
Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions

 ,
,  , ,
, ,
Abstract
:1. Introduction
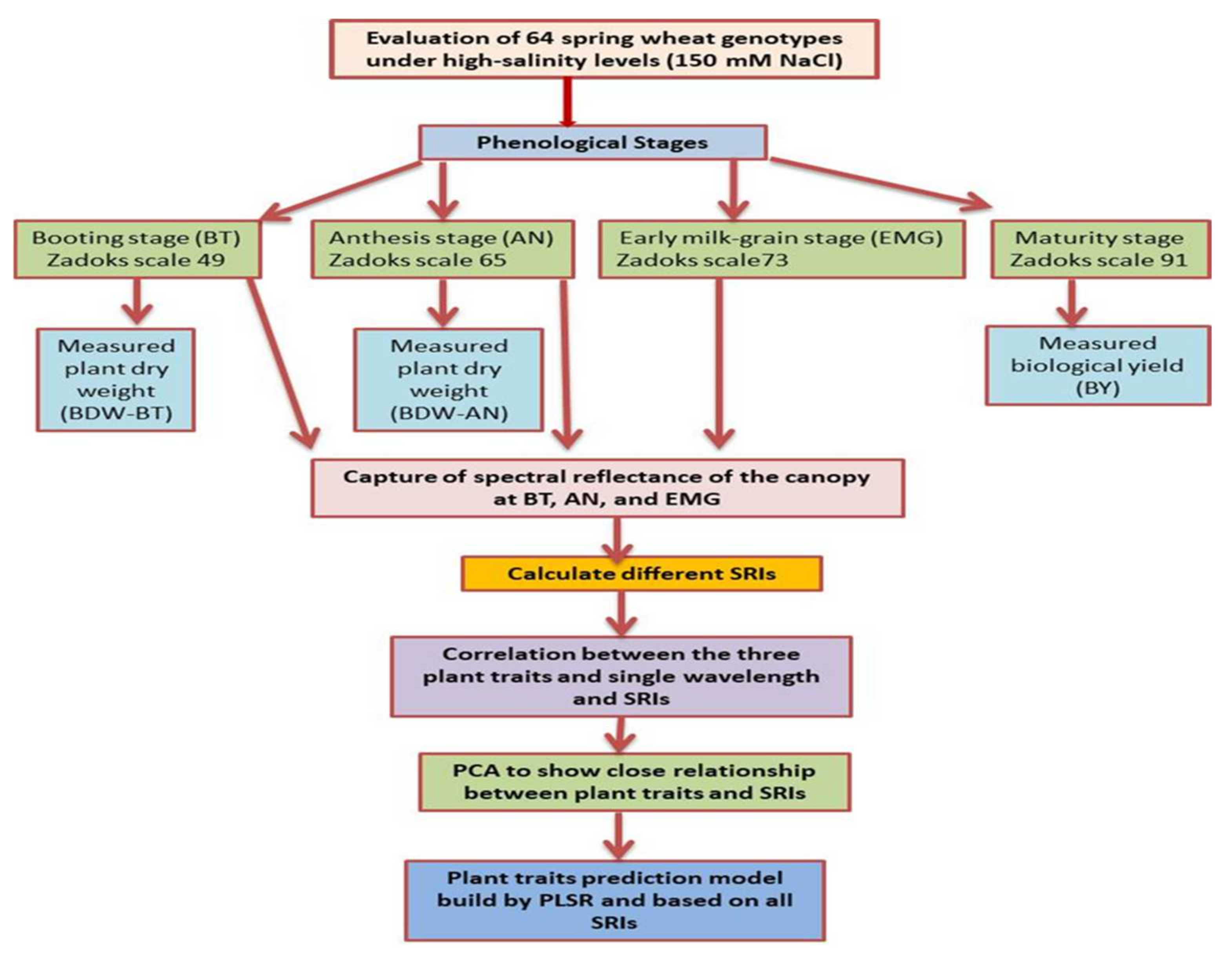
2. Materials and Methods
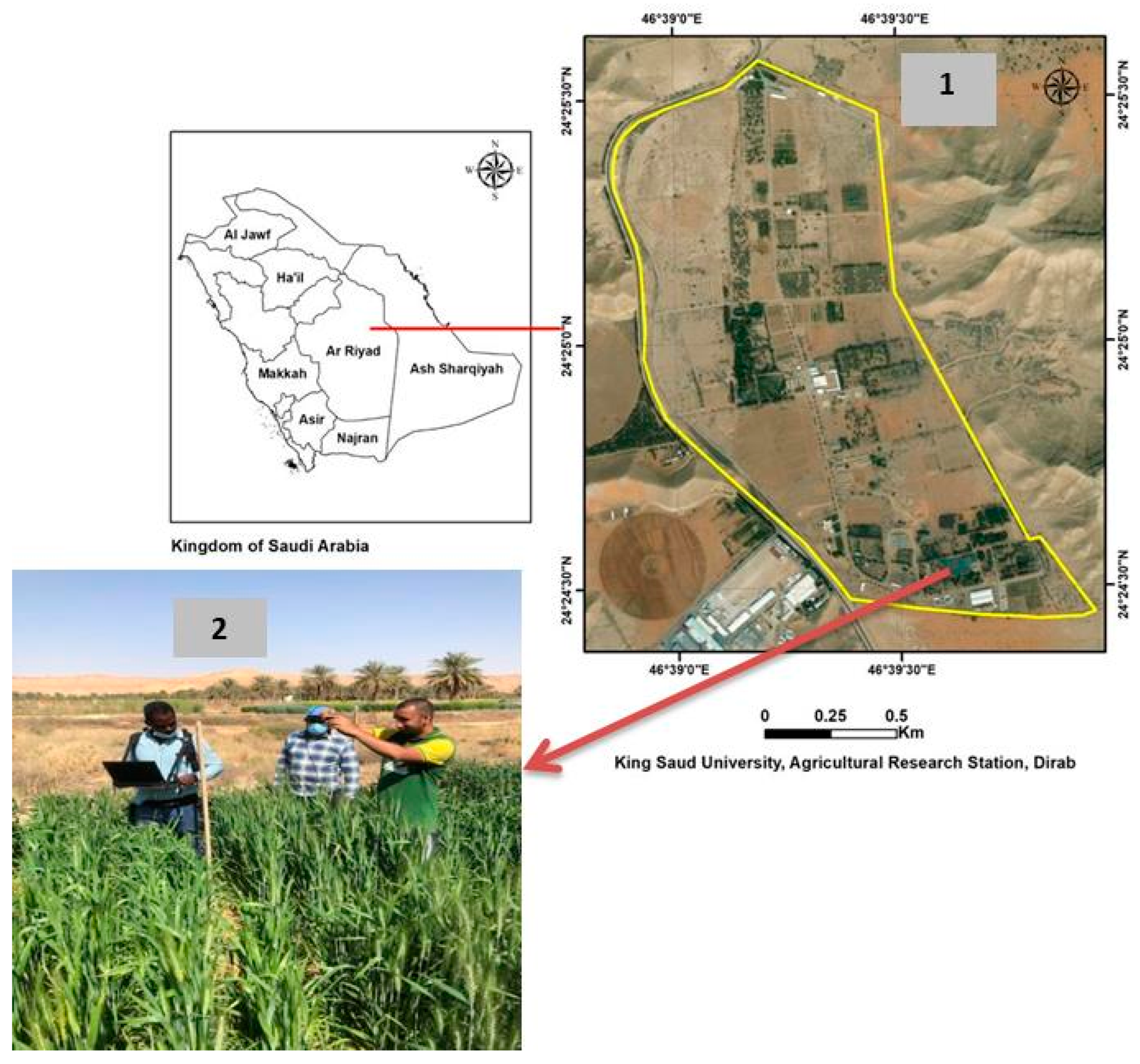
2.1. Plant Materials
2.2. Measurements of Biomass and Biological Yield
2.3. Spectroradiometric Data and Processing
2.4. Data Analysis
3. Results
3.1. Genotypic Variations for Biomass and Biological Yield
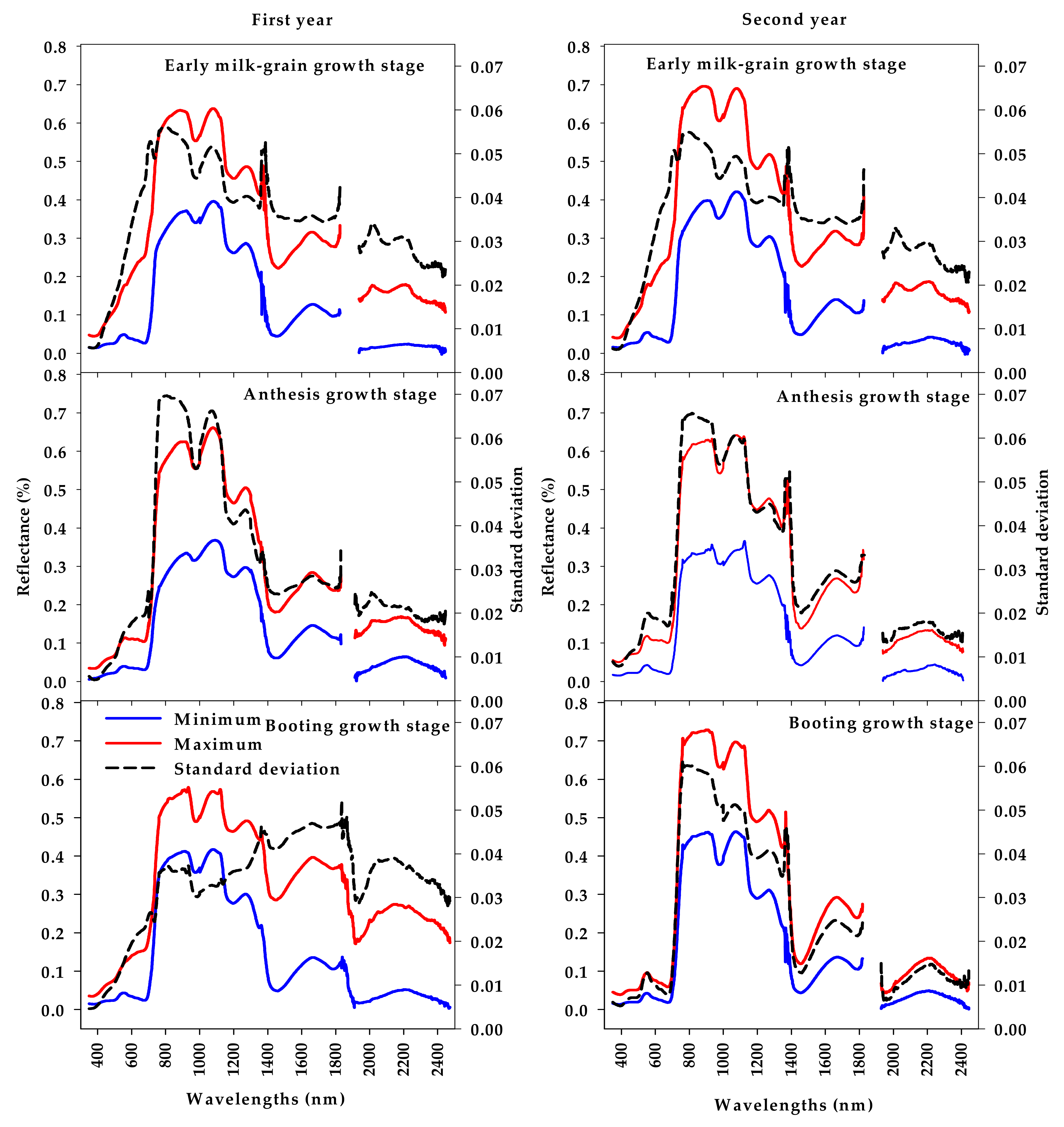
3.2. Analysis of Canopy Spectral Reflectance across Genotypes and Relationship with Biomass and Biological Yield at Different Growth Stages
3.3. Genotypic Variations for Spectral Reflectance Indices and Their Relationship with Biomass and Biological Yield
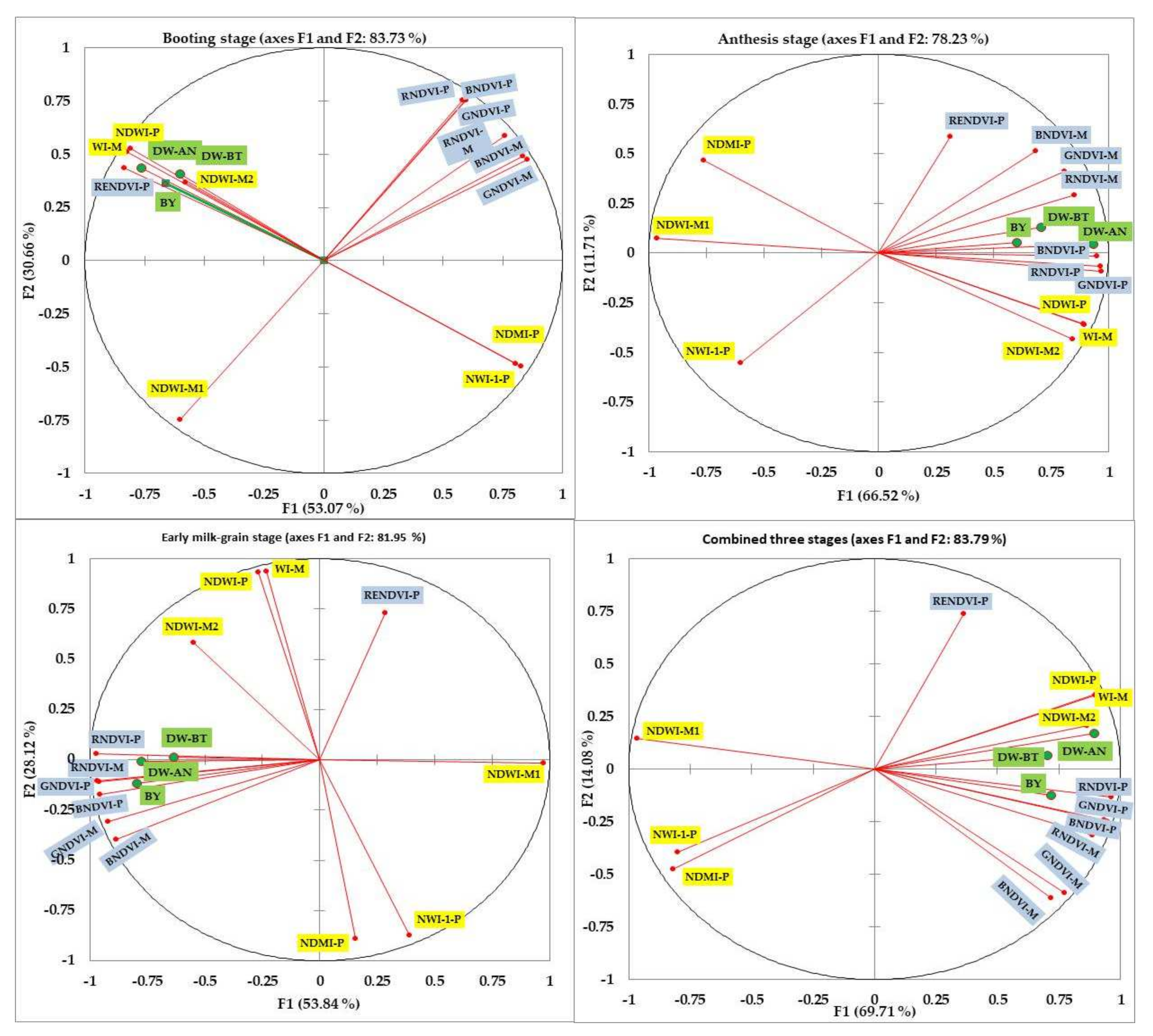
3.4. Principal Component Analysis
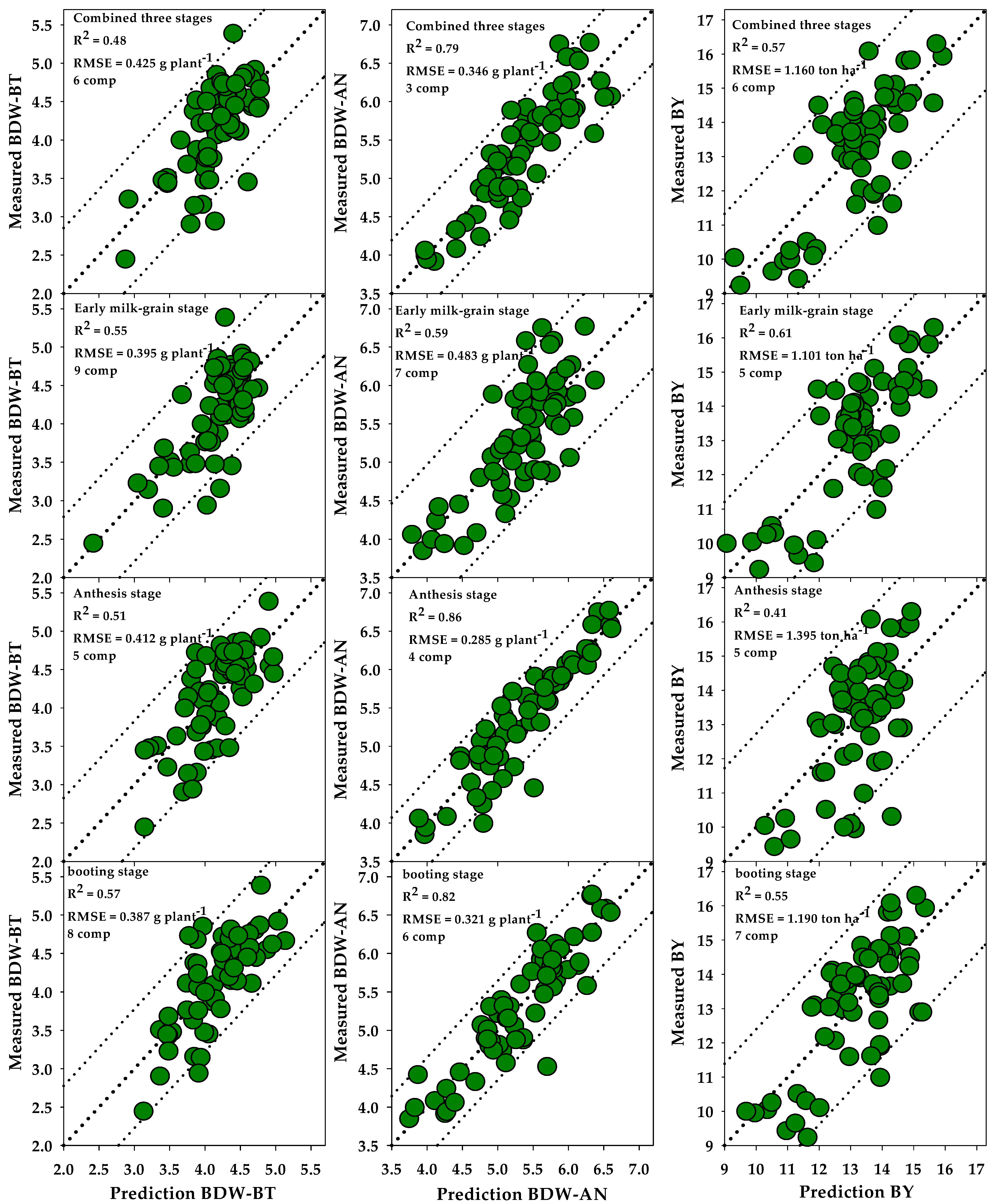
3.5. Prediction of Biomass and Biological Yield Based on All SRIs Using PLSR Models
4. Discussion
4.1. Identification of the Best Spectral Reflectance Indices and Growth Stage for Indirect Biomass Assessment
4.2. Prediction of Plant Biomass and Biological Yield Based on PLSR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Database, Rome. 2021. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 February 2021).
- Curtis, B.C. Wheat in the World. Available online: http://www.fao.org/3/y4011e/y4011e04.htm (accessed on 28 March 2019).
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Rengasamy, P. Soil processes affecting crop production in salt-affected soils. Funct. Plant Biol. 2010, 37, 613–620. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hassan, W.M.; Al-Suhaibani, N.A.; Refay, Y.; Abdella, K.A. Comparative performance of multivariable agro-physiological parameters for detecting salt tolerance of wheat cultivars under simulated saline field growing conditions. Front. Plant. Sci. 2017, 8, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Fan, B.; Guo, L. Soil salinization after long-term mulched drip irrigation poses a potential risk to agricultural sustainability. Eur. J. Soil Sci. 2019, 70, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Sheoran, P.; Kumar, A.; Sharma, R.; Prajapat, K.; Kumar, A.; Barman, A.; Raju, R.; Kumar, S.; Dar, Y.J.; Singh, R.K.; et al. Quantitative dissection of salt tolerance for sustainable wheat production in sodic agro-ecosystems through farmers participatory approach: An Indian experience. Sustainability 2021, 13, 3378. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Azzedine, F.; Gherroucha, H.; Baka, M. Improvement of salt tolerance in durum wheat by ascorbic acid application. J. Stress Physiol. Biochem. 2011, 7, 27–37. [Google Scholar]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants, 1st ed.; Kumar, V., Wani, S.H., Uprasanna, P., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2018; Volume 1, pp. 85–136. [Google Scholar]
- Minhas, P.S.; Bali, A.; Bhardwaj, A.K.; Singh, A.; Yadav, R.K. Structural stability and hydraulic characteristics of soils irrigated for two decades with water having residual alkalinity and its neutralization with gypsum and sulphuric acid. Agric. Water Manag. 2021, 244, 106609. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, M. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Hassan, W.; Dewir, Y.H.; El-Sayed, S.; Al-Ashkar, I.; Abdella, K.A.; Schmidhalter, U. Evaluation of wavelengths and spectral reflectance indices for high throughput assessment of growth, water relations and ion contents of wheat irrigated with saline water. Agric. Water Manag. 2019, 212, 358–377. [Google Scholar] [CrossRef]
- Sheoran, P.; Basak, N.; Kumar, A.; Yadav, R.K.; Singh, R.; Sharma, R.; Kumar, S.; Singh, R.K.; Sharma, P.C. Ameliorants and salt tolerant varieties improve rice-wheat production in soils undergoing sodification with alkali water irrigation in Indo–Gangetic Plains of India. Agric. Water Manag. 2021, 243, 106492. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Z.; Zhang, G.; Meng, Y.; Chen, B.; Wang, Y. Monitoring the leaf water content and specific leaf weight of cotton (Gossypium hirsutum L.) in saline soil using leaf spectral reflectance. Eur. J. Agron. 2012, 41, 103–117. [Google Scholar] [CrossRef]
