci. 2024, 62, 105321. [Google Scholar] [CrossRef]
- PN-EN ISO 10523:2012; Water Quality—Determination of pH. Polish Committee for Standardisation: Warsaw, Poland, 2012.
- PN-EN 26777:1999; Water Quality—Determination of Nitrites—Molecular Absorption Spectrometry Method. Polish Committee for Standardisation: Warsaw, Poland, 1999.
- PN-82/C-04576.08; Water and waste water—Tests for Nitrogen Compounds -Determination of Nitrate Nitrogen by the Colorimetric Method with Sodium Salicylate. Polish Committee for Standardisation: Warsaw, Poland, 2012.
- PN-ISO 7150-1:2002; Water Quality—Determination of Ammoniacal Nitrogen—Part 1: Manual Spectrometric Method. Polish Committee for Standardisation: Warsaw, Poland, 2002.
- PN-ISO 9297:1994; Water Quality—Determination of Chlorides—Method of Titration with Silver Nitrate in the Presence of Chromate as Indicator (Mohr Method). Polish Committee for Standardisation: Warsaw, Poland, 1994.
- PN-ISO 6059:1999; Water Quality—Determination of Total Calcium and Magnesium Content—Titration Method with EDTA. Polish Committee for Standardisation: Warsaw, Poland, 1999.
- PN-EN ISO 11885:2009; Water Quality—Determination of Selected Elements by Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES). Polish Committee for Standardisation: Warsaw, Poland, 2009.
- ISO 15923-1:2013; Water quality—Determination of selected parameters by discrete analysis systems—Part 1: Ammonium, nitrate, nitrite, chloride, orthophosphate, sulfate and silicate with photometric detection. International Organization for Standardization: Geneva, Switzerland, 2013.
- Papanikolaou, S.; Aggelis, G. Lipids of Oleaginous Yeasts. Part II: Technology and Potential Applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Taskin, M.; Saghafian, A.; Aydogan, M.N.; Arslan, N.P. Microbial Lipid Production by Cold-adapted Oleaginous Yeast Yarrowia lipolytica B9 in Non-sterile Whey Medium. Biofuels Bioprod. Biorefin. 2015, 9, 595–605. [Google Scholar] [CrossRef]
- Derewiaka, D.; Stepnowska, N.; Bryś, J.; Ziarno, M.; Ciecierska, M.; Kowalska, J. Chia Seed Oil as an Additive to Yogurt. Grasas Aceites 2019, 70, e302. [Google Scholar] [CrossRef]
- WisłaWarszawska.Pl. Available online: http://wislawarszawska.pl (accessed on 19 February 2025).
- Miejskie Przedsiębiorstwo Wodociągów i Kanalizacji. Available online: http://www.mpwik.com.pl (accessed on 19 February 2025).
- Official Portal of the City of Warsaw. Available online: http://um.warszawa.pl (accessed on 19 February 2025).
- Zdrowia, R.M. Warszawa, Dnia 11 Grudnia 2017 r. Poz. 2294. Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20170002294/O/D20172294.pdf (accessed on 9 June 2025). (In Polish)
- Dobrowolski, A.; Drzymała, K.; Rzechonek, D.A.; Mituła, P.; Mirończuk, A.M. Lipid Production from Waste Materials in Seawater-Based Medium by the Yeast Yarrowia lipolytica. Front. Microbiol. 2019, 10, 547. [Google Scholar] [CrossRef]
- Snopek, P.; Nowak, D.; Zieniuk, B.; Fabiszewska, A. Aeration and Stirring in Yarrowia lipolytica Lipase Biosynthesis during Batch Cultures with Waste Fish Oil as a Carbon Source. Fermentation 2021, 7, 88. [Google Scholar] [CrossRef]
- Fabiszewska, A.; Paplińska-Goryca, M.; Misiukiewicz-Stępień, P.; Wołoszynowska, M.; Nowak, D.; Zieniuk, B. Expression Profile of Selected Genes Involved in Storage Lipid Synthesis in a Model Oleaginous Yeast Species Yarrowia lipolytica. Int. J. Mol. Sci. 2022, 23, 1041. [Google Scholar] [CrossRef] [PubMed]
- Magdouli, S.; Brar, S.K.; Blais, J.F. Morphology and Rheological Behaviour of Yarrowia lipolytica: Impact of Dissolved Oxygen Level on Cell Growth and Lipid Composition. Process Biochem. 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zieniuk, B.; Kozłowska, M.; Mazurczak-Zieniuk, P.M.; Wołoszynowska, M.; Misiukiewicz-Stępień, P.; Nowak, D. Studies on Upgradation of Waste Fish Oil to Lipid-Rich Yeast Biomass in Yarrowia lipolytica Batch Cultures. Foods 2021, 10, 436. [Google Scholar] [CrossRef]
