Design and Application of Bionic Camouflage Materials Simulating Spectral Reflection Characteristics of Plants: A Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- The main factors affecting the spectral reflectance characteristics of plant leaves include pigment-visible light, structure-near infrared, and moisture-mid infrared, as well as leaf age, fertilizer (nutrient) depletion, and diseases. The PROSPECT model is a classic model that describes the radiation process of plant leaves in the solar spectral band and is widely used in the field of RS detection;
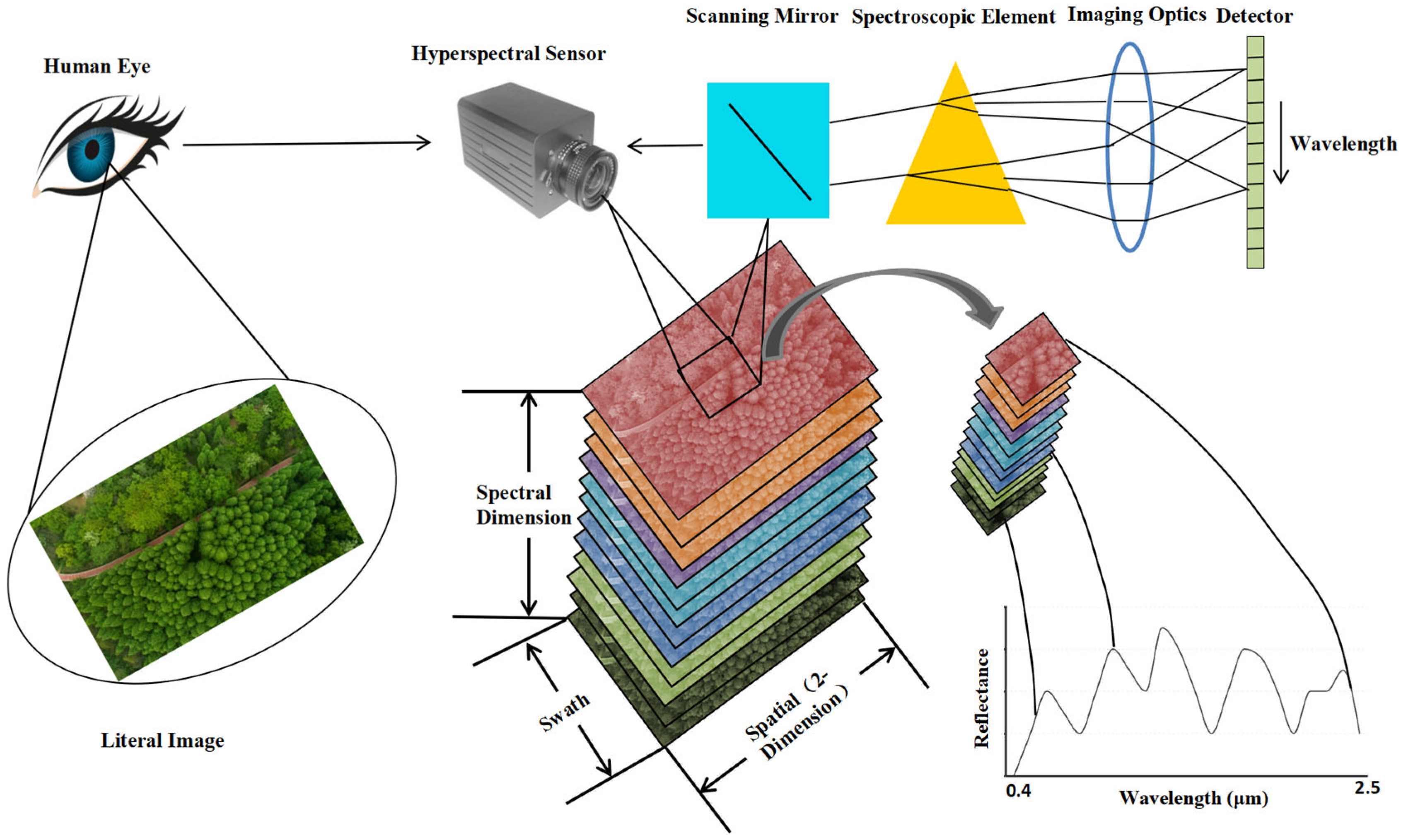
- Hyperspectral RS technology is widely used in fields such as vegetation monitoring [18], plant functional trait mapping [19], biodiversity estimation [20,21], and pest and disease control. Meanwhile, due to the strong ability of hyperspectral RS images to identify camouflage, they can distinguish real targets from decoy targets against the background of green vegetation and quickly detect small tactical targets against a desert background. The current military applications of hyperspectral RS mainly cover three aspects: detailed battlefield reconnaissance, identification of camouflaged targets, and detection and calculation of the target’s true temperature and emissivity;
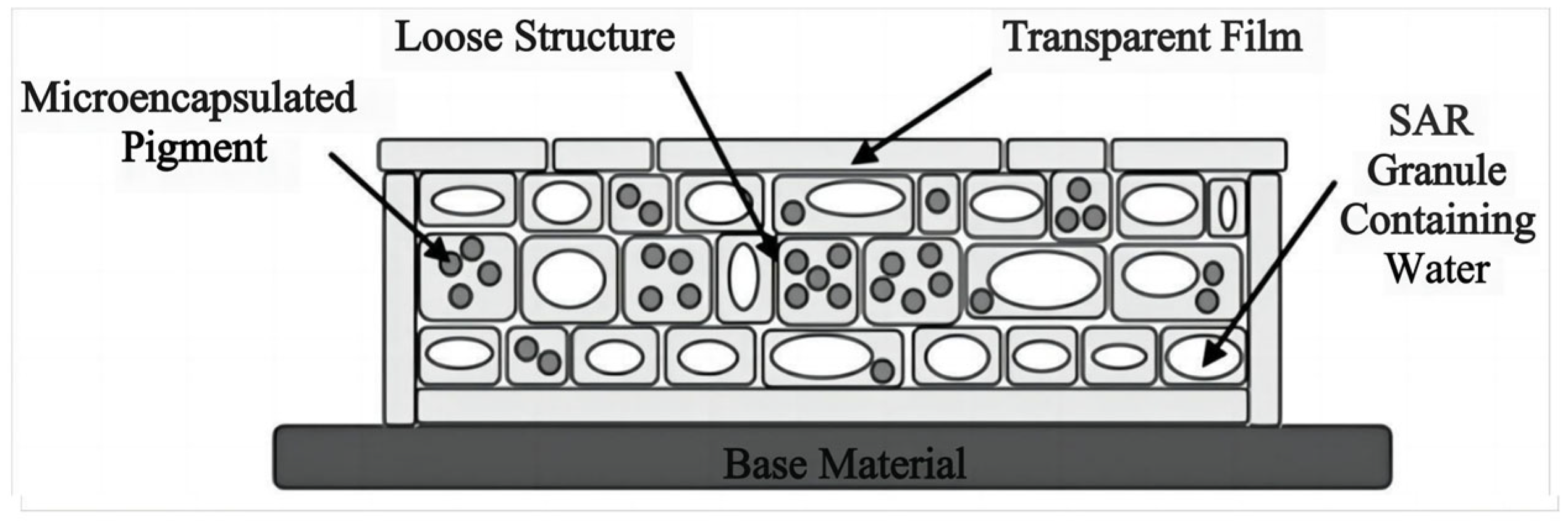
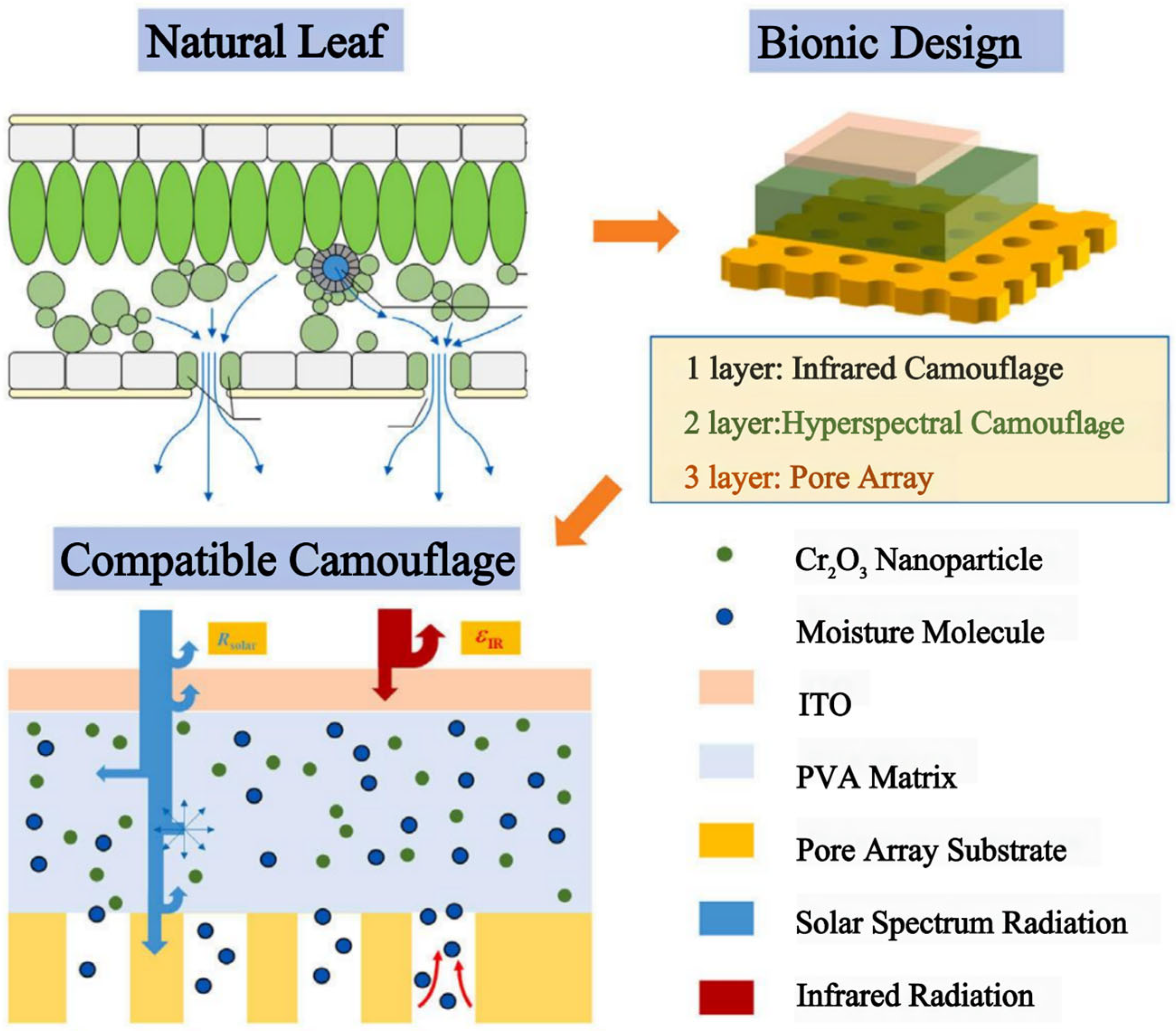
- Plant-mimicking camouflage materials mainly include bionic camouflage materials that can only imitate the color of vegetation and cannot resist hyperspectral RS detection, bionic camouflage materials based on inorganic pigments that can achieve “same color” but do not have the ability to completely achieve “same spectrum” with green vegetation; bionic materials with hyperspectral similarity containing chlorophyl; bionic materials with hyperspectral similarity of leaves at different growth stages, seasons, and species; and bionic materials with solar spectrum reflection characteristics and low infrared emissivity similar to natural leaves.
2. Main Factors Affecting Spectral Reflection Characteristics of Leaves
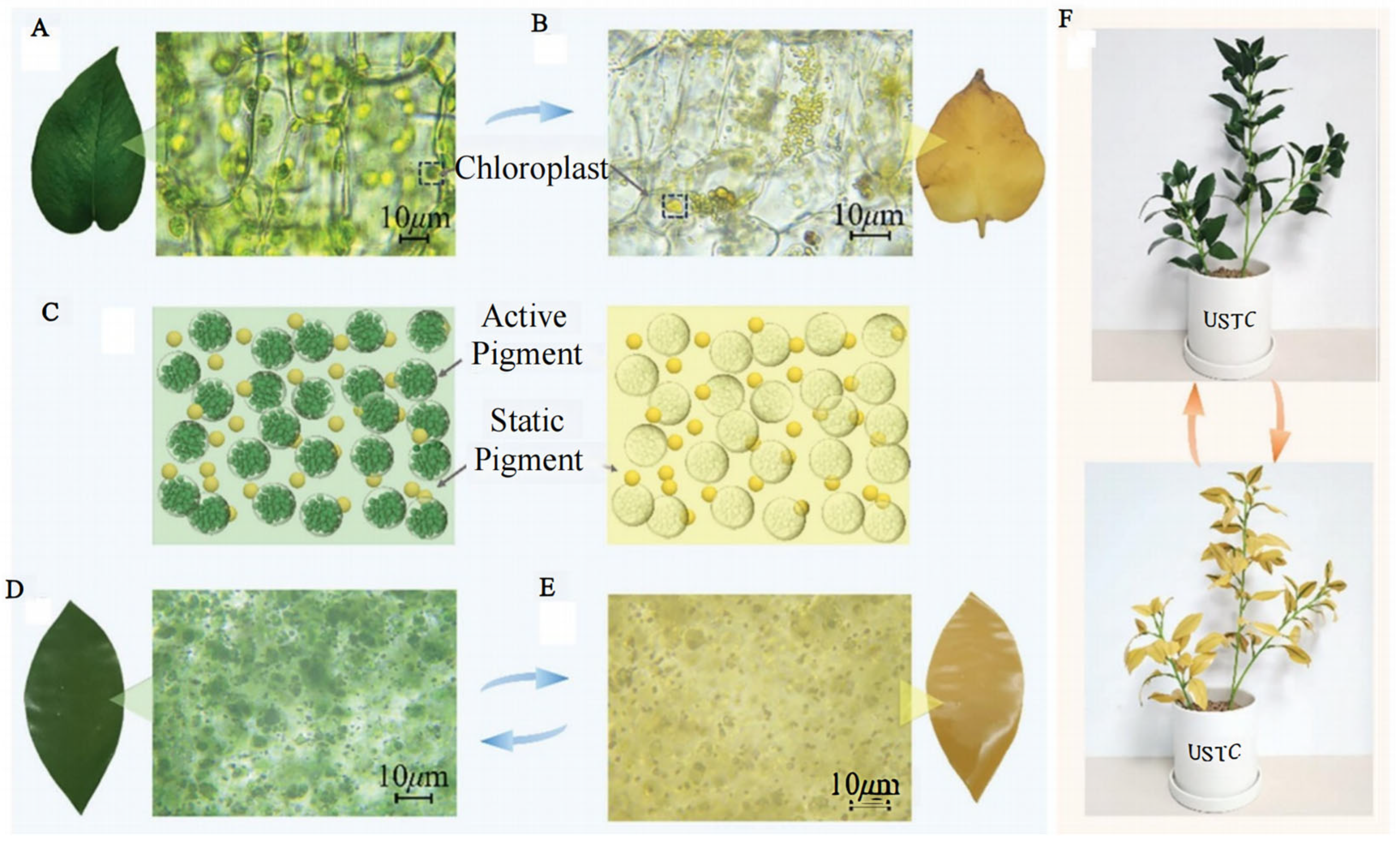
2.1. Pigments—Visible Light
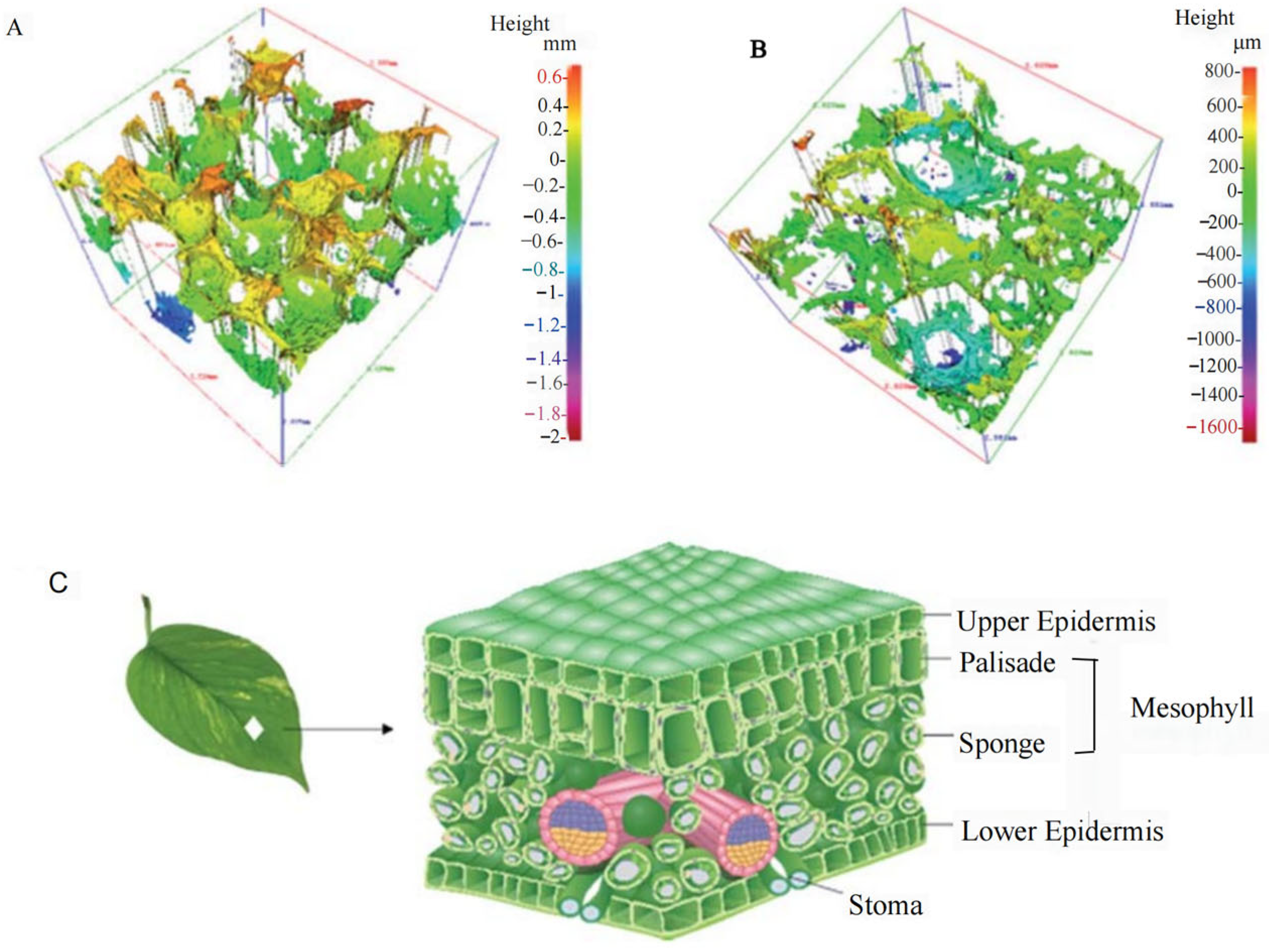
2.2. Structure—Near-Infrared
2.3. Moisture—Mid-Infrared
2.4. Other Influencing Factors
3. Application of Hyperspectral RS into Plant RS and Military Actions
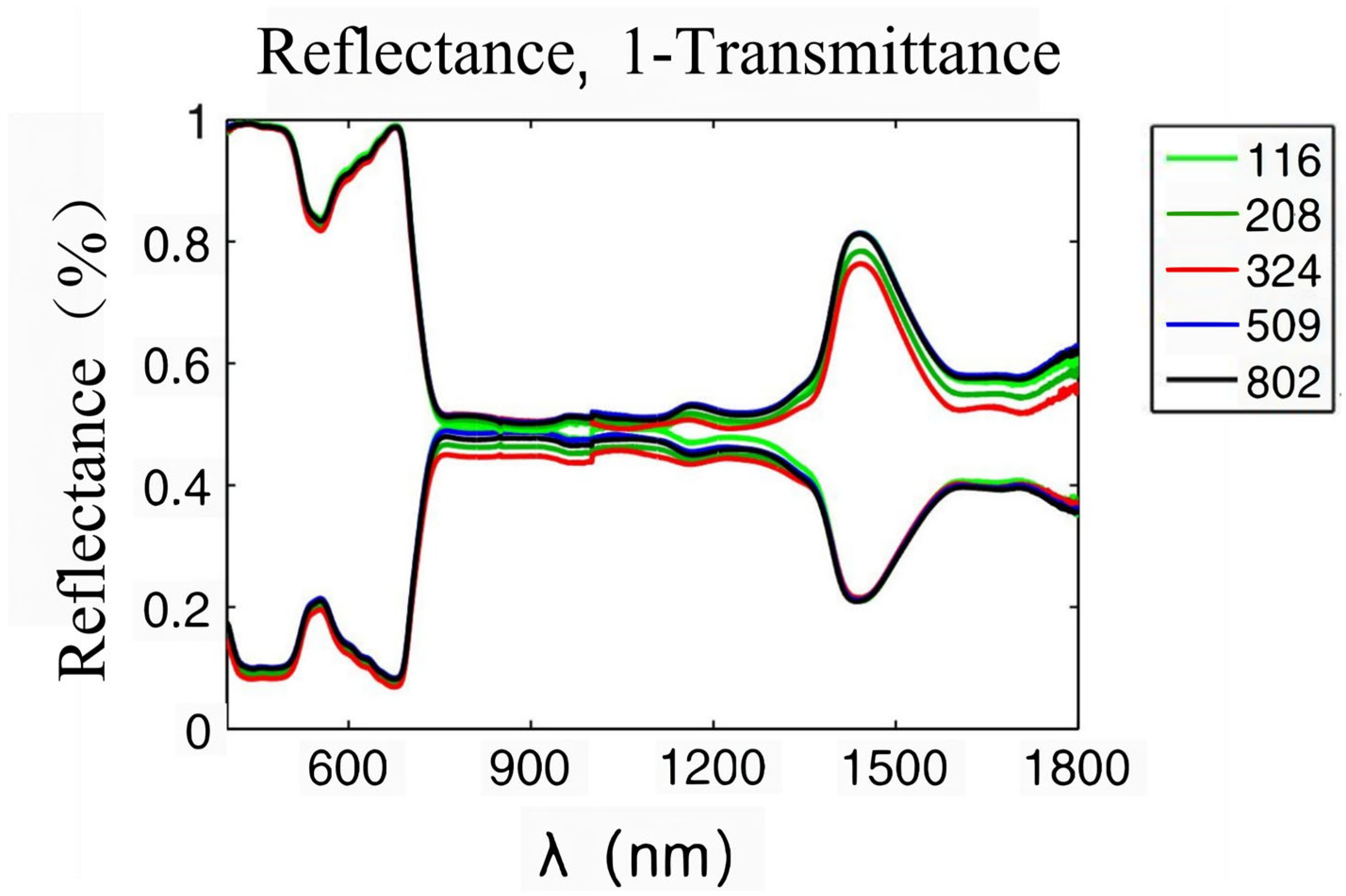
3.1. Research Advances of Plant Hyperspectral RS Technology
3.2. Military Application of Hyperspectral RS
- Detailed battlefield reconnaissance
- Identifying camouflage target
- Detecting and calculating the target’s true temperature and emissivity
4. Research Advances of Plant-Mimicking Camouflage Materials
4.1. Bionic Camouflage
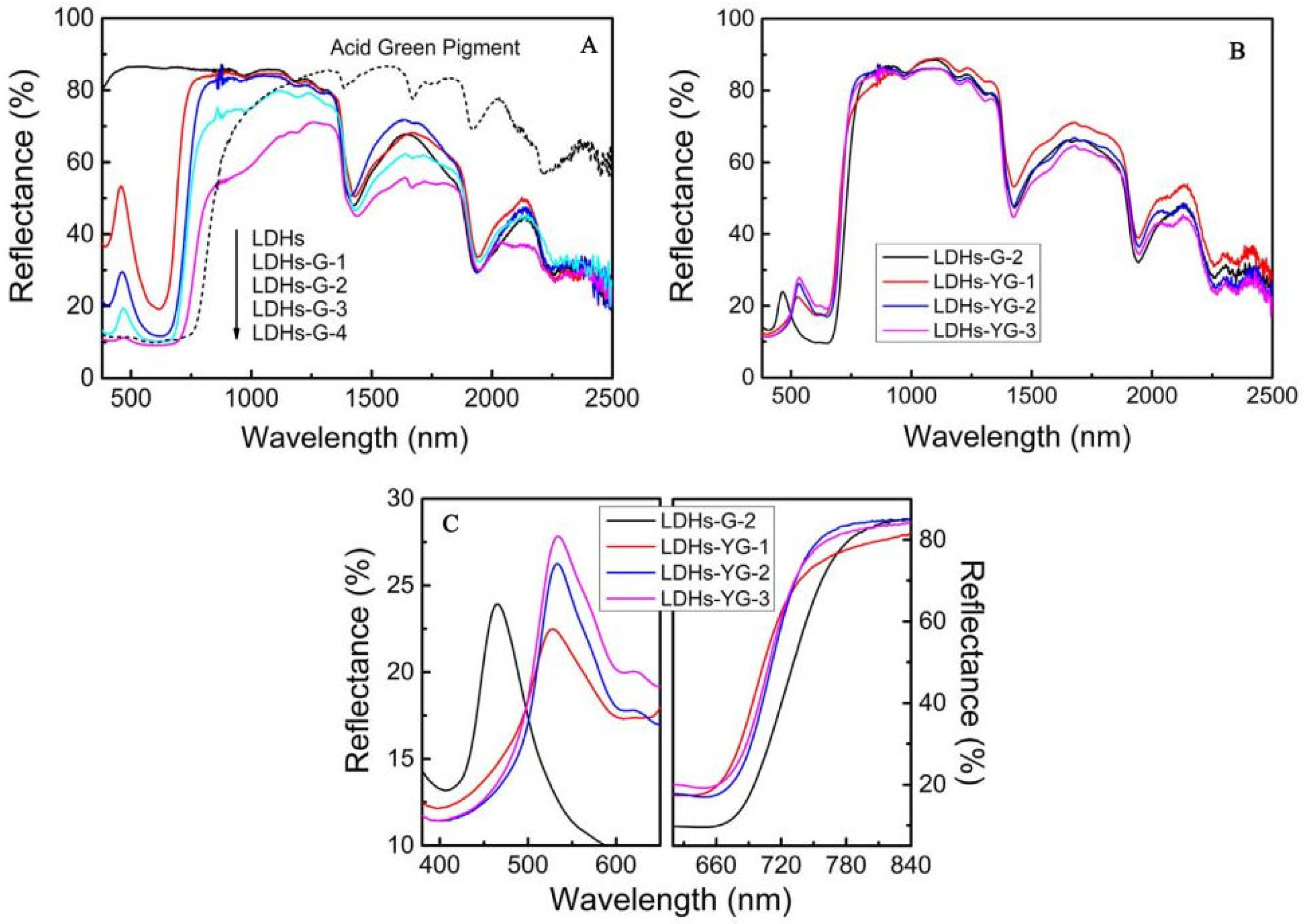
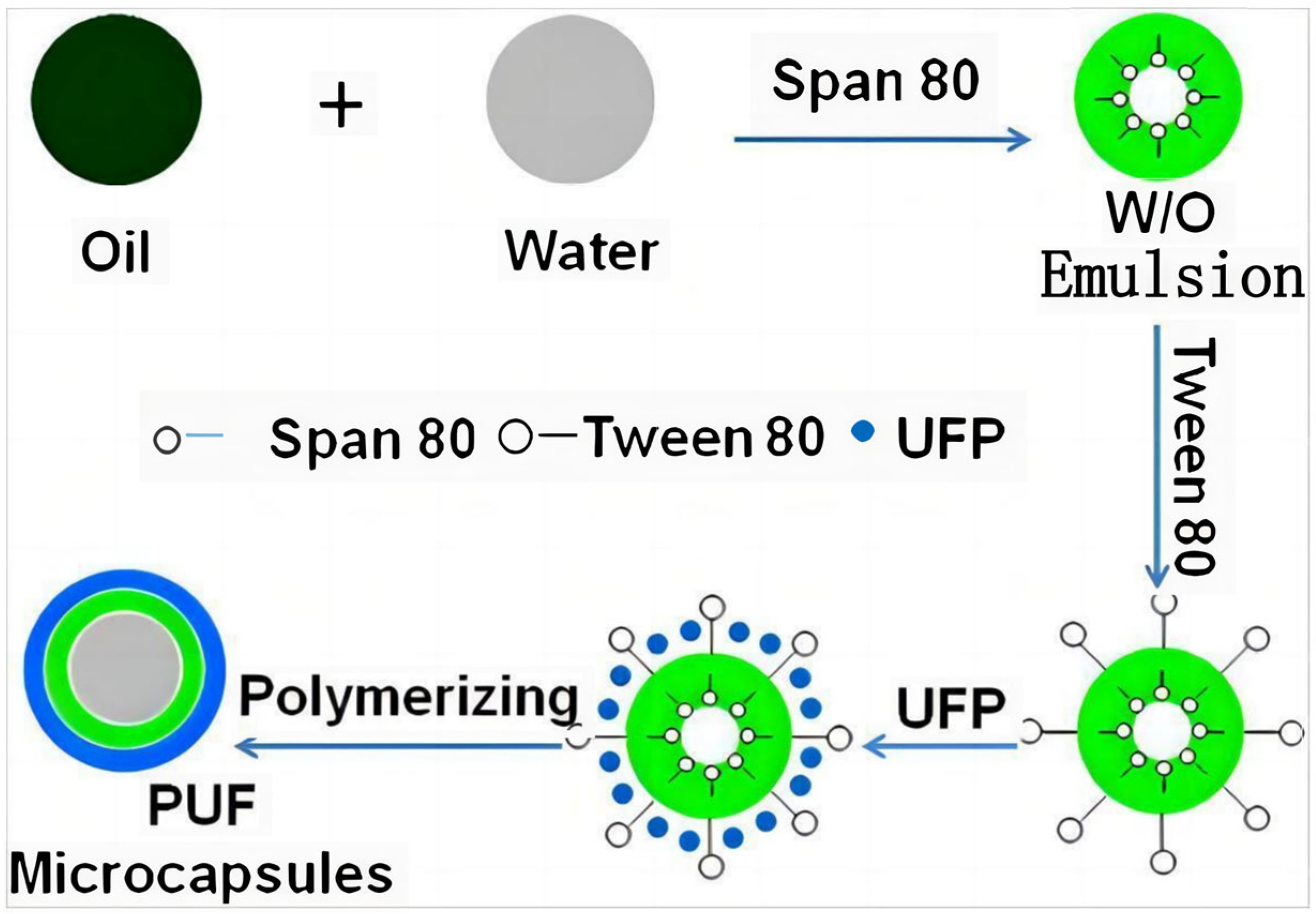
4.2. Bionic Materials Simulating Solar Reflection Characteristics of Leaves
5. Discussion and Prospects
- Improving the bionic camouflage technology
