
Protective Effects of Fermented Glasswort (Salicornia herbacea L.) on Aged Gut Induced by D-Galactose in Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Glasswort
2.2. Animal Study
2.3. TBARS Assay
2.4. Western Blotting
2.5. Analysis of SCFAs Using GC
2.6. Histology
2.7. Statistical Analysis
3. Results
3.1. General Characteristics
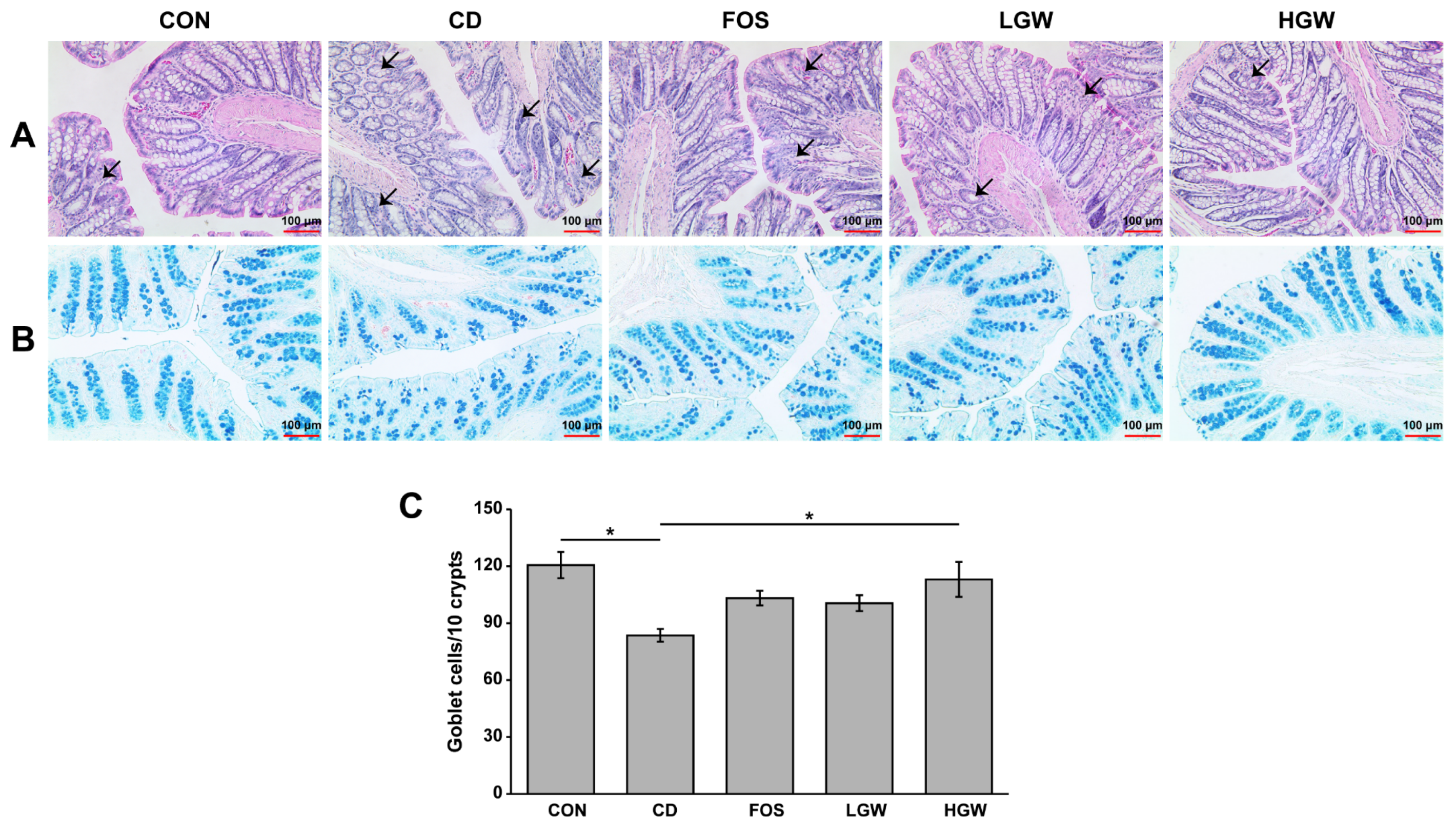
3.2. Histopathology and Number of Goblet Cells
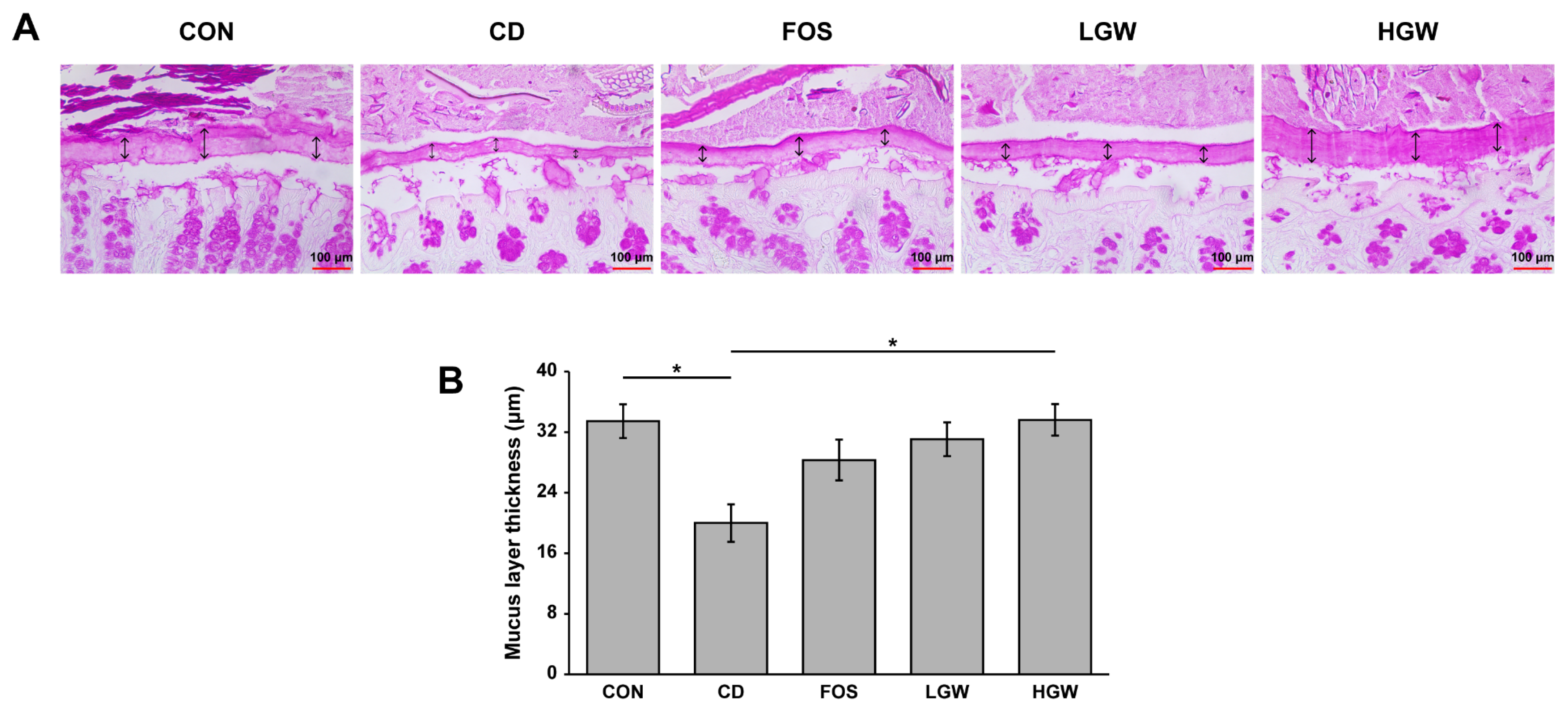
3.3. Mucus Layer Thickness of Colon
3.4. Short-Chain Fatty Acids (SCFAs)
3.5. Oxidative Stress-Related Markers
3.6. Expression Levels of Inflammation-Related Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hohman, L.S.; Osborne, L.C. A Gut-Centric View of Aging: Do Intestinal Epithelial Cells Contribute to Age-Associated Microbiota Changes, Inflammaging, and Immunosenescence? Aging Cell 2022, 21, e13700. [Google Scholar] [CrossRef] [PubMed]
- Wiertsema, S.P.; van Bergenhenegouwen, J.; Garssen, J.; Knippels, L.M.J. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients 2021, 13, 886. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vermeire, S. The Intestinal Barrier: A Fundamental Role in Health and Disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Elderman, M.; Sovran, B.; Hugenholtz, F.; Graversen, K.; Huijskes, M.; Houtsma, E.; Belzer, C.; Boekschoten, M.; De Vos, P.; Dekker, J.; et al. The Effect of Age on the Intestinal Mucus Thickness, Microbiota Composition and Immunity in Relation to Sex in Mice. PLoS ONE 2017, 12, e0184274. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Jeffery, I.B. Gut Microbiota and Aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef]
- Ren, M.; Li, M.Y.; Lu, L.Q.; Liu, Y.S.; An, F.K.; Huang, K.; Fu, Z. Arenga Pinnata Resistant Starch Modulate Gut Microbiota and Ameliorate Intestinal Inflammation in Aged Mice. Nutrients 2022, 14, 3931. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Min, J.-G.; Lee, D.-S.; Kim, T.-J.; Park, J.-H.; Cho, T.-Y.; Park, D.-I. Chemical Composition of S. herbacea L. Prev. Nutr. Food Sci. 2002, 7, 105–107. [Google Scholar] [CrossRef]
- Bailey, R.L.; West, K.P.; Black, R.E. The Epidemiology of Global Micronutrient Deficiencies. Ann. Nutr. Metab. 2015, 66, 22–33. [Google Scholar] [CrossRef]
