
Non-Invasive Physical Plasma Enhances the Membrane Permeability to Low Molecular Weight Compounds and Subsequently Leads to the Loss of Cellular ATP and the Devitalization of Epithelial Cancer Cells

 ,
,  , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
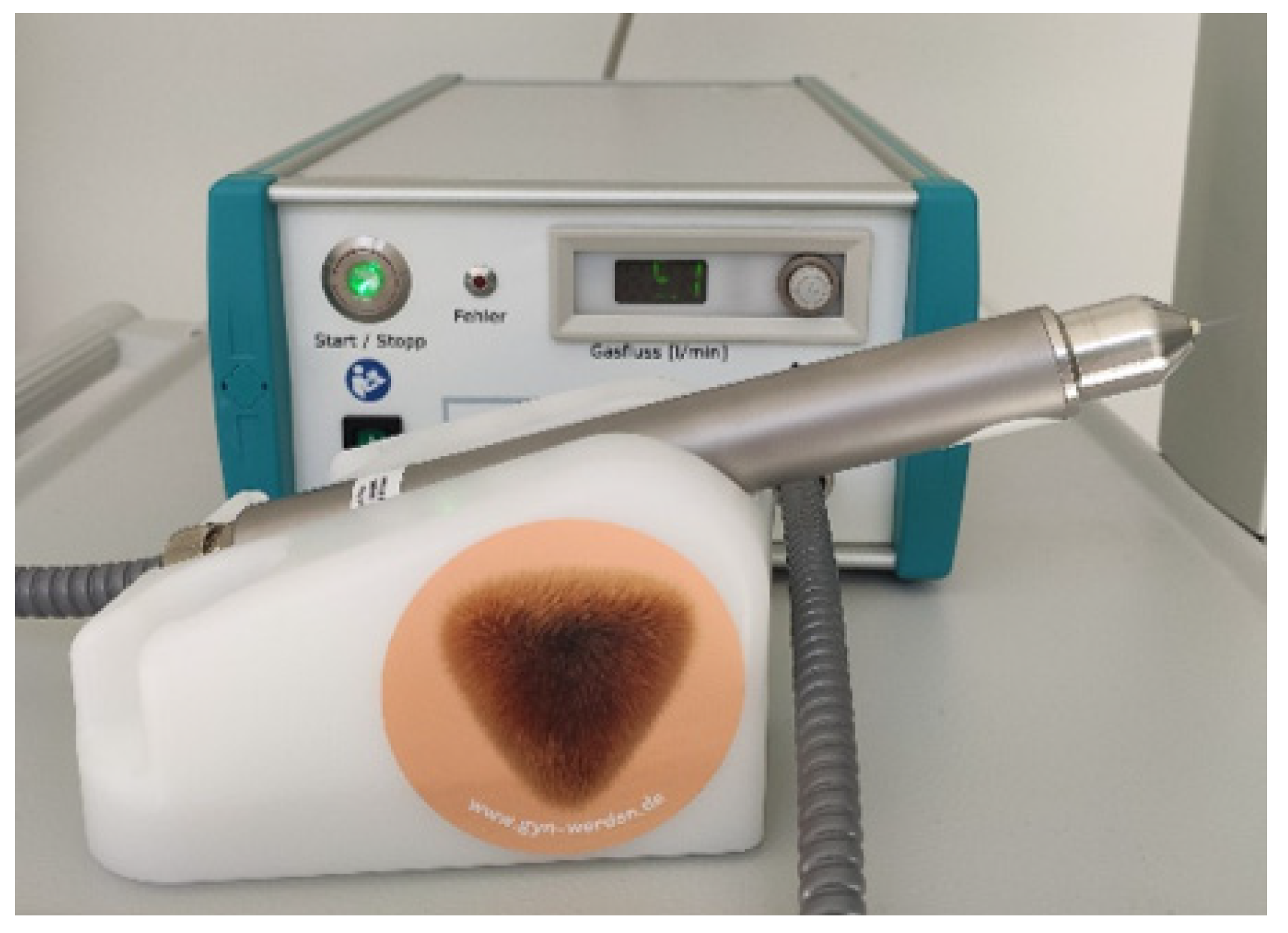
2.2. Non-Invasive Physical Plasma Treatment
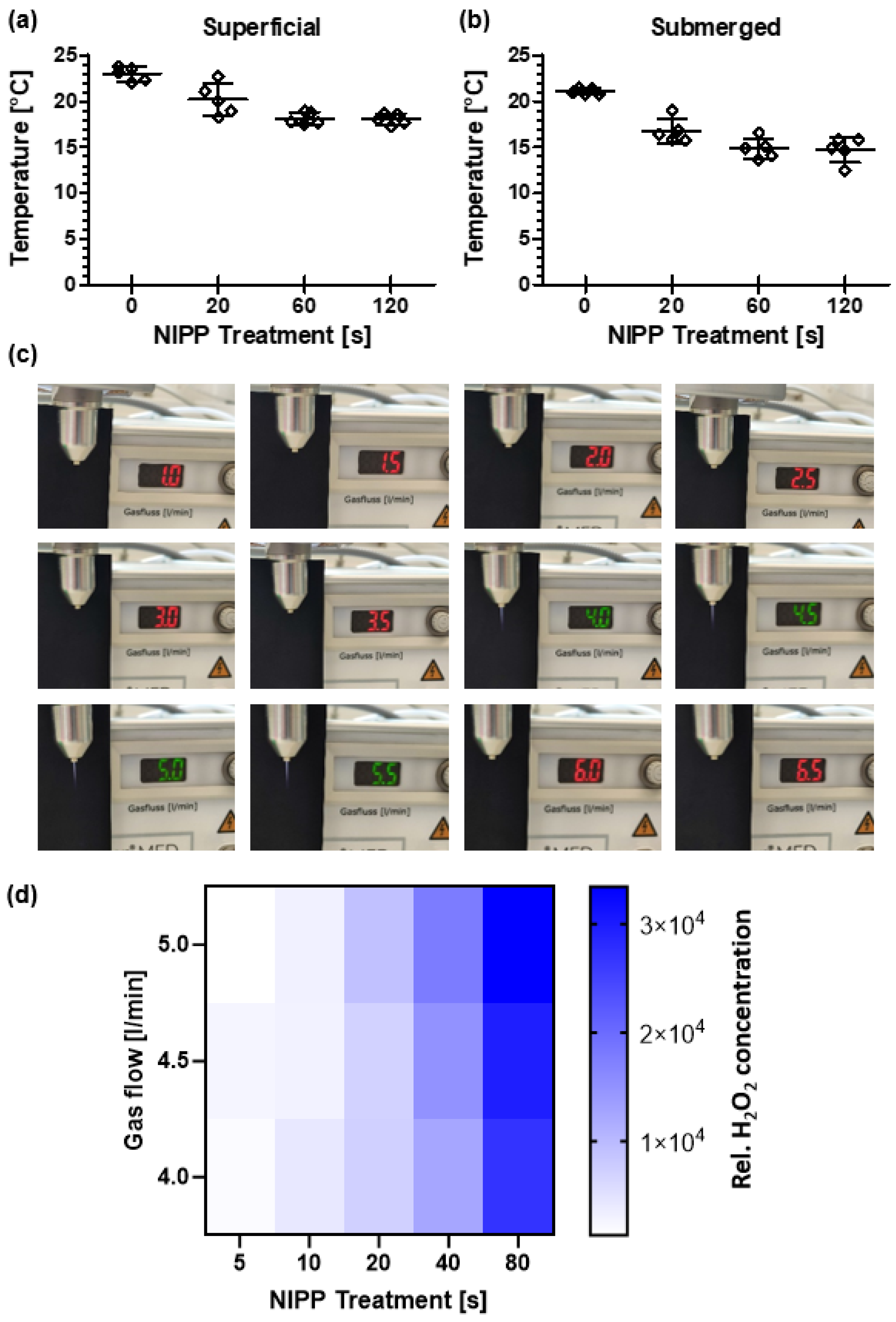
2.3. Temperature Measurement on Liquids
2.4. Detection of Plasma Ignition at Different Carrier Gas Flow Rates
2.5. Hydrogen Peroxide (H2O2) Assay
2.6. Microscopic Dextran-Fluorescein Isothiocyanate (FITC) Uptake Assay
2.7. Fluorescein Release Assay
2.8. ATP Release Assay
2.9. Caspase-3/7 (Casp-3/7) Apoptose Assay
2.10. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling Apoptose Assay
2.11. mRNA Analysis of the Pro-Apoptotic Regulators: Bcl-2 Antagonist of Cell Death (BAD) and Caspase-9 (Casp-9)
2.12. Cell Growth Assay
2.13. Statistics
3. Results
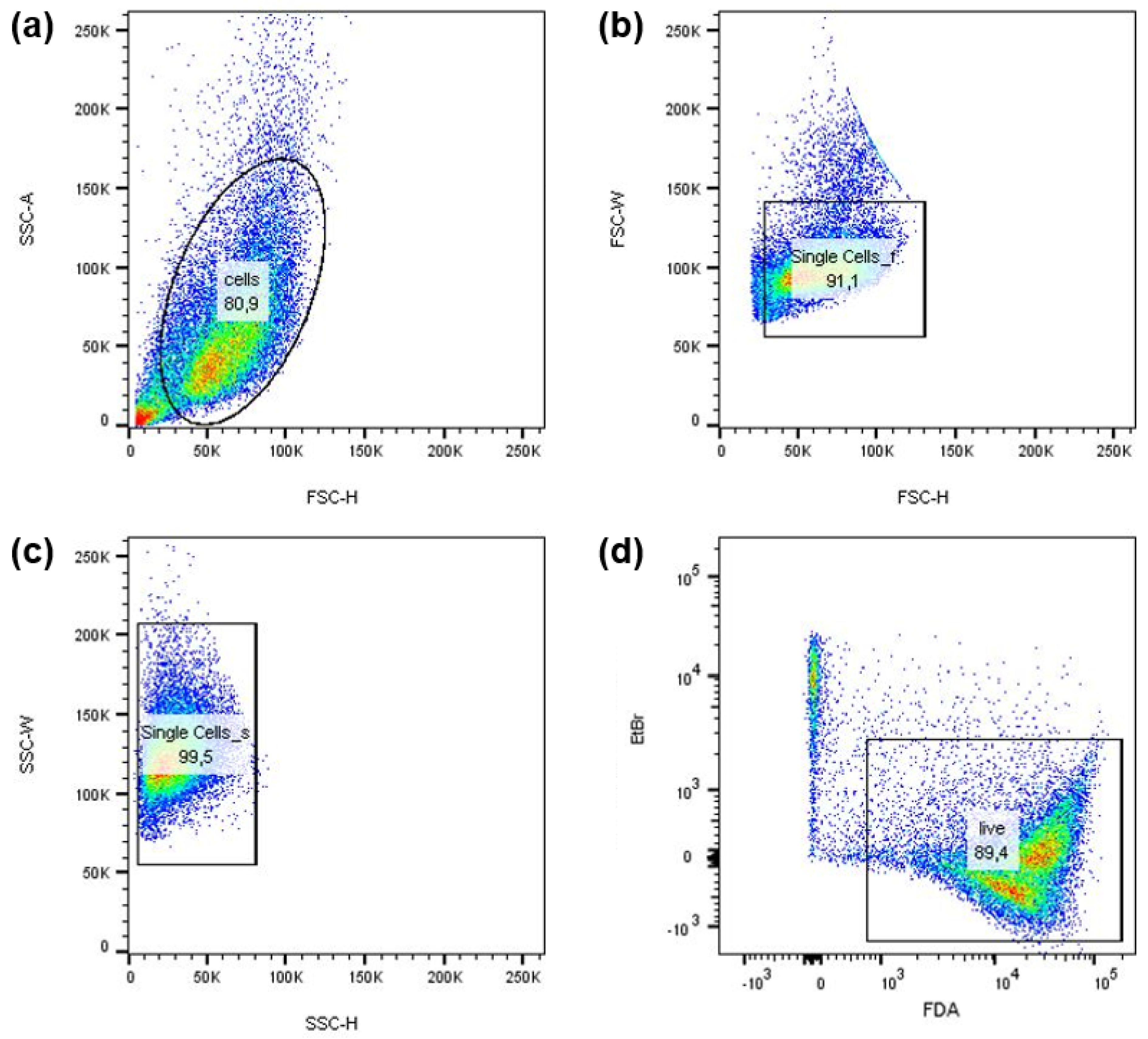
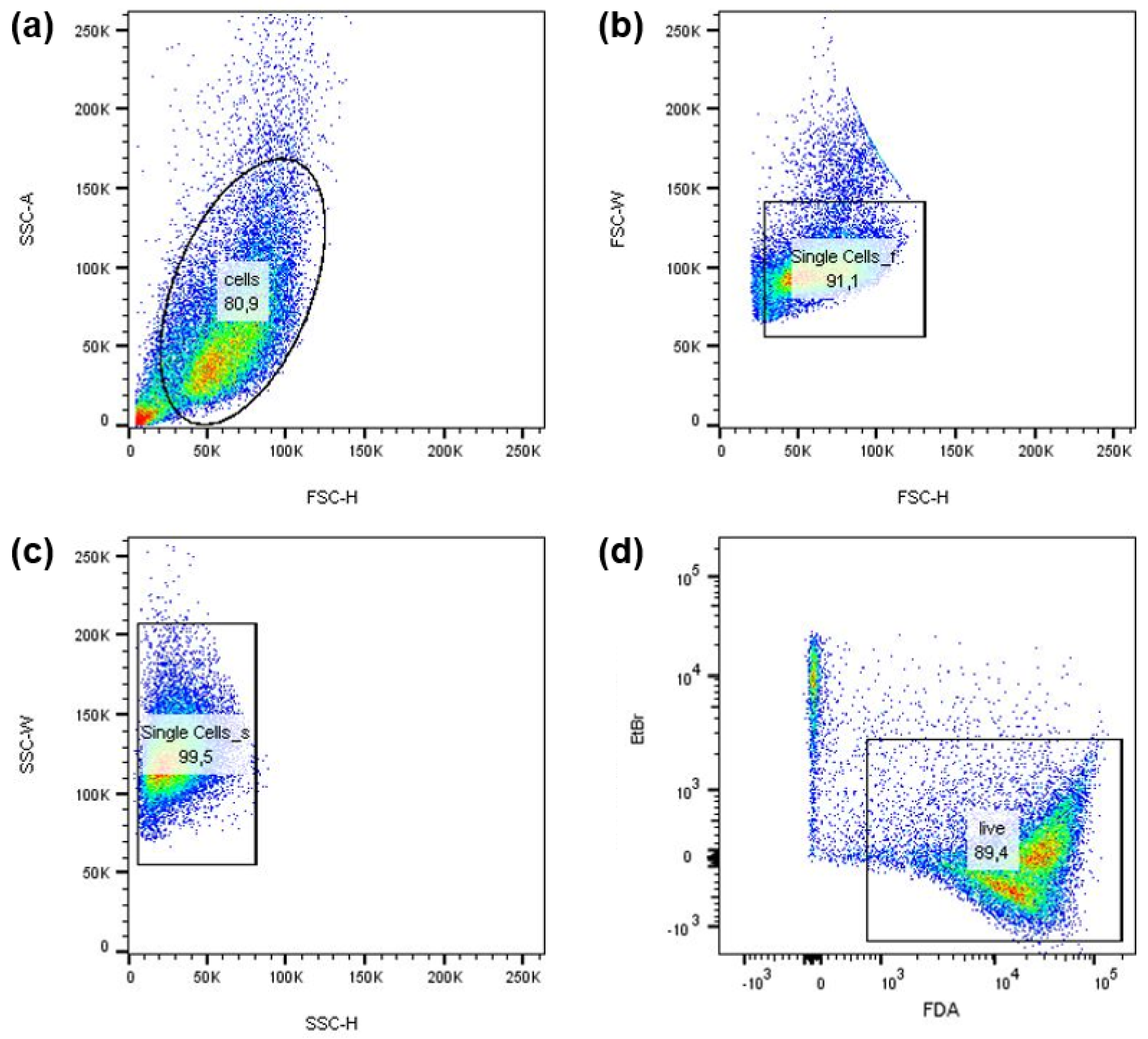
3.1. Basic Characterization of the NIPP Action in an In Vitro Cancer Cell Model
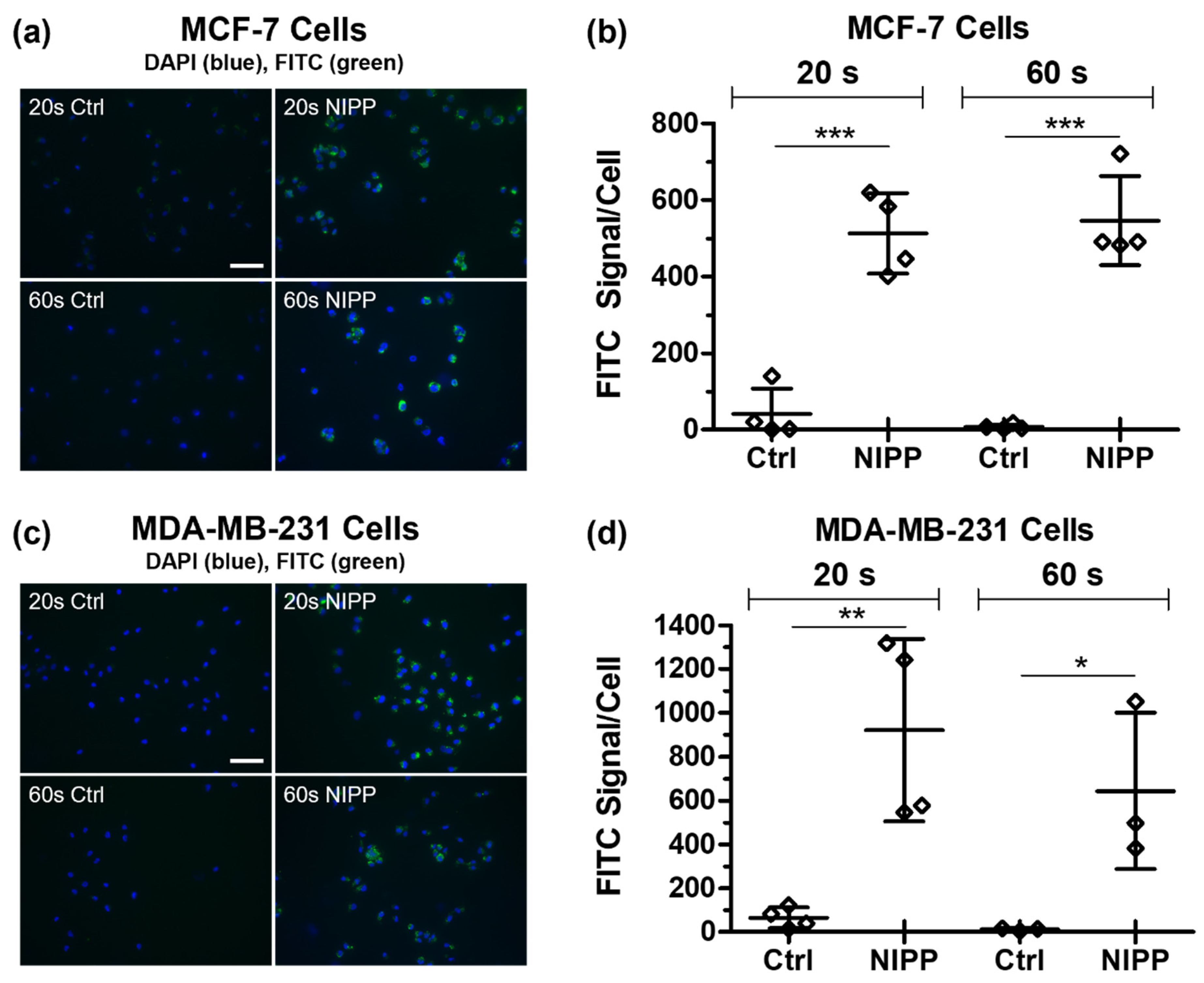
3.2. NIPP Affects the Cytoplasmic Membrane of Cancer Cells and Causes the Influx of FITC-Labeled Dextran
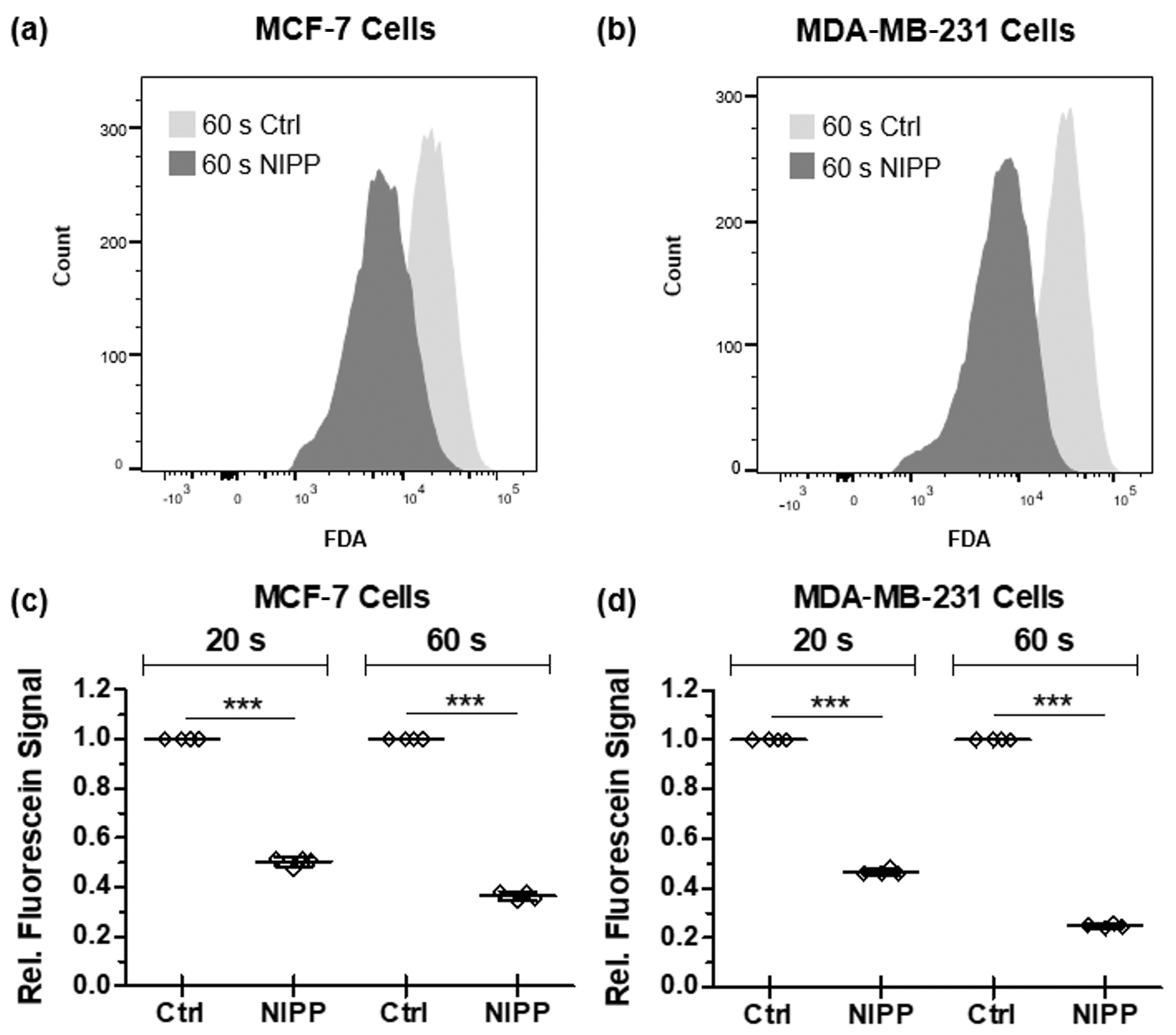
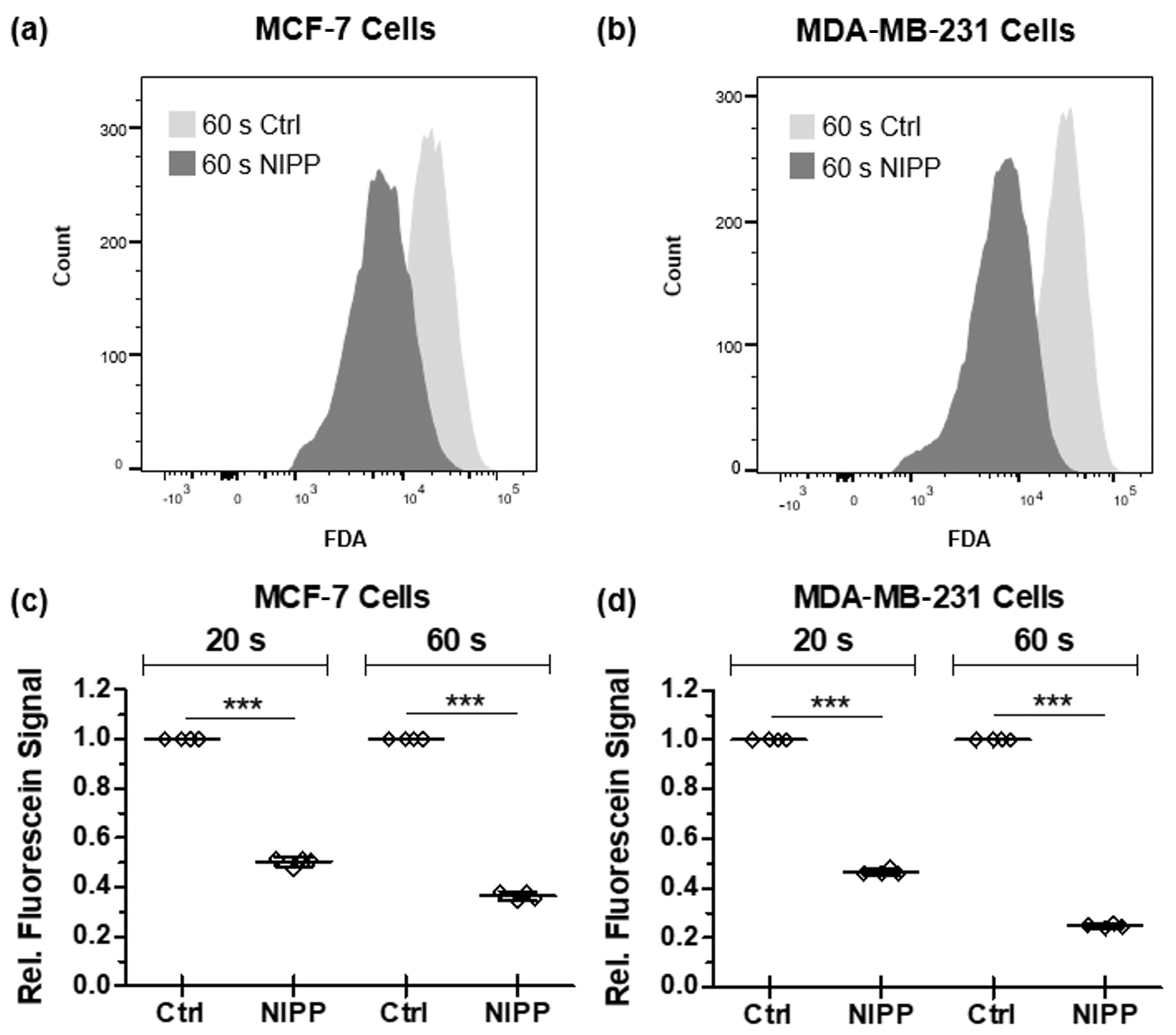
3.3. NIPP-Induced Alteration of the Cytoplasmic Membrane Enables the Efflux of Non-Membrane-Permeable Fluorescein
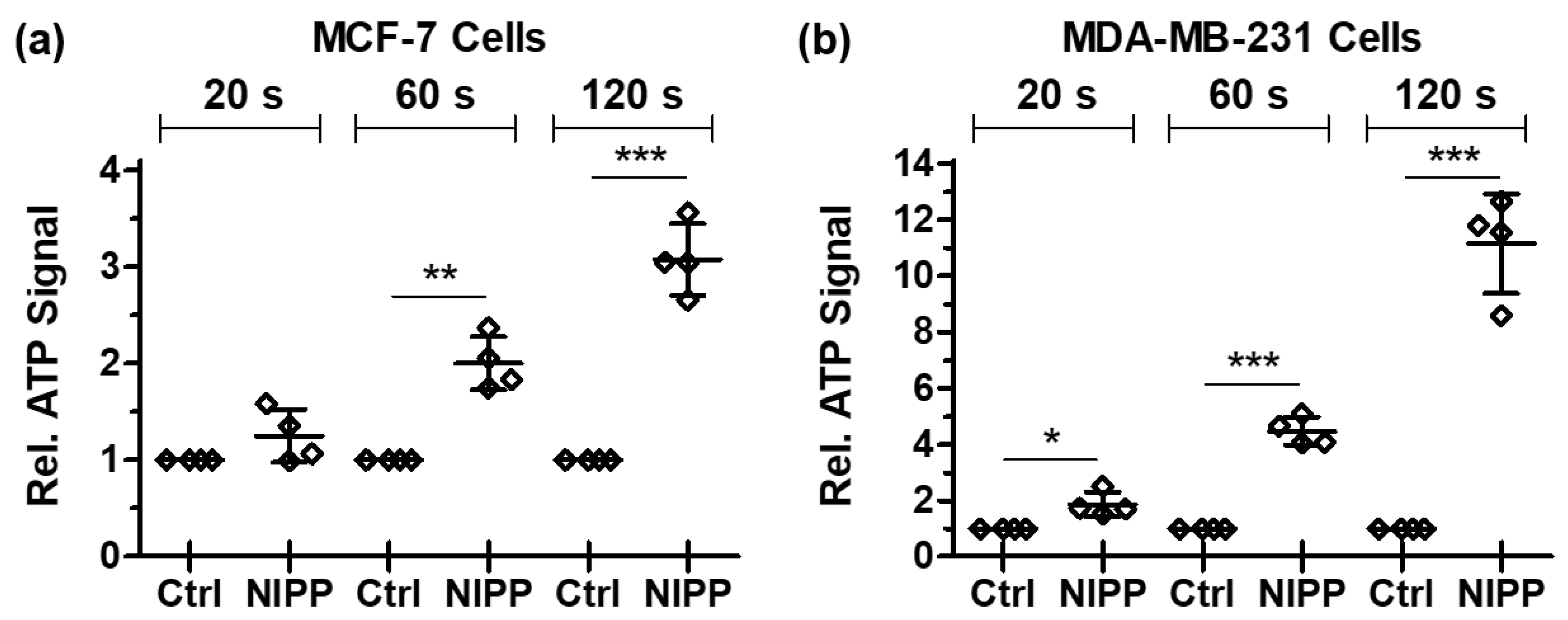
3.4. NIPP-Induced Alteration of the Cytoplasmic Membrane Enables the Efflux of Endogenous ATP Molecules
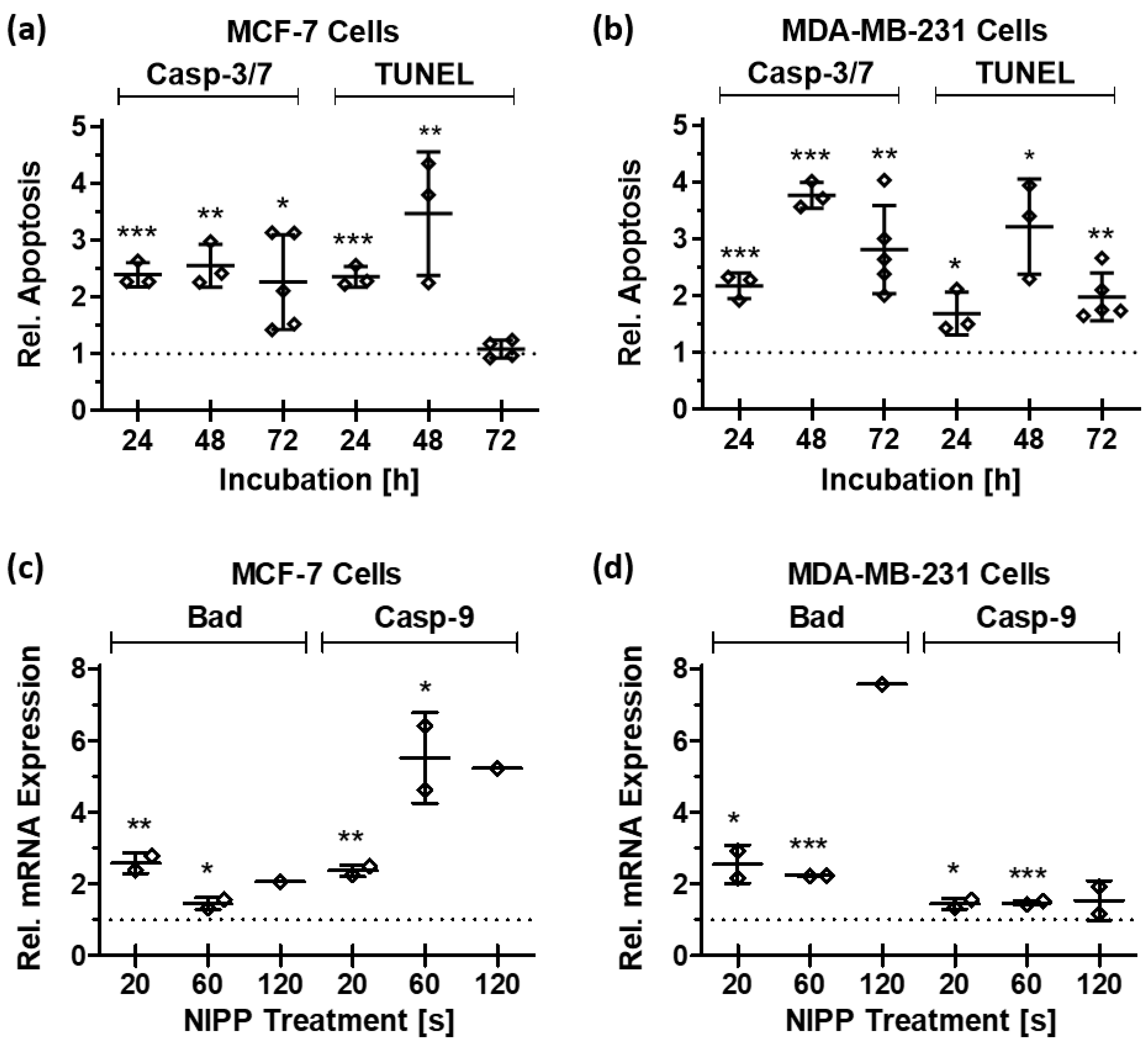
3.5. NIPP Treatment Leads to the Induction of the Apoptotic Machinery
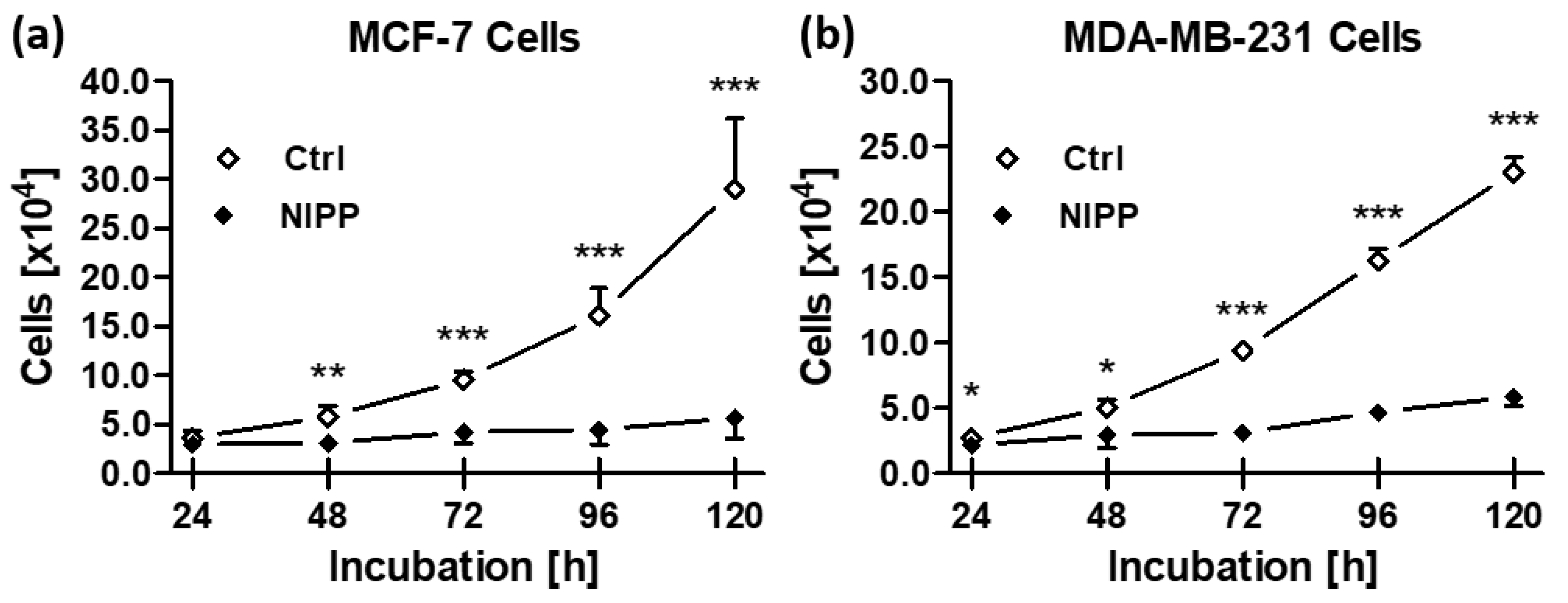
3.6. NIPP Efficacy on Cancer Cells Leads to Reduced Cell Growth
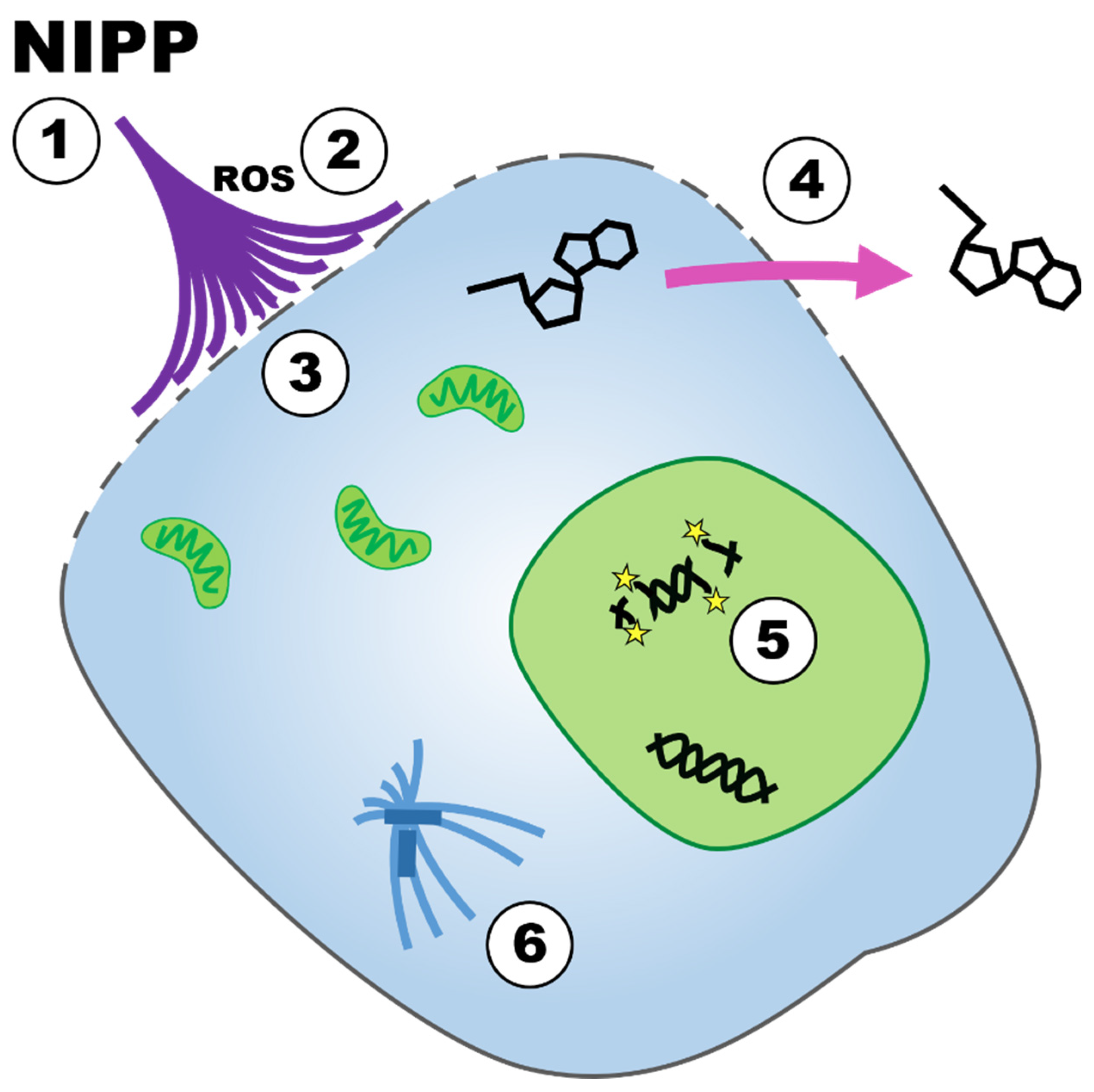
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kletschkus, K.; Haralambiev, L.; Mustea, A.; Bekeschus, S.; Stope, M.B. Review of innovative physical therapy methods: Introduction to the principles of cold physical plasma. In Vivo 2020, 34, 3103–3107. [Google Scholar] [CrossRef]