- Improving the performances of leaf-simulating camouflage materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murchie, S.; Arvidson, R.; Bedini, P.; Beisser, K.; Bibring, J.P.; Bishop, J. Compact reconnaissance imaging spectrometer for mars (CRISM) on mars reconnaissance orbiter (MRO). J. Geophys. Res. Atmos. 2007, 112, 431–433. [Google Scholar] [CrossRef]
- Jia, J.; Wang, Y.; Chen, J.; Guo, R.; Shu, R.; Wang, J. Status and application of advanced airborne hyperspectral imaging technology: A review. Infrared Phys. Technol. 2020, 104, 103115. [Google Scholar] [CrossRef]
- Wang, H.; Yan, C.; Yuan, J.; Lu, Q. Hyperspectral band selections for enhancing the discrimination of difficult targets using local band index and particle swarm optimization. Appl. Sci. 2022, 12, 3899. [Google Scholar] [CrossRef]
- Luo, J.; Cai, F.; Yao, X.; Li, J.; Huang, Q.; He, S. Experimental demonstration of an anti-shake hyperspectral imager of high spatial resolution and low cost. IEEE Sens. J. 2020, 20, 8082–8090. [Google Scholar] [CrossRef]
- Shen, F.; Deng, H.; Yu, L.; Cai, F. Open-source mobile multispectral imaging system and its applications in biological sample sensing. Spectrochim. Acta Part A 2022, 280, 121504. [Google Scholar] [CrossRef] [PubMed]
- Dian, R.; Li, S.; Sun, B.; Guo, A. Recent advances and new guidelines on hyperspectral and multispectral image fusion. Inform. Fusion 2021, 69, 40–51. [Google Scholar] [CrossRef]
- Shaw, G.; Manolakis, D. Signal processing for hyperspectral image exploitation. IEEE Signal Proc. Mag. 2002, 19, 12–16. [Google Scholar] [CrossRef]
- Tiwari, K.C.; Arora, M.K.; Singh, D. An assessment of independent component analysis for detection of military targets from hyperspectral images. Int. J. Appl. Earth Obs. 2011, 13, 730–740. [Google Scholar] [CrossRef]
- Makki, I.; Rafic, Y.; Clovis, F.; Tiziano, B.; Massimo, Z. A survey of landmine detection using hyperspectral imaging. ISPRS J. Photogramm. Remote Sens. 2017, 124, 40–53. [Google Scholar] [CrossRef]
- Lee, J.; Sul, H.; Jung, Y.; Kim, H.; Han, S.; Choi, J.H. Thermally controlled, active imperceptible artificial skin in visible-to-infrared range. Adv. Funct. Mater. 2020, 30, 2003328. [Google Scholar] [CrossRef]
- Xu, K.; Ye, H. Preparation and optimization of biomimetic materials simulating solar spectrum reflection characteristics of natural leaves. J. Mater. Sci. 2020, 55, 12848–12863. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.C. Near-infrared green camouflage of cotton fabrics using vat dyes. J. Text. Inst. 2008, 99, 83–88. [Google Scholar] [CrossRef]
- Qin, R.; Xu, G.; Guo, L.; Jiang, Y.; Ding, R. Preparation and characterization of a novel poly(urea-formaldehyde) microcapsules with similar reflectance spectrum to leaves in the UV-Vis-NIR region of 300–2500 nm. Mater. Chem. Phys. 2012, 136, 737–743. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, C.; Qing, X.; Bi, M.; Huang, G.; Weng, X. Synthesis and fine spectroscopy tuning of the hyperspectral simulation material based on organic anions intercalated mg-al layered double hydroxide. Infrared Phys. Technol. 2020, 107, 103328. [Google Scholar] [CrossRef]
