
Effect of Postharvest UV-C Radiation on Nutritional Quality, Oxidation and Enzymatic Browning of Stored Mature Date


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical, Reagents and Materials
2.2. Plant Material and Experimental Design
2.3. Visual Analysis and Determination of Water Content
2.4. Determination of Carotenoid Content
2.5. Determination of Total Sugar Content
2.6. Determination of Total Protein Content, PPO and POD Activities
2.7. Determination of Browning Index and Contents of Flavonoids and Condensed Tannins
2.8. Determination of Total Polyphenol Content
2.9. Evaluation of Antioxidant Activity
2.9.1. Anti-Free-Radical Activity (DPPH Test)
2.9.2. Ferric-Reducing Power Test
2.10. Statistical Analysis
3. Results and Discussion
3.1. Changes in External Color and in Carotenoid Content
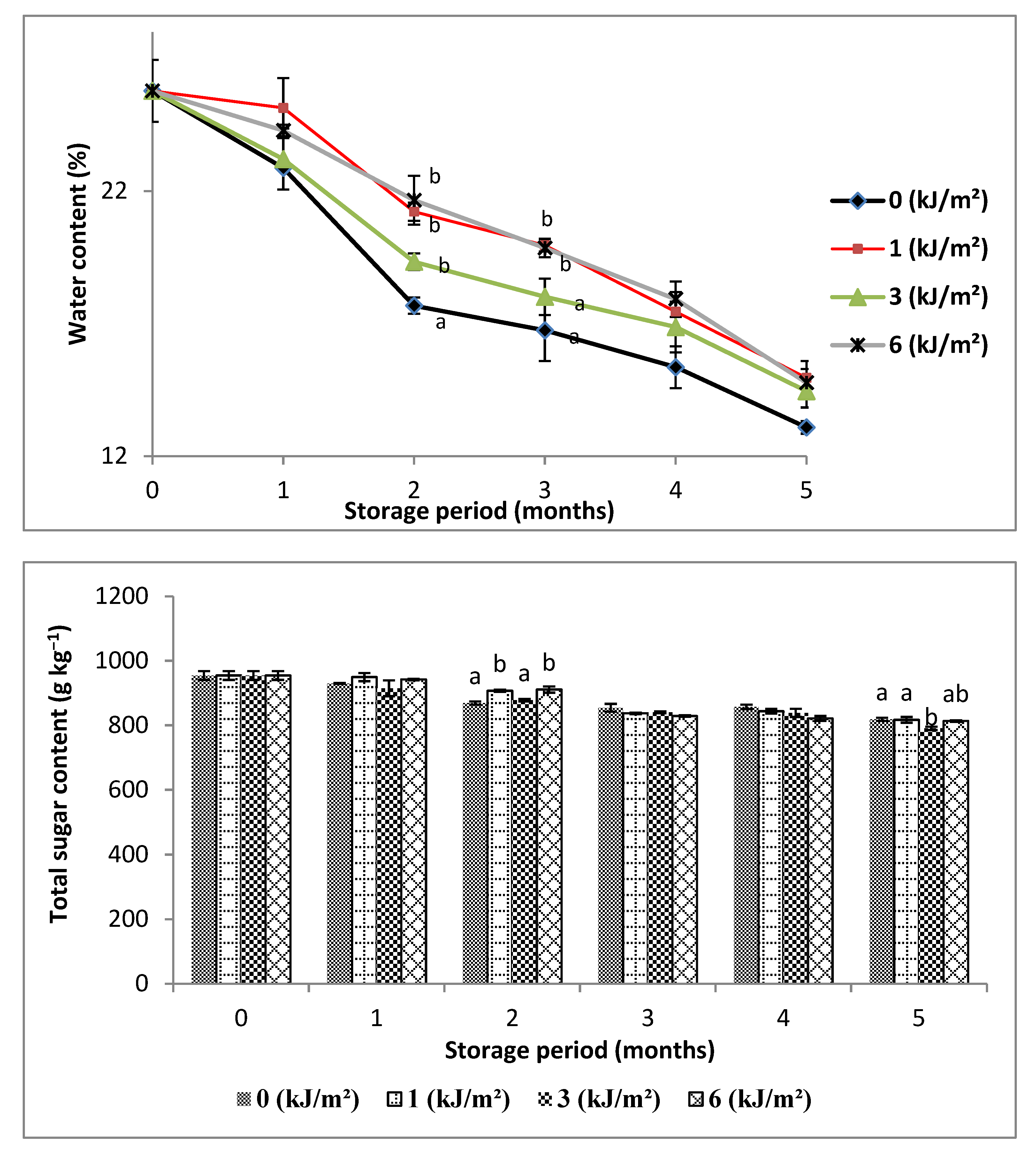
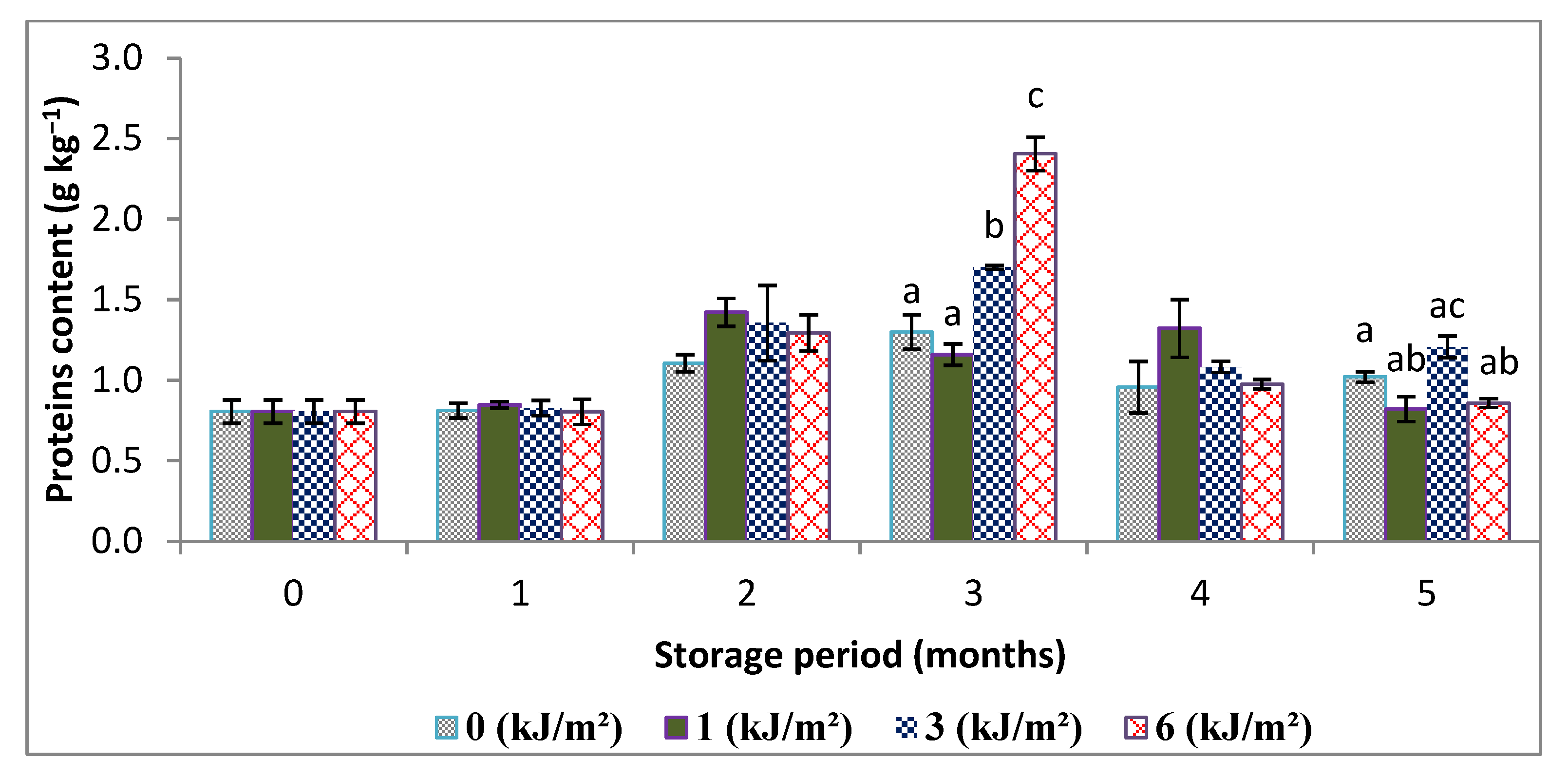
3.2. Changes in Moisture, Total Sugar and Protein Contents
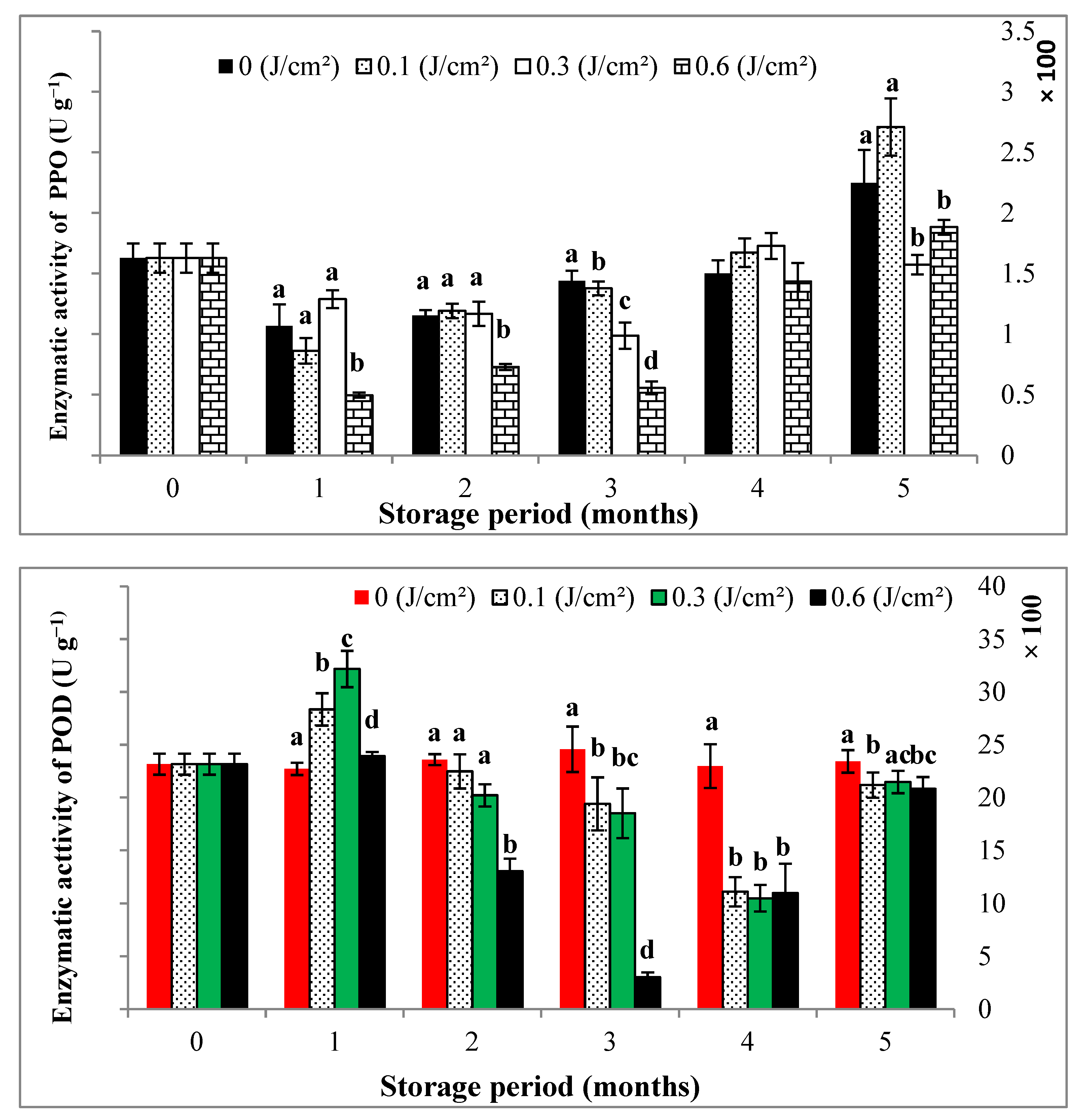
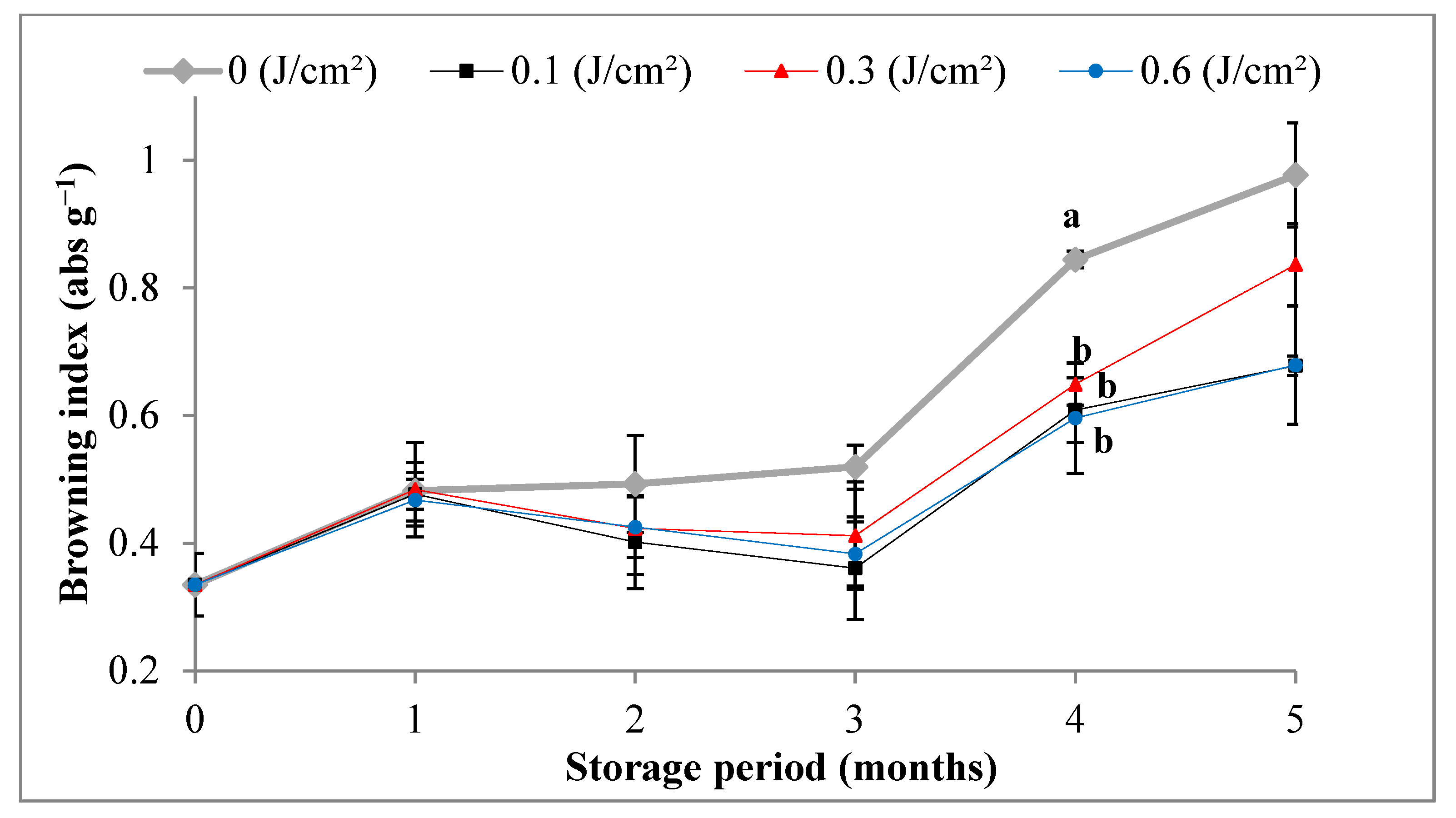
3.3. Enzymatic Activity of Polyphenoloxidase, Peroxidase and Browning Index
3.4. Total Polyphenol Content (TPC), Flavonoids and Condensed Tannins
3.5. Anti-Free-Radical Activity and Ferric-Reducing Power
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El Gharras, H. Polyphenols: Food sources, properties and applications—A review. Int. J. Food Sci. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Baliga, M.S.; Baliga, B.R.V.; Kandathil, S.M.; Bhat, H.P.; Vayalil, P.K. A review of the chemistry and pharmacology of the date fruits (Phoenix dactylifera L.). Food Res. Int. 2011, 44, 1812–1822. [Google Scholar] [CrossRef]
- Daas Amiour, S.; Alloui-Lombarkia, O.; Bouhdjila, F.; Ayachi, A.; Hambaba, L. Étude de l’implication des composés phénoliques des extraits de trois variétés de datte dans son activité antibactérienne. Phytothérapie 2014, 12, 135–142. [Google Scholar] [CrossRef]
- Dassamiour, S.; Vidal, V.; Laurent, S.; Sallanon, H.; Charles, F. Effect of gaseous pretreatment on enzymatic browning of mature date after cold storage. Fruits 2018, 73, 243–251. [Google Scholar] [CrossRef]
