
Targeted Protein Profiling of In Vivo NIPP-Treated Tissues Using DigiWest Technology

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro and In Vivo NIPP Treatment
2.2. Propagation of Cells
2.3. Protein Extraction from FFPE Tissue
2.4. Lysis of Cell Culture Pellets
2.5. Multiplex Protein Profiling via DigiWest
2.6. Statistical Analysis
3. Results
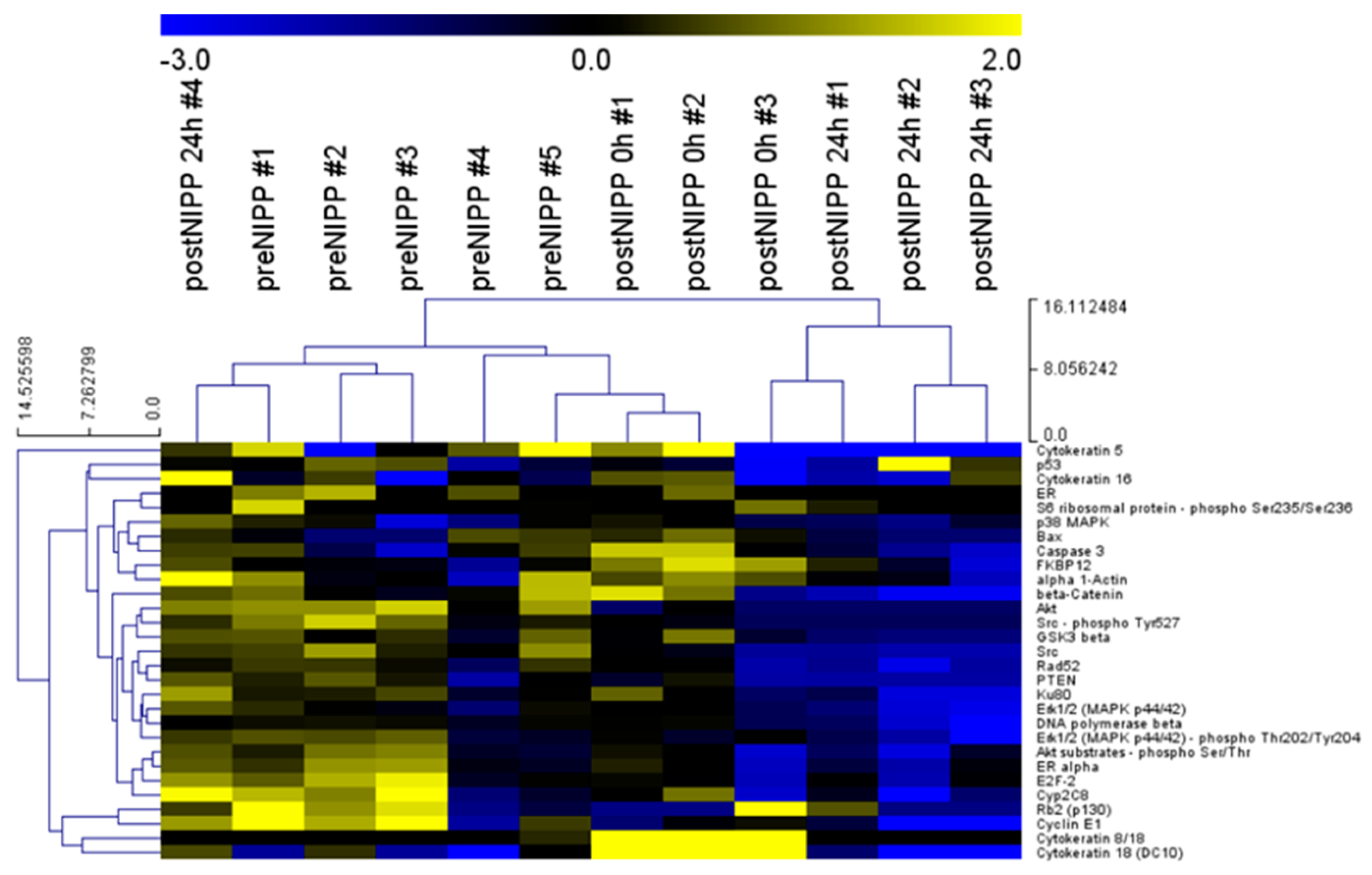
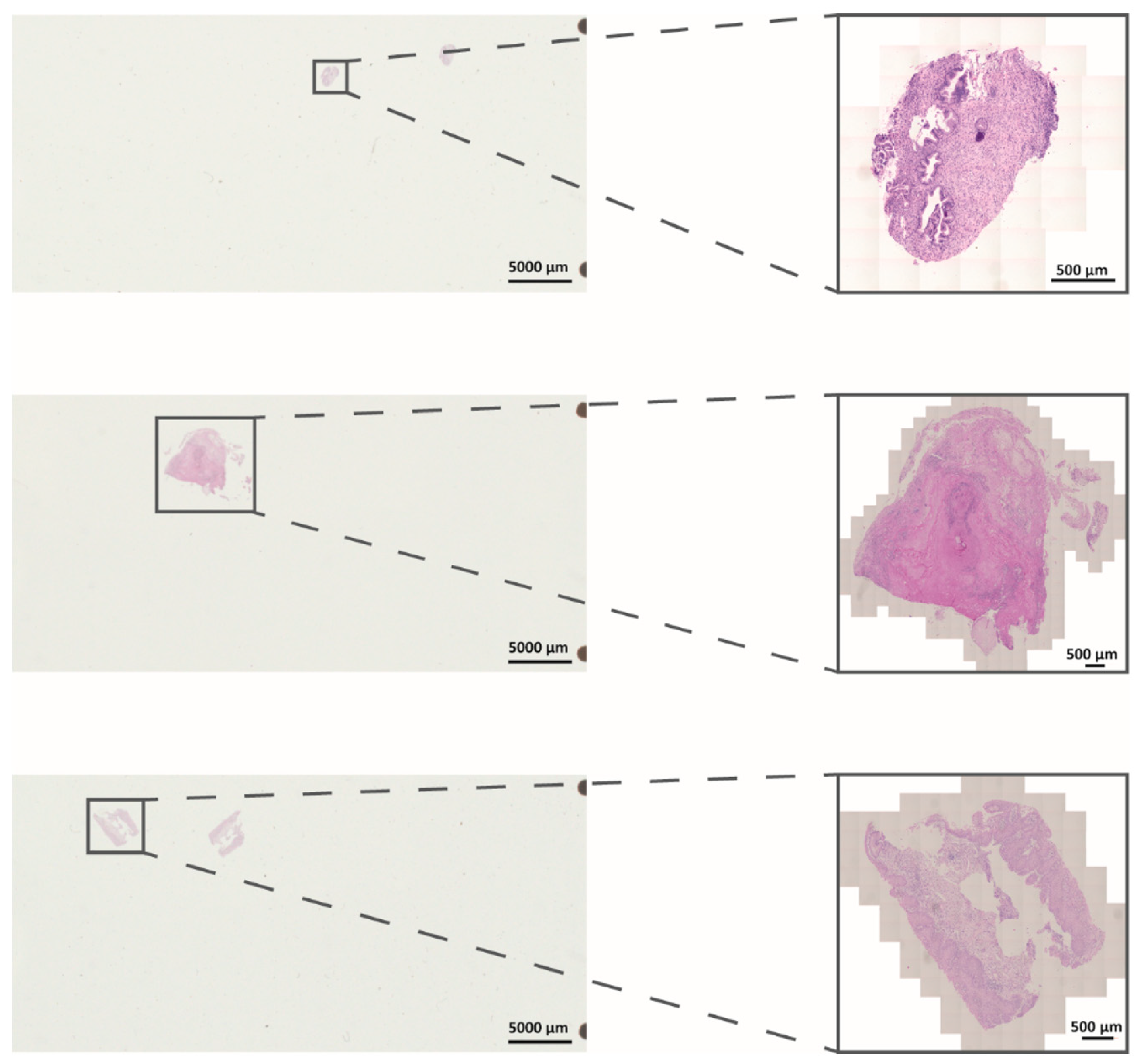
3.1. Establishment and Evaluation of DigiWest from FFPE Samples after In Vivo NIPP Treatment
3.2. Protein Profiles of Patients and Cell Culture following In Vivo and In Vitro NIPP Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirst, A.M.; Frame, F.M.; Arya, M.; Maitland, N.J.; O’Connell, D. Low temperature plasmas as emerging cancer therapeutics: The state of play and thoughts for the future. Tumour Biol. 2016, 37, 7021–7031. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Nguyen, L.N.; Akter, M.; Park, G.; Choi, E.H.; Kaushik, N.K. Impact of ros generated by chemical, physical, and plasma techniques on cancer attenuation. Cancers 2019, 11, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koensgen, D.; Besic, I.; Gumbel, D.; Kaul, A.; Weiss, M.; Diesing, K.; Kramer, A.; Bekeschus, S.; Mustea, A.; Stope, M.B. Cold atmospheric plasma (cap) and cap-stimulated cell culture media suppress ovarian cancer cell growth—A putative treatment option in ovarian cancer therapy. Anticancer Res. 2017, 37, 6739–6744. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Gumbel, D.; Gelbrich, N.; Brandenburg, L.O.; Mandelkow, R.; Zimmermann, U.; Ziegler, P.; Burchardt, M.; Stope, M.B. Inhibition of cell growth of the prostate cancer cell model lncap by cold atmospheric plasma. Vivo 2015, 29, 611–616. [Google Scholar]
- Weiss, M.; Gumbel, D.; Hanschmann, E.M.; Mandelkow, R.; Gelbrich, N.; Zimmermann, U.; Walther, R.; Ekkernkamp, A.; Sckell, A.; Kramer, A.; et al. Cold atmospheric plasma treatment induces anti-proliferative effects in prostate cancer cells by redox and apoptotic signaling pathways. PLoS ONE 2015, 10, e0130350. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, T.; Carvajal Berrio, D.A.; Daum, R.; Reisenauer, C.; Weltmann, K.D.; Wallwiener, D.; Brucker, S.Y.; Schenke-Layland, K.; Brauchle, E.M.; Weiss, M. Molecular effects and tissue penetration depth of physical plasma in human mucosa analyzed by contact- and marker-independent raman microspectroscopy. ACS Appl. Mater. Interfaces 2019, 11, 42885–42895. [Google Scholar] [CrossRef]
