
The Hormetic Effects of a Brassica Water Extract Triggered Wheat Growth and Antioxidative Defense under Drought Stress

 , ,
, ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Preparation of the Brassica Water Extract
2.3. Experimental Design and Treatments
2.4. Crop Husbandry
2.5. Data Recorded
2.5.1. Morphological Growth
2.5.2. Photosynthetic Pigments
2.5.3. Physiological Attributes
2.5.4. Enzymatic Antioxidants and H2O2 Content
2.6. Statistical Analysis
3. Results
3.1. Morphological Growth
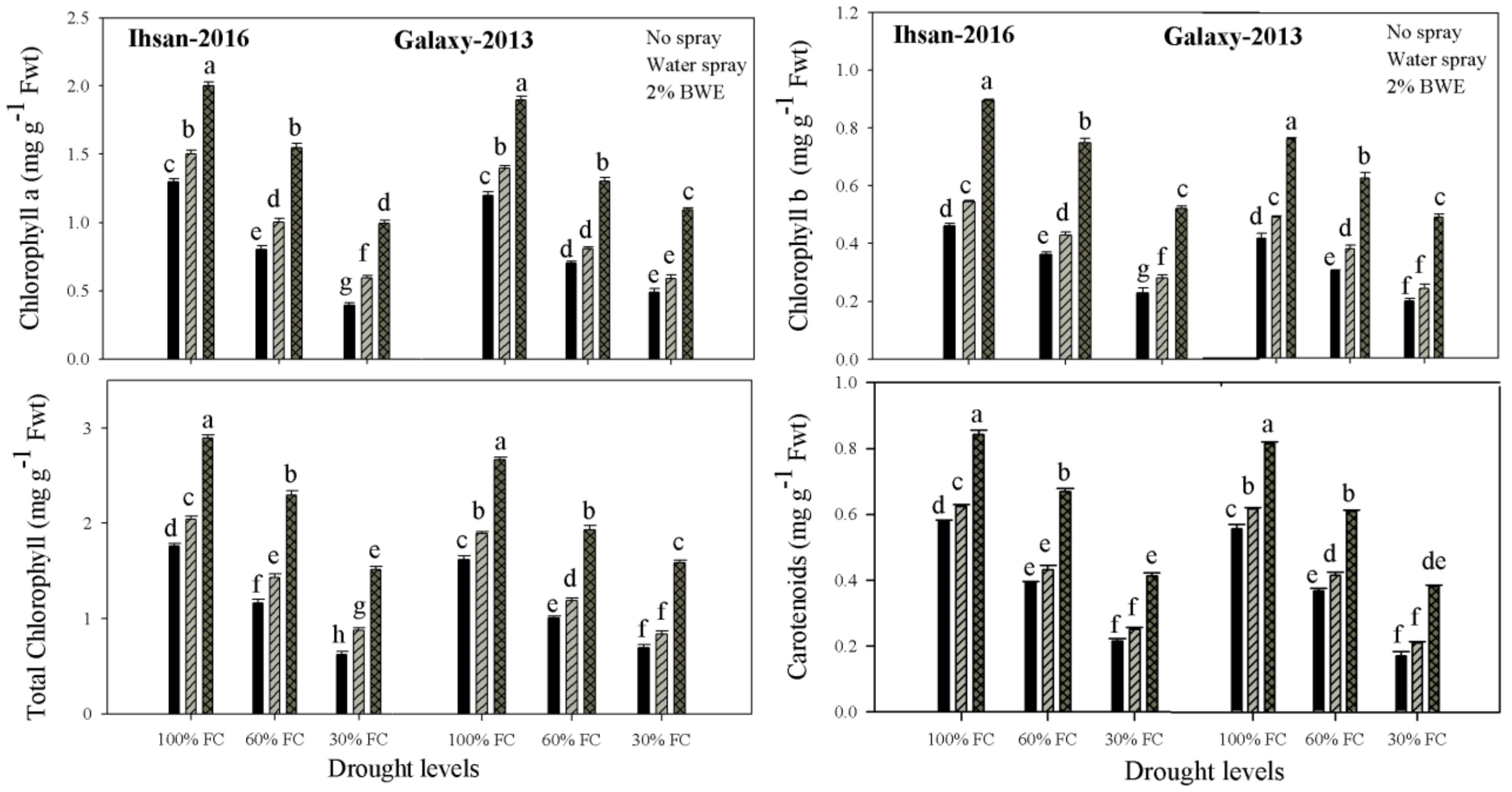
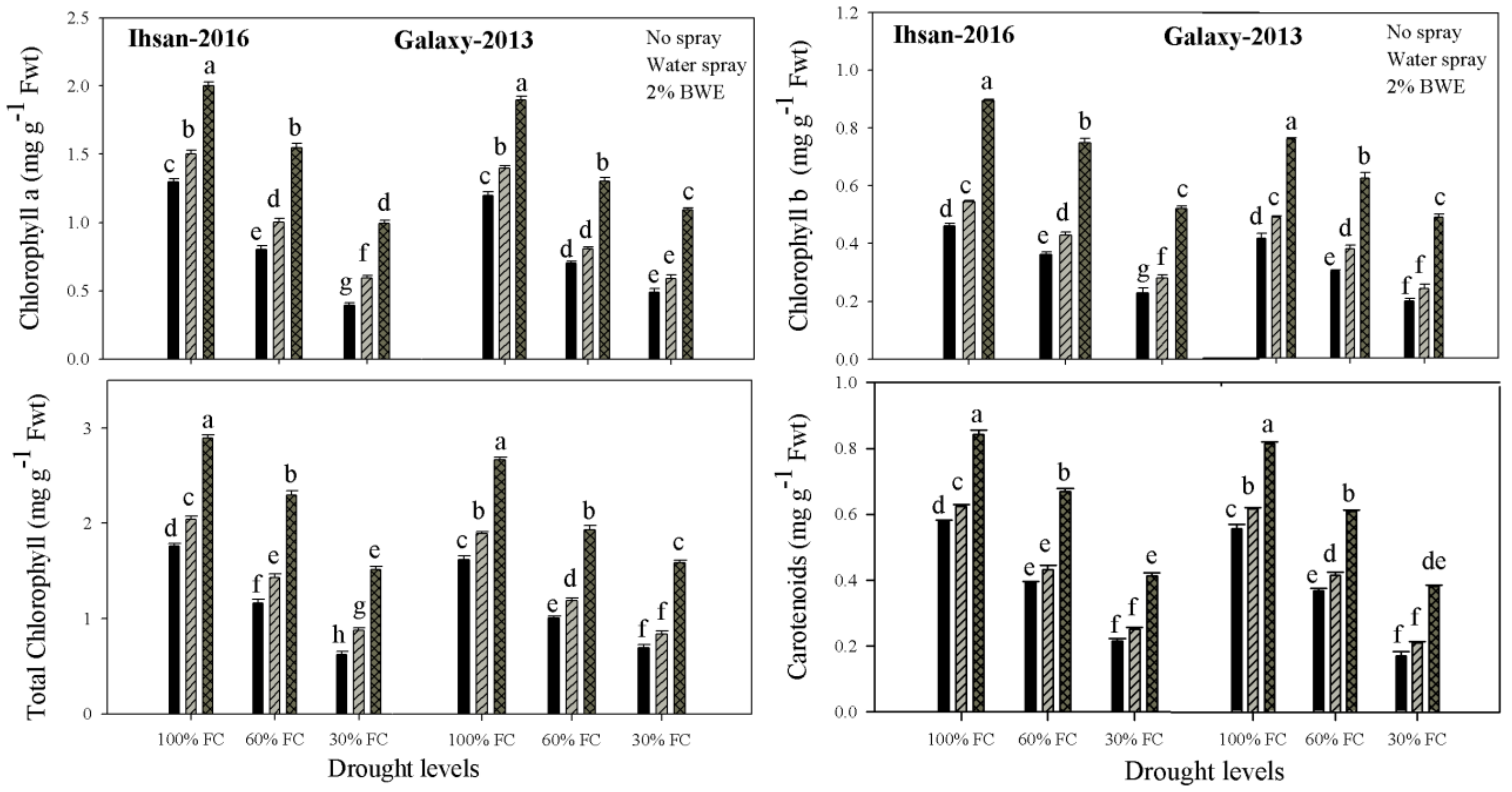
3.2. Photosynthetic Pigments
3.3. Physiological Attributes
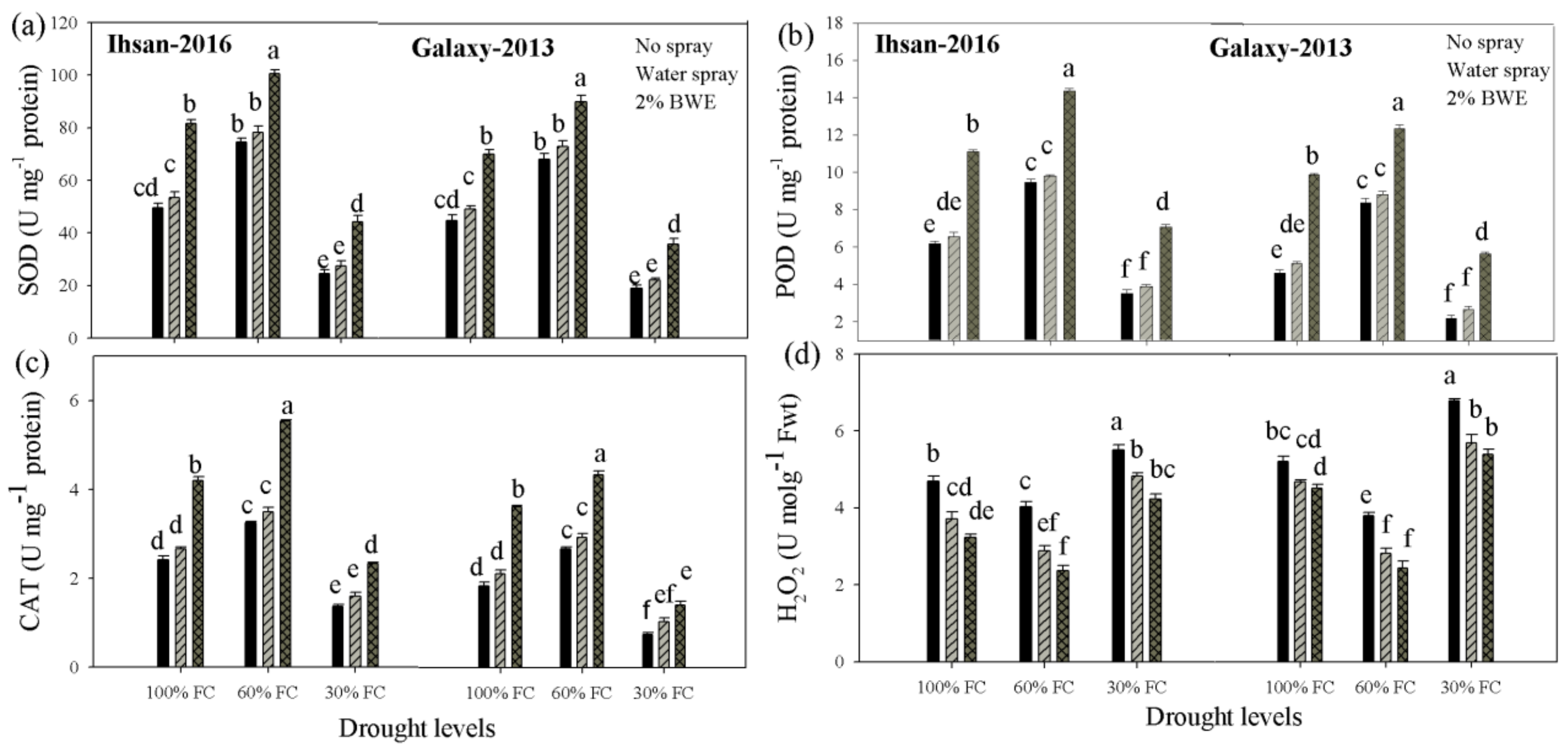
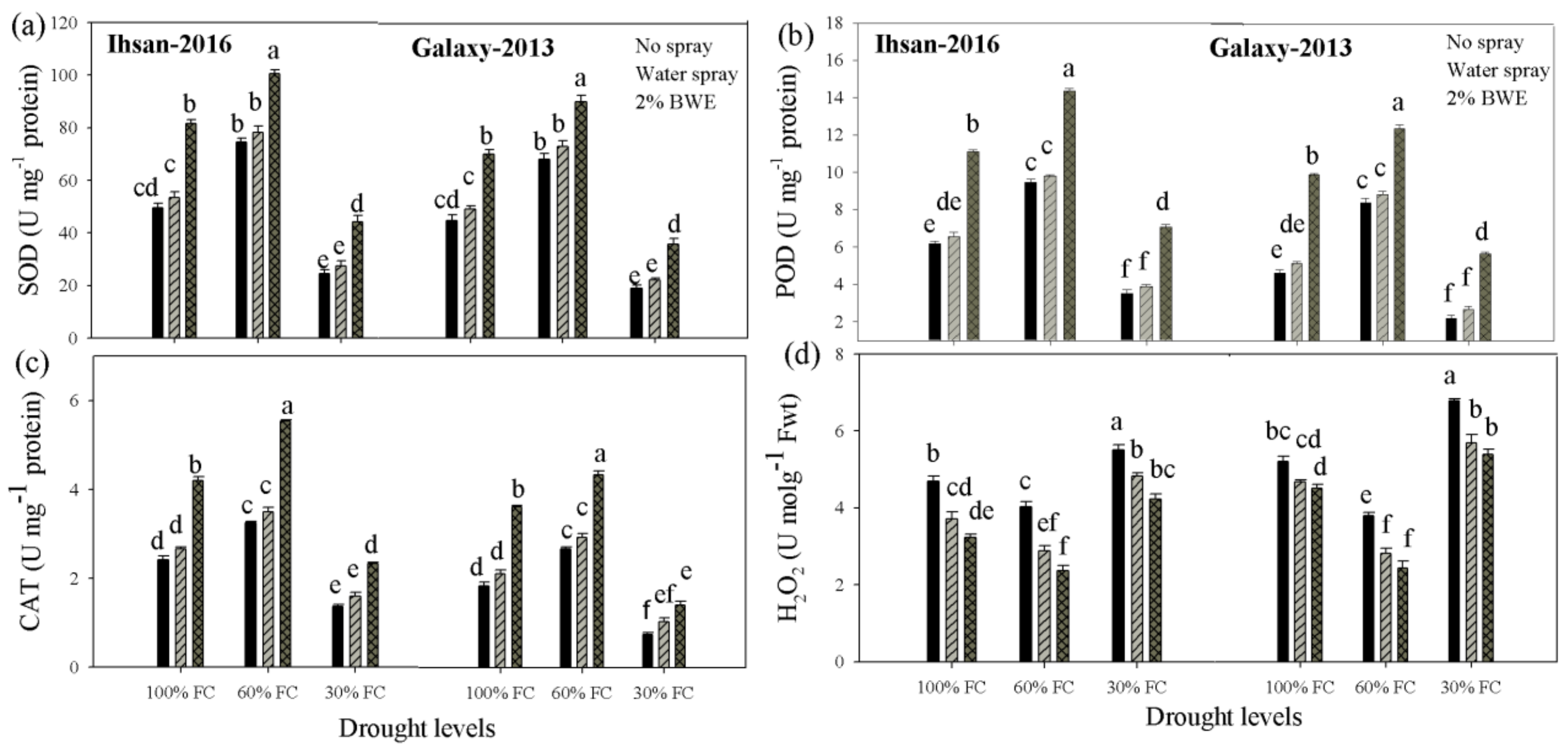
3.4. Antioxidant Enzymes
3.5. Hydrogen Peroxide (H2O2)
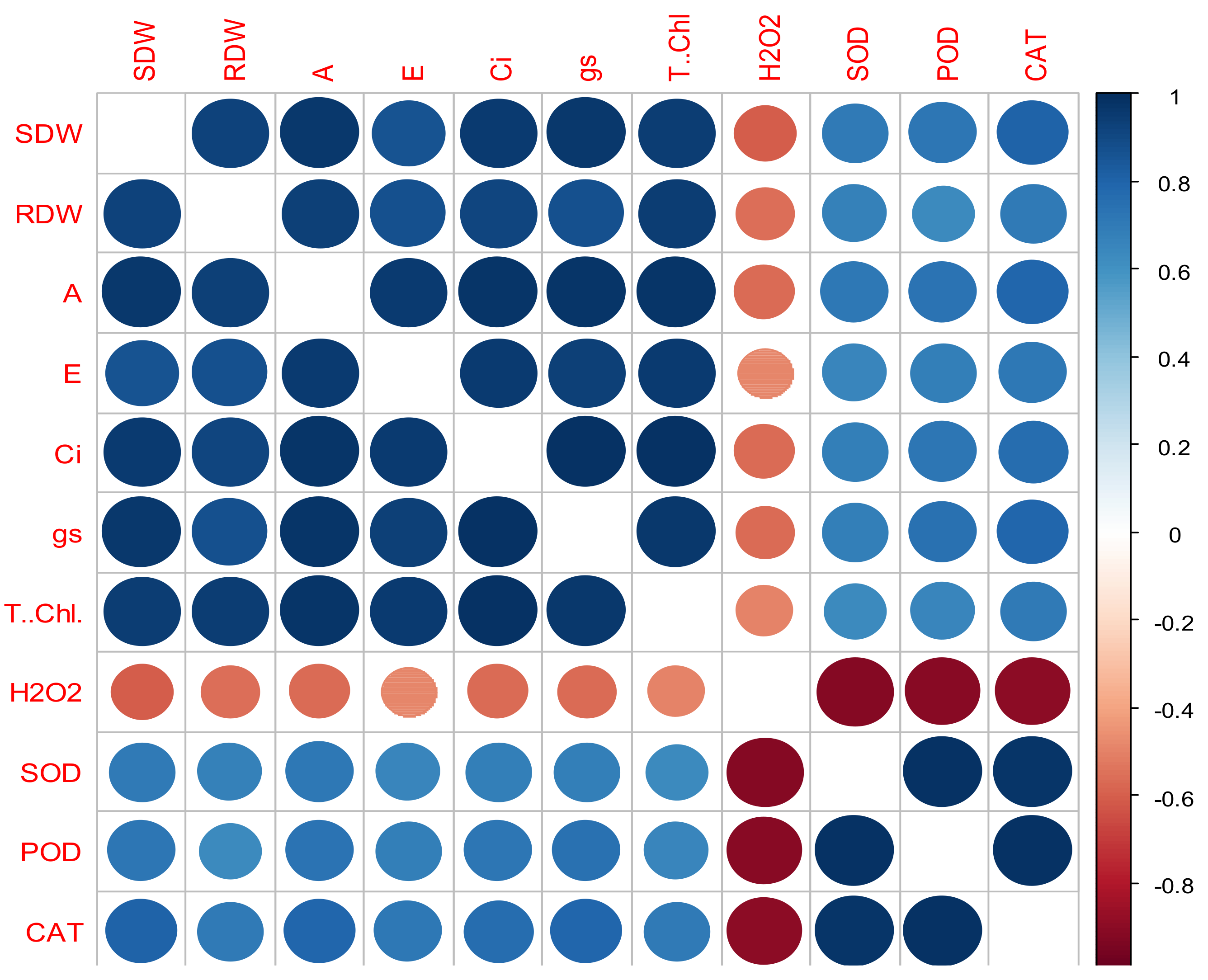
3.6. Correlation Analysis
4. Discussion
4.1. Drought Stress and Plant Performance
4.2. Foliar Application of the Brassica Water Extracts Lessened the Detrimental Effects of Drought
4.3. Drought Tolerance through Enhanced Physiological Attributes under the Foliar Application of Brassica Water Extracts
4.4. Brassica Water Extracts Strengthened the Antioxidant Defense System in Wheat under Drought Stress
4.5. Performance of Wheat Cultivars in Response to Drought and Brassica Water Extract Application
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, H.G.M.D.; Zeng, Y.; Yang, X.; Anwaar, H.A.; Mansha, M.Z.; Hanif, C.M.S.; Alghanem, S.M.S. Conferring drought-tolerant wheat genotypes through morpho-physiological and chlorophyll indices at seedling stage. Saudi J. Biol. Sci. 2020, 27, 2116–2123. [Google Scholar] [CrossRef]
- Kheir, A.M.; Alrajhi, A.A.; Ghoneim, A.M.; Ali, E.F.; Magrashi, A.; Zoghdan, M.G.; Abdelkhalik, S.A.; Fahmy, A.E.; Elnashar, A. Modeling deficit irrigation-based evapotranspiration optimizes wheat yield and water productivity in arid regions. Agric. Water Manag. 2021, 256, 107122. [Google Scholar] [CrossRef]
- Liu, H.; Sultan, M.A.R.F.; Liu, X.L.; Zhang, J.; Yu, F.; Zhao, H.X. Physiological and comparative proteomic analysis reveals different drought responses in roots and leaves of drought-tolerant wild wheat (Triticum boeoticum L.). PLoS ONE 2015, 10, 121–852. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhang, D.; Li, X.; Shi, Y.; Shao, Y.; Fang, B.; Cheng, H. Drought effects on photosynthetic performance of two wheat cultivars contrasting in drought. N. Z. J. Crop Hortic. Sci. 2021, 49, 17–29. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Jin, J. Effect of drought on agronomic traits of rice and wheat: A meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef] [Green Version]
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navab, A. A physio-morphological trait-based approach for breeding drought tolerant wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef]