- Gan, L.; Jiang, J.; Duan, J.W.; Wu, X.J.Z.; Zhang, S.; Duan, X.R.; Song, J.Q.; Chen, H.X. Cold atmospheric plasma applications in dermatology: A systematic review. J. Biophotonics 2021, 14, e202000415. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Zhang, S.; Poorun, D.; Liu, D.; Lu, X.; He, M.; Duan, X.; Chen, H. Medical applications of nonthermal atmospheric pressure plasma in dermatology. J. Dtsch. Dermatol. Ges. 2018, 16, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Sherman, J.H.; Keidar, M. Cold atmospheric plasma, a novel promising anti-cancer treatment modality. Oncotarget 2017, 8, 15977–15995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stope, M.B. Plasma oncology—Physical plasma as innovative tumor therapy. J. Cancer Biol. 2020, 1, 53–56. [Google Scholar] [CrossRef]
- Jacoby, J.M.; Strakeljahn, S.; Nitsch, A.; Bekeschus, S.; Hinz, P.; Mustea, A.; Ekkernkamp, A.; Tzvetkov, M.V.; Haralambiev, L.; Stope, M.B. An innovative therapeutic option for the treatment of skeletal sarcomas: Elimination of osteo- and Ewing’s sarcoma cells using physical gas plasma. Int. J. Mol. Sci. 2020, 21, 4460. [Google Scholar] [CrossRef]
- Koensgen, D.; Besic, I.; Gümbel, D.; Kaul, A.; Weiss, M.; Diesing, K.; Kramer, A.; Bekeschus, S.; Mustea, A.; Stope, M.B. Cold atmospheric plasma (CAP) and CAP-stimulated cell culture media suppress ovarian cancer cell growth—A putative treatment option in ovarian cancer therapy. Anticancer Res. 2017, 37, 6739–6744. [Google Scholar] [CrossRef]
- Weiss, M.; Gümbel, D.; Hanschmann, E.-M.; Mandelkow, R.; Gelbrich, N.; Zimmermann, U.; Walther, R.; Ekkernkamp, A.; Sckell, A.; Kramer, A.; et al. Cold atmospheric plasma treatment induces anti-proliferative effects in prostate cancer cells by redox and apoptotic signaling pathways. PLoS ONE 2015, 10, e0130350. [Google Scholar] [CrossRef]
- van Loenhout, J.; Flieswasser, T.; Freire Boullosa, L.; de Waele, J.; van Audenaerde, J.; Marcq, E.; Jacobs, J.; Lin, A.; Lion, E.; Dewitte, H.; et al. Cold atmospheric plasma-treated PBS eliminates immunosuppressive pancreatic Stellate cells and induces immunogenic cell death of pancreatic cancer cells. Cancers 2019, 11, 597. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Ballato, J.; Foy, P.; Hawkins, T.; Wei, Y.; Li, J.; Kim, S.-O. Apoptosis of lung carcinoma cells induced by a flexible optical fiber-based cold microplasma. Biosens. Bioelectron. 2011, 28, 333–338. [Google Scholar] [CrossRef]