- Ye, H.; Gao, Y.; Li, S.; Guo, L. Bionic leaves imitating the transpiration and solar spectrum reflection characteristics of natural leaves. J. Bionic Eng. 2015, 12, 109–116. [Google Scholar] [CrossRef]
- Xu, K.; Lin, Y.; Wei, X.; Ye, H. Biomimetic material simulating solar spectrum reflection characteristics of yellow leaf. J. Bionic Eng. 2018, 15, 741–750. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, B.; Ji, G.; Chen, K.; Wang, Z.; Ye, H. A camouflage coating with similar solar spectrum reflectance to leaves based on polymeric inorganic composite. Mater. Res. Express 2021, 8, 66404. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Smith, R.B.; De Pauw, E. Hyperspectral vegetation indices and their relationships with agricultural crop characteristics. Rem. Sens. Environ. 2000, 71, 158–182. [Google Scholar] [CrossRef]
- Roth, K.L.; Casas, A.; Huesca, M.; Ustin, S.L.; Alsina, M.M.; Mathews, S.A.; Whiting, M.L. Leaf spectral clusters as potential optical leaf functional types within California ecosystems. Remote Sens. Environ. 2016, 184, 229–246. [Google Scholar] [CrossRef]
- Pauli, D.; Chapman, S.C.; Bart, R.; Topp, C.N.; Lawrence-Dill, C.J.; Poland, J.; Gore, M.A. The quest for understanding phenotypic variation via integrated approaches in the field environment. Plant Physiol. 2016, 172, 622–634. [Google Scholar] [CrossRef]
- Kolmann, M.A.; Kalacska, M.; Lucanus, O.; Sousa, L.; Wainwright, D.; Arroyo-Mora, J.P.; Andrade, M.C. Hyperspectral data as a biodiversity screening tool can differentiate among diverse Neotropical fishes. Sci. Rep. 2021, 11, 16157. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, T.C. Leaf: Light Capture in the Photosynthetic Organ; Hohmann-Marriott, M.F., Ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Gerber, F.; Marion, R.; Olioso, A.; Jacquemoud, S.; Ribeiro da Luz, B.; Fabre, S. Modeling directional–hemispherical reflectance and transmittance of fresh and dry leaves from 0.4 μm to 5.7 μm with the PROSPECT-VISIR model. Remote Sens. Environ. 2011, 115, 404–414. [Google Scholar] [CrossRef]
- Vilfan, N.; van der Tol, C.; Muller, O.; Rascher, U.; Verhoef, W. Fluspect-B: A model for leaf fluorescence, reflectance and transmittance spectra. Remote Sens. Environ. 2016, 186, 596–615. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Gates, D.M.; Keegan, H.J.; Schleter, J.C.; Weidner, V.R. Spectrak properties of plants. Appl. Opt. 1965, 4, 11–20. [Google Scholar] [CrossRef]
- Peñuelas, J.; Inoue, Y. Reflectance indices indicative of changes in water and pigment contents of peanut and wheat leaves. Photosynthetica 1999, 36, 355–360. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and physiological basis for the reflectance of visible and near-infrared radiation from vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Hettinger, J.W.; Matthew de la Peña Mattozzi Myers, W.R.; Williams, M.E.; Reeves, A.; Parsons, R.L.; Haskell, R.C.; Petersen, D.C.; Wang, R.; Medford, J.I. Optical coherence microscopy. A technology for rapid, in vivo, non-destructive visualization of plants and plant cells. Plant Physiol. 2000, 123, 3–15. [Google Scholar] [CrossRef]