- Garriga, M.; Retamales, J.B.; Romero, S.; Caligari, P.D.S.; Lobos, G.A. Chlorophyll, anthocyanin, and gas exchange changes assessed by spectroradiometry in Fragaria chiloensis under salt stress. J. Integr. Plant. Biol. 2014, 56, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hackl, H.; Schmidhalter, U. Comparative performance of spectral and thermographic properties of plants and physiological traits for phenotyping salinity tolerance of wheat cultivars under simulated field conditions. Funct. Plant. Biol. 2016, 44, 134. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, A.; Yang, C.; Miller, M.E.; Kianian, S.F.; Marchetto, P.M. A Novel approach to assess salt stress tolerance in wheat using hyperspectral imaging. Front. Plant Sci. 2018, 9, 1182. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Dewir, Y.H.; El-Sayed, S.; Alotaibi, M.; Hassan, W.M.; Refay, Y.; Tahir, M.U. Ability of modified spectral reflectance indices for estimating growth and photosynthetic efficiency of wheat under saline field conditions. Agronomy 2019, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Calzone, A.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E. Hyperspectral detection and monitoring of salt stress in pomegranate cultivars. Agronomy 2021, 11, 1038. [Google Scholar] [CrossRef]
- Aparicio, N.; Villegas, D.; Casadesús, J.; Araus, J.L.; Royo, C. Spectral vegetation indices as nondestructive tools for determining durum wheat yield. Agron. J. 2000, 92, 83–91. [Google Scholar] [CrossRef]
- Pennacchi, J.P.; Carmo-Silva, E.; Andralojc, P.P.J.; Feuerhelm, D.; Powers, S.J.; Parry, M.A.J. Dissecting wheat grain yield drivers in a mapping population in the UK. Agronomy 2018, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Isla, R.; Filella, I.; Araus, J.L. Visible and near-infrared reflectance assessment of salinity effects on barley. Crop Sci. 1997, 37, 198–202. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salt tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Bazihizina, N.; Barrett-Lennard, E.G.; Colmer, T.D. Plant growth and physiology under heterogeneous salinity. Plant Soil. 2012, 354, 1–19. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Cao, F.; Zhang, M.; Chen, X.; Zhang, G.; Wu, F. Difference in yield and physiological features in response to drought and salinity combined stress during anthesis in Tibetan wild and cultivated barleys. PLoS ONE 2013, 8, e77869. [Google Scholar] [CrossRef] [PubMed]
- Oyiga, B.C.; Sharma, R.; Shen, J.; Baum, M.; Ogbonnaya, F.; Léon, J.; Ballvora, A. Identification and characterization of salt tolerance of wheat germplasm using a multivariable screening approach. J. Agron. Crop Sci. 2016, 202, 472–485. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; El-Hendawy, S.E.; Al-Suhaibani, N.; El-Kafafi, S.; Seleiman, M.F. Detecting salt tolerance in doubled haploid wheat Lines. Agronomys 2019, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wan, S.; Zhou, J.; Yang, Z.; Qin, P. Leaf chlorophyll fluorescence, hyperspectral reflectance, pigments content, malondialdehyde and proline accumulation responses of castor bean (Ricinus communis L.) seedlings to salt stress levels. Ind. Crops Prod. 2010, 31, 13–19. [Google Scholar] [CrossRef]
- Hamzeh, S.; Naseri, A.A.; AlaviPanah, S.K.; Mojaradi, B.; Bartholomeus, H.M.; Clevers, J.G.P.W.; Behzad, M. Estimating salinity stress in sugarcane fields with space borne hyperspectral vegetation indices. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 282–290. [Google Scholar] [CrossRef]
- Lara, M.A.; Diezma, B.; Lle’o, L.; Roger, J.M.; Garrido, Y.; Gil, M.I.; Ruiz-Altisent, M. Hyperspectral imaging to evaluate the effect of irrigation water salinity in lettuce. Appl. Sci. 2016, 6, 412. [Google Scholar] [CrossRef]
- Krezhova, D.; Kirova, E.; Yane, T.; Iliev, I. Effects of salinity on leaf spectral reflectance and biochemical parameters of nitrogen fixing soybean plants (Glycine max L.). In Proceedings of the AIP 7th General Conference of the Balkan Physical Union, Alexandroupolis, Greece, 9–13 September 2009; pp. 694–699. [Google Scholar]