- Daas Amiour, S.; Hambaba, L. Effect of pH, temperature and some chemicals on polyphenoloxidase and peroxidase activities in harvested Deglet Nour and Ghars dates. Postharvest Biol. Technol. 2016, 111, 77–82. [Google Scholar] [CrossRef]
- Xuetong, F.; Sommers, C.H. Irradiation of Fresh and Fresh-Cut Fruits and Vegetables: Quality and Shelf Life. Ch. 15. In Food Irradiation Research and Technology, 2nd ed.; Blackwell Publishing and the Institute of Food Technologists: Hoboken, NJ, USA, 2012; pp. 271–293. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lichanporn, I.; Techavuthiporn, C. The effects of nitric oxide and nitrous oxide on enzymatic browning in longkong (Aglaia dookkoo Griff.). Postharvest Biol. Technol. 2013, 86, 62–65. [Google Scholar] [CrossRef]
- Masuda, T.; Yonemori, S.; Oyama, Y.; Takeda, Y.; Tanaka, T.; Andoh, T.; Nakata, M. Evaluation of the antioxidant activity of environmental plants: Activity of the leaf extracts from seashore plants. J. Agric. Food Chem. 1999, 47, 1749–1754. [Google Scholar] [CrossRef]
- Oyaizu, M. Antioxidative activities of browning products of glucosamine fractionated by organic solvent and thin-layer chromatography. Nippon Shokuhin Kogyo Gakkaishi J. Jpn. Soc. Food Sci. 1988, 35, 771–775. [Google Scholar] [CrossRef]
- Alegria, C.; Pinheiro, J.; Duthoit, M.; Gonçalves, E.M.; Moldão-Martins, M.; Abreu, M. Fresh-cut carrot (cv. Nantes) quality as affected by abiotic stress (heat shock and UV-C irradiation) pre-treatments. LWT Food Sci. Technol. 2012, 48, 197–203. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Changes in carotenoids during processing and storage of foods. Arch. Latinoam. Nutr. 1999, 49, 38S–47S. [Google Scholar] [PubMed]
- Boudries, H.; Kefalas, P.; Hornero-Méndez, D. Carotenoid composition of Algerian date varieties (Phoenix dactylifera) at different edible maturation stages. Food Chem. 2007, 101, 1372–1377. [Google Scholar] [CrossRef]
- Freitas, A.; Moldão-Martins, M.; Costa, H.S.; Albuquerque, T.G.; Valente, A.; Sanches-Silva, A. Effect of UV-C radiation on bioactive compounds of pineapple (Ananas comosus L. Merr.) by-products. J. Sci. Food Agric. 2015, 95, 44–52. [Google Scholar] [CrossRef]
- Mutlak, H.H.; Mann, J. Darkening of dates: Control by microwave heating. Date Palm J. 1984, 3, 303–316. [Google Scholar]
