
Evaluation of Bacterial Diversity and Quality Features of Traditional Sichuan Bacon from Different Geographical Region

Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Physiochemical Characteristics of Sichuan Bacon
2.3. DNA Extraction and Amplicon Sequencing
2.4. Sequence Analyses
2.5. Statistical Analysis
3. Results
3.1. Physicochemical Parameters of Sichuan Bacon
3.2. Characteristics of the Sequencing Data
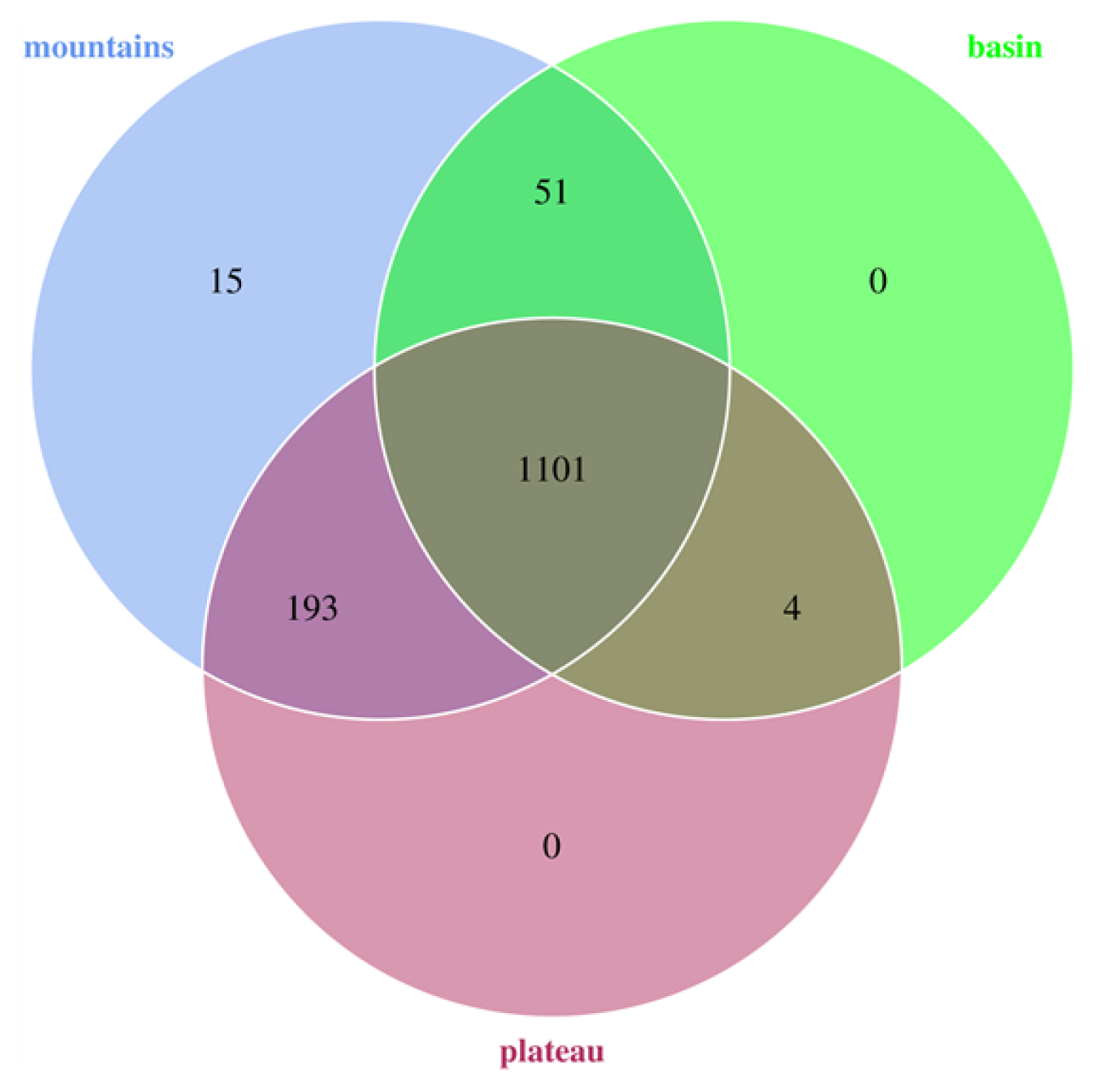
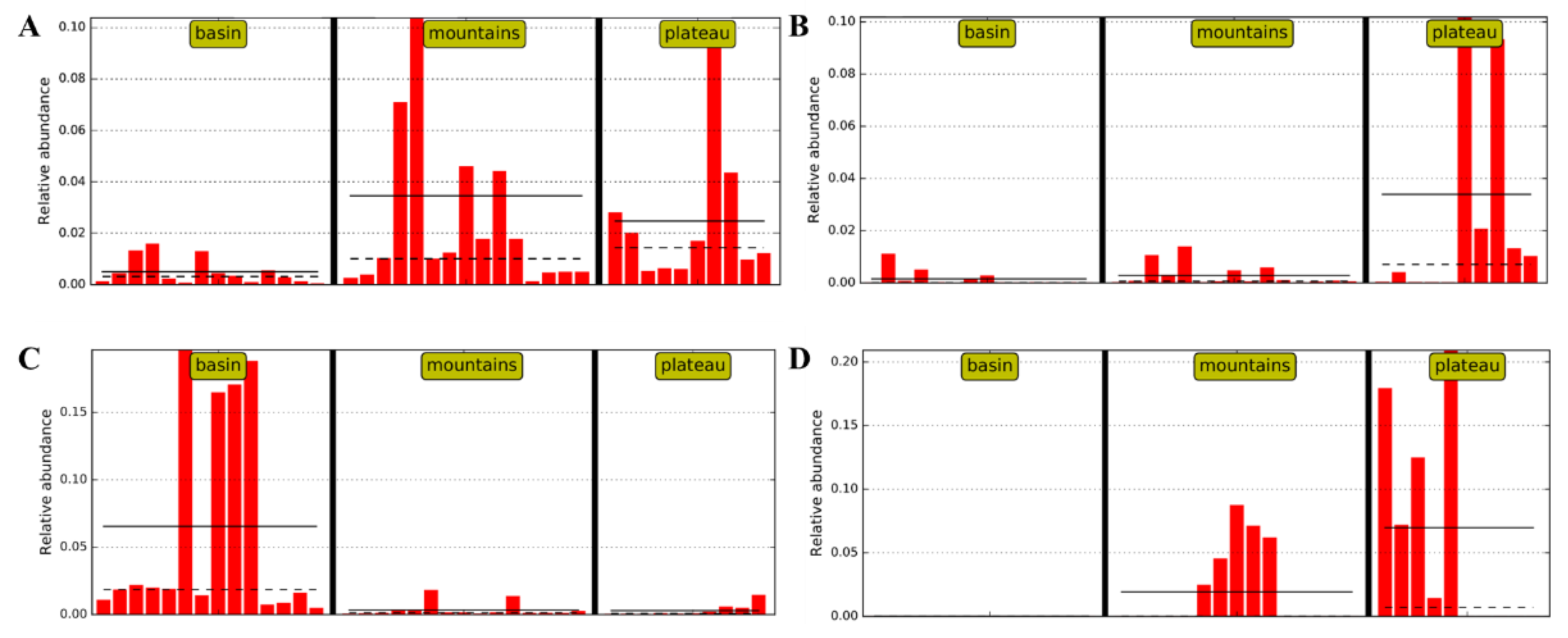
3.3. Bacterial Compositions of Sichuan Bacons
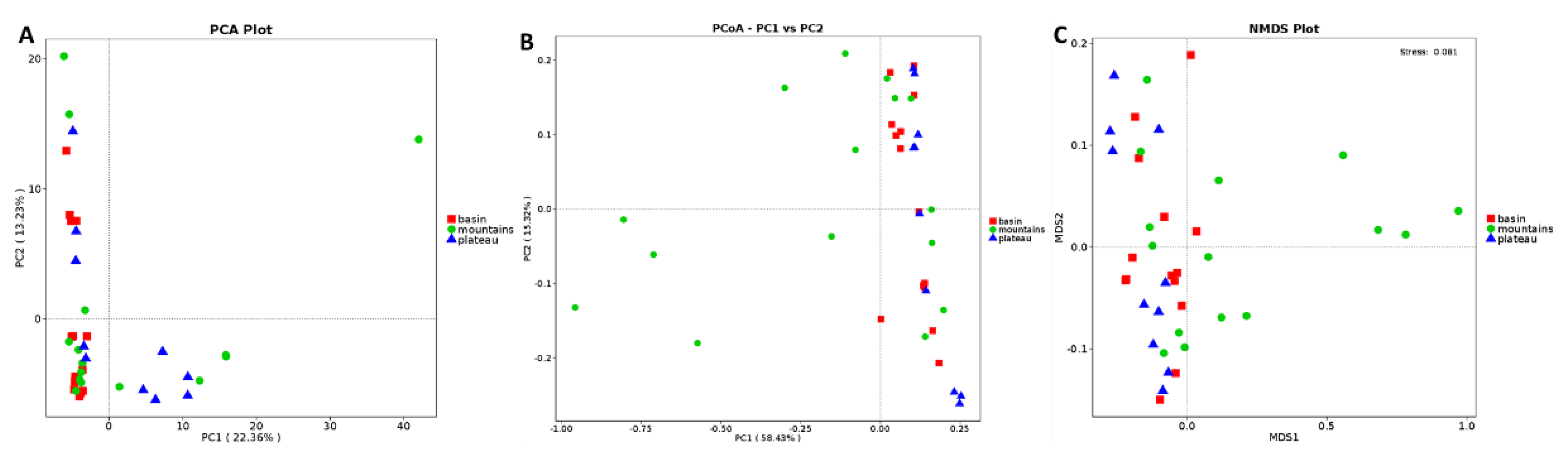
3.4. Analysis of Bacterial Communities of Sichuan Bacon in Different Geographical Regions
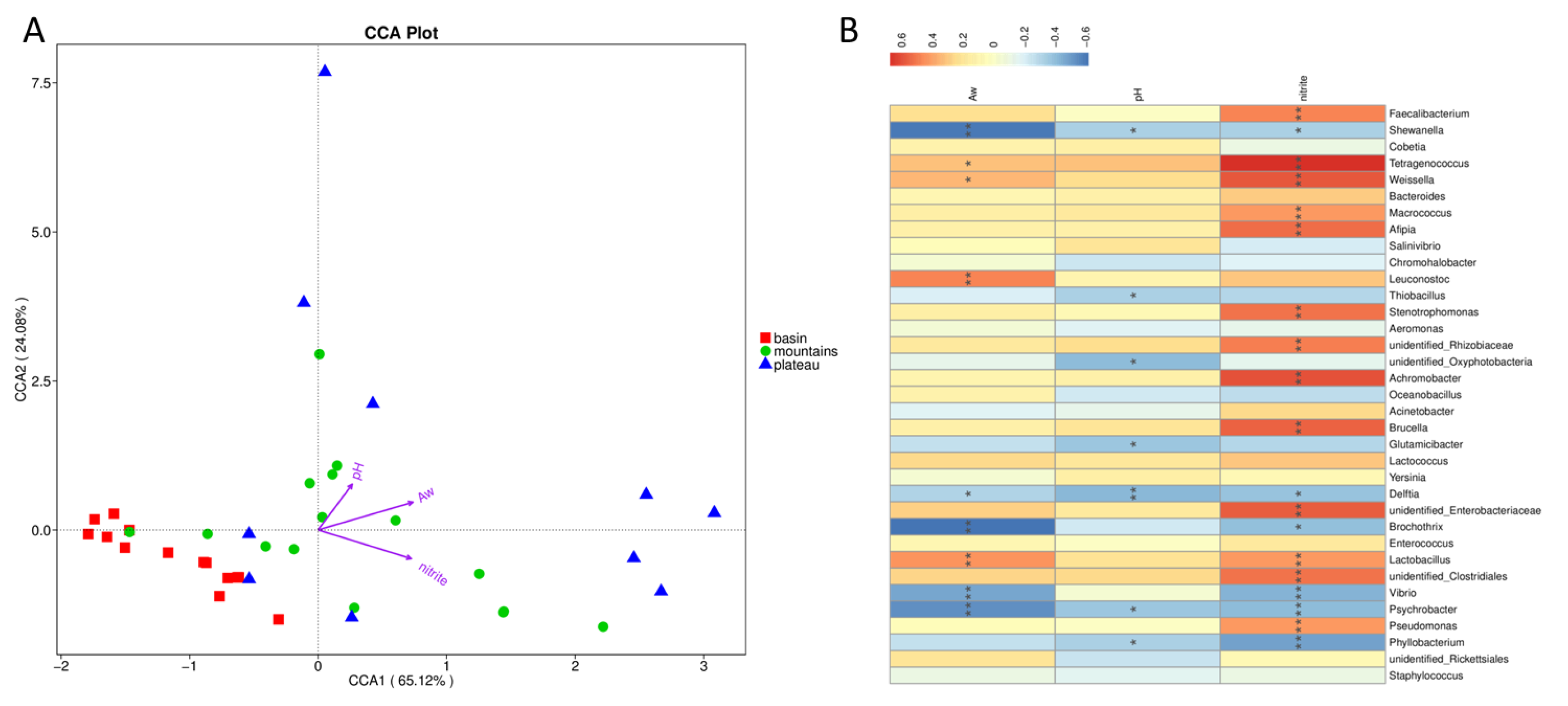
3.5. The Relationship between Top Genera and Environmental Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw PE (#) | Combined (#) | Qualified (#) | Nochime (#) | Avg Len (nt) | Effective % |
|---|---|---|---|---|---|---|
| BA1 | 98,835 | 96,773 | 95,663 | 93,267 | 252 | 94.37 |
| BA2 | 87,792 | 63,104 | 60,343 | 57,774 | 254 | 65.81 |
| BA3 | 86,549 | 84,950 | 83,974 | 80,927 | 252 | 93.5 |
| BA4 | 83,900 | 79,274 | 77,945 | 75,347 | 252 | 89.81 |
| BB1 | 84,898 | 82,385 | 81,136 | 73,781 | 253 | 86.91 |
| BB2 | 86,495 | 83,717 | 82,557 | 76,913 | 253 | 88.92 |
| BB3 | 95,010 | 90,558 | 89,135 | 84,040 | 253 | 88.45 |
| BB4 | 92,873 | 83,191 | 81,768 | 76,352 | 249 | 82.21 |
| BB5 | 87,069 | 75,142 | 73,213 | 70,254 | 254 | 80.69 |
| BC1 | 81,467 | 73,264 | 70,411 | 66,509 | 252 | 81.64 |
| BC2 | 98,199 | 94,404 | 93,045 | 87,076 | 253 | 88.67 |
| BC3 | 85,961 | 76,897 | 75,270 | 71,158 | 253 | 82.78 |
| BC4 | 94,842 | 80,522 | 77,919 | 72,685 | 253 | 76.64 |