- Rani, A.; Devi, P.; Jha, U.C.; Sharma, K.D.; Siddique, K.H.M.; Nayyar, H. Developing Climate-Resilient Chickpea Involving Physiological and Molecular Approaches with a Focus on Temperature and Drought Stresses. Front. Plant Sci. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Talbi, S.; Romero-Puertas, M.C.; Hernández, A.; Terrón, L.; Ferchichi, A.; Sandalio, L.M. Drought tolerance in a Saharian plant Oudneya africana: Role of antioxidant defences. Environ. Exp. Bot. 2015, 111, 114–126. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Farooq, M.; Nawaz, A. Seed priming with sorghum extracts and benzyl aminopurine improves the tolerance against salt stress in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2018, 24, 239–249. [Google Scholar] [CrossRef]
- Moghaddam, N.S.A.; Oskouie, M.N.; Butler, A.E.; Petit, P.X.; Barreto, G.E.; Sahebkar, A. Hormetic effects of curcumin: What is the evidence? J. Cell Physiol. 2019, 234, 10060–10071. [Google Scholar] [CrossRef]
- Rehman, S.; Shahzad, B.; Bajwa, A.A.; Hussain, S.; Rehman, A.; Cheema, S.A.; Li, P. Utilizing the allelopathic potential of Brassica species for sustainable crop production: A review. J. Plant Growth Regul. 2019, 38, 343–356. [Google Scholar] [CrossRef]
- Hussain, M.A.; Fahad, S.; Sharif, R.; Jan, M.F.; Mujtaba, M.; Ali, Q.; Ahmad, A.; Ahmad, H.; Amin, N.; Ajayo, B.S.; et al. Multifunctional role of brassinosteroid and its analogues in plants. Plant Growth Regul. 2020, 92, 141–156. [Google Scholar] [CrossRef]
- Latha, P.; Vardhini, B.V. Effect of homobrassinolide on bio-chemical activities and chlorophyll pigments of mustard plants grown in semi-arid tropics of Nizamabad. Eur. J. Biomed. Pharm. Sci. 2017, 4, 613–618. [Google Scholar]
- Kahlaoui, B.; Misle, E.; Khaskhoussy, K.; Jaouadi, I.; Hachicha, M. Brassinosteroids and drought tolerance in plants. Water Stress Crop Plants Sustain. Approach 2016, 2, 600–607. [Google Scholar]
- Sharma, A.; Yuan, H.; Kumar, V.; Ramakrishnan, M.; Kohli, S.K.; Kaur, R.; Zheng, B. Castasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Wang, Y.T.; Chen, Z.Y.; Jiang, Y.; Duan, B.B.; Xi, Z.M. Involvement of ABA and antioxidant system in brassinosteroid-induced water stress tolerance of grapevine (Vitis vinifera L.). Sci. Hortic. 2019, 256, 108596. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Brassinosteroids as a multidimensional regulator of plant physiological and molecular responses under various environmental stresses. Environ. Sci. Pollut. Res. 2021, 28, 44768–44779. [Google Scholar] [CrossRef]
- Ribeiro, D.S.D.G.; da Silva, B.R.S.; da Silva, L.A.K. Brassinosteroids induce tolerance to water deficit in soybean seedlings: Contributions linked to root anatomy and antioxidant enzymes. Acta Physiol. Plant. 2019, 41, 82. [Google Scholar] [CrossRef]