- Hibberd, M.C.; Wu, M.; Rodionov, D.A.; Li, X.; Cheng, J.; Griffin, N.W.; Barratt, M.J.; Giannone, R.J.; Hettich, R.L.; Osterman, A.L.; et al. The Effects of Micronutrient Deficiencies on Bacterial Species from the Human Gut Microbiota. Sci. Transl. Med. 2017, 9, eaal4069. [Google Scholar] [CrossRef]
- Venturelli, S.; Leischner, C.; Helling, T.; Renner, O.; Burkard, M.; Marongiu, L. Minerals and Cancer: Overview of the Possible Diagnostic Value. Cancers 2022, 14, 1256. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. Anti-Inflammatory Effects of Phytochemicals from Fruits, Vegetables, and Food Legumes: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Bachheti, R.K.; Worku, L.A.; Gonfa, Y.H.; Zebeaman, M.; Deepti; Pandey, D.P.; Bachheti, A. Prevention and Treatment of Cardiovascular Diseases with Plant Phytochemicals: A Review. Evid.-Based Complement. Altern. Med. 2022, 2022, 5741198. [Google Scholar] [CrossRef] [PubMed]
- Afrin, S.; Giampieri, F.; Gasparrini, M.; Forbes-Hernández, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Zhang, J.; Manna, P.P.; Daglia, M.; Atanasov, A.G.; et al. Dietary Phytochemicals in Colorectal Cancer Prevention and Treatment: A Focus on the Molecular Mechanisms Involved. Biotechnol. Adv. 2020, 38, 107322. [Google Scholar] [CrossRef] [PubMed]
- Dingeo, G.; Brito, A.; Samouda, H.; Iddir, M.; La Frano, M.R.; Bohn, T. Phytochemicals as Modifiers of Gut Microbial Communities. Food Funct. 2020, 11, 8444–8471. [Google Scholar] [CrossRef]
- Cho, J.Y.; Kim, J.Y.; Lee, Y.G.; Lee, H.J.; Shim, H.J.; Lee, J.H.; Kim, S.J.; Ham, K.S.; Moon, J.H. Four New Dicaffeoylquinic Acid Derivatives from Glasswort (Salicornia herbacea L.) and Their Antioxidative Activity. Molecules 2016, 21, 1097. [Google Scholar] [CrossRef]
- Park, S.Y.; Cho, J.Y.; Chung, D.O.; Ham, K.S. Physicochemical Characteristics and Physiological Activities of Naturally Fermented Glasswort (Salicornia herbacea L.) Juice. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1493–1500. [Google Scholar] [CrossRef]
- Seo, H.-B.; Kwak, Y.-Y.; Nam, J.-O.; Song, Y.-J.; Kim, B.-O.; Ryu, S.-P. Glasswort Powder Diet Activates Lipid Metabolism in Rat. J. Life Sci. 2012, 22, 478–485. [Google Scholar] [CrossRef]