- Woolley, J.T. Reflectance and transmittance of light by leaves. Plant Physiol. 1971, 47, 656–662. [Google Scholar] [CrossRef]
- Broge, N.H.; Mortensen, J.V. Deriving green crop area index and canopy chlorophyll density of winter wheat from spectral reflectance data. Remote Sens. Environ. 2002, 81, 45–57. [Google Scholar] [CrossRef]
- Hikosaka, K. Optimality of nitrogen distribution among leaves in plant canopies. J. Plant Res. 2016, 129, 299–311. [Google Scholar] [CrossRef]
- Muller, C.; Riederer, M. Plant surface properties in chemical ecology. J. Chem. Ecol. 2005, 31, 2621–2651. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Chivkunova, O.B.; Zhigalova, T.V.; Naqvi, K.R. Light absorption by isolated chloroplasts and leaves: Effects of scattering and packing. Photosynth. Res. 2009, 102, 31–41. [Google Scholar] [CrossRef]
- Huang, Z.; Shi, T.; Zhang, H.; Zhang, M.; Huttula, M.; Cao, W. A computational study of antireflection structures bio-mimicked from leaf surface morphologies. Sol. Energy 2016, 131, 131–137. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. PROSPECT: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Renzullo, L.J.; Blanchfield, A.L.; Guillermin, R.; Powell, K.S.; Held, A.A. Comparison of PROSPECT and HPLC estimates of leaf chlorophyll contents in a grapevine stress study. Int. J. Remote Sens. 2006, 27, 817–823. [Google Scholar] [CrossRef]
- Barry, K.M.; Newnham, G.J.; Stone, C. Estimation of chlorophyll content in eucalyptus globulus foliage with the leaf reflectance model PROSPECT. Agric. For. Meteorol. 2009, 149, 1209–1213. [Google Scholar] [CrossRef]
- Romero, A.; Aguado, I.; Yebra, M. Estimation of dry matter content in leaves using normalized indexes and prospect model inversion. Int. J. Remote Sens. 2012, 33, 396–414. [Google Scholar] [CrossRef]
- Yarbrough, S.; Caudill, T.; Kouba, E.; Osweiler, V.; Arnold, J. Mightysat ii.1 hyperspectral imager: Summary of on-orbit performance. In Proceedings of the SPIE-the International Society for Optical Engineering, Świnoujście, Poland, 23–27 September 2002; pp. 186–197. [Google Scholar]
- Goetz, A.F.H.; Vane, G.; Solomon, J.E.; Rock, B.N. Imaging spectrometry for earth remote sensing. Science 1985, 228, 1147–1153. [Google Scholar] [CrossRef]
- Goetz, A.F.H.; Wellman, J.B.; Barnes, W.L. Optical remote sensing of the earth. Proc. IEEE 1985, 73, 950–969. [Google Scholar] [CrossRef]
- Pinter, P.J.; Jackson, R.D.; Idso, S.B.; Reginato, R.J. Diurnal patterns of wheat spectral reflectances. IEEE Trans. Geosci. Remote Sens. 1983, GE-21, 156–163. [Google Scholar] [CrossRef]
- Curran, P.J.; Steven, M.D. Multispectral remote sensing for the estimation of green leaf area index [and discussion]. Philosophical Transactions of the Royal Society of London. Ser. A Math. Phys. Sci. 1983, 309, 257–270. [Google Scholar]
- Everitt, J.H.; Pettit, R.D.; Alaniz, M.A. Remote sensing of broom snakeweed (Gutierrezia sarothrae) and spiny aster (Aster spinosus). Weed Sci. 1987, 35, 295–302. [Google Scholar] [CrossRef]