| BC5 | 95,153 | 76,999 | 74,828 | 71,166 | 252 | 74.79 |
| MA1 | 82,378 | 76,064 | 74,786 | 72,287 | 249 | 87.75 |
| MA2 | 85,426 | 83,136 | 82,413 | 79,576 | 237 | 93.15 |
| MA3 | 97,461 | 88,505 | 87,206 | 85,441 | 234 | 87.67 |
| MA4 | 83,188 | 82,167 | 81,227 | 76,744 | 253 | 92.25 |
| MA5 | 92,712 | 87,305 | 85,919 | 82,609 | 253 | 89.1 |
| MB1 | 82,432 | 79,911 | 79,157 | 73,762 | 254 | 89.48 |
| MB2 | 89,052 | 84,857 | 83,635 | 79,450 | 254 | 89.22 |
| MB3 | 76,775 | 73,559 | 71,681 | 66,512 | 255 | 86.63 |
| MB4 | 88,555 | 85,598 | 83,973 | 80,118 | 250 | 90.47 |
| MB5 | 93,716 | 91,280 | 90,427 | 87,884 | 252 | 93.78 |
| MC1 | 92,442 | 87,084 | 85,654 | 79,627 | 252 | 86.14 |
| MC2 | 98,422 | 82,567 | 80,554 | 76,418 | 250 | 77.64 |
| MC3 | 84,825 | 80,792 | 79,890 | 77,608 | 245 | 91.49 |
| MC4 | 99,950 | 95,372 | 94,375 | 91,363 | 239 | 91.41 |
| MC5 | 90,170 | 88,416 | 87,902 | 86,234 | 229 | 95.63 |
| PA1 | 89,505 | 85,058 | 83,807 | 80,253 | 253 | 89.66 |
| PA2 | 96,874 | 88,771 | 86,815 | 81,797 | 253 | 84.44 |
| PA3 | 98,694 | 95,418 | 93,892 | 86,847 | 253 | 88 |
| PA4 | 86,633 | 82,189 | 80,884 | 76,942 | 253 | 88.81 |
| PA5 | 87,876 | 70,756 | 68,491 | 65,280 | 254 | 74.29 |
| PB1 | 80,545 | 77,321 | 76,581 | 74,041 | 253 | 91.93 |
| PB2 | 77,140 | 74,767 | 72,543 | 69,946 | 254 | 90.67 |
| PB3 | 88,085 | 84,479 | 82,851 | 77,546 | 253 | 88.04 |
| PB4 | 88,531 | 86,156 | 84,593 | 80,461 | 253 | 90.88 |
| PB5 | 76,757 | 74,594 | 73,210 | 69,141 | 254 | 90.08 |
| Sample | Observed Species | Shannon | Simpson | Chao1 | Ace | Good’s Coverage |
|---|---|---|---|---|---|---|
| BA1 | 346 | 1.5303 | 0.3083 | 357.413 | 368.0392 | 0.9991 |
| BA2 | 281 | 3.2806 | 0.7633 | 277.4737 | 279.2758 | 0.9998 |