- Rud, R.; Shoshany, M.; Alchanatis, V. Spectral indicators for salinity effects in crops: A comparison of a new green indigo ratio with existing indices. Remote Sens. Lett. 2011, 2, 289–298. [Google Scholar] [CrossRef]
- Zhang, T.; Zeng, S.; Gao, Y.; Ouyang, Z.; Li, B.; Fang, C.; Zhao, B. Using hyperspectral vegetation indices as a proxy to monitor soil salinity. Ecol. Indic. 2011, 11, 1552–1562. [Google Scholar] [CrossRef]
- Gutierrez, M.; Reynolds, M.P.; Raun, W.R.; Stone, M.L.; Klatt, A.R. Spectral water indices for assessing yield in elite bread wheat genotypes in well irrigated, water stressed, and high temperature conditions. Crop Sci. 2010, 50, 197–214. [Google Scholar] [CrossRef]
- Ajayi, S.; Reddy, S.K.; Gowda, P.H.; Xue, Q.; Rudd, J.C.; Pradhan, G.; Liu, B.A.; Stewart, C.; Biradar, K.E.; Jessup, K.E. Spectral reflectance models for characterizing winter wheat genotypes. J. Crop Improv. 2016, 30, 176–195. [Google Scholar] [CrossRef]
- Garriga, M.; Romero-Bravo, S.; Estrada, F.; Escobar, A.; Matus, I.A.; del Pozo, A.; Astudillo, C.A.; Lobos, G.A. Assessing wheat traits by spectral reflectance: Do we really need to focus on predicted trait-values or directly identify the elite genotypes group? Front. Plant Sci. 2017, 8, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garriga, M.; Romero-Bravo, S.; Estrada, F.; Méndez-Espinoza, A.M.; González-Martínez, L.; Matus, I.A.; Castillo, D.; Lobos, G.A.; Del Pozo, A. Estimating carbon isotope discrimination and grain yield of bread wheat grown under water-limited and full irrigation conditions by hyperspectral canopy reflectance and multilinear regression analysis. Intern. J. Remote Sens. 2021, 42, 2848–2871. [Google Scholar] [CrossRef]
- Weber, V.S.; Araus, J.L.; Cairns, J.E.; Sanchez, C.; Melchinger, A.E.; Orsini, E. Prediction of grain yield using reflectance spectra of canopy and leaves in maize plants grown under different water regimes. Field Crop. Res. 2012, 128, 82–90. [Google Scholar] [CrossRef]
- Silva-Perez, V.; Molero, G.; Serbin, S.P.; Condon, A.G.; Reynolds, M.P.; Furbank, R.T.; Evans, J.R. Hyperspectral reflectance as a tool to measure biochemical and physiological traits in wheat. J. Exp. Bot. 2018, 69, 483–496. [Google Scholar] [CrossRef] [Green Version]
- Lobos, G.A.; Escobar-Opazo, A.; Estrada, F.; Romero-Bravo, S.; Garriga, M.; del Pozo, A.; Poblete-Echeverría, C.; Gonzalez-Talice, J.; González-Martinez, L.; Caligari, P. Spectral reflectance modeling by wavelength selection: Studying the scope for blueberry physiological breeding under contrasting water supply and heat conditions. Remote Sens. 2019, 11, 329. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Al-Ashkar, I.; Alotaibi, M.; Tahir, M.U.; Solieman, T.; Hassan, W.M. Combining genetic analysis and multivariate modeling to evaluate spectral reflectance indices as indirect selection tools in wheat breeding under water deficit stress conditions. Remote Sens. 2020, 12, 1480. [Google Scholar] [CrossRef]
- Elmetwalli, A.H.; El-Hendawy, S.E.; Al-Suhaibani, N.; Alotaibi, M.; Tahir, M.U.; Mubushar, M.; Hassan, W.M.; El-Sayed, S. Potential of hyperspectral and thermal proximal sensing for estimating growth performance and yield of soybean exposed to different drip irrigation regimes under arid conditions. Sensors 2020, 20, 6569. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Elsayed, S.; Al-Suhaibani, N.; Alotaibi, M.; Tahir, M.U.; Mubushar, M.; Attia, A.; Hassan, W.M. Use of hyperspectral reflectance sensing for assessing growth and chlorophyll content of spring wheat grown under simulated saline field conditions. Plants 2021, 10, 101. [Google Scholar] [CrossRef]
- Yang, H.; Li, F.; Wang, W.; Yu, K. Estimating above-ground biomass of potato using random forest and optimized hyperspectral indices. Remote Sens. 2021, 13, 2339. [Google Scholar] [CrossRef]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Spatial variations in salinity stress across a coastal landscape using vegetation indices derived from hyperspectral imagery. Plant Ecol. 2009, 202, 285–297. [Google Scholar] [CrossRef]