- Curran, P.J.; Dungan, J.L.; Peterson, D.L. Estimating the foliar biochemical concentration of leaves with reflectance spectrometry: Testing the kokaly and clark methodologies. Remote Sens. Environ. 2001, 76, 349–359. [Google Scholar] [CrossRef]
- Jago, R.A.; Cutler, M.E.J.; Curran, P.J. Estimating canopy chlorophyll concentration from field and airborne spectra. Remote Sens. Environ. 1999, 68, 217–224. [Google Scholar] [CrossRef]
- Mcfeeters, S.K. The use of the normalized difference water index (NDWI) in the delineation of open water features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Casanova, D.; Epema, G.F.; Goudriaan, J. Monitoring rice reflectance at field level for estimating biomass and LAI. Field Crops Res. 1998, 55, 83–92. [Google Scholar] [CrossRef]
- Mistele, B.; Schmidhalter, U. Spectral measurements of the total aerial N and biomass dry weight in maize using a quadrilateral-view optic. Field Crops Res. 2008, 106, 94–103. [Google Scholar] [CrossRef]
- Tang, X.G.; Song, K.S.; Liu, D.W.; Wang, Z.M.; Zhang, B.; Du, J.; Zeng, L.H.; Jiang, G.J.; Wang, Y.D. Comparison of methods for estimating soybean chlorophyll content based on visual/near infrared reflection spectra. Spectrosc. Spect. Anal. 2011, 31, 371–374. [Google Scholar]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Tree species classification in the southern alps based on the fusion of very high geometrical resolution multispectral/hyperspectral images and LiDAR data. Remote Sens. Environ. 2012, 123, 258–270. [Google Scholar] [CrossRef]
- Kozoderov, V.V.; Dmitriev, E.V. Testing different classification methods in airborne hyperspectral imagery processing. Opt. Express 2016, 24, A956–A965. [Google Scholar] [CrossRef] [PubMed]
- Schull, M.A.; Knyazikhin, Y.; Xu, L.; Samanta, A.; Carmona, P.L.; Lepine, L.; Jenkins, J.P.; Ganguly, S.; Myneni, R.B. Canopy spectral invariants, part 2: Application to classification of forest types from hyperspectral data. J. Quant. Spectrosc. Radiat. Transf. 2011, 112, 736–750. [Google Scholar] [CrossRef]
- Ballanti, L.; Blesius, L.; Hines, E.; Kruse, B. Tree species classification using hyperspectral imagery: A comparison of two classifiers. Remote Sens. 2016, 8, 445. [Google Scholar] [CrossRef]
- Vangi, E.; D’amico, G.; Francini, S.; Giannetti, F.; Lasserre, B.; Marchetti, M.; Chirici, G. The new hyperspectral satellite prisma: Imagery for forest types discrimination. Sensors 2021, 21, 1182. [Google Scholar] [CrossRef] [PubMed]
- Modzelewska, A.; Kamińska, A.; Fassnacht, F.E.; Stereńczak, K. Multitemporal hyperspectral tree species classification in the białowieża forest world heritage site. Forestry 2021, 94, 464–476. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Lyon, J.G.; Huete, A. Fundamentals, Sensor Systems, Spectral Libraries, and Data Mining for Vegetation, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Durán, S.M.; Martin, R.E.; Díaz, S.; Maitner, B.S.; Malhi, Y.; Salinas, N.; Shenkin, A.; Silman, M.R.; Wieczynski, D.J.; Asner, G.P.; et al. Informing trait-based ecology by assessing remotely sensed functional diversity across a broad tropical temperature gradient. Sci. Adv. 2019, 5, eaaw8114. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Jiang, X.; Zhen, J.; Miao, J.; Zhao, D.; Wang, J.; Jia, S. Assessing mangrove leaf traits under different pest and disease severity with hyperspectral imaging spectroscopy. Ecol. Indic. 2021, 129, 107901. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, X.; Yang, G.; Zhu, C.; Huo, L.; Feng, H. Assessment of defoliation during the dendrolimus tabulaeformis tsai et liu disaster outbreak using UAV-based hyperspectral images. Remote Sens. Environ. 2018, 217, 323–339. [Google Scholar] [CrossRef]
- Ma, Y.; Lu, J.; Huang, X. Damage diagnosis of pinus yunnanensis canopies attacked by tomicus using UAV hyperspectral images. Forests 2023, 14, 61. [Google Scholar] [CrossRef]
- Zhang, X.; Han, L.; Dong, Y.; Shi, Y.; Huang, W.; Han, L.; González-Moreno, P.; Ma, H.; Ye, H.; Sobeih, T. A deep learning-based approach for automated yellow rust disease detection from high-resolution hyperspectral UAV images. Remote Sens. 2019, 11, 1554. [Google Scholar] [CrossRef]
- Xi, G.; Huang, X.; Xie, Y.; Gang, B.; Bao, Y.; Dashzebeg, G.; Nanzad, T.; Dorjsuren, A.; Enkhnasan, D.; Ariunaa, M. Detection of larch forest stress from jas’s larch inchworm (erannis jacobsoni djak) attack using hyperspectral remote sensing. Remote Sens. 2022, 14, 124. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Z.; Liu, X.; Yao, J.; Du, T.; Ma, Y.; Shi, L. Discriminant analysis of the damage degree caused by pine shoot beetle to yunnan pine using UAV-based hyperspectral images. Forests 2020, 11, 1258. [Google Scholar] [CrossRef]
- Gao, B.; Yu, L.; Ren, L.; Zhan, Z.; Luo, Y. Early detection of dendroctonus valens infestation at tree level with a hyperspectral UAV image. Remote Sens. 2023, 15, 407. [Google Scholar] [CrossRef]
- Bergman, S.M. The Utility of Hyperspectral Data to Detect and Discriminate Actual and Decoy Target Vehicles; Directorate for Information Operations and Reports: Washington, DC, USA, 1996. [Google Scholar]
- Li, X.P.; Chen, J.P.; Wang, X. Inversion of lunar nearside FeO and Al2O3 based on Chang’E-1 reflectance data. China Min. Mag. 2018, 27, 150–156. [Google Scholar]
- Bárta, V.; Racek, F.; Krejcí, J. NATO hyperspectral measurement of natural background. Int. Soc. Opt. Eng. 2018, 10794, 1–16. [Google Scholar]
- Stellman, C.M.; Hazel, G.G.; Schuler, J.M. Spectral Calibration, Spatial Mapping and Flat Fielding Studies of the Dark Horse 1(DH1) March Data Collection; Naval Research Laboratory: Washington, DC, USA, 1999. [Google Scholar]
- Zagolski, F.; Pinel, V.; Romier, J.; Alcayde, D.; Fontanari, J.; Gastellu-etchegorry, J.P.; Giordano, G.; Marty, G.; Mougin, E.; Joffre, R. Forest canopy chemistry with high spectral resolution remote sensing. Int. J. Remote Sens. 1996, 17, 1107–1128. [Google Scholar] [CrossRef]
- Junttila, M.L. Stationary fourier-transform spectrometer. Appl. Opt. 1992, 31, 4106–4112. [Google Scholar] [CrossRef]
- Incropera, F.P.; DeWitt, D.P. Fundamentals of Heat Transfer; Wiley: Hoboken, NJ, USA, 1981. [Google Scholar]
