
Carum carvi Modulates Acetaminophen-Induced Hepatotoxicity: Effects on TNF-α, NF-κB, and Caspases

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Procedure
2.3.1. Experiment 1: In Vitro Evaluation of Antioxidant Properties of Carum carvi
2.3.2. Experiment 2: To Study Effects of Carum carvi Extract on Acetaminophen-Induced Hepatotoxicity, Oxidative Stress, TNF-α, NF-κB, Caspases 3, 7, and 9, and Histopathological Changes
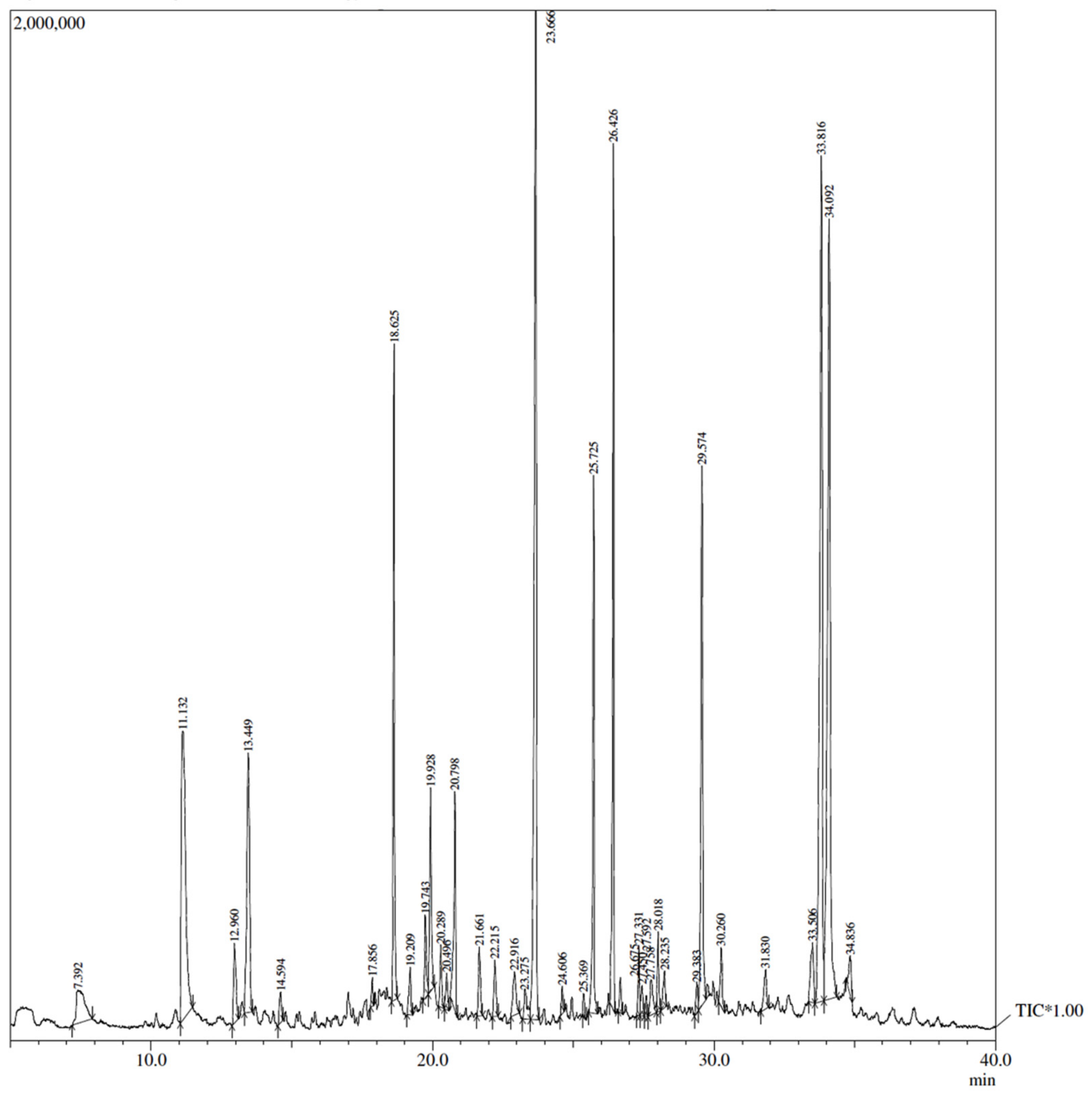
2.3.3. Analysis of the Carum carvi by GC (Gas Chromatography) Coupled with Mass Spectrometry
2.4. Post-Mitochondrial Supernatant (PMS) and Microsome Preparation
2.5. In Vitro Parameters
2.5.1. Total Phenolic Composition
2.5.2. Estimation of Lipid Peroxidation
2.5.3. Assay of DNA-Sugar Damage
2.5.4. DPPH-Scavenging Activity
2.5.5. Ferric Reducing Potential
2.6. In Vivo Parameters
2.6.1. In Vivo Enzyme Assays and Cytotoxicity Parameters
2.6.2. Cytochrome P450 Content
2.6.3. Quinone Reductase Activity
2.6.4. Expression of Caspases 3, 7, and 9
2.6.5. Tumor Necrosis Factor-Alpha (TNF-α)
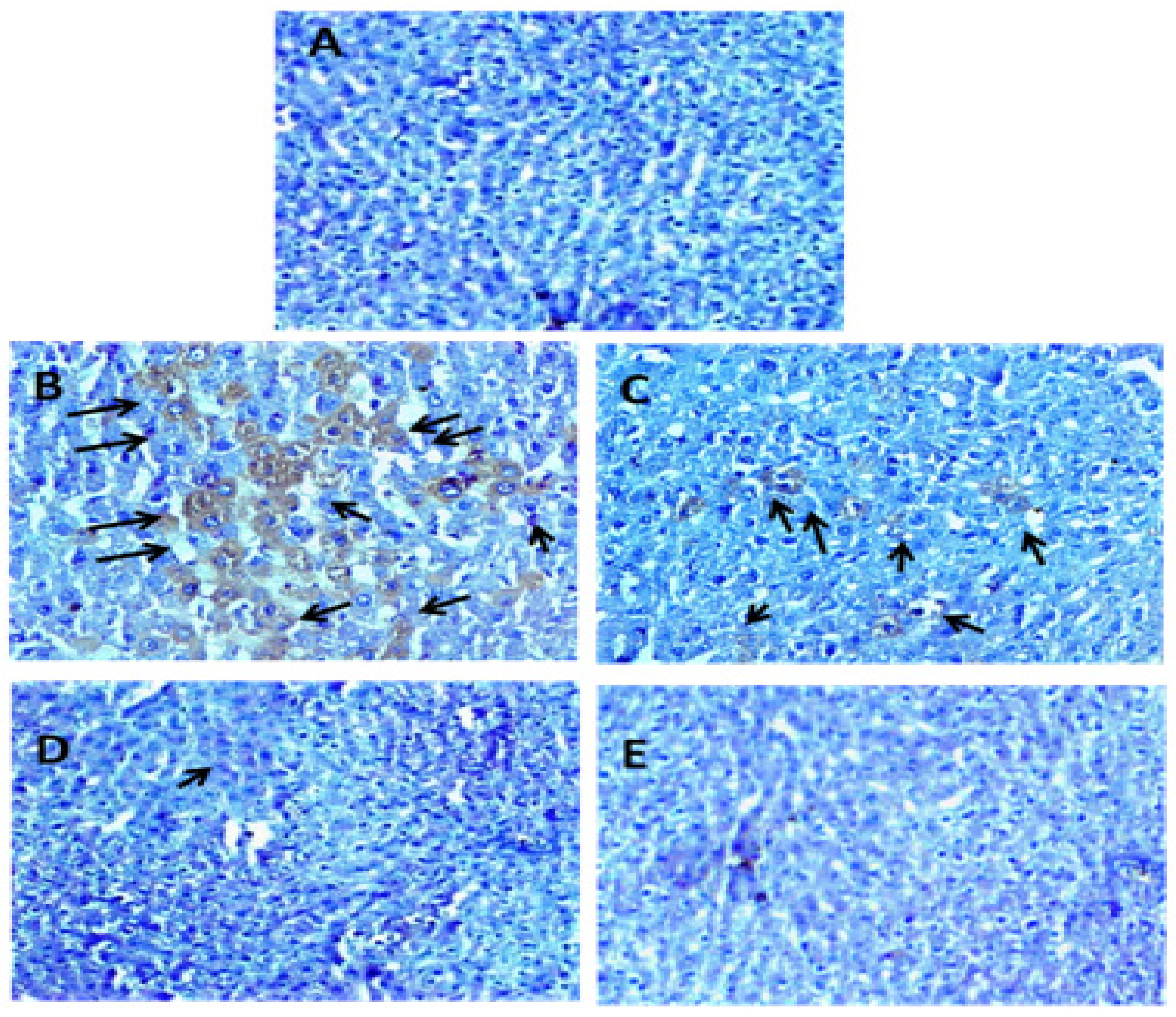
2.6.6. Immunohistochemical Staining of NF-κB and Histopathology
2.7. Statistical Analysis
3. Results
3.1. In Vitro Antioxidant Potential of Carum carvi
3.1.1. Total Phenolic Content and GC–MS Analysis
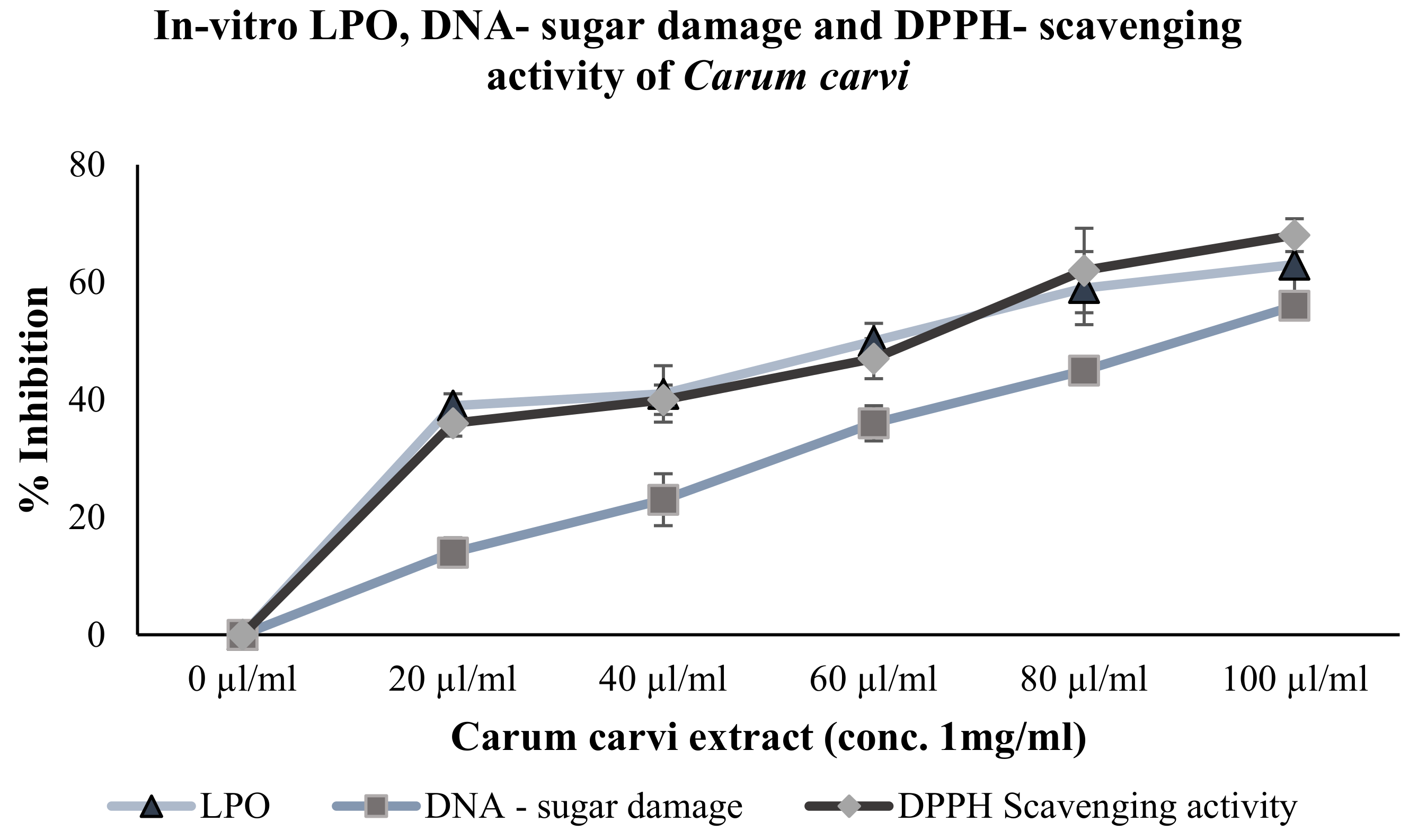
3.1.2. Inhibition of Lipid Peroxidation and DNA-Sugar Damage (In Vitro)
3.1.3. DPPH-Scavenging Activity
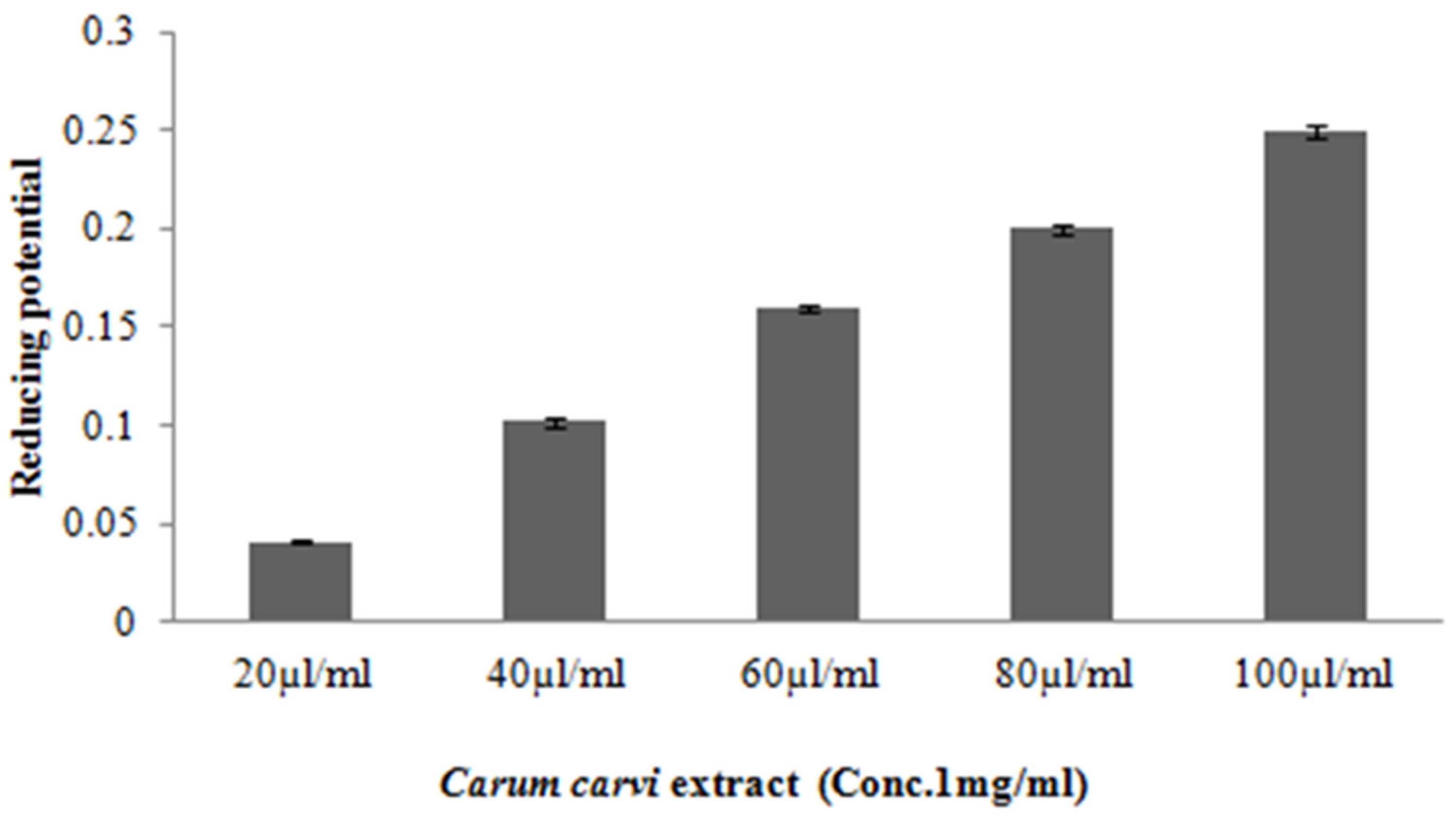
3.1.4. Ferric Reducing Potential
3.2. Effect of Carum carvi on In Vivo Antioxidant and Hepatoprotective Activities
3.2.1. Effect of Carum carvi on Liver Glutathione (Reduced) Levels, Lipid Peroxidation, and Cytochrome P450
3.2.2. Effect of Carum carvi on Intracellular Antioxidant and Phase II Drug-Metabolizing Enzymes such as Quinone Reductase (QR), Glutathione Reductase (GR), and Glutathione S-Transferase (GST) Activity
3.2.3. Carum carvi Administration Mediates Alteration of Antioxidant/Oxidant Enzymatic Activities in Liver Tissue
3.2.4. Levels of Serum Toxicity Markers in Rats Treated with Carum carvi
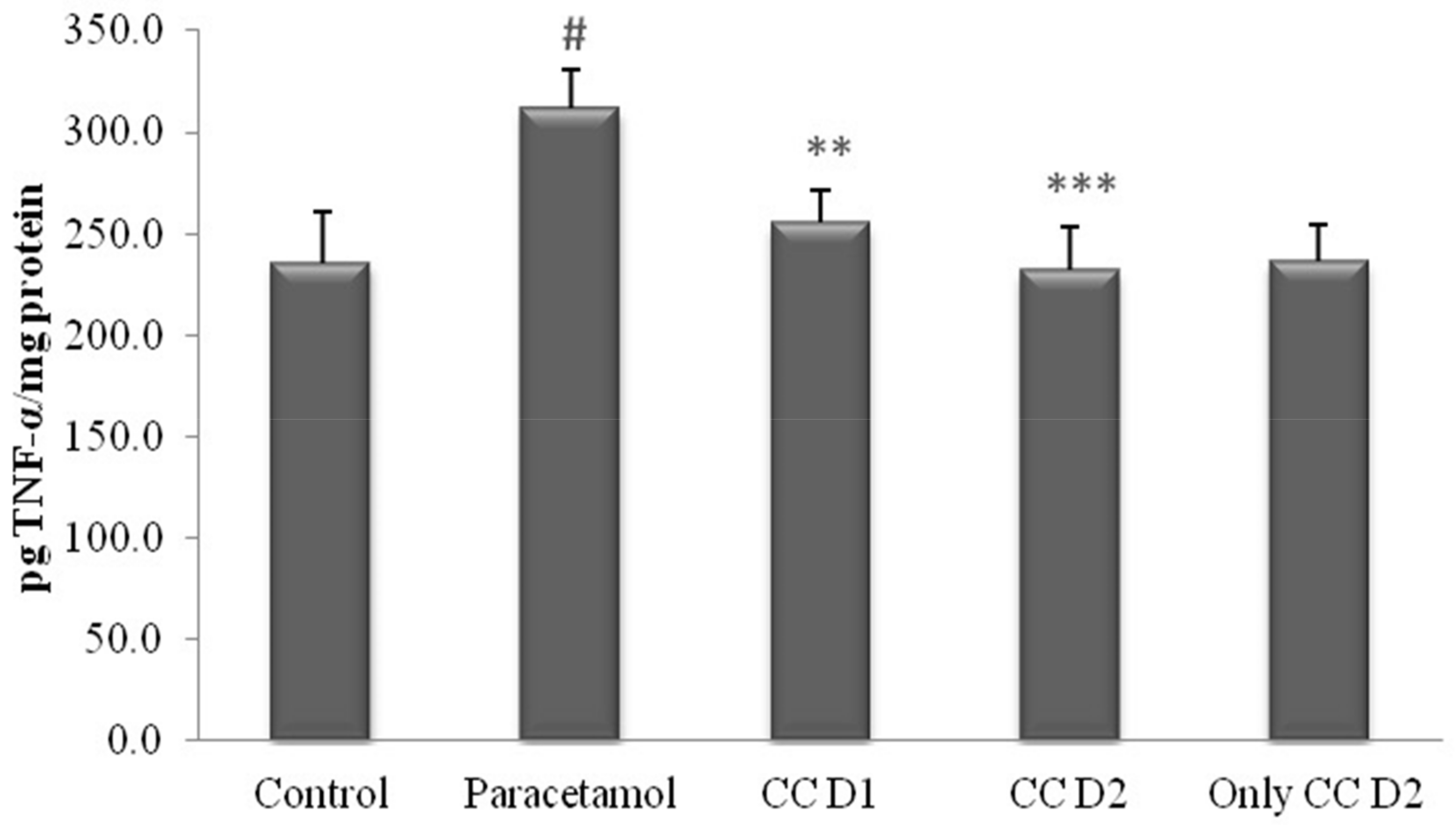
3.2.5. Effect of Carum carvi on Acetaminophen-Induced TNF-α Levels
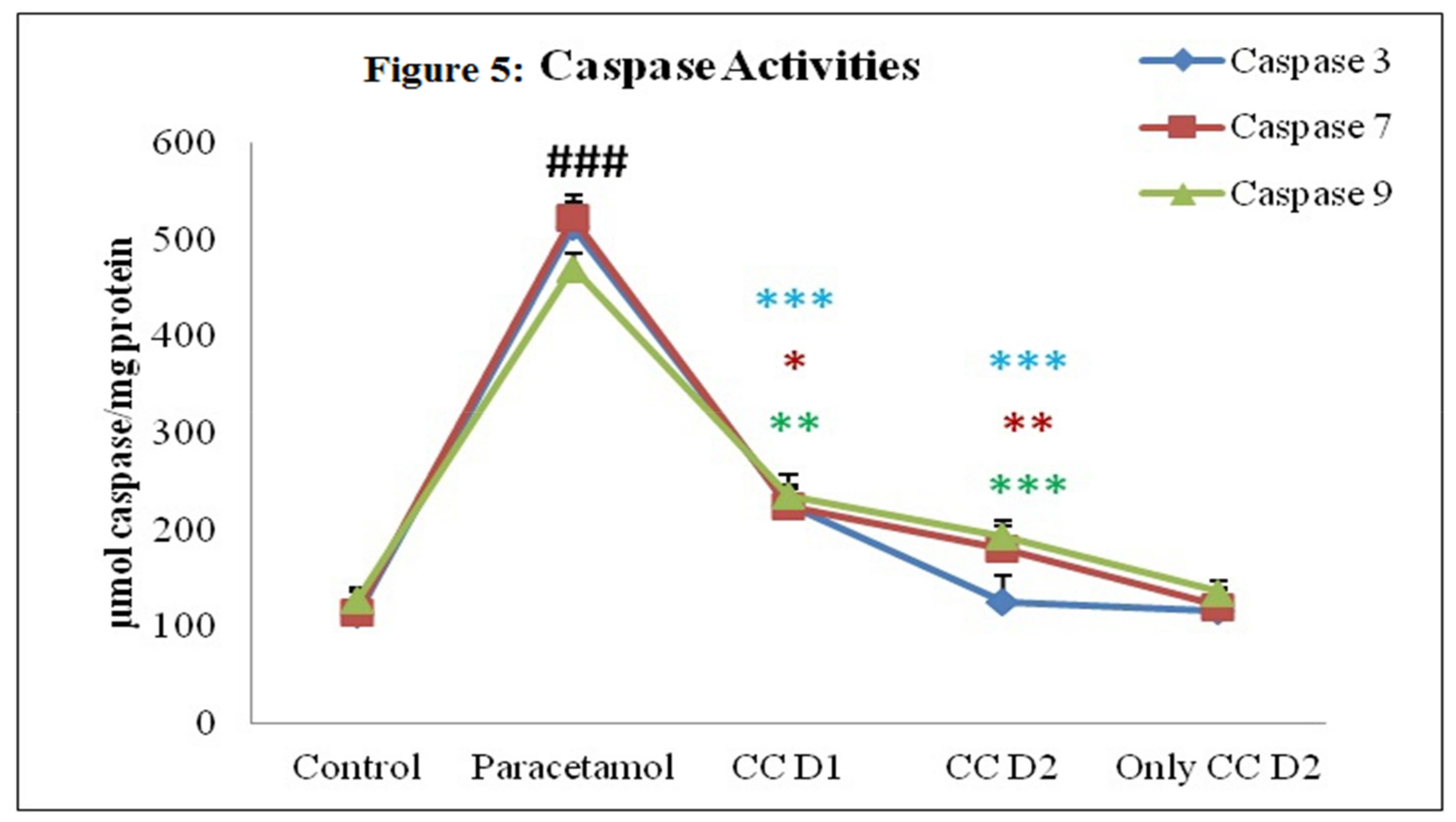
3.2.6. Effect of Carum carvi Extract on Caspase Activity