- Hediger, M.A.; Clémençon, B.; Burrier, R.E.; Bruford, E.A. The ABCs of membrane transporters in health and disease (SLC series): Introduction. Mol. Asp. Med. 2013, 34, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.J.; Hinner, M.J. Getting across the cell membrane: An overview for small molecules, peptides, and proteins. Methods Mol. Biol. 2015, 1266, 29–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Striesow, J.; Lackmann, J.-W.; Ni, Z.; Wenske, S.; Weltmann, K.-D.; Fedorova, M.; Woedtke, T.; von Wende, K. Oxidative modification of skin lipids by cold atmospheric plasma (CAP): A standardizable approach using RP-LC/MS(2) and DI-ESI/MS(2). Chem. Phys. Lipids 2020, 226, 104786. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, Y.; Liu, D.; Liu, Z.; Chen, C.; Xu, R.; Tian, M.; Wang, X.; Chen, H.; Kong, M.G. Cold atmospheric-pressure plasma induces DNA-protein crosslinks through protein oxidation. Free Radic. Res. 2018, 52, 783–798. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Fisusi, F.A.; Akala, E.O. Drug combinations in breast cancer therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Ravandeh, M.; Kahlert, H.; Jablonowski, H.; Lackmann, J.-W.; Striesow, J.; Agmo Hernández, V.; Wende, K. A combination of electrochemistry and mass spectrometry to monitor the interaction of reactive species with supported lipid bilayers. Sci. Rep. 2020, 10, 18683. [Google Scholar] [CrossRef] [PubMed]
- Bourke, P.; Ziuzina, D.; Han, L.; Cullen, P.J.; Gilmore, B.F. Microbiological interactions with cold plasma. J. Appl. Microbiol. 2017, 123, 308–324. [Google Scholar] [CrossRef] [Green Version]
- Nitsch, A.; Haralambiev, L.; Einenkel, R.; Muzzio, D.O.; Zygmunt, M.T.; Ekkernkamp, A.; Burchardt, M.; Stope, M.B. Determination of in vitro membrane permeability by analysis of intracellular and extracellular fluorescein signals in renal cells. In Vivo 2019, 33, 1767–1771. [Google Scholar] [CrossRef] [Green Version]
- Miyata, Y.; Matsuo, T.; Sagara, Y.; Ohba, K.; Ohyama, K.; Sakai, H. A Mini-review of reactive oxygen species in urological cancer: Correlation with NADPH oxidases, angiogenesis, and apoptosis. Int. J. Mol. Sci. 2017, 18, 2214. [Google Scholar] [CrossRef]
- Haralambiev, L.; Bandyophadyay, A.; Suchy, B.; Weiss, M.; Kramer, A.; Bekeschus, S.; Ekkernkamp, A.; Mustea, A.; Kaderali, L.; Stope, M.B. Determination of Immediate vs. Kinetic growth retardation in physically plasma-treated cells by experimental and modelling data. Anticancer Res. 2020, 40, 3743–3749. [Google Scholar] [CrossRef]