- Song, C.; White, B.L.; Heumann, B.W. Hyperspectral remote sensing of salinity stress on red (Rhizophora mangle) and white (Laguncularia racemosa) mangroves on Galapagos Islands. Remote Sens. Lett. 2011, 2, 221–230. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hassan, W.M.; Refay, Y.; Schmidhalter, U. On the use of spectral reflectance indices to assess agro-morphological traits of wheat plants grown under simulated saline field conditions. J. Agron. Crop Sci. 2017, 203, 406–428. [Google Scholar] [CrossRef]
- Hackl, H.; Mistele, B.; Hu, Y.; Schmidhalter, U. Spectral assessments of wheat plants grown in pots and containers under saline conditions. Funct. Plant Biol. 2013, 40, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Mistele, B.; Hu, Y.; Chen, X.; Schmidhalter, U. Reflectance estimation of canopy nitrogen content in winter wheat using optimized hyperspectral spectral indices and partial least squares regression. Euro. J. Agron. 2014, 52, 198–209. [Google Scholar] [CrossRef]
- Hansen, P.M.; Jorgensen, J.R.; Thomsen, A. Predicting grain yield and protein content in winter wheat and spring barley using repeated canopy reflectance measurements and partial least squares regression. J. Agric. Sci. Camb. 2003, 139, 307–318. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Villegas, D.; Zarco, J.; Aparicio, N.; Araus, J.L.; Royo, C. Assessment of durum wheat yield using visible and near-infrared reflectance spectra of canopies. Field Crops Res. 2005, 94, 126–148. [Google Scholar] [CrossRef]
- Elsayed, S.; Elhoweity, M.; Ibrahim, H.H.; Dewir, Y.H.; Migdadic, H.M.; Schmidhalter, U. Thermal imaging and passive reflectance sensing to estimate the water status and grain yield of wheat under different irrigation regimes. Agric. Water Manag. 2017, 189, 98–110. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, X.; Zhu, X.; Dong, Z.; Guo, W. Estimation of biomass in wheat using random forest regression algorithm and remote sensing data. Crop J. 2016, 4, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.X.; Zhang, L.Y.; Zhang, H.H.; Han, W.T.; Peng, X.S. Estimating above-ground biomass of maize using features derived from UAV-based RGB imagery. Remote Sens. 2019, 11, 1261. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Evaluation of genetic variation for improvement of salt tolerance in spring wheat. In Prospects for Saline Agriculture; Ahmed, R., Malik, K.A., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2002; pp. 131–137. [Google Scholar]
- El-Hendawy, S.E.; Hu, Y.C.; Yakout, G.M.; Awad, A.M.; Hafiz, S.E.; Schmidhalter, U. Evaluating salt tolerance of wheat genotypes using multiple parameters. Euro. J. Agron. 2005, 22, 243–253. [Google Scholar] [CrossRef]
- Mansour, E.; Moustafa, E.S.A.; Desoky, E.M.; Ali, M.M.A.; Yasin, M.A.T.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S.E. Multidimensional evaluation for detecting salt tolerance of bread wheat genotypes under actual saline field growing conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Al-Suhaibani, N.; Selim, M.; Alderfasi, A.; El-Hendawy, S. Comparative performance of integrated nutrient management between composted agricultural wastes, chemical fertilizers, and biofertilizers in improving soil quantitative and qualitative properties and crop yields under arid conditions. Agronomy 2020, 10, 1503. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Christenson, B.S.; Schapaugh, W.T.; Nan, J.R.; Price, K.P.; Prasad, V.; Fritz, A.K. Predicting soybean relative maturity and seed yield using canopy reflectance. Crop Sci. 2016, 56, 625–643. [Google Scholar] [CrossRef] [Green Version]
- Babar, M.A.; Reynolds, M.P.; van Ginkel, M.; Klatt, A.R.; Raun, W.R.; Stone, M.L. Spectral reflectance indices as a potential indirect selection criteria for wheat yield under irrigation. Crop Sci. 2006, 46, 578–588. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Lozano, F.J.; Suárez-Seoane, S.; de Luis, E. Assessment of several spectral indices derived from multi-temporal Landsat data for fire occurrence probability modeling. Remote Sens. Environ. 2007, 107, 533–544. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Avolio, M.L.; Hoffman, A.M.; Smith, M.D. Linking gene regulation, physiology, and plant biomass allocation in Andropogon gerardii in response to drought. Plant Ecol. 2018, 219, 1–15. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abo-Leila, B.H.; Metwally, S.A.; Leithy, S.Z. Anatomical structure of Jatropha leaves affected by proline and salinity conditions. J. Appl. Sci. Res. 2012, 8, 491–496. [Google Scholar]