3.2.7. Effect of Carum carvi Extract on Regulation of NF-κB as a Result of Paracetamol Treatment
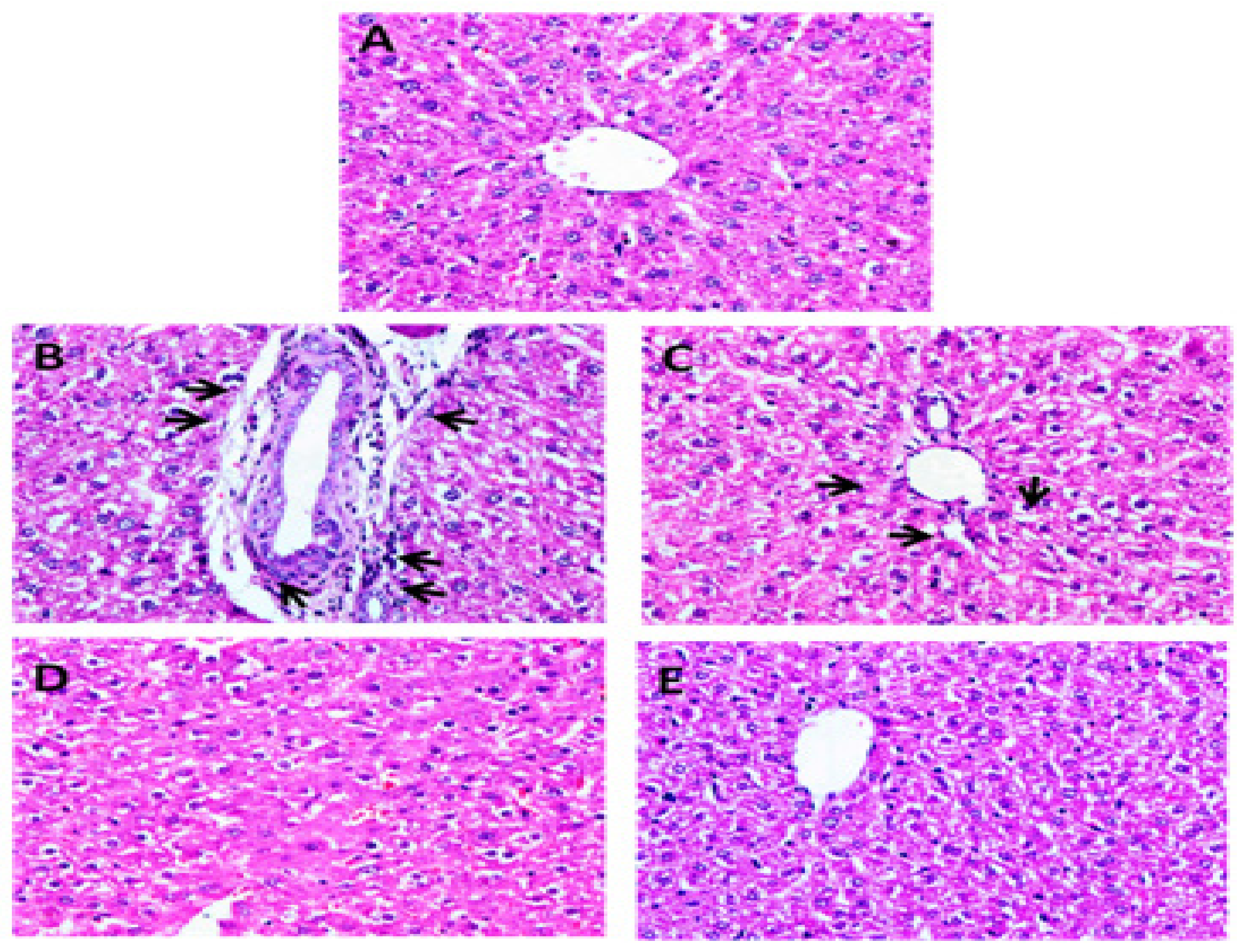
3.3. Histopathology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tujios, S.; Fontana, R.J. Mechanisms of drug-induced liver injury: From bedside to bench. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, X.; Ran, L.; Wan, J.; Wang, X.; Qin, Y.; Shu, F.; Gao, Y.; Yuan, Q.; Zhang, Q.; et al. Resveratrol improves insulin resistance, glucose and lipid metabolism in patients with non-alcoholic fatty liver disease: A randomized controlled trial. Dig. Liver Dis. 2015, 47, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Cho, W.C.; Upadhyay, G. Drug-Induced Liver Toxicity and Prevention by Herbal Antioxidants: An Overview. Front. Physiol. 2016, 6, 363. [Google Scholar] [CrossRef]
- Grattagliano, I.; Bonfrate, L.; Diogo, C.V.; Wang, H.H.; Wang, D.Q.; Portincasa, P. Biochemical mechanisms in drug-induced liver injury: Certainties and doubts. World J. Gastroenterol. WJG 2009, 15, 4865. [Google Scholar] [CrossRef]
- De Martin, E.; Michot, J.-M.; Rosmorduc, O.; Guettier, C.; Samuel, D. Liver toxicity as a limiting factor to the increasing use of immune checkpoint inhibitors. JHEP Rep. 2020, 2, 100170. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.; Gardner, C.R.; Dambach, D.M.; Brittingham, J.A.; Durham, S.K.; Laskin, J.D.; Laskin, D.L. Role of p55 tumor necrosis factor receptor 1 in acetaminophen-induced antioxidant defense. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 285, G959–G966. [Google Scholar] [CrossRef]
- Fisher, E.S.; Curry, S.C. Evaluation and treatment of acetaminophen toxicity. Adv. Pharmacol. 2019, 85, 263–272. [Google Scholar]
- Wong, L.T.; Whitehouse, L.W.; Solomonraj, G.; Paul, C.J. Pathways of disposition of acetaminophen conjugates in the mouse. Toxicol. Lett. 1981, 9, 145–151. [Google Scholar] [CrossRef]
- Bessems, J.G.M.; Vermeulen, N.P.E. Paracetamol (acetaminophen)-induced toxicity: Molecular and biochemical mechanisms, analogues and protective approaches. Crit. Rev. Toxicol. 2001, 31, 55–138. [Google Scholar] [CrossRef]
- Mazaleuskaya, L.L.; Sangkuhl, K.; Thorn, C.F.; FitzGerald, G.A.; Altman, R.B.; Klein, T.E. PharmGKB summary: Pathways of acetaminophen metabolism at the therapeutic versus toxic doses. Pharm. Genom. 2015, 25, 416–426. [Google Scholar] [CrossRef]
