
Evaluating Turkey-Derived Lactic-Acid-Producing Bacteria as Potential Probiotics for Use in Commercial Turkeys

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Acid Tolerance
2.3. Bile Salt Tolerance
2.4. Bile Salt Hydrolase Activity
2.5. Avian Cell Association Assay
2.6. Statistical Analysis
3. Results
3.1. Isolation and Species Identification of Turkey-Sourced Lactic Acid Bacteria
3.2. Turkey-Sourced LAB Vary in Their Ability to Grow in Acidic Conditions
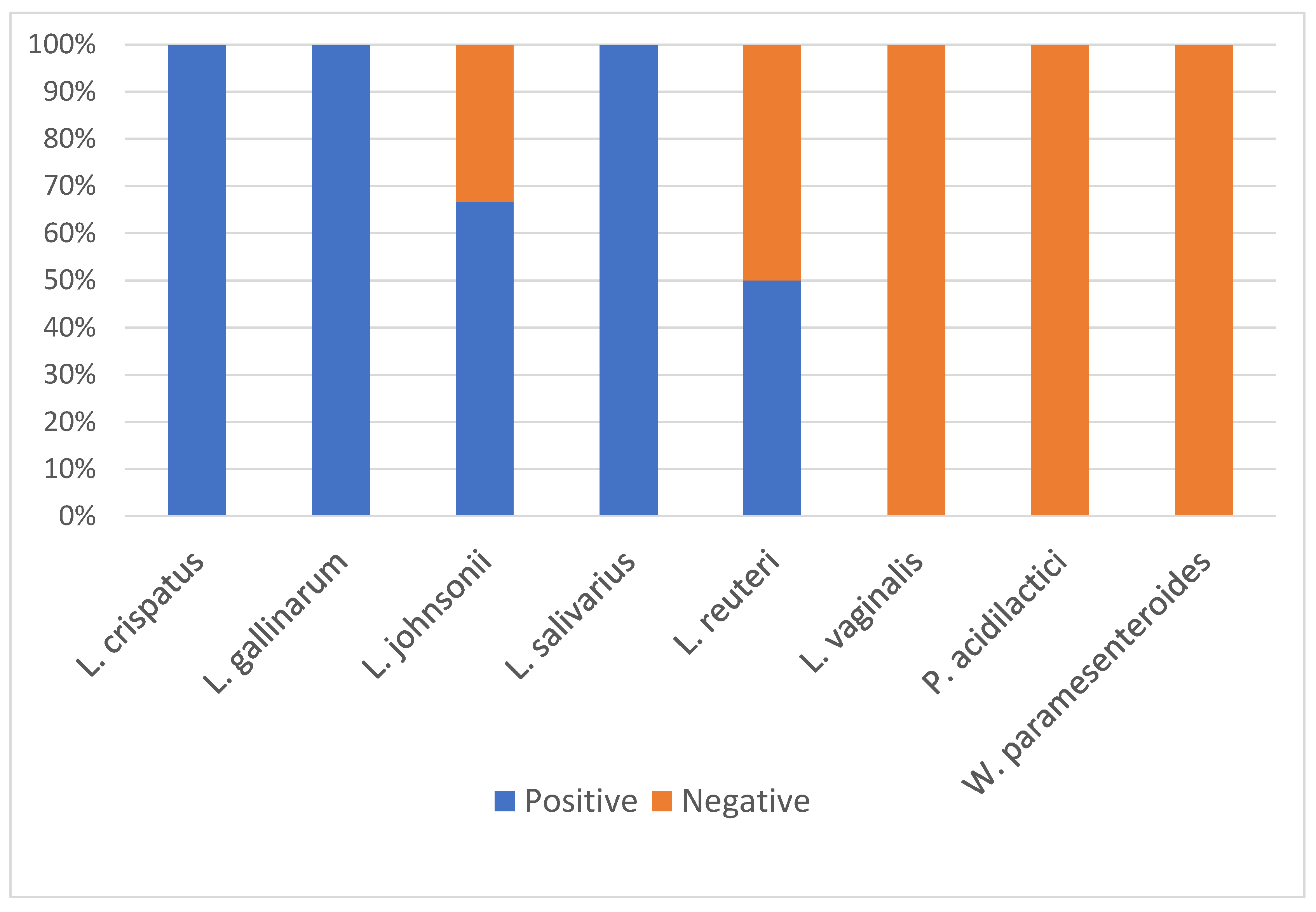
3.3. Turkey-Sourced LAB Vary in Their Response to Bile
3.4. Turkey-Sourced LAB Vary Individually, but Not by Species in Their Ability to Adhere to Avian Intestinal Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, R.S.; Porter, L.J.; Thomson, D.U.; Gage, M.; Beaudoin, A.; Wishnie, J.K. Raising Animals Without Antibiotics: U.S. Producer and Veterinarian Experiences and Opinions. Front. Vet. Sci. 2019, 6, 452. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Taha, A.E.; Soliman, S.M.; Ahmed, A.E.; El-Kott, A.F.; et al. Alternatives to antibiotics for organic poultry production: Types, modes of action and impacts on bird’s health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef]
- Johnson, A.; Miller, E.A.; Weber, B.; Figueroa, C.F.; Aguayo, J.M.; Johny, A.K.; Noll, S.; Brannon, J.; Kozlowicz, B.; Johnson, T.J. Evidence of host specificity in Lactobacillus johnsonii genomes and its influence on probiotic potential in poultry. Poult. Sci. 2023, 102, 102858. [Google Scholar] [CrossRef]
- Manes-Lazaro, R.; Van Diemen, P.M.; Pin, C.; Mayer, M.J.; Stevens, M.P.; Narbad, A. Administration of Lactobacillus johnsonii FI9785 to chickens affects colonisation by Campylobacter jejuni and the intestinal microbiota. Br. Poult. Sci. 2017, 58, 373–381. [Google Scholar] [CrossRef]