- Battie-Laclau, P.; Laclau, J.P.; Beri, C.; Mietton, L.; Muniz, M.R.; Arenque, B.C.; DE Cassia Piccolo, M.; Jordan-Meille, L.; Bouillet, J.P.; Nouvellon, Y. Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell. Environ. 2014, 37, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, H.; Chen, T.; Pen, J.; Yu, S.; Zhao, X. Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS ONE 2014, 9, e112807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayat, B.; van der Tol, C.; Verhoef, W. Remote sensing of grass response to drought stress using spectroscopic techniques and canopy reflectance model inversion. Remote Sens. 2016, 8, 557. [Google Scholar] [CrossRef] [Green Version]
- Maimaitiyiming, M.; Ghulam, A.; Bozzolo, A.; Wilkins, J.L.; Kwasniewski, M.T. Early detection of plant physiological responses to different levels of water stress using reflectance spectroscopy. Remote Sens. 2017, 9, 745. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.; Al-Suhaibani, N.; Mubushar, M.; Tahir, M.U.; Refay, Y.; Tola, E. Potential Use of Hyperspectral Reflectance as a High-Throughput Nondestructive Phenotyping Tool for Assessing Salt Tolerance in Advanced Spring Wheat Lines under Field Conditions. Plants 2021, 10, 2512. [Google Scholar] [CrossRef]
- Serbin, S.P.; Dillaway, D.N.; Kruger, E.L.; Townsend, P.A. Leaf optical properties reflect variation in photosynthetic metabolism and its sensitivity to temperature. J. Exp. Bot. 2012, 63, 489–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, E.R.; Doraiswamy, P.C.; McMurtrey, J.E.; Daughtry, C.S.T.; Perry, E.M.; Akhmedov, B. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. Geoinformation 2013, 21, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Barankova, B.; Lazar, D.; Naus, J. Analysis of the effect of chloroplast arrangement on optical properties of green tobacco leaves. Remote Sens. Environ. 2016, 174, 181–196. [Google Scholar] [CrossRef]
- Buitrago, M.F.; Groen, T.A.; Hecker, C.A.; Skidmore, A.K. Spectroscopic determination of leaf traits using infrared spectra. Int. J. Appl. Earth Obs. Geoinf. 2018, 69, 237–250. [Google Scholar] [CrossRef]
- Sun, J.; Yang, L.; Yang, X.; Wei, J.; Li, L.; Guo, E.; Kong, Y. Using spectral reflectance to estimate the leaf chlorophyll content of maize inoculated with arbuscular mycorrhizal fungi under water stress. Front. Plant Sci. 2021, 12, 646173. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Carver, B.F.; Stone, M.L.; Babar, M.A.; Raun, W.R.; Klatt, A.R. Genetic analysis of indirect selection for winter wheat grain yield using spectral reflectance indices. Crop Sci. 2007, 47, 1416–1425. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Alotaibi, M.; Al-Suhaibani, N.; Al-Gaadi, K.; Hassan, W.; Dewir, Y.H.; Emam, M.A.E.-G.; Elsayed, S.; Schmidhalter, U. Comparative performance of spectral reflectance indices and multivariate modeling for assessing agronomic parameters in advanced spring wheat lines under two contrasting irrigation regimes. Front. Plant Sci. 2019, 10, 1537. [Google Scholar] [CrossRef]
- Prasad, B.; Carver, B.F.; Stone, M.L.; Babar, M.A.; Raun, W.R.; Klatt, A.R. Potential use of spectral reflectance indices as a selection tool for grain yield in winter wheat under Great Plains conditions. Crop Sci. 2007, 47, 1426–1440. [Google Scholar] [CrossRef] [Green Version]