| BA3 | 357 | 2.9900 | 0.7031 | 358.4685 | 385.1882 | 0.9993 |
| BA4 | 448 | 3.1576 | 0.7523 | 463.6195 | 475.4967 | 0.9994 |
| BB1 | 306 | 3.6041 | 0.8341 | 312.2462 | 334.4232 | 0.9996 |
| BB2 | 349 | 3.4318 | 0.8120 | 362.2581 | 367.6659 | 0.9994 |
| BB3 | 389 | 3.6992 | 0.8556 | 400.7222 | 419.8041 | 0.9993 |
| BB4 | 749 | 4.1670 | 0.8242 | 755.6667 | 769.4436 | 0.9988 |
| BB5 | 53 | 1.8340 | 0.6303 | 52.88235 | 58.51154 | 0.9999 |
| BC1 | 680 | 3.8438 | 0.7982 | 696.4286 | 708.2509 | 0.9991 |
| BC2 | 683 | 3.3852 | 0.6842 | 683.7653 | 710.7533 | 0.9991 |
| BC3 | 706 | 3.7436 | 0.7757 | 713.7213 | 725.2032 | 0.9993 |
| BC4 | 557 | 3.8612 | 0.7721 | 565.3404 | 577.2322 | 0.9993 |
| BC5 | 534 | 3.6825 | 0.8016 | 562.6241 | 567.1605 | 0.9989 |
| MA1 | 284 | 3.0673 | 0.7926 | 294.9014 | 294.3744 | 0.9995 |
| MA2 | 312 | 3.6688 | 0.8350 | 321.0851 | 327.6277 | 0.9993 |
| MA3 | 377 | 2.8811 | 0.6826 | 379.6726 | 375.6161 | 0.9994 |
| MA4 | 511 | 3.1646 | 0.7491 | 513.4286 | 540.6371 | 0.9992 |
| MA5 | 374 | 3.1296 | 0.7168 | 372.0435 | 387.8292 | 0.9994 |
| MB1 | 610 | 4.8808 | 0.8810 | 608.28 | 633.0211 | 0.9993 |
| MB2 | 594 | 4.5379 | 0.8608 | 609.1866 | 612.6939 | 0.9995 |
| MB3 | 740 | 6.3633 | 0.9614 | 740.6842 | 740.852 | 0.9996 |
| MB4 | 617 | 4.5869 | 0.8680 | 627.4681 | 640.6145 | 0.9994 |
| MB5 | 476 | 3.1935 | 0.6678 | 501.5 | 497.8253 | 0.9989 |
| MC1 | 706 | 4.3571 | 0.8640 | 705.52 | 725.3433 | 0.9992 |
| MC2 | 769 | 4.5736 | 0.8899 | 772.4904 | 792.9436 | 0.9991 |
| MC3 | 464 | 3.1463 | 0.7101 | 475.0074 | 485.7014 | 0.9992 |
| MC4 | 470 | 3.5864 | 0.8186 | 498.1161 | 513.0337 | 0.9986 |
| MC5 | 368 | 2.2450 | 0.5116 | 386.7835 | 419.8829 | 0.9991 |