- Weiss, M.; Barz, J.; Ackermann, M.; Utz, R.; Ghoul, A.; Weltmann, K.D.; Stope, M.B.; Wallwiener, D.; Schenke-Layland, K.; Oehr, C.; et al. Dose-dependent tissue-level characterization of a medical atmospheric pressure argon plasma jet. ACS Appl. Mater. Interfaces 2019, 11, 19841–19853. [Google Scholar] [CrossRef]
- Braný, D.; Dvorská, D.; Halašová, E.Š.H. Cold atmospheric plasma: A powerful tool for modern medicine. Int. J. Mol. Sci. 2020, 21, 2932. [Google Scholar] [CrossRef] [Green Version]
- Weiss, M.; Utz, R.; Ackermann, M.; Taran, F.; Krämer, B.; Hahn, M.; Wallwiener, D.; Brucker, S.; Haupt, M.; Barz, J.; et al. Characterization of a non-thermally operated electrosurgical argon plasma source by electron spin resonance spectroscopy. Plasma Process. Polym. 2019, 16, 1800150. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Peto, J.; Gilham, C.; Deacon, J.; Taylor, C.; Evans, C.; Binns, W.; Haywood, M.; Elanko, N.; Coleman, D.; Yule, R.; et al. Cervical hpv infection and neoplasia in a large population-based prospective study: The manchester cohort. Br. J. Cancer 2004, 91, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Kyrgiou, M.; Athanasiou, A.; Kalliala, I.E.J.; Paraskevaidi, M.; Mitra, A.; Martin-Hirsch, P.P.; Arbyn, M.; Bennett, P.; Paraskevaidis, E. Obstetric outcomes after conservative treatment for cervical intraepithelial lesions and early invasive disease. Cochrane Database Syst Rev. 2017, 11, CD012847. [Google Scholar] [CrossRef]
- Sadler, L.; Saftlas, A.; Wang, W.; Exeter, M.; Whittaker, J.; McCowan, L. Treatment for cervical intraepithelial neoplasia and risk of preterm delivery. JAMA 2004, 291, 2100–2106. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, T.; Carvajal Berrio, D.A.; Reisenauer, C.; Layland, S.; Koch, A.; Wallwiener, D.; Brucker, S.Y.; Schenke-Layland, K.; Brauchle, E.M.; Weiss, M. Trans-mucosal efficacy of non-thermal plasma treatment on cervical cancer tissue and human cervix uteri by a next generation electrosurgical argon plasma device. Cancers 2020, 12, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specht, K.; Richter, T.; Muller, U.; Walch, A.; Werner, M.; Hofler, H. Quantitative gene expression analysis in microdissected archival formalin-fixed and paraffin-embedded tumor tissue. Am. J. Pathol. 