- Jemni, M.; Gómez, P.A.; Souza, M.; Chaira, N.; Ferchichi, A.; Otón, M.; Artés, F. Combined effect of UV-C, ozone and electrolyzed water for keeping overall quality of date palm. LWT Food Sci. Technol. 2014, 59, 649–655. [Google Scholar] [CrossRef]
- Lemoine, M.L.; Civello, P.M.; Martínez, G.A.; Chaves, A.R. Influence of postharvest UV-C treatment on refrigerated storage of minimally processed broccoli (Brassica oleracea var. Italica). J. Sci. Food Agric. 2007, 87, 1132–1139. [Google Scholar] [CrossRef]
- Pan, J.; Vicente, A.R.; Martínez, G.A.; Chaves, A.R.; Civello, P.M. Combined use of UV-C irradiation and heat treatment to improve postharvest life of strawberry fruit. J. Sci. Food Agric. 2004, 84, 1831–1838. [Google Scholar] [CrossRef]
- Yahia, E.M.; Kader, A.A. Date (Phoenix dactylifera L.). In Postharvest Biology and Technology of Tropical and Subtropical Fruits, 1st ed.; Woodhead Publishing: Sawston, UK, 2011. [Google Scholar] [CrossRef]
- Rygg, G.L. Date development, handling and packing in the United States. In Agricultural Handbook No. 482, 1st ed.; Agricultural Research Service, US Department of Agriculture Date Development: Washington, DC, USA, 1975. [Google Scholar]
- Hu, L.; Yang, C.; Zhang, L.; Feng, J.; Xi, W. Effect of light-emitting diodes and ultraviolet irradiation on the soluble sugar, organic acid, and carotenoid content of postharvest sweet oranges (Citrus sinensis (L.) Osbeck). Molecules 2019, 24, 3440. [Google Scholar] [CrossRef]
- Barka, E.A.; Kalantari, S.; Makhlouf, J.; Arul, J. Impact of UV-C irradiation on the cell wall-degrading enzymes during ripening of tomato (Lycopersicon esculentum L.) fruit. J. Agric. Food Chem. 2000, 48, 667–671. [Google Scholar] [CrossRef]
- Pickering, A.M.; Davies, K.J. Degradation of damaged proteins: The main function of the 20S proteasome. Prog. Mol. Biol. Transl. Sci. 2012, 109, 227–248. [Google Scholar] [CrossRef]
- Costa, L.; Vicente, A.R.; Civello, P.M.; Chaves, A.R.; Martínez, G.A. UV-C treatment delays postharvest senescence in broccoli florets. Postharvest Biol. Technol. 2006, 39, 204–210. [Google Scholar] [CrossRef]
- Xie, Z.; Fan, J.; Charles, M.T.; Charlebois, D.; Khanizadeh, S.; Rolland, D.; Roussel, D.; Zhang, Z. Preharvest ultraviolet-C irradiation: Influence on physicochemical parameters associated with strawberry fruit quality. Plant Physiol. Biochem. 2016, 108, 337–343. [Google Scholar] [CrossRef]