- Dehghan, M.; Balouchi, H.; Yadavi, A.; Zare, E. Improve wheat (Triticum aestivum L.) performance by brassinolide application under different irrigation regimes. S. Afr. J. Bot. 2020, 130, 259–267. [Google Scholar] [CrossRef]
- Talaat, N.B. 24-Epibrassinolide and Spermine combined treatment sustains maize (Zea mays L.) drought tolerance by improving photosynthetic efficiency and altering phytohormones profile. J. Soil Sci. Plant Nutr. 2020, 20, 516–529. [Google Scholar] [CrossRef]
- Zafari, M.; Ebadi, A.; Sedghi, M.; Jahanbakhsh, S.; Miransari, M.J. Alleviating effect of 24-epibrassinolide on seed oil content and fatty acid composition under drought stress in safflower. J. Food Compos. Anal. 2020, 92, 103544. [Google Scholar] [CrossRef]
- Farooq, O.; Ali, M.; Sarwar, N.; Rehman, I.M.M.; Naz, T.; Asghar, M.; Ehsan, F.; Nasir, M.; Hussain, Q.M.; Afzal, S. Foliar applied brassica water extract improves the seedling development of wheat and chickpea. Asian J. Agric. Biol. 2021, 1. [Google Scholar] [CrossRef]
- Cheema, Z.A.; Khaliq, A. Use of sorghum allelopathic properties to control weeds in irrigated wheat in semi-arid region of Punjab. Agric. Ecosyst. Environ. 2000, 79, 105–112. [Google Scholar] [CrossRef]
- Arnon, D.T. Copper enzyme in isolated chloroplasts polyphenols oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dhindsa, R.A.; Plumb-Dhindsa, P.; Thrope, T.A. Leaf senescence Correlated with increased permeability and lipid peroxidation and decreased levels of superoxide dimutase and catalase. J. Exp. Bot. 1981, 126, 93–101. [Google Scholar] [CrossRef]
- Putter, J. Peroxidases. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie, Weinheim. Rade and Markets Division (EST); Elsevier: Rome, Italy, 1974; pp. 685–690. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw Hill Book Co. Inc.: New York, NY, USA, 1997; pp. 400–428. [Google Scholar]
- Camaille, M.; Fabre, N.; Clément, C.; Ait Barka, E. Advances in wheat physiology in response to drought and the role of plant growth promoting rhizobacteria to trigger drought tolerance. Microorganisms 2021, 9, 687. [Google Scholar] [CrossRef]
- He, J.X.; Wang, J.; Liang, H.G. Effects of water stress on photochemical function and protein metabolism of photosystem II in wheat leaves. Physiol. Plant. 2010, 93, 771–777. [Google Scholar] [CrossRef]
- Asghar, H.N.; Zahir, Z.A.; Akram, M.A.; Ahmad, H.T.; Hussain, M.B. Isolation and screening of beneficial bacteria to ameliorate drought stress in wheat. Soil Environ. 2015, 34, 100–110. [Google Scholar]
- Miao, Y.; Zhu, Z.; Guo, Q.; Ma, H.; Zhu, L. Alternate wetting and drying irrigation-mediated changes in the growth, photosynthesis and yield of the medicinal plant Tulipa edulis. Ind. Crop. Prod. 2015, 66, 81–88. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.H.P.J.C.; Harris, P.J. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Jourdan, N.F.; Martino, C.; El-Esawi, M.; Witczak, J.; Bouchet, P.E.; d’Harlingu, A.; Ahmad, M. Blue-light dependent ROS formation by Arabidopsis cryptochrome-2 may contribute toward its signaling role. Plant Signal. Behav. 2015, 10, 1042647. [Google Scholar] [CrossRef] [Green Version]