- Gao, P.; Ma, C.; Sun, Z.; Wang, L.; Huang, S.; Su, X.; Xu, J.; Zhang, H. Feed-additive probiotics accelerate yet antibiotics delay intestinal microbiota maturation in broiler chicken. Microbiome 2017, 5, 91. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Kollanoor Johny, A. Characterizing the Antimicrobial Function of a Dairy-Originated Probiotic, Propionibacterium freudenreichii, Against Multidrug-Resistant Salmonella enterica Serovar Heidelberg in Turkey Poults. Front. Microbiol. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Hashemi, S.M.B. Lactic acid production-producing microorganisms and substrates sources-state of art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.L.; Weber, B.P.; Mendoza, K.M.; Danzeisen, J.L.; Llop, K.; Lang, K.; Clayton, J.B.; Grace, E.; Brannon, J.; Radovic, I.; et al. Antibiotics and Host-Tailored Probiotics Similarly Modulate Effects on the Developing Avian Microbiome, Mycobiome, and Host Gene Expression. mBio 2019, 10, e02171-19. [Google Scholar] [CrossRef] [PubMed]
- Casas, I.; Edens, F.; Parkhurst, C.; Dobrogosz, W. Probiotic Administrations of Lactobacillus Reuteri Moderate Avian Growth Depression in Turkeys. Biosci. Microflora 1998, 17, 125–131. [Google Scholar] [CrossRef]
- Khabirov, A.; Khaziakhmetov, F.; Rebezov, Y.; Gorelik, O.; Derkho, M.; Fedoseeva, N.; Lykasova, N.; Kuznetsov, A.; Samorodvoa, I. Effect of Feeding Diet Containing Probiotics on Growth Rate and Hematological Changes in the Blood of Turkeys. Int. J. Pharm. Res. 2020, 12, 1454. [Google Scholar]
- Partners, T.I. Poultry Probiotic Market to Reach $188 Million by 2030-Exclusive Report by the Insight Partners. Available online: https://www.globenewswire.com/news-release/2023/11/08/2776279/0/en/Poultry-Probiotic-Market-to-Reach-188-Million-by-2030-Exclusive-Report-by-The-Insight-Partners.html (accessed on 10 January 2024).