- Adamson, G.M.; Harman, A.W. Oxidative stress in cultured hepatocytes exposed to acetaminophen. Biochem. Pharmacol. 1993, 45, 2289–2294. [Google Scholar] [CrossRef]
- Kamiyama, T.; Sato, C.; Liu, J.; Tajiri, K.; Miyakawa, H.; Marumo, F. Role of lipid peroxidation in acetaminophen-induced hepatotoxicity: Comparison with carbon tetrachloride. Toxicol. Lett. 1993, 66, 7–12. [Google Scholar] [CrossRef]
- Blazka, M.E.; Germolec, D.R.; Simeonova, P.; Bruccoleri, A.; Pennypacker, K.R.; Luster, M.I. Acetaminophen-induced hepatotoxicity is associated with early changes in NF-kB and NF-IL6 DNA binding activity. J. Inflamm. 1995, 47, 138–150. [Google Scholar]
- Laskin, D.L.; Gardner, C.R. Nonparenchymal cells, inflammatory macrophages, and hepatotoxicity. In Drug-Induced Liver Diseases; Marcel Dekker: New York, NY, USA, 2003; pp. 183–212. [Google Scholar]
- Laskin, D.L.; Pilaro, A.M.; Ji, S. Potential role of activated macrophages in acetaminophen hepatotoxicity: II. Mechanism of macrophage accumulation and activation. Toxicol. Appl. Pharmacol. 1986, 86, 216–226. [Google Scholar] [CrossRef]
- Holtmann, G.; Haag, S.; Adam, B.; Funk, P.; Wieland, V.; Heydenreich, C.-J. Effects of a fixed combination of peppermint oil and caraway oil on symptoms and quality of life in patients suffering from functional dyspepsia. Phytomedicine 2003, 10, 56–57. [Google Scholar] [CrossRef]
- Johri, R.K. Cuminum cyminum and Carum carvi: An update. Pharmacogn. Rev. 2011, 5, 63–72. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Wright, J.R.; Colby, H.D.; Miles, P.R. Cytosolic factors which affect microsomal lipid peroxidation in lung and liver. Arch. Biochem. Biophys. 1981, 206, 296–304. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Formation of a thiobarbituric-acid-reactive substance from deoxyribose in the presence of iron salts: The role of superoxide and hydroxyl radicals. FEBS Lett. 1981, 128, 347–352. [Google Scholar] [CrossRef]
- Fenglin, H.; Ruili, L.; Liang, M. Free radical scavenging activity of extracts prepared from fresh leaves of selected Chinese medicinal plants. Fitoterapia 2004, 75, 14–23. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Tahir, M.; Sultana, S. Chrysin modulates ethanol metabolism in Wistar rats: A promising role against organ toxicities. Alcohol Alcohol. 2011, 46, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Omufca, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. J. Biol. Chem. 1964, 239, 2379–2385. [Google Scholar]