| PA1 | 363 | 3.0520 | 0.6912 | 362.2222 | 375.9001 | 0.9995 |
| PA2 | 646 | 3.2985 | 0.7065 | 642.3623 | 664.7128 | 0.9991 |
| PA3 | 544 | 3.8095 | 0.8344 | 571.2261 | 579.1615 | 0.9990 |
| PA4 | 646 | 3.3790 | 0.7589 | 637.9827 | 669.531 | 0.9992 |
| PA5 | 382 | 3.7926 | 0.8719 | 398.703 | 414.029 | 0.9992 |
| PB1 | 544 | 4.1810 | 0.8650 | 544.175 | 563.6767 | 0.9993 |
| PB2 | 416 | 2.6434 | 0.6151 | 436.2581 | 435.0155 | 0.9993 |
| PB3 | 539 | 4.5015 | 0.9082 | 554.5118 | 559.1409 | 0.9992 |
| PB4 | 522 | 4.4449 | 0.8858 | 533.8261 | 533.6284 | 0.9994 |
| PB5 | 474 | 4.6674 | 0.9015 | 491.8583 | 493.4462 | 0.9993 |

References
- Wang, H.; Zhang, X.; Suo, H.; Zhao, X.; Kan, J. Aroma and flavor characteristics of commercial Chinese traditional bacon from different geographical regions. J. Sens. Stud. 2019, 34, e12475. [Google Scholar] [CrossRef]
- Huang, Y.; Li, H.; Huang, T.; Li, F.; Sun, J. Lipolysis and lipid oxidation during processing of Chinese traditional smoke-cured bacon. Food Chem. 2014, 149, 31–39. [Google Scholar] [CrossRef]
- Lu, S.; Ji, H.; Wang, Q.; Li, B.; Li, K.; Xu, C.; Jiang, C. The effects of starter cultures and plant extracts on the biogenic amine accumulation in traditional Chinese smoked horsemeat sausages. Food Control 2015, 50, 869–875. [Google Scholar] [CrossRef]
- Swetwiwathana, A.; Visessanguan, W. Potential of bacteriocin-producing lactic acid bacteria for safety improvements of traditional Thai fermented meat and human health. Meat Sci. 2015, 109, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Talon, R.; Leroy, S.; Lebert, I. Microbial ecosystems of traditional fermented meat products: The importance of indigenous starters. Meat Sci. 2007, 77, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; Verluyten, J.; De Vuyst, L. Functional meat starter cultures for improved sausage fermentation. Int. J. Food Microbiol. 