- Dolphin, C.T.; Caldwell, J.; Smith, R.L. Depression of oxidative metabolism of aspirin in mice via an interferon-associated mechanism in relation to Reye’s syndrome. J. Pharm. Pharmacol. 1987, 39, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.S.; Huang, C.; Hsieh, F.-H.; Huff, H.; Duan, Y. Bacterial inactivation using a low-temperature atmospheric plasma brush sustained with argon gas. J. Biomed. Mater. Res. B Appl. Biomater. 2007, 80, 211–219. [Google Scholar] [CrossRef]
- Haralambiev, L.; Nitsch, A.; Jacoby, J.M.; Strakeljahn, S.; Bekeschus, S.; Mustea, A.; Ekkernkamp, A.; Stope, M.B. Cold atmospheric plasma treatment of chondrosarcoma cells affects proliferation and cell membrane permeability. Int. J. Mol. Sci. 2020, 21, 2291. [Google Scholar] [CrossRef] [Green Version]
- Haralambiev, L.; Nitsch, A.; Einenkel, R.; Muzzio, D.O.; Gelbrich, N.; Burchardt, M.; Zygmunt, M.; Ekkernkamp, A.; Stope, M.B.; Gümbel, D. The effect of cold atmospheric plasma on the membrane permeability of human osteosarcoma cells. Anticancer Res. 2020, 40, 841–846. [Google Scholar] [CrossRef]
- Dezest, M.; Chavatte, L.; Bourdens, M.; Quinton, D.; Camus, M.; Garrigues, L.; Descargues, P.; Arbault, S.; Burlet-Schiltz, O.; Casteilla, L.; et al. Mechanistic insights into the impact of cold atmospheric pressure plasma on human epithelial cell lines. Sci. Rep. 2017, 7, 41163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, T.; Carvajal Berrio, D.A.; Daum, R.; Reisenauer, C.; Weltmann, K.D.; Wallwiener, D.; Brucker, S.Y.; Schenke-Layland, K.; Brauchle, E.M.; Weiss, M. Molecular effects and tissue penetration depth of physical plasma in human mucosa analyzed by contact- and marker-independent Raman microspectroscopy. ACS Appl. Mater. Interfaces 2019, 11, 42885–42895. [Google Scholar] [CrossRef] [PubMed]
- Miller, V.; Lin, A.; Fridman, A. Why target immune cells for plasma treatment of cancer. Plasma Chem. Plasma Process 2016, 36, 259–268. [Google Scholar] [CrossRef]
- Azzariti, A.; Iacobazzi, R.M.; Di Fonte, R.; Porcelli, L.; Gristina, R.; Favia, P.; Fracassi, F.; Trizio, I.; Silvestris, N.; Guida, G.; et al. Plasma-activated medium triggers cell death and the presentation of immune activating danger signals in melanoma and pancreatic cancer cells. Sci. Rep. 2019, 9, 4099. [Google Scholar] [CrossRef]