- Hussain, I.; Ashraf, M.A.; Anwar, F.; Rasheed, R.; Niaz, M.; Wahid, A. Biochemical characterization of maize (Zea mays L.) for salt tolerance. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2014, 148, 1016–1026. [Google Scholar]
- Ahammed, G.J.; Li, X.; Liu, A.; Chen, S. Brassinosteroids in plant tolerance to abiotic stress. J. Plant Growth Regul. 2020, 39, 1451–1464. [Google Scholar] [CrossRef]
- Nie, S.; Huang, S.; Wang, S.; Mao, Y.; Liu, J.; Ma, R.; Wang, X. Enhanced brassinosteroid signaling intensity via SlBRI1 overexpression negatively regulates drought resistance in a manner opposite of that via exogenous BR application in tomato. Plant Physiol. Biochem. 2019, 138, 36–47. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [Green Version]
- Peres, A.L.G.; Soares, J.S.; Tavares, R.G.; Righetto, G.; Zullo, M.A.; Mandava, N.B.; Menossi, M. Brassinosteroids, the sixth class of phytohormones: A molecular view from the discovery to hormonal interactions in plant development and stress adaptation. Int. J. Mol. Sci. 2019, 20, 331. [Google Scholar] [CrossRef] [Green Version]
- Maghsoudi, K.; Emam, Y.; Ashraf, M. Influence of foliar application of silicon on chlorophyll fluorescence, photosynthetic pigments, and growth in water-stressed wheat cultivars differing in drought tolerance. Turk. J. Bot. 2015, 39, 625–634. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef]
- Liu, S.; Che, Z.; Chen, G. Multiple-fungicide resistance to carbendazim, diethofencarb, procymidone, and pyrimethanil in field isolates of Botrytis cinerea from tomato in Henan Province, China. Crop Prot. 2016, 84, 56–61. [Google Scholar] [CrossRef]
- Da Fonseca, S.S.; da Silva, B.R.S.; Lobato, A.K.D.S. 24-Epibrassinolide positively modulate leaf structures, antioxidant system and photosynthetic machinery in rice under simulated acid rain. J. Plant Growth Regul. 2020, 39, 1559–1576. [Google Scholar] [CrossRef]
- Rajewska, I.; Talarek, M.; Bajguz, A. Brassinosteroids and response of plants to heavy metals action. Front. Plant Sci. 2016, 7, 629. [Google Scholar] [CrossRef] [Green Version]
- Talaat, N.B.; Shawky, B.T.; Ibrahim, A.S. Alleviation of drought-induced oxidative stress in maize (Zea mays L.) plants by dual application of 24-epibrassinolide and spermine. Environ. Exp. Bot. 2015, 113, 47–58. [Google Scholar] [CrossRef]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Urban, L.; Aarrouf, J.; Bidel, L.P. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Seed priming with brassinosteroids alleviates aluminum toxicity in rice via improving antioxidant defense system and suppressing aluminum uptake. Environ. Sci. Pollut. Res. 2022, 29, 10183–10197. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef]