- Yannam, S.K.; Patras, A.; Pendyala, B.; Vergne, M.; Ravi, R.; Gopisetty, V.; Sasges, M. Effect of UV-C irradiation on the inactivation kinetics of oxidative enzymes, essential amino acids and sensory properties of coconut water. J. Food Sci. Technol. 2020, 57, 3564–3572. [Google Scholar] [CrossRef]
- Lei, J.; Li, B.; Zhang, N.; Yan, R.; Guan, W.; Brennan, C.S.; Peng, B. Effects of UV-C treatment on browning and the expression of polyphenol oxidase (PPO) genes in different tissues of Agaricusbisporus during cold storage. Postharvest Biol. Technol. 2018, 139, 99–105. [Google Scholar] [CrossRef]
- Mohammadi, N.; Mohammadi, S.; Abdossi, V.; Akbar-Boojar, M.A. Effect of UV-C radiation on antioxidant enzymes in strawberry fruit (Fragaria x ananassa cv. Camarosa). J. Agric. Biol. Sci. 2012, 7, 860–864. [Google Scholar]
- Kim, J.; Marshall, M.R.; Wei, C. Polyphenoloxidase. In Seafood Enzymes Utilization and Influence on Postharvest Seafood Quality; Haard, N.F., Simpson, B.K., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 271–315. [Google Scholar]
- Gonzalez-Aguilar, G.; Wang, C.Y.; Buta, G.J. UV-C irradiation reduces breakdown and chilling injury of peaches during cold storage. J. Sci. Food Agric. 2004, 84, 415–422. [Google Scholar] [CrossRef]
- Akgün, M.P.; Ünlütürk, S. Effects of ultraviolet light emitting diodes (LEDs) on microbial and enzyme inactivation of apple juice. Int. J. Food Microbiol. 2017, 260, 65–74. [Google Scholar] [CrossRef]
- Teoh, L.S.; Lasekan, O.; Adzahan, N.M.; Hashim, N. The effect of ultraviolet treatment on enzymatic activity and total phenolic content of minimally processed potato slices. J. Food Sci. Technol. 2016, 53, 3035–3042. [Google Scholar] [CrossRef]
- Wang, D.; Chen, L.; Ma, Y.; Zhang, M.; Zhao, Y.; Zhao, X. Effect of UV-C treatment on the quality of fresh-cut lotus (Nelumbo nucifera Gaertn.) root. Food Chem. 2019, 278, 659–664. [Google Scholar] [CrossRef]
- Winter, T.R.; Rostás, M. Ambient ultraviolet radiation induces protective responses in soybean but does not attenuate indirect defense. Environ. Pollut. 2008, 155, 290–297. [Google Scholar] [CrossRef]