- Im, S.A.; Kim, K.; Lee, C.K. Immunomodulatory Activity of Polysaccharides Isolated from Salicornia herbacea. Int. Immunopharmacol. 2006, 6, 1451–1458. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and Vegetables, as a Source of Nutritional Compounds and Phytochemicals: Changes in Bioactive Compounds during Lactic Fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Hsiao, Y.H.; Ho, C.T.; Pan, M.H. Bioavailability and Health Benefits of Major Isoflavone Aglycones and Their Metabolites. J. Funct. Foods 2020, 74, 104164. [Google Scholar] [CrossRef]
- Baruah, R.; Ray, M.; Halami, P.M. Preventive and Therapeutic Aspects of Fermented Foods. J. Appl. Microbiol. 2022, 132, 3476–3489. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-Microbiota-Targeted Diets Modulate Human Immune Status. Cell 2021, 184, 4137–4153. [Google Scholar] [CrossRef] [PubMed]
- Park, D.H.; Kothari, D.; Niu, K.M.; Han, S.G.; Yoon, J.E.; Lee, H.G.; Kim, S.K. Effect of Fermented Medicinal Plants as Dietary Additives on Food Preference and Fecal Microbial Quality in Dogs. Animals 2019, 9, 690. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.H.; Kim, K.R. The Proximate and Antioxidative Effect of Dried and Fermented Salicornia herbacea. J. Investig. Cosmetol. 2011, 7, 425–431. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of Malonaldehyde Precursor in Tissues by Thiobarbituric Acid Test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Sangpreecha, N.; Chanmuang, S.; Park, K.H.; Sangar, M.; Sharma, D.; Song, D.; Park, Y.J.; Sung, H.M.; Promyo, K.; Ham, K.S. Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats. Appl. Sci. 2023, 13, 1590. [Google Scholar] [CrossRef]
- Erben, U.; Christoph Loddenkemper, K.D.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A Guide to Histomorphological Evaluation of Intestinal Inflammation in Mouse Models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Ballard, J.W.O.; Towarnicki, S.G. Mitochondria, the Gut Microbiome and ROS. Cell. Signal. 2020, 75, 109737. [Google Scholar] [CrossRef]
- Martel, J.; Chang, S.H.; Ko, Y.F.; Hwang, T.L.; Young, J.D.; Ojcius, D.M. Gut Barrier Disruption and Chronic Disease. Trends Endocrinol. Metab. 2022, 33, 247–265. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. D-Galactose-Induced Accelerated Aging Model: An Overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef] [PubMed]