- Benson, A.M.; Hunkeler, M.J.; Talalay, P. Increase of NAD (P) H: Quinone reductase by dietary antioxidants: Possible role in protection against carcinogenesis and toxicity. Proc. Natl. Acad. Sci. USA 1980, 77, 5216–5220. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Rehman, M.U.; Lateef, A.; Khan, R.; Khan, A.Q.; Qamar, W.; Ali, F.; O’Hamiza, O.; Sultana, S. Diosmin protects against ethanol-induced hepatic injury via alleviation of inflammation and regulation of TNF-α and NF-κB activation. Alcohol 2013, 47, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-C.; Shiau, C.-Y.; Chen, H.-M.; Chiou, T.-K. Antioxidant activities of carnosine, anserine, some free amino acids and their combination. J. Food Drug Anal. 2003, 11, 13. [Google Scholar] [CrossRef]
- Chandrasekar, M.J.N.; Bommu, P.; Nanjan, M.L.J.; Suresh, B. Chemoprotective effect of Phyllanthus maderaspatensis. In modulating cisplatin-induced nephrotoxicity and genotoxicity. Pharm. Biol. 2006, 44, 100–106. [Google Scholar] [CrossRef]
- Przybyła, G.W.; Szychowski, K.A.; Gmiński, J. Paracetamol—An old drug with new mechanisms of action. Clin. Exp. Pharmacol. Physiol. 2020, 48, 3–19. [Google Scholar] [CrossRef]
- Sabir, S.M.; Rocha, J.B.T. Water-extractable phytochemicals from Phyllanthus niruri exhibit distinct in vitro antioxidant and in vivo hepatoprotective activity against paracetamol-induced liver damage in mice. Food Chem. 2008, 111, 845–851. [Google Scholar] [CrossRef]
- Nagi, M.N.; Almakki, H.A.; Sayed-Ahmed, M.M.; Al-Bekairi, A.M. Thymoquinone supplementation reverses acetaminophen-induced oxidative stress, nitric oxide production and energy decline in mice liver. Food Chem. Toxicol. 2010, 48, 2361–2365. [Google Scholar] [CrossRef]
- Hinson, J.A.; Bucci, T.J.; Irwin, L.K.; Michael, S.L.; Mayeux, P.R. Effect of inhibitors of nitric oxide synthase on acetaminophen-induced hepatotoxicity in mice. Nitric Oxide 2002, 6, 160–167. [Google Scholar] [CrossRef]