- Jagadeesh, S.L.; Charles, M.T.; Gariepy, Y.; Goyette, B.; Raghavan, G.S.V.; Vigneault, C. Influence of postharvest UV-C hormesis on the bioactive components of tomato during post-treatment handling. Food Bioproc. Technol. 2011, 4, 1463–1472. [Google Scholar] [CrossRef]
- Bravo, S.; García-Alonso, J.; Martín-Pozuelo, G.; Gómez, V.; García-Valverde, V.; Navarro-González, I.; Periago, M.J. Effects of postharvest UV-C treatment on carotenoids and phenolic compounds of vine-ripe tomatoes. Int. J. Food Sci. 2013, 48, 1744–1749. [Google Scholar] [CrossRef]
- Sheng, K.; Zheng, H.; Shui, S.; Yan, L.; Liu, C.; Zheng, L. Comparison of postharvest UV-B and UV-C treatments on table grape: Changes in phenolic compounds and their transcription of biosynthetic genes during storage. Postharvest Biol. Technol. 2018, 138, 74–81. [Google Scholar] [CrossRef]
- Wu, X.; Guan, W.; Yan, R.; Lei, J.; Xu, L.; Wang, Z. Effects of UV-C on antioxidant activity, total phenolics and main phenolic compounds of the melanin biosynthesis pathway in different tissues of button mushroom. Postharvest Biol. Technol. 2016, 118, 51–58. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Howard, L. Blueberry fruit response to postharvest application of ultraviolet radiation. Postharvest Biol. Technol. 2008, 47, 280–285. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Villegas-Ochoa, M.A.; Martínez-Téllez, M.A.; Gardea, A.A.; Ayala-Zavala, J.F. Improving Antioxidant Capacity of Fresh-Cut Mangoes Treated with UV-C. J. Food Sci. 2007, 72, S197–S202. [Google Scholar] [CrossRef]
- Pombo, M.A.; Rosli, H.G.; Martínez, G.A.; Civello, P.M. UV-C treatment affects the expression and activity of defense genes in strawberry fruit (Fragaria × ananassa, Duch.). Postharvest Biol. Technol. 2011, 59, 94–102. [Google Scholar] [CrossRef]
- Kim, H.J.; Fonseca, J.M.; Kubota, C.; Kroggel, M.; Choi, J.H. Quality of fresh-cut tomatoes as affected by salt content in irrigation water and post-processing ultraviolet-C treatment. J. Sci. Food Agric. 2008, 88, 1969–1974. [Google Scholar] [CrossRef]
- Qin, G.Q.; Yan, C.L.; Wei, L.L. Effect of cadmium stress on the contents of tannin, soluble sugar and proline in Kandeliacandel (L.) Druce seedlings. Acta Ecol. Sin. 2006, 36, 112–123. [Google Scholar] [CrossRef]