- Kepp, O.; Senovilla, L.; Vitale, I.; Vacchelli, E.; Adjemian, S.; Agostinis, P.; Apetoh, L.; Aranda, F.; Barnaba, V.; Bloy, N.; et al. Consensus guidelines for the detection of immunogenic cell death. Oncoimmunology 2014, 3, 955691. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Truong, B.; Patel, S.; Kaushik, N.; Choi, E.H.; Fridman, G.; Fridman, A.; Miller, V. Nanosecond-pulsed DBD plasma-generated reactive oxygen species trigger immunogenic cell death in A549 lung carcinoma cells through intracellular oxidative stress. Int. J. Mol. Sci. 2017, 18, 966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, M.R.; Chekeni, F.B.; Trampont, P.C.; Lazarowski, E.R.; Kadl, A.; Walk, S.F.; Park, D.; Woodson, R.L.; Ostankovich, M.; Sharma, P.; et al. Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic clearance. Nature 2009, 461, 282–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Wang, B.; Xu, Y.; Chen, Z.; Cui, Q.; Yang, Y.; Chen, H.; Kong, M.G. Intracellular ROS mediates gas plasma-facilitated cellular transfection in 2D and 3D cultures. Sci. Rep. 2016, 6, 27872. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Naidis, G.V.; Laroussi, M.; Reuter, S.; Graves, D.B.; Ostrikov, K. Reactive species in non-equilibrium atmospheric-pressure plasmas: Generation, transport, and biological effects. Phys. Rep. 2016, 630, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Park, H.J.; Yang, S.S.; Choi, K.S.; Lee, J.-S. Anti-cancer efficacy of nonthermal plasma dissolved in a liquid, liquid plasma in heterogeneous cancer cells. Sci. Rep. 2016, 6, 29020. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, S.; Levchenko, I.; Beilis, I.I.; Keidar, M. In vitro demonstration of cancer inhibiting properties from stratified self-organized plasma-liquid interface. Sci. Rep. 2017, 7, 12163. [Google Scholar] [CrossRef]
- Ninomiya, K.; Ishijima, T.; Imamura, M.; Yamahara, T.; Enomoto, H.; Takahashi, K.; Tanaka, Y.; Uesugi, Y.; Shimizu, N. Evaluation of extra- and intracellular OH radical generation, cancer cell injury, and apoptosis induced by a non-thermal atmospheric-pressure plasma jet. J. Phys. D Appl. Phys. 2013, 46, 425401. [Google Scholar] [CrossRef]