2006, 106, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Franciosa, I.; Alessandria, V.; Dolci, P.; Rantsiou, K.; Cocolin, L. Sausage fermentation and starter cultures in the era of molecular biology methods. Int. J. Food Microbiol. 2018, 279, 26–32. [Google Scholar] [CrossRef]
- Landeta, G.; Curiel, J.A.; Carrascosa, A.V.; Muñoz, R.; de Las Rivas, B. Technological and safety properties of lactic acid bacteria isolated from Spanish dry-cured sausages. Meat Sci. 2013, 95, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, I. Presence, changes and technological properties of yeast species during processing of pastirma, a Turkish dry-cured meat product. Food Control 2015, 50, 76–84. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, X.; Shan, C.; Xia, X.; Wang, Y.; Dong, M.; Zhou, J. Novel bacteriocin produced by Lactobacillus alimentarius FM-MM4 from a traditional Chinese fermented meat Nanx Wudl: Purification, identification and antimicrobial characteristics. Food Control 2017, 77, 290–297. [Google Scholar] [CrossRef]
- Sun, J.; Cao, C.; Feng, M.; Xu, X.; Zhou, G. Technological and safety characterization of coagulase-negative staphylococci with high protease activity isolated from Traditional Chinese fermented sausages. Lwt—Food Sci. Technol. 2019, 114, 108371. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. New trends in technology and identity of traditional dairy and fermented meat production processes: Preservation of typicality and hygiene. Trends Food Sci. Technol. 2014, 37, 51–58. [Google Scholar] [CrossRef]