- Zhang, L.H.; Shao, H.B.; Ye, G.F.; Lin, Y.M. Effects of fertilization and drought stress on tannin biosynthesis of Casuarina equisetifolia seedlings branchlets. Acta Physiol. Plant. 2012, 34, 1639–1649. [Google Scholar] [CrossRef][Green Version]
- Rabelo, M.C.; Bang, W.Y.; Nair, V.; Alves, R.E.; Jacobo-Velázquez, D.A.; Sreedharan, S.; de Miranda, M.R.A.; Cisneros-Zevallos, L. UVC light modulates vitamin C and phenolic biosynthesis in acerola fruit: Role of increased mitochondria activity and ROS production. Sci. Rep. 2020, 10, 21972–21985. [Google Scholar] [CrossRef]
- Meitha, K.; Pramesti, Y.; Suhandono, S. Reactive Oxygen Species and Antioxidants in Postharvest Vegetables and Fruits. Int. J. Food Sci. 2020, 2020, 8817778–8817788. [Google Scholar] [CrossRef]
- Park, M.H.; Kim, J.G. Low-dose UV-C irradiation reduces the microbial population and preserves antioxidant levels in peeled garlic (Allium sativum L.) during storage. Postharvest Biol. Technol. 2015, 100, 109–112. [Google Scholar] [CrossRef]
- Wallace, T.C.; Bailey, R.L.; Blumberg, J.B.; Burton-Freeman, B.; Chen, C.O.; Crowe-White, K.M.; Drewnowski, A.; Hooshmand, S.; Johnson, E.; Lewis, R.; et al. Fruits, vegetables, and health: A comprehensive narrative, umbrella review of the science and recommendations for enhanced public policy to improve intake. Crit. Rev. Food Sci. Nutr. 2020, 60, 2174–2211. [Google Scholar] [CrossRef]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Lemoine, M.L.; Civello, P.M.; Chaves, A.R.; Martínez, G.A. Influence of a combined hot air and UV-C treatment on quality parameters of fresh-cut broccoli florets at 0 °C. Int. J. Food Sci. 2010, 45, 1212–1218. [Google Scholar] [CrossRef]
- Martínez-Hernández, G.B.; Gómez, P.A.; Pradas, I.; Artés, F.; Artés-Hernández, F. Moderate UV-C pretreatment as a quality enhancement tool in fresh-cut Bimi® broccoli. Postharvest Biol. Technol. 2011, 62, 327–337. [Google Scholar] [CrossRef]
- Vicente, A.R.; Pineda, C.; Lemoine, L.; Civello, P.M.; Martinez, G.A.; Chaves, A.R. UV-C treatments reduce decay, retain quality and alleviate chilling injury in pepper. Postharvest Biol. Technol. 2005, 35, 69–78. [Google Scholar] [CrossRef]