- Kozer, E.; Evans, S.; Barr, J.; Greenberg, R.; Soriano, I.; Bulkowstein, M.; Petrov, I.; Chen-Levi, Z.; Barzilay, B.; Berkovitch, M. Glutathione, glutathione-dependent enzymes and antioxidant status in erythrocytes from children treated with high-dose paracetamol. Br. J. Clin. Pharmacol. 2003, 55, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Ajiboye, T.O.; Salau, A.K.; Yakubu, M.T.; Oladiji, A.T.; Akanji, M.A.; Okogun, J.I. Acetaminophen perturbed redox homeostasis in Wistar rat liver: Protective role of aqueous Pterocarpus osun leaf extract. Drug Chem. Toxicol. 2010, 33, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Kon, K.; Kim, J.-S.; Jaeschke, H.; Lemasters, J.J. Mitochondrial permeability transition in acetaminophen-induced necrosis and apoptosis of cultured mouse hepatocytes. Hepatology 2004, 40, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Song, C.; Chen, J.; Zhou, L.; Jiang, X.; Cao, X.; Zhang, Q.I. Limonin ameliorates acetaminophen-induced hepatotoxicity by activating Nrf2 antioxidative pathway and inhibiting NF-κB inflammatory response via upregulating Sirt1. Phytomedicine 2020, 69, 153211. [Google Scholar] [CrossRef]
- Jaeschke, H.; Knight, T.R.; Bajt, M.L. The role of oxidant stress and reactive nitrogen species in acetaminophen hepatotoxicity. Toxicol. Lett. 2003, 144, 279–288. [Google Scholar] [CrossRef]
- Ren, S.; Leng, J.; Xu, X.Y.; Jiang, S.; Wang, Y.P.; Yan, X.T.; Li, W. Ginsenoside Rb1, a major saponin from Panax ginseng, exerts protective effects against acetaminophen-induced hepatotoxicity in mice. Am. J. Chin. Med. 2019, 47, 1815–1831. [Google Scholar] [CrossRef]
- Lin, H.-H.; Hsu, J.-Y.; Tseng, C.-Y.; Huang, X.-Y.; Tseng, H.-C.; Chen, J.-H. Hepatoprotective Activity of Nelumbo nucifera Gaertn. Seedpod Extract Attenuated Acetaminophen-Induced Hepatotoxicity. Molecules 2022, 27, 4030. [Google Scholar] [CrossRef]
- Yi, R.K.; Song, J.L.; Lim, Y.I.; Kim, Y.K.; Park, K.Y. Preventive effect of the Korean traditional health drink (Taemyeongcheong) on acetaminophen-induced hepatic damage in ICR Mice. Prev. Nutr. Food Sci. 2015, 20, 52. [Google Scholar] [CrossRef]
- Keshavarz, A.; Minaiyan, M.; Ghannadi, A.; Mahzouni, P. Effects of Carum carvi L. (Caraway) extract and essential oil on TNBS-induced colitis in rats. Res. Pharm. Sci. 2013, 8, 1–8. [Google Scholar]
- De Carvalho, C.C.C.R.; da Fonseca, M.M.R. Carvone: Why and how should one bother to produce this terpene. Food Chem. 2006, 95, 413–422. [Google Scholar] [CrossRef]