- Matt, S.M.; Allen, J.M.; Lawson, M.A.; Mailing, L.J.; Woods, J.A.; Johnson, R.W. Butyrate and Dietary Soluble Fiber Improve Neuroinflammation Associated with Aging in Mice. Front. Immunol. 2018, 9, 1832. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.M.; Pham, P.T.; Bach, T.Q.; Ngo, A.T.L.; Nguyen, Q.T.; Phan, T.T.K.; Nguyen, G.H.; Le, P.T.T.; Hoang, V.T.; Forsyth, N.R.; et al. Stem Cell-Based Therapy for Human Diseases. Signal Transduct. Target. Ther. 2022, 7, 272. [Google Scholar] [CrossRef] [PubMed]
- Nalapareddy, K.; Zheng, Y.; Geiger, H. Aging of Intestinal Stem Cells. Stem Cell Rep. 2022, 17, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Gallego, P.; Garcia-Bonete, M.J.; Trillo-Muyo, S.; Recktenwald, C.V.; Johansson, M.E.V.; Hansson, G.C. The Intestinal MUC2 Mucin C-Terminus Is Stabilized by an Extra Disulfide Bond in Comparison to von Willebrand Factor and Other Gel-Forming Mucins. Nat. Commun. 2023, 14, 1969. [Google Scholar] [CrossRef] [PubMed]
- Sovran, B.; Hugenholtz, F.; Elderman, M.; Van Beek, A.A.; Graversen, K.; Huijskes, M.; Boekschoten, M.V.; Savelkoul, H.F.J.; De Vos, P.; Dekker, J.; et al. Age-Associated Impairment of the Mucus Barrier Function Is Associated with Profound Changes in Microbiota and Immunity. Sci. Rep. 2019, 9, 1437. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Y.; Ma, F.; Sun, M.; Song, Y.; Xu, D.; Mu, G.; Tuo, Y. Lactobacillus Plantarum Y44 Alleviates Oxidative Stress by Regulating Gut Microbiota and Colonic Barrier Function in Balb/C Mice with Subcutaneous d-Galactose Injection. Food Funct. 2021, 12, 373–386. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, Y.; Li, B.; Xie, Y.; Shi, Y.; Le, G. Dietary methionine restriction improves gut microbiota composition and prevents cognitive impairment in D-galactose-induced aging mice. Food Funct. 2022, 13, 12896–12914. [Google Scholar] [CrossRef]
- On, J.Y.; Kim, S.H.; Kim, J.M.; Park, S.; Kim, K.H.; Lee, C.H.; Kim, S.K. Effects of Fermented Artemisia annua L. and Salicornia herbacea L. on Inhibition of Obesity In Vitro and In Mice. Nutrients 2023, 15, 2022. [Google Scholar] [CrossRef]
- Hossen, I.; Hua, W.; Ting, L.; Mehmood, A.; Jingyi, S.; Duoxia, X.; Yanping, C.; Hongqing, W.; Zhipeng, G.; Kaiqi, Z.; et al. Phytochemicals and Inflammatory Bowel Disease: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1321–1345. [Google Scholar] [CrossRef]