2001, 158, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Matyunina, L.V.; Walker, L.D.; Chen, W.; Xiao, H.; Benigno, B.B.; Wu, R.; McDonald, J.F. Evidence for the importance of post-transcriptional regulatory changes in ovarian cancer progression and the contribution of mirnas. Sci. Rep. 2017, 7, 8171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bockmayr, T.; Erdmann, G.; Treue, D.; Jurmeister, P.; Schneider, J.; Arndt, A.; Heim, D.; Bockmayr, M.; Sachse, C.; Klauschen, F. Multiclass cancer classification in fresh frozen and formalin-fixed paraffin-embedded tissue by digiwest multiplex protein analysis. Lab. Investig. 2020, 100, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Treindl, F.; Ruprecht, B.; Beiter, Y.; Schultz, S.; Dottinger, A.; Staebler, A.; Joos, T.O.; Kling, S.; Poetz, O.; Fehm, T.; et al. A bead-based western for high-throughput cellular signal transduction analyses. Nat. Commun. 2016, 7, 12852. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. Tm4 microarray software suite. Methods Enzym. 2006, 411, 134–193. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; App, H.; Zhang, X.F.; Banerjee, P.; Brautigan, D.L.; Rapp, U.R.; Avruch, J. Raf-1 activates map kinase-kinase. Nature 1992, 358, 417–421. [Google Scholar] [CrossRef]
- Shaul, Y.D.; Seger, R. The mek/erk cascade: From signaling specificity to diverse functions. Biochim. Biophys. Acta 2007, 1773, 1213–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeyrathna, P.; Su, Y. The critical role of akt in cardiovascular function. Vasc. Pharm. 2015, 74, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Ram, P.T.; Iyengar, R. G protein coupled receptor signaling through the src and stat3 pathway: Role in proliferation and transformation. Oncogene 2001, 20, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.L.; Guan, Y.J.; Wang, L.; Wei, W.; Kane, A.B.; Chin, Y.E. Central role of the threonine residue within the p+1 loop of receptor tyrosine kinase in stat3 constitutive phosphorylation in metastatic cancer cells. Mol. Cell Biol. 2004, 24, 9390–9400. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. P21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Adami, G.R.; Wei, N.; Keyomarsi, K.; Elledge, S.J. The p21 cdk-interacting protein cip1 is a potent inhibitor of g1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- Weng, Y.R.; Cui, Y.; Fang, J.Y. Biological functions of cytokeratin 18 in cancer. Mol. Cancer Res. 2012, 10, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dowbenko, D.; Lasky, L.A. Akt/pkb phosphorylation of p21cip/waf1 enhances protein stability of p21cip/waf1 and promotes cell survival. J. Biol. Chem. 2002, 277, 11352–11361. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, K.M.; Anderson, N.G. The protein kinase b/akt signalling pathway in human malignancy. Cell Signal. 2002, 14, 381–395. [Google Scholar] [CrossRef]
- Feil, L.; Koch, A.; Utz, R.; Ackermann, M.; Barz, J.; Stope, M.; Kramer, B.; Wallwiener, D.; Brucker, S.Y.; Weiss, M. Cancer-selective treatment of cancerous and non-cancerous human cervical cell models by a non-thermally operated electrosurgical argon plasma device. Cancers 2020, 12, 1037. [Google Scholar] [CrossRef] [Green Version]
- Bayer, M.; Angenendt, L.; Schliemann, C.; Hartmann, W.; Konig, S. Are formalin-fixed and paraffin-embedded tissues fit for proteomic analysis? J. Mass Spectrom. 2020, 55, e4347. [Google Scholar] [CrossRef]
- Dubuc, A.; Monsarrat, P.; Virard, F.; Merbahi, N.; Sarrette, J.P.; Laurencin-Dalicieux, S.; Cousty, S. Use of cold-atmospheric plasma in oncology: A concise systematic review. Adv. Med. Oncol. 2018, 10, 1758835918786475. [Google Scholar] [CrossRef]
- Cheng, Y.; Qin, K.; Huang, N.; Zhou, Z.; Xiong, H.; Zhao, J.; Zhang, Y.; Yu, S. Cytokeratin 18 regulates the transcription and alternative splicing of apoptoticrelated genes and pathways in hela cells. Oncol. Rep. 2019, 42, 301–312. [Google Scholar] [CrossRef]
- Sirnio, P.; Vayrynen, J.P.; Mutt, S.J.; Herzig, K.H.; Walkowiak, J.; Klintrup, K.; Makela, J.; Karttunen, T.J.; Makinen, M.J.; Tuomisto, A. Systemic inflammation is associated with circulating cell death released keratin 18 fragments in colorectal cancer. Oncoimmunology 2020, 9, 1783046. [Google Scholar] [CrossRef]
- Abraham, M.C.; Shaham, S. Death without caspases, caspases without death. Trends Cell Biol. 2004, 14, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Guo, Y.; Stacey, W.C.; Harwalkar, J.; Fretthold, J.; Hitomi, M.; Stacey, D.W. Glycogen synthase kinase 3 has a limited role in cell cycle regulation of cyclin d1 levels. BMC Cell Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Echalier, A.; Endicott, J.A.; Noble, M.E. Recent developments in cyclin-dependent kinase biochemical and structural studies. Biochim. Biophys. Acta 2010, 1804, 511–519. [Google Scholar] [CrossRef]