- Yi, L.; Su, G.; Hu, G.; Peng, Q. Diversity study of microbial community in bacon using metagenomic analysis. J. Food Saf. 2017, 37, e12334. [Google Scholar] [CrossRef]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef]
- Raimondi, S.; Amaretti, A.; Rossi, M.; Fall, P.A.; Tabanelli, G.; Gardini, F.; Montanari, C. Evolution of microbial community and chemical properties of a sourdough during the production of Colomba, an Italian sweet leavened baked product. Lwt—Food Sci. Technol. 2017, 86, 31–39. [Google Scholar] [CrossRef]
- Duru, I.C.; Laine, P.; Andreevskaya, M.; Paulin, L.; Kananen, S.; Tynkkynen, S.; Auvinen, P.; Smolander, O. Metagenomic and metatranscriptomic analysis of the microbial community in Swiss-type Maasdam cheese during ripening. Int. J. Food Microbiol. 2018, 281, 10–22. [Google Scholar] [CrossRef]
- Jung, M.Y.; Kim, T.; Lee, C.; Kim, J.Y.; Song, H.S.; Kim, Y.B.; Ahn, S.W.; Kim, J.S.; Roh, S.W.; Lee, S.H. Role of jeotgal, a Korean traditional fermented fish sauce, in microbial dynamics and metabolite profiles during kimchi fermentation. Food Chem. 2018, 265, 135–143. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Ren, H.; Zhan, Y. Comparison of bacterial diversity profiles and microbial safety assessment of salami, Chinese dry-cured sausage and Chinese smoked-cured sausage by high-throughput sequencing. Lwt—Food Sci. Technol. 2018, 90, 108–115. [Google Scholar] [CrossRef]
- Jin, G.; Zhang, J.; Yu, X.; Zhang, Y.; Lei, Y.; Wang, J. Lipolysis and lipid oxidation in bacon during curing and drying-ripening. Food Chem. 2010, 123, 465–471. [Google Scholar] [CrossRef]