- Molan, A.L.; Flanagan, J.; Wei, W.; Moughan, P.J. Selenium-containing green tea has higher antioxidant and prebiotic activities than regular green tea. Food Chem. 2009, 114, 829–835. [Google Scholar] [CrossRef]
- Hosseini, F.S.; Akhavan, H.R.; Maghsoudi, H.; Hajimohammadi-Farimani, R.; Balvardi, M. Effects of a rotational UV-C irradiation system and packaging on the shelf life of fresh pistachio. J. Sci. Food Agric. 2019, 99, 5229–5238. [Google Scholar] [CrossRef]
- Bhat, R. Impact of ultraviolet radiation treatments on the quality of freshly prepared tomato (Solanum lycopersicum) juice. Food Chem. 2016, 213, 635–640. [Google Scholar] [CrossRef]
- Niu, L.; Wu, Z.; Yang, L.; Wang, Y.; Xiang, Q.; Bai, Y. Antimicrobial Effect of UVC Light-Emitting Diodes against Saccharomyces cerevisiae and Their Application in Orange Juice Decontamination. J. Food Prot. 2021, 84, 139–146. [Google Scholar] [CrossRef]
- Unluturk, S.; Atilgan, M.R. Microbial Safety and Shelf Life of UV-C Treated Freshly Squeezed White Grape Juice. J. Food Sci. 2015, 80, M1831–M1841. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Period (Months) | Irradiation Doses (kJ m−2) | Carotenoid Content (mg kg−1) |
|---|---|---|
| 0 | - | 3.46 ± 0.33 |
| 1 | 0 | 3.41 ± 0.22 |
| 1 | 3.49 ± 0.20 | |
| 3 | 3.47 ± 0.10 | |
| 6 | 3.62 ± 0.07 | |
| 2 | 0 | 3.45 ± 0.12 a |
| 1 | 4.17 ± 0.07 b | |
| 3 | 3.51 ± 0.21 a | |
| 6 | 4.02 ± 0.06 b | |
| 3 | 0 | 3.35 ± 0.16 |
| 1 | 3.58 ± 0.02 | |
| 3 | 3.48 ± 0.25 | |
| 6 | 3.77 ± 0.17 | |
| 4 | 0 | 3.33 ± 0.25 |
| 1 | 3.44 ± 0.25 | |
| 3 | 3.00 ± 0.17 | |
| 6 | 3.56 ± 0.13 | |
| 5 | 0 | 3.20 ± 0.17 |
| 1 | 3.06 ± 0.03 | |
| 3 | 2.86 ± 0.06 | |
| 6 | 2.97 ± 0.08 |
| Storage Period (Months) | Irradiation Doses (kJ m−2) | TP Content (g kg−1) | Flavonoids Content (g kg−1) | Tannins Content (g kg−1) |
|---|---|---|---|---|
| 0 | - | 0.274 ± 0.014 | 0.034 ± 0.003 | 0.109 ± 0.010 |
| 1 | 0 | 0.288 ± 0.027 a | 0.035 ± 0.003 | 0.131 ± 0.003 a |
| 1 | 0.469 ± 0.007 b | 0.038 ± 0.004 | 0.299 ± 0.003 b | |
| 3 | 0.453 ± 0.009 b | 0.046 ± 0.003 | 0.370 ± 0.002 c | |
| 6 | 0.537 ± 0.014 c | 0.039 ± 0.002 | 0.283 ± 0.020 b | |
| 2 | 0 | 0.274 ± 0.021 a | 0.027 ± 0.003 | 0.134 ± 0.019 a |
| 1 | 0.306 ± 0.014 a | 0.030 ± 0.004 | 0.251 ± 0.011 b | |
| 3 | 0.339 ± 0.018 b | 0.032 ± 0.003 | 0.272 ± 0.011 c | |