- Mitra, S.; Nguyen, L.N.; Akter, M.; Park, G.; Choi, E.H.; Kaushik, N.K. Impact of ROS generated by chemical, physical, and llasma techniques on cancer attenuation. Cancers 2019, 11, 1030. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Lin, L.; Ding, C.B.; Jin, T.; Han, X.H.; Zhou, H.; Wu, Z.W.; Pan, Y.Y. A meaningful attempt: Applying dielectric barrier discharge plasma to induce apoptosis of MDA-MB-231 cells via regulating HIF-1α/VEGFA expression. Surf. Coat. Technol. 2020, 41, 126293. [Google Scholar] [CrossRef]
- Adachi, T.; Tanaka, H.; Nonomura, S.; Hara, H.; Kondo, S.-I.; Hori, M. Plasma-activated medium induces A549 cell injury via a spiral apoptotic cascade involving the mitochondrial-nuclear network. Free Radic. Biol. Med. 2015, 79, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, H.; Bae, H.; Choi, E.H.; Kim, S.J. Epigenetic silencing of miR-19a-3p by cold atmospheric plasma contributes to proliferation inhibition of the MCF-7 breast cancer cell. Sci. Rep. 2016, 6, 30005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.B.; Kim, B.; Bae, H.; Lee, H.; Lee, S.; Choi, E.H.; Kim, S.J. Differential epigenetic effects of atmospheric cold plasma on MCF-7 and MDA-MB-231 breast cancer cells. PLoS ONE 2015, 10, e0129931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalghatgi, S.; Kelly, C.M.; Cerchar, E.; Torabi, B.; Alekseev, O.; Fridman, A.; Friedman, G.; Azizkhan-Clifford, J. Effects of non-thermal plasma on mammalian cells. PLoS ONE 2011, 6, e16270. [Google Scholar] [CrossRef] [Green Version]
- Kalghatgi, S.; Fridman, A.; Azizkhan-Clifford, J.; Friedman, G. DNA damage in mammalian cells by non-thermal atmospheric pressure microsecond pulsed dielectric barrier discharge plasma is not mediated by ozone. Plasma Process. Polym. 2012, 9, 726–732. [Google Scholar] [CrossRef]
- Schuster, M.; Rutkowski, R.; Hauschild, A.; Shojaei, R.K.; Woedtke, T.; Rana, A.; Bauer, G.; Metelmann, P.; Seebauer, C. Side effects in cold plasma treatment of advanced oral cancer—Clinical data and biological interpretation. Clin Plasma Med. 2018, 10, 9–15. [Google Scholar] [CrossRef]
- Weiss, M.; Stope, M.B. Physical plasma: A new treatment option in gynecological oncology. Arch. Gynecol. Obstet. 2018, 298, 853–855. [Google Scholar] [CrossRef]