- Seddighfar, M.; Mirghazanfari, S.M.; Dadpay, M. Analgesic and anti-inflammatory properties of hydroalcoholic extracts of Malva sylvestris, Carum carvi or Medicago sativa, and their combination in a rat model. J. Integr. Med. 2020, 18, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, P.; Singh, D.K. A review on the pharmacological aspects of Carum carvi. J. Biol. Earth Sci. 2014, 4, M1–M13. [Google Scholar]
- Saleh, M.A.; Clark, S.; Woodard, B. Antioxidant and free radical scavenging activities of essential oils. Ethn. Dis. 2010, 20, 78–82. [Google Scholar]
- Khayyal, M.T.; El-Ghazaly, M.A.; Kenawy, S.A.; Seif-El-Nasr, M.; Mahran, L.G.; Kafafi, Y.A.; Okpanyi, S.N. Antiulcerogenic effect of some gastrointestinally acting plant extracts and their combination. Arzneimittelforschung 2001, 51, 545–553. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | R. Time | Area | Area% | Name |
|---|---|---|---|---|
| 1 | 7.392 | 1,369,937 | 1.83 | Acetic acid |
| 2 | 11.132 | 5,981,212 | 7.99 | Propanedioc acid |
| 3 | 12.960 | 903,621 | 1.21 | Cyclohexanone |
| 4 | 13.449 | 3,505,814 | 4.68 | 1,6-diphenyl-1,5-hexadiene-3,4-diol |
| 5 | 14.594 | 302,041 | 0.40 | 1-acetylcyclohexene |
| 6 | 17.856 | 178,691 | 0.24 | 1,7-octadiyne |
| 7 | 18.625 | 4,052,419 | 5.41 | 4-methyl-benzenepropanol |
| 8 | 19.209 | 475,531 | 0.64 | 1-methylencyclopropyl-ethanol |
| 9 | 19.743 | 701,253 | 0.94 | 4-(3-methyl-2-butenyl)-4-cyclopentene-1,3-dione |
| 10 | 19.928 | 1,684,050 | 2.25 | 1-hydroxy-2-methyl-5-isopropylbenzene |
| 11 | 20.289 | 518,031 | 0.69 | 1-hydroxy-2-methyl-5-isopropylbenzene |
| 12 | 20.496 | 332,170 | 0.44 | 1-Bromo-2,2,3,3-tetramethyl-1-(1-propynyl)cyclopropane |
| 13 | 20.798 | 1,847,276 | 2.47 | Methyl eicosanoate |
| 14 | 21.661 | 641,813 | 0.86 | 1,3-di-o-methylpyrogallol |
| 15 | 22.215 | 535,091 | 0.71 | 2-nitrocumene |
| 16 | 22.916 | 662,203 | 0.88 | 2,3-dihydroxypropanal |
| 17 | 23.275 | 511,835 | 0.68 | 1(2H)-quinoline carboxylic acid |
| 18 | 23.666 | 10,890,129 | 14.55 | 3-Ethyl-3-phenyl-2,6-piperidinedione |
| 19 | 24.606 | 218,246 | 0.29 | 2-Methyl-4-[5-(4-methyl-3-nitrophenyl)-2-furyl]-n-phenyl-1,4-dihydropyrimido [1,2-a]benzimidazole-3-carboxamide |
| 20 | 25.369 | 145,663 | 0.19 | 4-Methylpentanoic acid |