- De Assis-Gomes, M.D.M.; Pinheiro, D.T.; Bressan-Smith, R.; Campostrini, E. Exogenous brassinosteroid application delays senescence and promotes hyponasty in Carica papaya L. leaves. Theor. Exp. Plant Physiol. 2018, 30, 193–201. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, X.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Enhanced photosynthetic capacity and antioxidant potential mediate brassinosteriod-induced phenanthrene stress tolerance in tomato. Environ. Pollut. 2015, 201, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Paciolla, C.; Paradiso, A.; De Pinto, M.C. Cellular redox homeostasis as central modulator in plant stress response. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Springer: Cham, Switzerland, 2016; pp. 1–23. [Google Scholar]
- Al Mahmud, J.; Bhuyan, M.B.; Anee, T.I.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Reactive oxygen species metabolism and antioxidant defense in plants under metal/metalloid stress. In Plant Abiotic Stress Tolerance; Springer: Cham, Switzerland, 2019; pp. 221–257. [Google Scholar]
- Kaya, C.; Ashraf, M.; Wijaya, L.; Ahmad, P. The putative role of endogenous nitric oxide in brassinosteroid-induced antioxidant defense system in pepper (Capsicum annuum L.) plants under water stress. Plant Physiol. Biochem. 2019, 143, 119–128. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Traits | Source of Variation | |||||
|---|---|---|---|---|---|---|
| Drought Stress (DS) | Brassica Water Extract (BWE) | DS × BWE | ||||
| Ihsan-2016 | Galaxy-2013 | Ihsan-2016 | Galaxy-2013 | Ihsan-2016 | Galaxy-2013 | |
| Shoot length (cm) | 1228.73 ** | 1206.12 ** | 69.26 ** | 68.70 ** | 0.21 ns | 0.98 ns |
| Root length (cm) | 813.37 ** | 848.62 ** | 88.41 ** | 85.14 ** | 0.66 ns | 0.33 ns |
| Shoot fresh weight (g) | 402.29 ** | 324.48 ** | 10.32 ** | 14.39 ** | 1.07 ns | 0.54 ns |
| Root fresh weight (g) | 120.47 ** | 97.66 ** | 13.81 ** | 4.40 ** | 0.18 ** | 0.03 ns |
| Shoot dry weight (g) | 10.63 ** | 10.55 ** | 1.09 ** | 0.87 ** | 0.06 ** | 0.03 ** |
| Root dry weight (g) | 8.55 ** | 7.87 ** | 0.37 ** | 0.19 ** | 0.003 ns | 0.002 ns |
| Chlorophyll a (mg g−1 FW) | 1.98 ** | 1.43 ** | 1.10 ** | 1.008 ** | 0.005 * | 0.003 ns |
| Chlorophyll b (mg g−1 FW) | 0.019 ** | 0.13 ** | 0.35 ** | 0.25 ** | 0.004 ns | 0.0006 ns |
| Total chlorophyll (mg g−1 FW) | 3.40 ** | 2.42 ** | 2.69 ** | 2.28 ** | 0.01 ** | 0.003 ns |
| Carotenoids (mg g−1 FW) | 0.34 ** | 0.38 ** | 0.15 ** | 0.14 ** | 0.001 ** | 0.0004 ns |
| Photosynthesis rate (µmol Co2 m−2 s−1) | 51.42 ** | 48.64 ** | 5.76 ** | 5.12 ** | 0.55 ** | 0.50 ** |