- Giacomobono, R.; Albergo, R.; Valerio, V.; Caporusso, A.; De Bari, I. Modelling of the Citric Acid Production from Crude Glycerol by Wild-Type Yarrowia lipolytica DSM 8218 Using Response Surface Methodology (RSM). Life 2022, 12, 621. [Google Scholar] [CrossRef]
- Pereira, A.S.; Lopes, M.; Miranda, S.M.; Belo, I. Bio-Oil Production for Biodiesel Industry by Yarrowia lipolytica from Volatile Fatty Acids in Two-Stage Batch Culture. Appl. Microbiol. Biotechnol. 2022, 106, 2869–2881. [Google Scholar] [CrossRef]
- Louhasakul, Y.; Cheirsilp, B.; Maneerat, S.; Prasertsan, P. Potential Use of Flocculating Oleaginous Yeasts for Bioconversion of Industrial Wastes into Biodiesel Feedstocks. Renew. Energy 2019, 136, 1311–1319. [Google Scholar] [CrossRef]
- Juszczyk, P.; Rymowicz, W.; Kita, A.; Rywińska, A. Biomass Production by Yarrowia lipolytica Yeast Using Waste Derived from the Production of Ethyl Esters of Polyunsaturated Fatty Acids of Flaxseed Oil. Ind. Crops Prod. 2019, 138, 111590. [Google Scholar] [CrossRef]
- Karamerou, E.E.; Parsons, S.; McManus, M.C.; Chuck, C.J. Using Techno-Economic Modelling to Determine the Minimum Cost Possible for a Microbial Palm Oil Substitute. Biotechnol. Biofuels 2021, 14, 57. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; Zhang, B.; Chen, H.; He, Y.; Bai, Z.; He, Q. Techno-economic evaluation of large-scale SCP production by Pichia pastoris using methanol as sole carbon source. Bioresour. Technol. 2023, 368, 128331. [Google Scholar] [CrossRef]
- Khot, M.; Raut, G.; Ghosh, D.; Alarcón-Vivero, M.; Contreras, D.; Ravikumar, A. Lipid Recovery from Oleaginous Yeasts: Perspectives and Challenges for Industrial Applications. Fuel 2020, 259, 116292. [Google Scholar] [CrossRef]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid Production in Yarrowia lipolytica Is Maximized by Engineering Cytosolic Redox Metabolism. Nat. Biotechnol. 2017, 35, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Qiao, K.; Ahn, W.S.; Stephanopoulos, G. Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. USA 2016, 113, 10848–10853. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Nielsen, J. Production of advanced biofuels in yeast. Biotechnol. J. 2021, 16, 2000304. [Google Scholar]
- Uğur, Ş.; Zieniuk, B.; Fabiszewska, A. Nutritional and Medicinal Properties of Microbial Oil. Appl. Sci. 2024, 14, 4232. [Google Scholar] [CrossRef]
- Mazurczak, P.; Białecka-Florjańczyk, E.; Fabiszewska, A.; Nowak, D.; Wołoszynowska, M.; Zieniuk, B. Utylizacja odpadów pochodzących z zakładów przemysłu spożywczego i paliwowego z wykorzystaniem lipolitycznych drożdży Yarrowia lipolytica. Zesz. Probl. Postępów Nauk. Rol. 2017, nr 588, 15–24. [Google Scholar] [CrossRef]
- Wierzchowska, K.; Pakulska, A.; Derewiaka, D.; Piasecka, I.; Zieniuk, B.; Nowak, D.; Fabiszewska, A. Concept of Batch and Fed-Batch Cultures of Yarrowia lipolytica as a Valuable Source of Sterols with Simultaneous Valorization of Molasses and Post-Frying Rapeseed Oil. Appl. Sci. 2022, 12, 12877. [Google Scholar] [CrossRef]
- Bryś, A.; Bryś, J.; Mellado, Á.F.; Głowacki, S.; Tulej, W.; Ostrowska-Ligęza, E.; Koczoń, P. Characterization of Oil from Roasted Hemp Seeds Using the PDSC and FTIR Techniques. J. Therm. Anal. Calorim. 2019, 138, 2781–2786. [Google Scholar] [CrossRef]
- Ciemniewska-Żytkiewicz, H.; Ratusz, K.; Bryś, J.; Reder, M.; Koczoń, P. Determination of the Oxidative Stability of Hazelnut Oils by PDSC and Rancimat Methods. J. Therm. Anal. Calorim. 2014, 118, 875–881. [Google Scholar] [CrossRef]
- Islam, M.; Kaczmarek, A.; Tomaszewska-Gras, J. Differential Scanning Calorimetry as a Tool to Assess the Oxidation State of Cold-Pressed Oils during Shelf-Life. J. Food Meas. Charact. 2023, 17, 6639–6651. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2023/915 of 25 April 2023 on Maximum Levels for Certain Contaminants in Food and Repealing Regulation (EC) No 1881/2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32023R0915 (accessed on 19 May 2025).