- Gizaw, S.A.; Garland-Campbell, K.; Carter, A.H. Evaluation of agronomic traits and spectral reflectance in Pacific Northwest winter wheat under rain-fed and irrigated conditions. Field Crops Res. 2016, 196, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Bowman, B.C.; Chen, J.; Zhang, J.; Wheeler, J.; Wang, Y.; Zhao, W.; Nayak, S.; Heslot, N.; Bockelman, H.; Bonman, J.M. Evaluating grain yield in spring wheat with canopy spectral reflectance. Crop Sci. 2015, 55, 1881–1890. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Y.; Bao, Y.; Luo, J.; Jin, X.; Xu, X.; Song, X.; Yang, G. Exploring the best hyperspectral features for LAI estimation using partial least squares regression. Remote Sens. 2014, 6, 6221–6241. [Google Scholar] [CrossRef] [Green Version]
- Barmeier, G.; Schmidhalter, U. High-throughput field phenotyping of leaves, leaf sheaths, culms and ears of spring barley cultivars at anthesis and dough ripeness. Front. Plant Sci. 2017, 8, 1920. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Feng, H.; Yang, G.; Li, Z. A Comparison of regression techniques for estimation of above-ground winter wheat biomass using near-surface spectroscopy. Remote Sens. 2018, 10, 66. [Google Scholar] [CrossRef] [Green Version]
- Din, M.; Ming, J.; Hussain, S.; Ata-Ul-Karim, S.T.; Rashid, M.; Tahir, M.N.; Hua, S.; Wang, S. Estimation of dynamic canopy variables using hyperspectral derived vegetation indices under varying N rates at diverse phenological stages of rice. Front. Plant Sci. 2019, 9, 1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, S.; El-Hendawy, S.; Dewir, Y.H.; Schmidhalter, U.; Ibrahim, H.H.; Ibrahim, M.M.; Elsherbiny, O.; Farouk, M. Estimating the leaf water status and grain yield of wheat under different irrigation regimes using optimized two- and three-band hyperspectral indices and multivariate regression models. Water 2021, 13, 2666. [Google Scholar] [CrossRef]
- Sun, H.; Feng, M.; Xiao, L.; Yang, W.; Ding, G.; Wang, C.; Jia, X.; Wu, G.; Zhang, S. Potential of multivariate statistical technique based on the effective spectra bands to estimate the plant water content of wheat under different irrigation regimes. Front. Plant Sci. 2021, 12, 631573. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name and Abbreviation of SRIs | Equation | References |
|---|---|---|
| Vegetation-SRIs | ||
| Blue normalized difference vegetation index (BNDVI-P) | (R1245 − R415)/(R1245 + R415) | [62] |
| Blue normalized difference vegetation index (BNDVI-M) | (R1640 − R415)/(R1640 + R415) | This study |
| Green normalized difference vegetation index (GNDVI-P) | (R1245 − R550)/(R1245 + R550) | [62] |
| Green normalized difference vegetation index (GNDVI-M) | (R1640 − R550)/(R1640 + R550) | This study |
| Red normalized difference vegetation index (RNDVI-P) | (R1245 − R680)/(R1245 + R680) | [62] |
| Red normalized difference vegetation index (RNDVI-M) | (R1640 − R680)/(R1640 + R680) | This study |
| Red-edge normalized difference vegetation index (RENDVI-P) | (R1100 − R715)/(R1100 + R715) | [62] |
| Water-SRIs | ||
| Water index (WI-M) | (R860/R1640) | This study |
| Normalized water index -1 (NWI-1-P) | (R970 − R850)/(R970 + R850) | [63] |
| Normalized difference water index (NDWI-P) | (R860 − R1640)/(R860 + R1640) | [64] |
| Normalized difference moisture index (NDMI-P) | (R2200 − R1100)/(R2200 + R1100) | [65] |
| Normalized difference moisture index (NDWI-M1) | (R669 − R1300)/(R669 + R1300) | This study |
| Normalized difference water index (NDWI-M2) | (R737 − R1360)/(R737 + R1360) | This study |
| Effect | Y | GS | GS × Y | G | G × Y | G × GS | G × GS × Y |
|---|---|---|---|---|---|---|---|
| dF | 1 | 2 | 2 | 63 | 63 | 126 | 126 |
| Vegetation-SRIs | |||||||
| BNDVI-P | 0.0114 ** | 0.0817 *** | 0.0731 *** | 0.0120 *** | 0.0032 *** | 0.0066 *** | 0.0034 *** |
| BNDVI-M | 0.1969 ** | 0.0366 *** | 0.2666 *** | 0.0160 *** | 0.0103 *** | 0.0202 *** | 0.0104 *** |
| GNDVI-P | 0.0011 ns | 1.9338 *** | 0.0500 *** | 0.0578 *** | 0.0120 *** | 0.0305 *** | 0.0123 *** |
| GNDVI-M | 0.2608 ** | 2.0079 *** | 0.1929 *** | 0.0530 *** | 0.0294 *** | 0.0596 *** | 0.0293 *** |
| RNDVI-P | 0.8169 ** | 5.6678 *** | 0.2006 *** | 0.0922 *** | 0.0148 *** | 0.0524 *** | 0.0164 *** |
| RNDVI-M | 1.1368 * | 8.8531 *** | 0.3262 *** | 0.1073 *** | 0.0365 *** | 0.1005 *** | 0.0348 *** |
| RENDVI-P | 0.1400 ** | 4.6975 *** | 0.0295 ** | 0.0130 *** | 0.0102 *** | 0.0143 *** | 0.0097 *** |
| Water-SRIs | |||||||