| Transpiration rate (mmol H2O m−2 s−1) | 28.06 ** | 29.96 ** | 4.28 ** | 5.92 ** | 0.16 ** | 0.21 ** |
| Internal carbon dioxide (µmol CO2 mol air−1) | 31,573.6 ** | 34,934.4 ** | 7054.5 ** | 6248.2 ** | 62.2 ns | 59.4 ns |
| Stomatal conductance (mol H2O m−2 s−1) | 0.09 ** | 0.09 ** | 0.02 ** | 0.02 ** | 0.0008 ** | 0.001 ** |
| Superoxidase dismutase (U mg−1 protein) | 6205.92 ** | 5911.06 ** | 1785.36 ** | 1143.50 ** | 36.19 * | 16.40 ns |
| Peroxidase (U mg−1 protein) | 91.60 ** | 92.22 ** | 55.04 ** | 48.90 ** | 0.62 ** | 0.83 ** |
| Catalase (U mg−1 protein) | 12.28 ** | 11.69 ** | 7.42 ** | 4.73 ** | 0.43 ** | 0.39 ** |
| Hydrogen peroxide (umol g−1 FW) | 6.97 ** | 20.02 ** | 4.97 ** | 3.27 ** | 0.04 ns | 0.13 ns |
| Ihsan-2016 | Galaxy-2013 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Length (cm) | Root Length (cm) | Shoot Fresh Weight (g) | Root Fresh Weight (g) | Shoot Dry Weight (g) | Root Dry Weight (g) | Shoot Length (cm) | Root Length (cm) | Shoot Fresh Weight (g) | Root Fresh Weight (g) | Shoot Dry Weight (g) | Root Dry Weight (g) | ||
| 100% FC | Control | 53.41 a–e | 22.15 g–i | 15.82 c–f | 5.93 g | 2.56 g–i | 1.79 c–f | 54.41 a–h | 21.15 ef | 14.80 a–e | 5.28 de | 2.18 gh | 1.69 d |
| Water spray | 54.41 a–d | 23.15 f–h | 16.50 b–f | 6.46 fg | 2.68 fg | 1.99 b–e | 56.41 a–f | 22.15 de | 15.36 a–c | 5.77 cd | 2.32 f–h | 1.79 cd | |
| 0.5% BWE | 56.41 a–c | 25.15 d–g | 17.43 a–e | 7.24 de | 2.93 d–g | 2.17 a–c | 58.41 a–d | 24.15 cd | 16.10 ab | 6.37 bc | 2.52 d–g | 1.89 bc | |
| 1.0% BWE | 58.41 ab | 27.15 b–e | 18.20 a–d | 8.07 c | 3.28 b–d | 2.29 ab | 59.41 a–d | 26.15 c | 17.00 a | 6.88 ab | 2.72 b–e | 1.97 b | |
| 1.5% BWE | 59.41 ab | 29.15 a–c | 19.27 a–c | 8.87 ab | 3.39 a–c | 2.39 a | 61.41 a–c | 28.15 ab | 17.53 a | 7.07 ab | 2.90 a–c | 2.09 a | |
| 2.0% BWE | 61.41 a | 32.15 a | 20.40 a | 9.46 a | 3.72 a | 2.49 a | 63.41 a | 30.15 a | 17.79 a | 7.43 a | 3.11 a | 2.19 a | |
| 2.5% BWE | 60.41 a | 30.15 ab | 19.8 0 ab | 9.07 ab | 3.62 ab | 2.37 a | 62.41 ab | 28.15 ab | 17.80 a | 6.87 ab | 3.01 ab | 2.13 a | |
| 3.0% BWE | 58.41 ab | 28.15 b–d | 19.46 a–c | 8.46 bc | 3.43 a–c | 2.14 a–c | 59.41 a–d | 27.15 b | 17.37 a | 6.64 ab | 2.80 a–d | 2.08 a | |
| 60% FC | Control | 47.41 c–h | 17.15 j–l | 13.40 f–i | 3.93 jk– | 2.02 j–l | 1.29 h–k | 48.41 e–j | 16.15 h–j | 11.40 e–g | 3.46 gh | 1.51 k–m | 1.19 i |
| Water spray | 48.4 c–f1 | 18.15 jk | 13.81 e–h | 4.36 h–j | 2.28 h–j | 1.39 g–j | 50.41 d–i | 17.15 g–i | 11.92 d–g | 3.86 g | 1.59 kl | 1.29 hi | |
| 0.5% BWE | 50.41 b–g | 20.15 h–j | 14.60 d–g | 5.06 h | 2.59 g–i | 1.59 f–i | 52.41 c–i | 19.15 fg | 12.21 c–g | 4.16 fg | 1.81 i–k | 1.36 gh | |
| 1.0% BWE | 52.41 a–f | 22.15 g–i | 15.16 d–f | 6.67 ef | 2.81 e–g | 1.69 e–g | 53.41 b–i | 21.15 ef | 13.00 b–f | 4.67 ef | 1.98 h–j | 1.46 fg | |