- Donati Zeppa, S.; Agostini, D.; Ferrini, F.; Gervasi, M.; Barbieri, E.; Bartolacci, A.; Piccoli, G.; Saltarelli, R.; Sestili, P.; Stocchi, V. Interventions on Gut Microbiota for Healthy Aging. Cells 2023, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, H.; Wang, R.; Yu, Y.; Liu, X.; Tian, Z. Protective Effect of Sodium Butyrate on Intestinal Barrier Damage and Uric Acid Reduction in Hyperuricemia Mice. Biomed. Pharmacother. 2023, 161, 114568. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- On, J.Y.; Kim, J.M.; Kothari, D.; Kim, S.K. Antioxidant Properties and Kidney Cell Protection by the Extracts of Curcuma Longa, Artemisia Princeps, Salicornia herbacea, and Schisandra chinesis. Fermentation 2022, 8, 702. [Google Scholar] [CrossRef]
- Altay, A.; Celep, G.S.; Yaprak, A.E.; Baskose, I.; Bozoglu, F. Glassworts as Possible Anticancer Agents Against Human Colorectal Adenocarcinoma Cells with Their Nutritive, Antioxidant and Phytochemical Profiles. Chem. Biodivers. 2017, 14, e1600290. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.T.; Vasconcelos, Q.D.J.S.; Aragão, G.F. Fructooligosaccharides on Inflammation, Immunomodulation, Oxidative Stress, and Gut Immune Response: A Systematic Review. Nutr. Rev. 2022, 80, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, D.; Sangpreecha, N.; Chanmuang, S.; Park, Y.-K.; Ham, K.-S. Protective Effects of Fermented Glasswort (Salicornia herbacea L.) on Aged Gut Induced by D-Galactose in Rats. Appl. Sci. 2023, 13, 8386. https://doi.org/10.3390/app13148386
Song D, Sangpreecha N, Chanmuang S, Park Y-K, Ham K-S. Protective Effects of Fermented Glasswort (Salicornia herbacea L.) on Aged Gut Induced by D-Galactose in Rats. Applied Sciences. 2023; 13(14):8386. https://doi.org/10.3390/app13148386
Chicago/Turabian StyleSong, Doyoung, Neeracha Sangpreecha, Saoraya Chanmuang, Yang-Kyun Park, and Kyung-Sik Ham. 2023. "Protective Effects of Fermented Glasswort (Salicornia herbacea L.) on Aged Gut Induced by D-Galactose in Rats" Applied Sciences 13, no. 14: 8386. https://doi.org/10.3390/app13148386
APA StyleSong, D., Sangpreecha, N., Chanmuang, S., Park, Y.-K., & Ham, K.-S. (2023). Protective Effects of Fermented Glasswort (Salicornia herbacea L.) on Aged Gut Induced by D-Galactose in Rats. Applied Sciences, 13(14), 8386. https://doi.org/10.3390/app13148386