| WI-M | 68.3056 ** | 34.2353 *** | 16.7869 *** | 1.8441 *** | 0.3190 *** | 0.7575 *** | 0.6815 *** |
| NWI-1-P | 0.1229 ** | 0.0291 *** | 0.0390 *** | 0.0016 *** | 0.0007 *** | 0.0007 *** | 0.0008 *** |
| NDWI-P | 1.3212 *** | 0.6691 *** | 0.3359 *** | 0.0471 *** | 0.0103 *** | 0.0183 *** | 0.0172 *** |
| NDMI-P | 0.9310 ** | 0.3763 *** | 0.2314 *** | 0.0295 *** | 0.0119 *** | 0.0143 *** | 0.0141 *** |
| NDWI-M1 | 0.7614 ** | 5.4684 *** | 0.1913 *** | 0.0896 *** | 0.0149 *** | 0.0516 *** | 0.0161 *** |
| NDWI-M2 | 0.7219 ** | 0.1230 *** | 0.4228 *** | 0.0354 *** | 0.0072 *** | 0.0194 *** | 0.0133 *** |
| SRIs | Booting Stage | Anthesis Stage | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | MSE | LSD | Min | Max | Mean | MSE | LSD | |
| BNDVI-P | 0.768 | 0.917 | 0.864 | 0.003 | 0.047 *** | 0.720 | 0.931 | 0.857 | 0.005 | 0.026 *** |
| BNDVI-M | 0.551 | 0.838 | 0.740 | 0.007 | 0.092 *** | 0.601 | 0.832 | 0.740 | 0.005 | 0.059 *** |
| GNDVI-P | 0.547 | 0.810 | 0.705 | 0.006 | 0.084 *** | 0.458 | 0.828 | 0.676 | 0.010 | 0.047 *** |
| GNDVI-M | 0.219 | 0.651 | 0.478 | 0.011 | 0.140 *** | 0.257 | 0.609 | 0.456 | 0.008 | 0.089 *** |
| RNDVI-P | 0.584 | 0.891 | 0.806 | 0.007 | 0.077 *** | 0.551 | 0.890 | 0.745 | 0.011 | 0.051 *** |
| RNDVI-M | 0.254 | 0.797 | 0.641 | 0.012 | 0.130 *** | 0.315 | 0.730 | 0.563 | 0.011 | 0.085 *** |
| RENDVI-P | 0.453 | 0.662 | 0.587 | 0.006 | 0.056 *** | 0.474 | 0.639 | 0.555 | 0.005 | 0.040 *** |
| WI-M | 1.626 | 3.515 | 2.812 | 0.049 | 0.542 *** | 1.480 | 3.940 | 2.644 | 0.070 | 0.590 *** |
| NWI-1-P | −0.086 | −0.022 | −0.063 | 0.002 | 0.015 *** | −0.075 | −0.007 | −0.056 | 0.002 | 0.012 *** |
| NDWI-P | 0.238 | 0.557 | 0.466 | 0.008 | 0.078 *** | 0.193 | 0.595 | 0.436 | 0.011 | 0.071 *** |
| NDMI-P | −0.799 | −0.487 | −0.712 | 0.007 | 0.074 *** | −0.809 | −0.531 | −0.696 | 0.008 | 0.058 *** |
| NDWI-M1 | −0.890 | −0.580 | −0.803 | 0.007 | 0.078 *** | −0.890 | −0.553 | −0.744 | 0.011 | 0.051 *** |
| NDWI-M2 | −0.080 | 0.117 | 0.030 | 0.004 | 0.060 *** | −0.233 | 0.234 | 0.053 | 0.012 | 0.098 *** |
| Early milk-grain stage | Across three stages | |||||||||
| Min | Max | Mean | MSE | LSD | Min | Max | Mean | MSE | LSD | |
| BNDVI-P | 0.701 | 0.919 | 0.834 | 0.005 | 0.025 *** | 0.792 | 0.904 | 0.853 | 0.003 | 0.020 *** |
| BNDVI-M | 0.496 | 0.853 | 0.721 | 0.009 | 0.043 *** | 0.649 | 0.807 | 0.736 | 0.004 | 0.039 *** |
| GNDVI-P | 0.266 | 0.773 | 0.569 | 0.013 | 0.056 *** | 0.512 | 0.768 | 0.649 | 0.007 | 0.037 *** |
| GNDVI-M | −0.051 | 0.623 | 0.343 | 0.016 | 0.077 *** | 0.257 | 0.568 | 0.422 | 0.007 | 0.061 *** |
| RNDVI-P | 0.196 | 0.837 | 0.570 | 0.019 | 0.070 *** | 0.480 | 0.852 | 0.700 | 0.009 | 0.038 *** |
| RNDVI-M | −0.079 | 0.737 | 0.351 | 0.024 | 0.091 *** | 0.289 | 0.712 | 0.499 | 0.011 | 0.060 *** |
| RENDVI-P | 0.275 | 0.527 | 0.379 | 0.007 | 0.036 *** | 0.423 | 0.568 | 0.500 | 0.003 | 0.026 *** |
| WI-M | 1.551 | 2.969 | 2.252 | 0.032 | 0.222 *** | 1.798 | 3.142 | 2.521 | 0.037 | 0.276 *** |
| NWI-1-P | −0.077 | −0.001 | −0.044 | 0.001 | 0.010 *** | −0.074 | −0.028 | −0.055 | 0.001 | 0.007 *** |
| NDWI-P | 0.216 | 0.496 | 0.381 | 0.006 | 0.043 *** | 0.285 | 0.517 | 0.427 | 0.006 | 0.038 *** |
| NDMI-P | −0.766 | −0.511 | −0.650 | 0.006 | 0.510 *** | −0.761 | −0.551 | −0.685 | 0.005 | 0.036 *** |
| NDWI-M1 | −0.838 | −0.205 | −0.572 | 0.019 | 0.069 *** | −0.851 | −0.482 | −0.700 | 0.009 | 0.038 *** |
| NDWI-M2 | −0.050 | 0.176 | 0.061 | 0.005 | 0.040 *** | −0.064 | 0.136 | 0.048 | 0.005 | 0.041 *** |
| SRIs | Booting Stage | Across Stages | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BDW-BT | BDW-AN | BY | ||||||||
| 1st Year | 2nd Year | Com. | 1st Year | 2nd Year | Com. | 1st Year | 2nd Year | Com. | BDW-BT | |
| BNDVI-P | −0.04 | −0.14 | −0.09 | −0.19 | −0.07 | −0.14 | −0.13 | −0.03 | −0.14 | 0.61 |
| BNDVI-M | −0.24 | −0.23 | −0.29 | −0.43 | −0.16 | −0.39 | −0.31 | −0.13 | −0.35 | 0.44 |
| GNDVI-P | −0.03 | −0.13 | −0.10 | −0.16 | −0.09 | −0.14 | −0.10 | −0.06 | −0.14 | 0.61 |
| GNDVI-M | −0.24 | −0.22 | −0.31 | −0.40 | −0.18 | −0.40 | −0.30 | −0.15 | −0.36 | 0.48 |