- Lichtenthaler, H.K.; Gitelson, A.; Lang, M. Non-destructive determination of chlorophyll content of leaves of a green and an aurea mutant of tobacco by reflectance measurements. J. Plant Physiol. 1996, 148, 483–493. [Google Scholar] [CrossRef]
- Bank, S.; Cumming, R.T.; Li, Y.; Henze, K.; Le Tirant, S.; Bradleret, S. A tree of leaves: Phylogeny and historical biogeography of the leaf insects (Phasmatodea: Phylliidae). Commun. Biol. 2021, 4, 932. [Google Scholar] [CrossRef]
- Villoutreix, R.; de Carvalho, C.F.; Feder, J.L.; Gompert, Z.; Nosil, P. Disruptive selection and the evolution of discrete color morphs in timema stick insects. Sci. Adv. 2023, 9, eabm8157. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.T.; Zhu, K.; Petracca, J.; Wurtzel, E.T. Analysis of plant-derived carotenoids in camouflaging stick and leaf insects (phasmatodea). Method Enzymol. 2022, 670, 499–524. [Google Scholar]
- Hui, Z.; Jianchun, Z. Near-infrared green camouflage of PET fabrics using disperse dyes. Sen’i Gakkaishi 2007, 63, 223–229. [Google Scholar] [CrossRef]
- Zhou, L.; Shao, J.Z.; Chai, L.Q. Study on the uv-protective properties and near infrared camouflage of lithospermum. J. Fiber Bioeng. Inform. 2009, 2, 177–181. [Google Scholar]
- Khajeh Mehrizi, M.; Mortazavi, S.M.; Mallakpour, S.; Bidoki, S.M.; Vik, M.; Vikova, M. Effect of carbon black nanoparticles on reflective behavior of printed cotton/nylon fabrics in visible/near infrared regions. Fibers Polym. 2012, 13, 501–506. [Google Scholar] [CrossRef]
- Winkelmann, M. Analysis of exploitable spectral features of target and background materials. In Proceedings of the SPIE Defence and Security Toulouse, Toulouse, France, 21–23 September 2015; p. 9653. [Google Scholar]
- Lv, C.; Zu, M.; Xie, D.; Cheng, H. Emulating solar spectral reflectance of natural leaf with bionic leaf prepared from 4a zeolite-derived ultramarine green pigment. Materials 2021, 14, 1406. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ye, H. Bionic membrane simulating solar spectrum reflection characteristics of natural leaf. Int. J. Heat Mass Transf. 2017, 114, 115–124. [Google Scholar] [CrossRef]
- Ai, S.; Zheng, H.; Yu, J. Preparation and reflectance spectrum modulation of Cr2O3 green pigment by solution combustion synthesis. Materials 2020, 13, 1540. [Google Scholar] [CrossRef]
- Xie, D.; Lü, C.; Zu, M.; Cheng, H. Research progress of bionic materials simulating vegetation visible-near infrared reflectance spectra. Spectrosc. Spect. Anal. 2021, 41, 1032–1038. [Google Scholar]
- Lu, Q.; Li, M.; Tian, A.; Fu, S. Green plant leaf-inspired smart camouflage fabrics for visible light and near-infrared stealth. J. Bionic Eng. 2022, 19, 788–798. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, W.; Hu, B. Design of biomimetic camouflage materials based on angiosperm leaf organs. Sci. China Ser. E 2008, 51, 1902–1910. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Z.; Hu, B.; Wu, W. Design of plant leaf bionic camouflage materials based on spectral analysis. Spectrosc. Spect. Anal. 2011, 31, 1668–1672. [Google Scholar]