- Directive 2009/32/EC of the European Parliament and of the Council of 23 April 2009 on the Approximation of the Laws of the Member States on Extraction Solvents Used in the Production of Foodstuffs and Food Ingredients. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ%3AL%3A2009%3A141%3A0003%3A0011%3AEN%3APDF (accessed on 19 May 2025).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Parameter | Unit | Water C | Water P | Water B | Water U | Water Parameters and Acceptable Parametric Values [35] |
|---|---|---|---|---|---|---|
| pH (at 20 °C) | - | 7.0 ± 0.2 | 8.1 ± 0.2 | 7.0 ± 0.2 | 7.7 ± 0.2 | 6.5–9.5 |
| Electrical conductivity (ɣ25) | µS/cm | 3.0 ± 1.0 | 1021.0 ± 31.0 | 831.0 ± 25.0 | 857.0 ± 26.0 | 2500.0 |
| Nitrites (NO2) | mg/L | <0.033 (0.033 ± 0.005) | <0.033 (0.033 ± 0.005) | <0.033 (0.033 ± 0.005) | <0.033 (0.033 ± 0.005) | 0.50 |
| Nitrates (NO3) | mg/L | <0.1 | 5.41 ± 0.76 | 5.31 ± 0.75 | 0.71 ± 0.10 | 50.0 |
| Ammonium ion (NH4) | mg/L | <0.060 (0.060 ± 0.013) | <0.060 (0.060 ± 0.013) | <0.060 (0.060 ± 0.013) | <0.060 (0.060 ± 0.013) | 0.50 |
| Hardness | mg/L | <1.0 | 252 ± 38 | 230 ± 35 | 400 ± 60 | 60–500 |
| Chlorides | mg/L | <1.0 | 186 ± 11 | 132 ± 8 | 97 ± 6 | 250.0 |
| Iron | µg/L | <5.0 (5.0 ± 0.5) | <5.0 (5.0 ± 0.5) | <5.0 (5.0 ± 0.5) | 18 ± 2 | 200.0 |
| Manganese | µg/L | <0.1 | 1.4 ± 0.1 | 2.6 ± 0.3 | 10 ± 1 | 50.0 |
| Oxidisability in KMnO4 (permanganate index) | mg/L | <0.50 (0.50 ± 0.08) | <0.50 (0.50 ± 0.08) | <0.50 (0.50 ± 0.08) | <0.50 (0.50 ± 0.08) | 5.0 |
| Sulfates | mg/L | <5.0 | 65.2 ± 9.8 | 72.7 ± 11.0 | 162.0 ± 25.0 | 250.0 |
| Water Used for the Culture Medium | Time | Yeast Biomass Yield [g/dm3] | Lipid Content [%] | Residual Oil in Medium [%] |
|---|---|---|---|---|
| Tap water (U) | 43 | 8.17 (±0.73) | 27 | 0.44 |
| 65 | 22.33 (±4.56) | 33 | 1.44 | |
| Distilled water | 43 | 9.21 (±0.76) | 26 | 4.44 |
| 65 | 24.89 (±0.94) | 30 | 2.89 | |
| Tap water (U) | 20 | 16.44 (±1.57) | 39 | 36.22 |
| Water Used for the Culture Medium | Time | Protein Content [% DM] | Lipid Content [% DM] | Content of Other Cell Constituents * [% DM] |
|---|---|---|---|---|
| Distilled water | 65 | 24.16(±1.50) a | 39.43 (±2.22) a | 36.41 |
| Tap water (U) | 65 | 27.03(±1.75) a | 48.95 (±2.90) b | 24.02 |
| Tap water (U) | 20 | 25.49 (±1.67) a | 50.53 (±2.16) b | 23.98 |
| Parameter | Unit | Water Used for the Culture Medium | ||||
|---|---|---|---|---|---|---|
| Distilled Water | Tap Water (U)—65 h | Tap Water (U)—20 h | ||||
| Initial concentration of carbon source [S] | g/dm3 | 75 | 125 | 75 | 50 | |
| Time [t] | h | 43 | 65 | 43 | 65 | 20 |
| Duration of lag phase [tlag] | h | 11.1 | 11.4 | 11.7 | ||