| RNDVI-P | 0.02 | −0.11 | −0.03 | −0.11 | −0.09 | −0.11 | −0.06 | −0.02 | −0.08 | 0.65 |
| RNDVI-M | −0.11 | −0.18 | −0.18 | −0.26 | −0.17 | −0.30 | −0.17 | −0.10 | −0.24 | 0.60 |
| RENDVI-P | 0.60 | 0.62 | 0.71 | 0.77 | 0.75 | 0.88 | 0.65 | 0.60 | 0.71 | 0.39 |
| WI-M | 0.63 | 0.46 | 0.65 | 0.77 | 0.51 | 0.82 | 0.57 | 0.52 | 0.68 | 0.60 |
| NWI-1-P | −0.64 | −0.45 | −0.64 | −0.74 | −0.53 | −0.79 | −0.56 | −0.51 | −0.66 | −0.57 |
| NDWI-P | 0.61 | 0.42 | 0.62 | 0.75 | 0.48 | 0.80 | 0.57 | 0.52 | 0.68 | 0.60 |
| NDMI-P | −0.63 | −0.47 | −0.66 | −0.75 | −0.66 | −0.83 | −0.60 | −0.54 | −0.68 | −0.52 |
| NDWI-M1 | −0.01 | 0.12 | 0.04 | 0.12 | 0.10 | 0.13 | 0.07 | 0.03 | 0.09 | −0.65 |
| NDWI-M2 | 0.49 | −0.02 | 0.35 | 0.62 | 0.03 | 0.48 | 0.46 | 0.03 | 0.43 | 0.55 |
| Anthesis Stage | BDW-AN | |||||||||
| BNDVI-P | 0.63 | 0.43 | 0.65 | 0.74 | 0.69 | 0.83 | 0.50 | 0.45 | 0.54 | 0.77 |
| BNDVI-M | 0.52 | 0.26 | 0.55 | 0.46 | 0.48 | 0.58 | 0.30 | 0.30 | 0.38 | 0.52 |
| GNDVI-P | 0.67 | 0.46 | 0.67 | 0.83 | 0.71 | 0.88 | 0.53 | 0.46 | 0.55 | 0.80 |
| GNDVI-M | 0.60 | 0.35 | 0.65 | 0.62 | 0.58 | 0.75 | 0.35 | 0.36 | 0.45 | 0.60 |
| RNDVI-P | 0.61 | 0.46 | 0.65 | 0.80 | 0.72 | 0.88 | 0.50 | 0.42 | 0.55 | 0.82 |
| RNDVI-M | 0.57 | 0.41 | 0.64 | 0.65 | 0.69 | 0.80 | 0.38 | 0.35 | 0.46 | 0.74 |
| RENDVI-P | 0.27 | 0.06 | 0.12 | 0.45 | 0.36 | 0.34 | 0.08 | 0.13 | 0.19 | 0.53 |
| WI-M | 0.46 | 0.40 | 0.51 | 0.76 | 0.73 | 0.84 | 0.39 | 0.39 | 0.47 | 0.88 |
| NWI-1-P | −0.45 | −0.13 | −0.34 | −0.66 | −0.43 | −0.60 | −0.23 | −0.15 | −0.29 | −0.78 |
| NDWI-P | 0.47 | 0.41 | 0.51 | 0.78 | 0.70 | 0.83 | 0.41 | 0.41 | 0.49 | 0.86 |
| NDMI-P | −0.40 | −0.30 | −0.40 | −0.66 | −0.53 | −0.68 | −0.32 | −0.22 | −0.34 | −0.76 |
| NDWI-M1 | −0.62 | −0.46 | −0.65 | −0.79 | −0.72 | −0.88 | −0.51 | −0.42 | −0.55 | −0.81 |
| NDWI-M2 | 0.42 | 0.38 | 0.50 | 0.68 | 0.62 | 0.76 | 0.34 | 0.39 | 0.42 | 0.80 |
| Early Milk-Grain Stage | BY | |||||||||
| BNDVI-P | 0.36 | 0.54 | 0.54 | 0.54 | 0.66 | 0.70 | 0.61 | 0.71 | 0.74 | 0.65 |
| BNDVI-M | 0.26 | 0.51 | 0.48 | 0.43 | 0.63 | 0.64 | 0.54 | 0.68 | 0.72 | 0.52 |
| GNDVI-P | 0.34 | 0.53 | 0.52 | 0.53 | 0.62 | 0.68 | 0.63 | 0.69 | 0.72 | 0.66 |
| GNDVI-M | 0.24 | 0.51 | 0.47 | 0.43 | 0.61 | 0.64 | 0.56 | 0.67 | 0.71 | 0.56 |
| RNDVI-P | 0.41 | 0.56 | 0.57 | 0.56 | 0.67 | 0.71 | 0.65 | 0.73 | 0.73 | 0.71 |
| RNDVI-M | 0.34 | 0.56 | 0.54 | 0.51 | 0.68 | 0.69 | 0.63 | 0.73 | 0.74 | 0.68 |
| RENDVI-P | 0.09 | −0.24 | −0.09 | −0.04 | −0.23 | −0.16 | −0.17 | −0.30 | −0.31 | 0.26 |
| WI-M | 0.19 | −0.10 | 0.12 | 0.23 | −0.13 | 0.16 | 0.08 | −0.08 | 0.02 | 0.55 |
| NWI-1-P | −0.20 | 0.04 | −0.23 | −0.22 | 0.06 | −0.25 | −0.26 | −0.06 | −0.23 | −0.55 |
| NDWI-P | 0.23 | −0.08 | 0.18 | 0.25 | −0.10 | 0.20 | 0.14 | −0.03 | 0.08 | 0.58 |
| NDMI-P | −0.10 | 0.14 | −0.02 | −0.12 | 0.14 | −0.07 | −0.09 | 0.09 | −0.02 | −0.50 |
| NDWI-M1 | −0.41 | −0.56 | −0.57 | −0.56 | −0.67 | −0.71 | −0.65 | −0.73 | −0.74 | −0.71 |
| NDWI-M2 | 0.19 | 0.06 | 0.31 | 0.31 | 0.07 | 0.38 | 0.36 | 0.21 | 0.38 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hendawy, S.; Al-Suhaibani, N.; Mubushar, M.; Tahir, M.U.; Marey, S.; Refay, Y.; Tola, E. Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions. Appl. Sci. 2022, 12, 1983. https://doi.org/10.3390/app12041983
El-Hendawy S, Al-Suhaibani N, Mubushar M, Tahir MU, Marey S, Refay Y, Tola E. Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions. Applied Sciences. 2022; 12(4):1983. https://doi.org/10.3390/app12041983
Chicago/Turabian StyleEl-Hendawy, Salah, Nasser Al-Suhaibani, Muhammad Mubushar, Muhammad Usman Tahir, Samy Marey, Yahya Refay, and ElKamil Tola. 2022. "Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions" Applied Sciences 12, no. 4: 1983. https://doi.org/10.3390/app12041983
APA StyleEl-Hendawy, S., Al-Suhaibani, N., Mubushar, M., Tahir, M. U., Marey, S., Refay, Y., & Tola, E. (2022). Combining Hyperspectral Reflectance and Multivariate Regression Models to Estimate Plant Biomass of Advanced Spring Wheat Lines in Diverse Phenological Stages under Salinity Conditions. Applied Sciences, 12(4), 1983. https://doi.org/10.3390/app12041983