- Yang, Y.; Liu, Z.; Hu, B.; Man, Y.; Wu, W. Bionic composite material simulating the optical spectra of plant leaves. J. Bionic Eng. 2010, 7 (Suppl. 4), S43–S49. [Google Scholar] [CrossRef]
- Hu, A.; Li, M.; Zhang, L.; Wang, C.; Fu, S. Polyurethane-based bionic material simulating the vis-NIR spectrum and thermal infrared properties of vegetation. RSC Adv. 2019, 9, 41438–41446. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wang, S.; Wang, L.; Ye, H. Bionic coating imitating reflection characteristics of plant leaf in solar spectrum waveband for hyperspectral camouflage. Infrared Phys. Technol. 2022, 127, 104477. [Google Scholar] [CrossRef]
- Zhang, X.; Goldberg, M.D. Monitoring fall foliage coloration dynamics using time-series satellite data. Remote Sens. Environ. 2011, 115, 382–391. [Google Scholar] [CrossRef]
- Rozenstein, O.; Adamowski, J. Linking spaceborne and ground observations of autumn foliage senescence in southern québec, Canada. Remote Sens. 2017, 9, 630. [Google Scholar] [CrossRef]
- Xie, Y.; Civco, D.L.; Silander, J.A. Species-specific spring and autumn leaf phenology captured by time-lapse digital cameras. Ecosphere 2018, 9, e02089. [Google Scholar] [CrossRef]
- Anav, A.; Liu, Q.; De Marco, A.; Proietti, C.; Savi, F.; Paoletti, E.; Piao, S. The role of plant phenology in stomatal ozone flux modeling. Glob. Chang. Biol. 2018, 24, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Zu, M.; Li, M.; Liu, D.; Wang, Z.; Li, Q.; Cheng, H. A hyperspectral camouflage colorant inspired by natural leaves. Adv. Mater. 2023, 35, e2302973. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Long, L.; Gao, Y.; Tang, Z.; Zhang, J.; Xu, K.; Ye, H.; Liu, M. A color-changing biomimetic material closely resembling the spectral characteristics of vegetation foliage. Small 2023, 20, e2303966. [Google Scholar] [CrossRef]
- Xu, K.; Long, L.; Yang, W.; Huang, Z.; Ye, H. Bionic metamaterial for multispectral-compatible camouflage of solar spectrum and infrared in the background of vegetation. Cell Rep. Phys. Sci. 2024, 5, 101798. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Ren, L.; Yang, X.; Yuan, H. Design and Application of Bionic Camouflage Materials Simulating Spectral Reflection Characteristics of Plants: A Review. Appl. Sci. 2024, 14, 4404. https://doi.org/10.3390/app14114404
Lin Y, Ren L, Yang X, Yuan H. Design and Application of Bionic Camouflage Materials Simulating Spectral Reflection Characteristics of Plants: A Review. Applied Sciences. 2024; 14(11):4404. https://doi.org/10.3390/app14114404
Chicago/Turabian StyleLin, Yanping, Luquan Ren, Xiaodong Yang, and Hengyi Yuan. 2024. "Design and Application of Bionic Camouflage Materials Simulating Spectral Reflection Characteristics of Plants: A Review" Applied Sciences 14, no. 11: 4404. https://doi.org/10.3390/app14114404
APA StyleLin, Y., Ren, L., Yang, X., & Yuan, H. (2024). Design and Application of Bionic Camouflage Materials Simulating Spectral Reflection Characteristics of Plants: A Review. Applied Sciences, 14(11), 4404. https://doi.org/10.3390/app14114404