| 6 | 0.345 ± 0.018 b | 0.035 ± 0.001 | 0.274 ± 0.015 c | |
| 3 | 0 | 0.308 ± 0.025 a | 0.027 ± 0.003 | 0.141 ± 0.015 a |
| 1 | 0.317 ± 0.013 a | 0.027 ± 0.004 | 0.149 ± 0.016 a | |
| 3 | 0.388 ± 0.012 b | 0.034 ± 0.005 | 0.236 ± 0.005 b | |
| 6 | 0.400 ± 0.006 b | 0.038 ± 0.003 | 0.238 ± 0.003 b | |
| 4 | 0 | 0.329 ± 0.010 a | 0.030 ± 0.002 a | 0.147 ± 0.023 a |
| 1 | 0.366 ± 0.020 a | 0.050 ± 0.004 b | 0.144 ± 0.013 a | |
| 3 | 0.364 ± 0.030 a | 0.045 ± 0.003 b | 0.162 ± 0.010 b | |
| 6 | 0.437 ± 0.012 b | 0.046 ± 0.002 b | 0.164 ± 0.006 b | |
| 5 | 0 | 0.337 ± 0.013 a | 0.032 ± 0.005 a | 0.139 ± 0.008 a |
| 1 | 0.411 ± 0.020 b | 0.045 ± 0.003 b | 0.147 ± 0.014 a | |
| 3 | 0.409 ± 0.012 b | 0.041 ± 0.005 b | 0.164 ± 0.007 b | |
| 6 | 0.417 ± 0.036 b | 0.050 ± 0.006 b | 0.135 ± 0.019 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dassamiour, S.; Boujouraf, O.; Sraoui, L.; Bensaad, M.S.; Derardja, A.e.; Alsufyani, S.J.; Sami, R.; Algarni, E.; Aljumayi, H.; Aljahani, A.H. Effect of Postharvest UV-C Radiation on Nutritional Quality, Oxidation and Enzymatic Browning of Stored Mature Date. Appl. Sci. 2022, 12, 4947. https://doi.org/10.3390/app12104947
Dassamiour S, Boujouraf O, Sraoui L, Bensaad MS, Derardja Ae, Alsufyani SJ, Sami R, Algarni E, Aljumayi H, Aljahani AH. Effect of Postharvest UV-C Radiation on Nutritional Quality, Oxidation and Enzymatic Browning of Stored Mature Date. Applied Sciences. 2022; 12(10):4947. https://doi.org/10.3390/app12104947
Chicago/Turabian StyleDassamiour, Saliha, Ourida Boujouraf, Linda Sraoui, Mohamed Sabri Bensaad, Ala eddine Derardja, Sultan J. Alsufyani, Rokayya Sami, Eman Algarni, Huda Aljumayi, and Amani H. Aljahani. 2022. "Effect of Postharvest UV-C Radiation on Nutritional Quality, Oxidation and Enzymatic Browning of Stored Mature Date" Applied Sciences 12, no. 10: 4947. https://doi.org/10.3390/app12104947
APA StyleDassamiour, S., Boujouraf, O., Sraoui, L., Bensaad, M. S., Derardja, A. e., Alsufyani, S. J., Sami, R., Algarni, E., Aljumayi, H., & Aljahani, A. H. (2022). Effect of Postharvest UV-C Radiation on Nutritional Quality, Oxidation and Enzymatic Browning of Stored Mature Date. Applied Sciences, 12(10), 4947. https://doi.org/10.3390/app12104947