- Jin, G.; He, L.; Li, C.; Zhao, Y.; Chen, C.; Zhang, Y.; Zhang, J.; Ma, M. Effect of pulsed pressure-assisted brining on lipid oxidation and volatiles development in pork bacon during salting and drying-ripening. Lwt—Food Sci. Technol. 2015, 64, 1099–1106. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, M.; Chen, J.; Luo, Y.; Ye, F.; Jiao, S.; Hu, X.; Zhang, J.; Lü, X. Bacterial diversity in traditional sourdough from different regions in China. Lwt—Food Sci. Technol. 2018, 96, 251–259. [Google Scholar] [CrossRef]
- Mariutti, L.R.B.; Bragagnolo, N. Influence of salt on lipid oxidation in meat and seafood products: A review. Food Res. Int. 2017, 94, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Ras, G.; Leroy, S.; Talon, R. Nitric oxide synthase: What is its potential role in the physiology of staphylococci in meat products? Int. J. Food Microbiol. 2018, 282, 28–34. [Google Scholar] [CrossRef]
- Cardinali, F.; Milanović, V.; Osimani, A.; Aquilanti, L.; Taccari, M.; Garofalo, C.; Polverigiani, S.; Clementi, F.; Franciosi, E.; Tuohy, K.; et al. Microbial dynamics of model Fabriano-like fermented sausages as affected by starter cultures, nitrates and nitrites. Int. J. Food Microbiol. 2018, 278, 61–72. [Google Scholar] [CrossRef]
- Ge, Q.; Gu, Y.; Zhan, W.; Yin, Y.; Yu, H.; Wu, M.; Wang, Z.; Zhou, G. Comparison of microbial communities from different Jinhua ham factories. AMB Express 2017, 7, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, L.; Han, Y.; Zhong, S.; Liu, H.; Han, T.; Xie, Y. Isolation and partial characterization of a bacteriocin produced by Lactobacillus plantarum BM-1 isolated from a traditionally fermented Chinese meat product. Microbiol. Immunol. 2013, 57, 746–755. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats: A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef]