| 21 | 25.725 | 3,607,495 | 4.82 | Methyl 5-(2-undecylcyclopropyl) pentanoate |
| 22 | 26.426 | 4,901,363 | 6.55 | Methyl linolelaidate |
| 23 | 26.675 | 282,385 | 0.38 | 2-Methyl-6-methylene-7-octen-2-ol |
| 24 | 27.331 | 452,416 | 0.60 | Methyl (5E,8E)-5,8-octadecadienoate |
| 25 | 27.450 | 294,650 | 0.39 | 7,12-Dihydro-6,7-bis(4-hydroxyphenyl)-6H-[1,2,4]triazolo[1’,5’:1,2]pyrimido[5,4-c]chromen-2-ol |
| 26 | 27.592 | 133,563 | 0.18 | 5,9-Dimethyl-1-decanol |
| 27 | 27.758 | 553,642 | 0.74 | 3-Methyl-3-buten-2-ol |
| 28 | 28.018 | 596,527 | 0.80 | Isobutyric acid, 2-ethylhexyl ester |
| 29 | 28.235 | 380,720 | 0.51 | 2,6-Dimethyl-1,5,7-octatriene |
| 30 | 29.383 | 285,521 | 0.38 | 11-Cyclopentylundecanoic acid |
| 31 | 29.574 | 4,695,539 | 6.27 | 1-[4-(Methoxymethyl)phenyl]ethanol |
| 32 | 30.260 | 562,950 | 0.75 | 3-Methyl-2H-indazol-2-ol |
| 33 | 31.830 | 486,928 | 0.65 | 1-Methyl-3-cyclohexenol |
| 34 | 33.506 | 839,142 | 1.12 | Pelargonic alcohol |
| 35 | 33.816 | 10,015,857 | 13.38 | Stearic acid |
| 36 | 34.092 | 10,848,949 | 14.49 | 1,2-Bis(2,6-dimethylphenyl)diazene 1,2-dioxide |
| 37 | 34.836 | 470,680 | 0.63 | 3-(2-Isopropyl-5-methylcyclohexyloxycarbonylmethyl)-2-(4-methoxyphenyl)-1-methylbenzimidazolium ion |
| CYP 450 (nmol/mg protein) | Lipid Peroxidation (nmol MDA formed/g tissue) | Catalase (nmol H2O2 consumed/min/mg protein) | Quinone Reductase (nmol DCPIP red/min/mg protein) | Xanthine Oxidase (µg of Uric Acid formed/min/mg protein) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Control | 13.91 | 0.83 | 6.7 | 0.19 | 42.27 | 1.29 | 602.12 | 25.85 | 0.11 | 0.06 |
| Acetaminophen | 28.64 a | 0.63 | 14.03 a | 1.15 | 29.41 a | 2.74 | 350.17 a | 46.5 | 0.21a | 0.08 |
| Carum carvi D1 + Acetaminophen | 18.41 ** | 0.45 | 9.8 ** | 0.7 | 38.4 * | 2.6 | 464.15 * | 39.82 | 0.15** | 0.04 |
| Carum carvi D2 + Acetaminophen | 14.21 *** | 0.87 | 7.5 *** | 0.5 | 41.24 ** | 2.1 | 510.7 ** | 49.07 | 0.12 *** | 0.03 |
| Only Carum carvi D2 | 14.01 | 0.68 | 6.4 | 0.5 | 41.39 | 3.3 | 605.83 | 50.95 | 0.12 | 0.06 |
| Glutathione Peroxidase (nmol NADPH oxidized/min/mg protein) | Glutathione Reductase (nmol NADPH oxidized/min/mg protein) | Reduced Glutathione (µmol GSH conjugate/g tissue) | Glutathione S-Transferase (nmol CDNB conjugate formed/min/mg protein) | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SD | Mean | SEM | |