- Stope, M.B.; Benouahi, R.; Sander, C.; Haralambiev, L.; Nitsch, A.; Egger, E.; Mustea, A. Protherapeutic effects and inactivation of mammary carcinoma cells by a medical argon plasma device. Anticancer Res. 2020, 40, 6205–6212. [Google Scholar] [CrossRef]
- Wenzel, T.; Carvajal Berrio, D.A.; Reisenauer, C.; Layland, S.; Koch, A.; Wallwiener, D.; Brucker, S.Y.; Schenke-Layland, K.; Brauchle, E.-M.; Weiss, M. Trans-mucosal efficacy of non-thermal plasma treatment on cervical cancer tissue and human cervix uteri by a next generation electrosurgical argon plasma device. Cancers 2020, 12, 267. [Google Scholar] [CrossRef] [Green Version]
- Feil, L.; Koch, A.; Utz, R.; Ackermann, M.; Barz, J.; Stope, M.; Krämer, B.; Wallwiener, D.; Brucker, S.Y.; Weiss, M. Cancer-selective treatment of cancerous and non-cancerous human cervical cell models by a non-thermally operated electrosurgical argon plasma device. Cancers 2020, 12, 1037. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Mizuno, M.; Ishikawa, K.; Toyokuni, S.; Kajiyama, H.; Kikkawa, F.; Hori, M. New hopes for plasma-based cancer treatment. Plasma 2018, 155. [Google Scholar] [CrossRef] [Green Version]
- Sagwal, S.K.; Pasqual-Melo, G.; Bodnar, Y.; Gandhirajan, R.K.; Bekeschus, S. Combination of chemotherapy and physical plasma elicits melanoma cell death via upregulation of SLC22A16. Cell Death Dis. 2018, 9, 1179. [Google Scholar] [CrossRef] [PubMed]
- Daeschlein, G.; Hillmann, A.; Gümbel, D.; Sicher, C.; Podewils, S.; Stope, M.B.; Jünger, M. Enhanced anticancer efficacy by drug chemotherapy and cold atmospheric plasma against melanoma and glioblastoma cell lines in vitro. IEEE Trans. Radiat. Plasma Med. Sci. 2018, 2, 153–159. [Google Scholar] [CrossRef]
- Tempfer, C.B.; Hilal, Z.; Dogan, A.; Petersen, M.; Rezniczek, G.A. Concentrations of cisplatin and doxorubicin in ascites and peritoneal tumor nodules before and after pressurized intraperitoneal aerosol chemotherapy (PIPAC) in patients with peritoneal metastasis. Eur. J. Surg. Oncol. 2018, 44, 1112–1117. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Assay | Neg Ctrl | Pos Ctrl | Ctrl | NIPP | ||||
|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | ||||
| MCF-7 | Casp-3/7 | 1.5 × 10−1 ±9.8 × 10−2 | 2.1 × 100 ±3.2 × 10−1 | 1.9 × 10−1 ±1.2 × 10−2 | 2.5 × 10−1 ±4.2 × 10−2 | 2.6 × 10−1 ±1.7 × 10−1 | 4.4 × 10−1 ±4.6 × 10−2 | 5.9 × 10−1 ±1.4 × 10−1 | 5.9 × 10−1 ±3.5 × 10−1 |