| Biomass yield [X] | gd.m./dm3 | 9.21 (±0.76) | 24.89 (±0.94) | 8.17 (±0.73) | 22.33 (±4.56) | 16.44 (±1.57) |
| Maximum concentration of lipids produced [Lmax] | g/dm3 | 2.39 | 7.47 | 2.20 | 7.37 | 6.41 |
| Conversion yield of biomass per carbon substrate [YX/S] | gd.m./g | 0.12 | 0.2 | 0.11 | 0.3 | 0.33 |
| Conversion yield of storage lipids per biomass formed [YL/X] | g/gd.m. | 0.26 | 0.30 | 0.27 | 0.33 | 0.39 |
| Conversion yield of storage lipids per carbon substrate [YL/S] | g/g | 0.03 | 0.06 | 0.03 | 0.1 | 0.13 |
| Volumetric rate of storage lipids production [qLv] | g/dm3/h | 0.06 | 0.11 | 0.05 | 0.11 | 0.32 |
| Specific rate of storage lipid production [qL] | g/gd.m./h | 0.006 | 0.005 | 0.006 | 0.005 | 0.02 |
| Maximum concentration of lipids produced [Lmax]—in the dry biomass | g/dm3 | - | 9.81 | - | 10.93 | 8.31 |
| Conversion yield of storage lipids per biomass formed [YL/X] | g/gd.m. | - | 0.39 | - | 0.50 | 0.51 |
| Conversion yield of storage lipids per carbon substrate [YL/S] | g/g | - | 0.08 | - | 0.15 | 0.17 |
| Volumetric rate of storage lipids production [qLv] | g/dm3/h | - | 0.15 | - | 0.17 | 0.41 |
| Specific rate of storage lipid production [qL] | g/gd.m./h | - | 0.006 | - | 0.008 | 0.03 |
| Maximum concentration of protein content [Lmax] | g/dm3 | - | 6.01 | - | 6.04 | 4.19 |
| Conversion yield of protein content per biomass formed [YL/X] | g/gd.m. | - | 0.24 | - | 0.27 | 0.25 |
| Conversion yield of protein content per carbon substrate [YL/S] | g/g | - | 0.05 | - | 0.08 | 0.08 |
| Volumetric rate of protein content [qLv] | g/dm3/h | - | 0.09 | - | 0.09 | 0.21 |
| Specific rate of storage protein content [qL] | g/gd.m./h | - | 0.004 | - | 0.004 | 0.01 |
| Maximum concentration of content of other cell constituents [Lmax] | g/dm3 | - | 9.06 | - | 5.36 | 3.94 |
| Conversion yield of content of other cell constituents per biomass formed [YL/X] | g/gd.m. | - | 0.36 | - | 0.24 | 0.24 |
| Conversion yield of content of other cell constituents per carbon substrate [YL/S] | g/g | - | 0.07 | - | 0.07 | 0.08 |
| Volumetric rate of content of other cell constituents [qLv] | g/dm3/h | - | 0.14 | - | 0.08 | 0.20 |
| Specific rate of storage content of other cell constituents [qL] | g/gd.m./h | - | 0.006 | - | 0.004 | 0.01 |
| Fatty Acid | Symbol | Waste Oil (Carbon Source) | 65 h Fed-Batch Culture in Distilled Water-Based Medium | 65 h Fed-Batch Culture in Tap Water-Based Medium | 20 h Fed-Batch Culture in Tap Water-Based Medium | Shaken Cultures in Rich YPO Medium |
|---|---|---|---|---|---|---|
| Myristic acid | 14:0 | 0.10 | 0.00 | 0.07 (±0.02) | 0.11 (±0.01) | 0.00 |