| 1.5% BWE | 53.41 a–e | 24.15 e–g | 15.70 c–f | 7.14 ef | 2.99 d–f | 1.74 e–g | 55.41 a–g | 22.15 de | 14.50 a–e | 5.03 de | 2.40 e–g | 1.51 ef | |
| 2.0% BWE | 55.41 a–c | 26.15 c–f | 16.54 a–f | 7.87 cd | 3.31 b–d | 1.84 c–f | 57.41 a–e | 24.15 cd | 15.76 a–c | 5.68 cd | 2.61 c–f | 1.59 e | |
| 2.5% BWE | 54.41 a–d | 24.15 e–g | 14.94 d–g | 7.04 ef | 3.11 c–e | 1.79 d–f | 55.41 a–g | 23.15 de | 15.30 a–d | 5.03 de | 2.41 e–g | 1.55 ef | |
| 3.0% BWE | 52.41 a–f | 23.15 f–h | 15.10 d–f | 6.46 fg | 2.80 e–g | 1.63 e–h | 54.41 a–h | 21.15 ef | 14.76 a–e | 4.77 ef | 2.22 gh | 1.49 ef | |
| 30% FC | Control | 39.41 h | 11.15 m | 9.07 j | 1.87 n | 1.52 m | 0.69 m | 41.41 j | 9.15 m | 7.90 h | 1.47 k | 1.02 n | 0.60 n |
| Water spray | 40.41 h | 12.15 m | 9.60 ij | 2.36 mn | 1.77 lm | 0.79 lm | 42.41 j | 10.15 lm | 8.23 h | 1.73 k | 1.13 mn | 0.68 mn | |
| 0.5% BWE | 42.41 gh | 14.15 lm | 10.08 h–j | 2.96 lm | 1.80 lm | 0.95 k–m | 44.41 ij | 12.15 kl | 8.86 gh | 2.17 jk | 1.31 l–n | 0.78 lm | |
| 1.0% BWE | 43.41 f–h | 16.15 kl | 10.50 h–j | 3.57 kl | 1.91 j–l | 1.09 j–l | 46.41 g–j | 14.15 jk | 9.30 gh | 2.64 ij | 1.42 lm | 0.85 kl | |
| 1.5% BWE | 45.41 d–h | 18.15 jk | 10.61 h–j | 4.17 i–k | 2.02 j–l | 1.14 j–l | 47.41 f–j | 16.15 h–j | 10.0 f–h4 | 3.01 hi | 1.51 k–m | 0.95 jk | |
| 2.0% BWE | 47.41 c–h | 19.15 i–k | 11.20 g–j | 4.87 hi | 2.21 i–k | 1.24 i–k | 48.41 e–j | 18.15 gh | 10.70 f–h | 3.55 gh | 1.62 j–l | 0.98 j | |
| 2.5% BWE | 45.41 d–h | 17.15 j–l | 10.50 h–j | 4.23 i–k | 2.01 j–l | 1.19 jk | 46.41 g–j | 17.15 g–i | 10.30 f–h | 2.86 h–j | 1.52 kl | 0.95 jk | |
| 3.0% BWE | 44.41 e–h | 16.15 kl | 10.06 h–j | 3.83 jk | 1.89 k–m | 0.99 k–m | 45.41 h–j | 15.15 ij | 9.80 f–h | 2.57 ij | 1.42 lm | 0.88 j–l | |
| Ihsan-2016 | Galaxy-2013 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| A (µmol Co2 m−2 s−1) | E (mmol H2o m−2 s−1) | Ci (µmol Co2 mol−2 air−1) | Gs (mmol H2o m−2 s−1) | A (µmol Co2 m−2 s−1) | E (mmol H2o m−2 s−1) | Ci (µmol Co2 mol−2 air−1) | Gs (mmol H2o m−2 s−1) | ||
| 100% FC | Control | 4.07 gh | 3.28 g–i | 148 g–i | 0.19 h–j | 3.57 f | 3.36 f–h | 135 g–i | 0.15 gh |
| Water priming | 4.26 f–h | 3.40 f–i | 160 e–h | 0.20 g–i | 3.87 ef | 3.90 ef | 145 f–h | 0.17 fg | |
| 0.5% BWE | 4.86 e–g | 3.90 d–g | 175 c–f | 0.22 f–h | 4.36 de | 4.30 de | 165 d–f | 0.20 ef | |
| 1.0% BWE | 5.34 c–e | 4.30 cd | 195 b–d | 0.26 d–f | 4.86 cd | 4.80 cd | 185 a–d | 0.24 b–e | |
| 1.5% BWE | 6.07 a–d | 4.80 ac | 215 ab | 0.31 a–c | 5.57 ab | 5.40 ab | 200 ab | 0.26 a–c | |
| 2.0% BWE | 6.64 a | 5.40 a | 225 a | 0.35 a | 6.16 a | 5.90 a | 210 a | 0.30 a | |
| 2.5% BWE | 6.44 ab | 5.10 ab | 215 ab | 0.33 ab | 5.77 a | 5.60 a | 200 ab | 0.28 ab | |
| 3.0% BWE | 6.13 a–c | 4.80 a–c | 200 a–c | 0.31 a–c | 5.56 ab | 5.40 ab | 195 a–c | 0.25 b–d | |
| 60% FC | Control | 2.84 i–k | 2.36 jk | 110 jk | 0.13 k–m | 2.46 h | 2.42 j–l | 100 j–l | 0.09 i–k |