- Rodriguez, R.L.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The Microbial Genomes Atlas (MiGA) webserver: Taxonomic and gene diversity analysis of Archaea and Bacteria at the whole genome level. Nucleic Acids Res. 2018, 46 (W1), W282–W288. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Kwarteng, J.; Tano-Debrah, K.; Akabanda, F.; Jespersen, L. Technological properties and probiotic potential of Lactobacillus fermentum strains isolated from West African fermented millet dough. BMC Microbiol. 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.V.; Nair, D.V.T.; Noll, S.; Johnson, T.J.; Cardona, C.; Johny, A.K. Effect of Turkey-Derived Beneficial Bacteria Lactobacillus salivarius and Lactobacillus ingluviei on a Multidrug-Resistant Salmonella Heidelberg Strain in Turkey Poults. J. Food Prot. 2019, 82, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Sprouffske, K.; Wagner, A. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef]
- Johnson, T.J.; Youmans, B.P.; Noll, S.; Cardona, C.; Evans, N.P.; Karnezos, T.P.; Ngunjiri, J.M.; Abundo, M.C.; Lee, C.W. A Consistent and Predictable Commercial Broiler Chicken Bacterial Microbiota in Antibiotic-Free Production Displays Strong Correlations with Performance. Appl. Environ. Microbiol. 2018, 84, e00362-18. [Google Scholar] [CrossRef]
- Danzeisen, J.L.; Calvert, A.J.; Noll, S.L.; McComb, B.; Sherwood, J.S.; Logue, C.M.; Johnson, T.J. Succession of the turkey gastrointestinal bacterial microbiome related to weight gain. PeerJ 2013, 1, e237. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, K.; Zhang, Y.; Li, Y.; Zhou, N.; Li, G. Probiotic characteristics and whole-genome sequence analysis of Pediococcus acidilactici isolated from the feces of adult beagles. Front. Microbiol. 2023, 14, 1179953. [Google Scholar]
- Diaz, M.; Del Rio, B.; Ladero, V.; Redruello, B.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. Histamine production in Lactobacillus vaginalis improves cell survival at low pH by counteracting the acidification of the cytosol. Int. J. Food Microbiol. 2020, 321, 108548. [Google Scholar] [CrossRef]
- Olajugbagbe, T.E.; Elugbadebo, O.E.; Omafuvbe, B.O. Probiotic potentials of Pediococuss acidilactici isolated from wara; A Nigerian unripened soft cheese. Heliyon 2020, 6, e04889. [Google Scholar] [CrossRef]
- Sulistiani; Novarina, I.; Inawati; Dinoto, A.; Julistiono, H.; Handayani, R.; Saputra, S. Assessment of Potential Probiotic Lactic Acid Bacteria from Tempe and Tape. IOP Conf. Ser. Earth Environ. Sci. 2020, 572, 012026. [Google Scholar] [CrossRef]
- Li, L.; Zhang, N.; Kong, L.; Zhao, W.; Xiao, Y.; Li, B.; Han, X. Identification and Selection of Lactic Acid Bacteria Resistance to Acid and Bile Salt Isolated from Corn Silage. J. Pure Appl. Microbiol. 2015, 9, 13–23. [Google Scholar]
- Sayers, E.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Conner, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Pabari, K.; Pithva, S.; Kothari, C.; Purama, R.K.; Kondepudi, K.K.; Vyas, B.R.M.; Kothari, R.; Ambalam, P. Evaluation of Probiotic Properties and Prebiotic Utilization Potential of Weissella paramesenteroides Isolated From Fruits. Probiotics Antimicrob. Proteins 2020, 12, 1126–1138. [Google Scholar] [CrossRef] [PubMed]