| Palmitic acid | 16:0 | 11.50 | 5.07 (±0.06) | 4.46 (±0.14) | 3.71 (±0.14) | 5.39 (±0.01) |
| Palmitoleic acid | 16:1 | 0.66 | 0.07 (±0.03) | 0.00 | 0.22 (±0.03) | 0.50 (±0.03) |
| Stearic acid | 18:0 | 4.32 | 2.00 (±0.03) | 2.05 (±0.02) | 2.55(0.14) | 1.25 (±0.05) |
| Oleic acid | 18:1 | 59.79 | 66.44 (±0.06) | 65.36 (±0.86) | 66.55 (±0.49) | 62.73 (±1.32) |
| Linoleic acid | 18:2 | 16.58 | 19.40 (±0.28) | 19.52 (±0.69) | 18.88 (±0.28) | 21.40 (±0.66) |
| Linolenic acid | 18:3 | 5.22 | 5.56 (±0.20) | 6.77 (±0.46) | 6.14 (±0.07) | 8.72 (±1.95) |
| Arachidic acid | 20:0 | 0.89 | 0.29 (±0.01) | 0.50 (±0.03) | 0.41 (±0.07) | 0.00 |
| Isocenic acid | 20:1 | 0.94 | 1.02 (±0.05) | 1.14 (±0.08) | 0.87 (±0.02) | 0.00 |
| Erucic acid | 22:1 | 0.00 | 0.14 (±0.05) | 0.12 (±0.03) | 0.58 (±0.05) | 0.00 |
| Biological Material | Yeast Biomass * | Microbial Oil | European Regulations [4,57,58] | Reference |
|---|---|---|---|---|
| Sum of PAHs: benzo(a) pyrene, benz(a) anthracene, benzo(b) fluoranthene and chrysene [μg/kg] | n.a. | 4.12 | 10.0 | [20] |
| Benzo(a) pyrene [μg/kg] | n.a. | 0.61 | 2.0 | [20] |
| Hexane residues (mg/kg) | n.a. | 0.0007 | 1.0 | [20] |
| Heavy metals | 0 | 0 | 0.1 | [20] |
| Total bacterial counts [CFU/g] | 5.8 × 101 | 1.4 × 101 | ≤5 × 103 | |
| Number of yeast counts[CFU/g] | 0 | 0 | ≤1 × 102 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabiszewska, A.U.; Kobus, J.; Górnicka, M.; Piotrowicz, A.; Piasecka, I.; Nowak, D. Valorisation of Waste Oils Through Oleaginous Yarrowia lipolytica Yeast: Insights into Lipid Stability and Nutritive Properties of Lipid-Rich Biomass. Appl. Sci. 2025, 15, 6796. https://doi.org/10.3390/app15126796
Fabiszewska AU, Kobus J, Górnicka M, Piotrowicz A, Piasecka I, Nowak D. Valorisation of Waste Oils Through Oleaginous Yarrowia lipolytica Yeast: Insights into Lipid Stability and Nutritive Properties of Lipid-Rich Biomass. Applied Sciences. 2025; 15(12):6796. https://doi.org/10.3390/app15126796
Chicago/Turabian StyleFabiszewska, Agata Urszula, Joanna Kobus, Magdalena Górnicka, Aleksandra Piotrowicz, Iga Piasecka, and Dorota Nowak. 2025. "Valorisation of Waste Oils Through Oleaginous Yarrowia lipolytica Yeast: Insights into Lipid Stability and Nutritive Properties of Lipid-Rich Biomass" Applied Sciences 15, no. 12: 6796. https://doi.org/10.3390/app15126796
APA StyleFabiszewska, A. U., Kobus, J., Górnicka, M., Piotrowicz, A., Piasecka, I., & Nowak, D. (2025). Valorisation of Waste Oils Through Oleaginous Yarrowia lipolytica Yeast: Insights into Lipid Stability and Nutritive Properties of Lipid-Rich Biomass. Applied Sciences, 15(12), 6796. https://doi.org/10.3390/app15126796