| Control | 242.8 | 19.5 | 318.8 | 7.7 | 0.056 | 0.01 | 57.4 | 7.5 |
| Acetaminophen | 138.0 a | 16.8 | 218.5 a | 7.6 | 0.03 a | 0.004 | 33.25 a | 5.8 |
| Carum carvi D1 + Acetaminophen | 198.6 ** | 25.4 | 282.1 ** | 9.2 | 0.046 ** | 0.004 | 45.3 ** | 4.8 |
| Carum carvi D2 + Acetaminophen | 232.1 ** | 32.4 | 320.4 *** | 11.5 | 0.052 *** | 0.005 | 55.4 *** | 7.01 |
| Only Carum carvi D2 | 246.5 | 21.8 | 319.5 | 8.7 | 0.058 | 0.003 | 58.2 | 8.4 |
| Aspartate Aminotransferase (AST or) (IU/L) | Alanine Aminotransferase (ALT or SGPT) (IU/L) | Lactate Dehydrogenase (LDH) (nmol NADH oxidised/min/mg protein) | ||||
|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | |
| Control | 47.39 | 3.44 | 10.36 | 0.9 | 165.04 | 16.3 |
| Acetaminophen | 76.5 | 5.8 a | 19.3 | 2.3 a | 268.3 a | 23.5 |
| Carum carvi D1 + Acetaminophen | 62.1 | 5.4 ** | 14.6 * | 1.33 | 208.5 *** | 24.6 |
| Carum carvi D2 + Acetaminophen | 50.47 | 6.8 *** | 11.5 ** | 1.14 | 174.2 *** | 19.8 |
| Only Carum carvi D2 | 47.07 | 3.8 | 10.2 | 1.6 | 169.8 | 21.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mir, T.M.; Rehman, M.U.; Ashfaq, M.K.; Qamar, W.; Khan, R.; Ali, A.; Almuqbil, M.; Alshehri, S.; Sultana, S. Carum carvi Modulates Acetaminophen-Induced Hepatotoxicity: Effects on TNF-α, NF-κB, and Caspases. Appl. Sci. 2022, 12, 11010. https://doi.org/10.3390/app122111010
Mir TM, Rehman MU, Ashfaq MK, Qamar W, Khan R, Ali A, Almuqbil M, Alshehri S, Sultana S. Carum carvi Modulates Acetaminophen-Induced Hepatotoxicity: Effects on TNF-α, NF-κB, and Caspases. Applied Sciences. 2022; 12(21):11010. https://doi.org/10.3390/app122111010
Chicago/Turabian StyleMir, Tahir Maqbool, Muneeb U Rehman, Mohammad Khalid Ashfaq, Wajhul Qamar, Rehan Khan, Aarif Ali, Mansour Almuqbil, Sultan Alshehri, and Sarwat Sultana. 2022. "Carum carvi Modulates Acetaminophen-Induced Hepatotoxicity: Effects on TNF-α, NF-κB, and Caspases" Applied Sciences 12, no. 21: 11010. https://doi.org/10.3390/app122111010
APA StyleMir, T. M., Rehman, M. U., Ashfaq, M. K., Qamar, W., Khan, R., Ali, A., Almuqbil, M., Alshehri, S., & Sultana, S. (2022). Carum carvi Modulates Acetaminophen-Induced Hepatotoxicity: Effects on TNF-α, NF-κB, and Caspases. Applied Sciences, 12(21), 11010. https://doi.org/10.3390/app122111010