| Water priming | 3.06 ij | 2.50 jk | 120 j | 0.15 j–l | 2.67 gh | 2.70 i–l | 110 i–k | 0.10 i–k | |
| 0.5% BWE | 3.44 hi | 2.90 h–j | 135 h–j | 0.20 g–i | 3.26 fg | 3.00 hi | 125 h–j | 0.13 g–i | |
| 1.0% BWE | 4.23 f–h | 3.50 e–h | 165 e–g | 0.24 e–g | 3.86 ef | 3.70 fg | 145 f–h | 0.17 fg | |
| 1.5% BWE | 5.04 ef | 4.10 de | 185 c–e | 0.28 c–e | 4.76 cd | 4.50 cd | 165 d–f | 0.22 c–e | |
| 2.0% BWE | 5.63 b–e | 4.50 b–d | 195 b–d | 0.30 b–d | 5.04 bc | 5.00 bc | 180 b–e | 0.26 a–c | |
| 2.5% BWE | 5.24 de | 4.30 cd | 180 c–e | 0.27 c–e | 4.76 cd | 4.70 cd | 170 c–f | 0.24 b–e | |
| 3.0% BWE | 5.03 ef | 4.00 d–f | 170 d–g | 0.25 ef | 4.56 cd | 4.40 de | 155 e–g | 0.21 d–f | |
| 30% FC | Control | 1.87 l | 1.40 m | 78 l | 0.09 m | 1.57 j | 1.70 m | 70 m | 0.06 k |
| Water priming | 2.17 kl | 1.70 lm | 90 kl | 0.10 m | 1.77 ij | 1.90 lm | 75 lm | 0.07 jk | |
| 0.5% BWE | 2.57 j–l | 2.10 kl | 110 jk | 0.12 lm | 2.17 h–j | 2.30 kl | 90 k–m | 0.10 i–k | |
| 1.0% BWE | 2.77 i–k | 2.40 jk | 125 ij | 0.15 j–l | 2.27 hi | 2.90 h–j | 110 i–k | 0.12 hi | |
| 1.5% BWE | 2.87 i–k | 2.60 jk | 135 h–j | 0.17 i–k | 2.37 hi | 3.1 hi | 120 h–j | 0.13 g–i | |
| 2.0% BWE | 2.96 i–k | 2.80 ij | 150 f–i | 0.19 h–j | 2.57 h | 3.20 g–i | 130 g–i | 0.15 gh | |
| 2.5% BWE | 2.77 i–k | 2.50 jk | 135 h–j | 0.18 h–j | 2.37 hi | 3.00 hi | 120 h–j | 0.13 g–i | |
| 3.0% BWE | 2.56 j–l | 2.30 j–l | 130 ij | 0.16 i–l | 2.09 h–j | 2.70 i–k | 110 i–k | 0.11 h–j | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khaliq, A.; Ibrahim, M.U.; Hussain, S.; Zia Ul Haq, M.; Al-Huqail, A.A.; Nawaz, M.; Ali, B.; Khan, F.; Ali, H.M.; Siddiqui, M.H. The Hormetic Effects of a Brassica Water Extract Triggered Wheat Growth and Antioxidative Defense under Drought Stress. Appl. Sci. 2022, 12, 4582. https://doi.org/10.3390/app12094582
Khaliq A, Ibrahim MU, Hussain S, Zia Ul Haq M, Al-Huqail AA, Nawaz M, Ali B, Khan F, Ali HM, Siddiqui MH. The Hormetic Effects of a Brassica Water Extract Triggered Wheat Growth and Antioxidative Defense under Drought Stress. Applied Sciences. 2022; 12(9):4582. https://doi.org/10.3390/app12094582
Chicago/Turabian StyleKhaliq, Abdul, Muhammad Usman Ibrahim, Saddam Hussain, Muhammad Zia Ul Haq, Asma A. Al-Huqail, Muhammad Nawaz, Basharat Ali, Faheema Khan, Hyassam M. Ali, and Manzer H. Siddiqui. 2022. "The Hormetic Effects of a Brassica Water Extract Triggered Wheat Growth and Antioxidative Defense under Drought Stress" Applied Sciences 12, no. 9: 4582. https://doi.org/10.3390/app12094582
APA StyleKhaliq, A., Ibrahim, M. U., Hussain, S., Zia Ul Haq, M., Al-Huqail, A. A., Nawaz, M., Ali, B., Khan, F., Ali, H. M., & Siddiqui, M. H. (2022). The Hormetic Effects of a Brassica Water Extract Triggered Wheat Growth and Antioxidative Defense under Drought Stress. Applied Sciences, 12(9), 4582. https://doi.org/10.3390/app12094582