- Bartova, I.; Otyepka, M.; Kriz, Z.; Koca, J. Activation and inhibition of cyclin-dependent kinase-2 by phosphorylation; a molecular dynamics study reveals the functional importance of the glycine-rich loop. Protein. Sci. 2004, 13, 1449–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthet, C.; Aleem, E.; Coppola, V.; Tessarollo, L.; Kaldis, P. Cdk2 knockout mice are viable. Curr. Biol. 2003, 13, 1775–1785. [Google Scholar] [CrossRef] [Green Version]
- Dehm, S.M.; Bonham, K. Src gene expression in human cancer: The role of transcriptional activation. Biochem. Cell. Biol. 2004, 82, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Przystal, J.M.; Becker, H.; Canjuga, D.; Tsiami, F.; Anderle, N.; Keller, A.L.; Pohl, A.; Ries, C.H.; Schmittnaegel, M.; Korinetska, N.; et al. Targeting csf1r alone or in combination with pd1 in experimental glioma. Cancers 2021, 13, 2400. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruoff, F.; Henes, M.; Templin, M.; Enderle, M.; Bösmüller, H.; Wallwiener, D.; Brucker, S.Y.; Schenke-Layland, K.; Weiss, M. Targeted Protein Profiling of In Vivo NIPP-Treated Tissues Using DigiWest Technology. Appl. Sci. 2021, 11, 11238. https://doi.org/10.3390/app112311238
Ruoff F, Henes M, Templin M, Enderle M, Bösmüller H, Wallwiener D, Brucker SY, Schenke-Layland K, Weiss M. Targeted Protein Profiling of In Vivo NIPP-Treated Tissues Using DigiWest Technology. Applied Sciences. 2021; 11(23):11238. https://doi.org/10.3390/app112311238
Chicago/Turabian StyleRuoff, Felix, Melanie Henes, Markus Templin, Markus Enderle, Hans Bösmüller, Diethelm Wallwiener, Sara Y. Brucker, Katja Schenke-Layland, and Martin Weiss. 2021. "Targeted Protein Profiling of In Vivo NIPP-Treated Tissues Using DigiWest Technology" Applied Sciences 11, no. 23: 11238. https://doi.org/10.3390/app112311238
APA StyleRuoff, F., Henes, M., Templin, M., Enderle, M., Bösmüller, H., Wallwiener, D., Brucker, S. Y., Schenke-Layland, K., & Weiss, M. (2021). Targeted Protein Profiling of In Vivo NIPP-Treated Tissues Using DigiWest Technology. Applied Sciences, 11(23), 11238. https://doi.org/10.3390/app112311238