| TUNEL | 1.5 × 10−5 ±1.0 × 10−5 | 8.4 × 10−5 ±8.8 × 10−5 | 8.2 × 10−6 ±2.6 × 10−6 | 9.6 × 10−6 ±6.2 × 10−6 | 1.9 × 10−5 ±6.3 × 10−6 | 1.7 × 10−5 ±4.2 × 10−6 | 2.2 × 10−5 ±8.6 × 10−6 | 3.2 × 10−5 ±9.9 × 10−6 | |
| MDA-MB-231 | Casp-3/7 | 8.3 × 10−2 ±5.2 × 10−2 | 1.4 × 100 ±6.8 × 10−1 | 2.8 × 10−1 ±9.4 × 10−2 | 3.2 × 10−1 ±1.3 × 10−1 | 2.3 × 10−1 ±5.6 × 10−2 | 5.7 × 10−1 ±1.3 × 10−1 | 9.7 × 10−1 ±3.9 × 10−1 | 6.1 × 10−1 ±1.0 × 10−1 |
| TUNEL | 1.9 × 10−6 ±1.2 × 10−6 | 2.3 × 10−5 ±2.5 × 10−5 | 4.1 × 10−6 ±1.6 × 10−6 | 4.3 × 10−6 ±2.2 × 10−6 | 5.4 × 10−6 ±7.5 × 10−7 | 6.1 × 10−6 ±2.7 × 10−6 | 1.1 × 10−5 ±3.7 × 10−6 | 1.0 × 10−5 ±3.5 × 10−6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sander, C.; Nitsch, A.; Erb, H.H.H.; Egger, E.K.; Haralambiev, L.; Eggers, B.; Kramer, F.-J.; Weiss, M.; Mustea, A.; Stope, M.B. Non-Invasive Physical Plasma Enhances the Membrane Permeability to Low Molecular Weight Compounds and Subsequently Leads to the Loss of Cellular ATP and the Devitalization of Epithelial Cancer Cells. Appl. Sci. 2021, 11, 9801. https://doi.org/10.3390/app11219801
Sander C, Nitsch A, Erb HHH, Egger EK, Haralambiev L, Eggers B, Kramer F-J, Weiss M, Mustea A, Stope MB. Non-Invasive Physical Plasma Enhances the Membrane Permeability to Low Molecular Weight Compounds and Subsequently Leads to the Loss of Cellular ATP and the Devitalization of Epithelial Cancer Cells. Applied Sciences. 2021; 11(21):9801. https://doi.org/10.3390/app11219801
Chicago/Turabian StyleSander, Caroline, Andreas Nitsch, Holger H. H. Erb, Eva K. Egger, Lyubomir Haralambiev, Benedikt Eggers, Franz-Josef Kramer, Martin Weiss, Alexander Mustea, and Matthias B. Stope. 2021. "Non-Invasive Physical Plasma Enhances the Membrane Permeability to Low Molecular Weight Compounds and Subsequently Leads to the Loss of Cellular ATP and the Devitalization of Epithelial Cancer Cells" Applied Sciences 11, no. 21: 9801. https://doi.org/10.3390/app11219801
APA StyleSander, C., Nitsch, A., Erb, H. H. H., Egger, E. K., Haralambiev, L., Eggers, B., Kramer, F.-J., Weiss, M., Mustea, A., & Stope, M. B. (2021). Non-Invasive Physical Plasma Enhances the Membrane Permeability to Low Molecular Weight Compounds and Subsequently Leads to the Loss of Cellular ATP and the Devitalization of Epithelial Cancer Cells. Applied Sciences, 11(21), 9801. https://doi.org/10.3390/app11219801