- Merati, R.; Mohamed, A.A.A.-F.; Berrama, Z.; Aggad, H.; Hammoudi, A.; Temim, S. The effects of Pediococcus acidilactici and Saccharomyces cerevisiae on broiler chickens challenged with Clostridium perfringens induced subclinical necrotic enteritis. Vet. Arh. 2021, 91, 389–397. [Google Scholar] [CrossRef]
- Rahmani Alizadeh, M.; Aliakbarpour, H.-R.; Hashemi Karouei, S.M. Effect of dietary supplementation of Iranian multi-strain probiotic or P. acidilactici of camel milk isolate on broilers performance, blood parameters, intestinal histology, and microbiota. Ital. J. Anim. Sci. 2023, 22, 660–665. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Closest Species Match | ANI (%) | |
|---|---|---|
| MOF2W3B1T2 | Lactobacillus crispatus | 98.0 |
| MOF2W6B4C10 | Lactobacillus crispatus | 97.8 |
| MOF2W6B4C8 | Lactobacillus crispatus | 97.7 |
| SNF2W3B2M1 | Lactobacillus crispatus | 97.8 |
| SNF2W6B1L3 | Lactobacillus crispatus | 97.9 |
| MOF2W10B1T3 | Lactobacillus gallinarum | 97.7 |
| MOF2W1B3C5 | Lactobacillus gallinarum | 97.7 |
| MOF2W3B5T3 | Lactobacillus gallinarum | 97.9 |
| MOF2W5B1C1 | Lactobacillus gallinarum | 98.1 |
| SNF2W3B1L8 | Lactobacillus gallinarum | 97.8 |
| UMNLJ21 | Lactobacillus johnsonii | 97.5 |
| SNF2W1B5L4 | Lactobacillus johnsonii | 97.7 |
| SNF2W3B3L6 | Lactobacillus johnsonii | 97.8 |
| MOF2W3B1T3 | Ligilactobacillus salivarius | 97.2 |
| MOF2W6B4C3 | Ligilactobacillus salivarius | 97.2 |
| MOF2W3B1C8 | Limosilactobacillus reuteri | 95.8 |
| SNF2W3B2M2 | Limosilactobacillus reuteri | 96.0 |
| SNF2W3B1L7 | Limosilactobacillus vaginalis | 98.8 |
| MOF2W1LC6 | Pediococcus acidilactici | 98.6 |
| MOF2W1LT10 | Pediococcus acidilactici | 98.7 |
| MOF2W5B3T3 | Pediococcus acidilactici | 98.5 |
| MOF2W1LC8 | Weissella paramesenteroides | 98.0 |
| MOF2W2LT2 | Weissella paramesenteroides | 98.2 |
| MOF2W3LC2 | Weissella paramesenteroides | 98.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.; Weber, B.P.; Nair, D.T.; Singer, R.S.; Kollanoor Johny, A.; Johnson, T.J. Evaluating Turkey-Derived Lactic-Acid-Producing Bacteria as Potential Probiotics for Use in Commercial Turkeys. Appl. Sci. 2024, 14, 2010. https://doi.org/10.3390/app14052010
Johnson A, Weber BP, Nair DT, Singer RS, Kollanoor Johny A, Johnson TJ. Evaluating Turkey-Derived Lactic-Acid-Producing Bacteria as Potential Probiotics for Use in Commercial Turkeys. Applied Sciences. 2024; 14(5):2010. https://doi.org/10.3390/app14052010
Chicago/Turabian StyleJohnson, Abigail, Bonnie P. Weber, Divek T. Nair, Randall S. Singer, Anup Kollanoor Johny, and Timothy J. Johnson. 2024. "Evaluating Turkey-Derived Lactic-Acid-Producing Bacteria as Potential Probiotics for Use in Commercial Turkeys" Applied Sciences 14, no. 5: 2010. https://doi.org/10.3390/app14052010
APA StyleJohnson, A., Weber, B. P., Nair, D. T., Singer, R. S., Kollanoor Johny, A., & Johnson, T. J. (2024). Evaluating Turkey-Derived Lactic-Acid-Producing Bacteria as Potential Probiotics for Use in Commercial Turkeys. Applied Sciences, 14(5), 2010. https://doi.org/10.3390/app14052010