- Janssens, M.; Myter, N.; De Vuyst, L.; Leroy, F. Community dynamics of coagulase-negative staphylococci during spontaneous artisan-type meat fermentations differ between. smoking and moulding treatments. Int. J. Food Microbiol. 2013, 166, 168–175. [Google Scholar] [CrossRef]
- Casaburi, A.; Piombino, P.; Nychas, G.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef]
- Yang, X.; Zhu, L.; Zhang, Y.; Liang, R.; Luo, X. Microbial community dynamics analysis by high-throughput sequencing in chilled beef longissimus steaks packaged under modified atmospheres. Meat Sci. 2018, 141, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Aurélie, R.; Vicky, M.; Kalliopi, G.; Didier, R. Intracellular Rickettsiales: Insights into manipulators of eukaryotic cells. Trends Mol. Med. 2011, 17, 573–583. [Google Scholar] [CrossRef]
- Jin, G.; Zhu, Y.; Xu, Y. Mystery behind Chinese liquor fermentation. Trends Food Sci. Technol. 2017, 63, 18–28. [Google Scholar] [CrossRef]
- Tidona, F.; Meucci, A.; Povolo, M.; Pelizzola, V.; Zago, M.; Contarini, G.; Carminati, D.; Giraffa, G. Applicability of Lactococcus hircilactis and Lactococcus laudensis as dairy cultures. Int. J. Food Microbiol. 2018, 271, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aspri, M.; Bozoudi, D.; Tsaltas, D.; Hill, C.; Papademas, P. Raw donkey milk as a source of Enterococcus diversity: Assessment of their technological properties and safety characteristics. Food Control 2017, 73, 81–90. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Yu, D.; Gao, P.; Jiang, Q.; Yang, F. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 2018, 111, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Alahakoon, A.U.; Jayasena, D.D.; Ramachandra, S.; Jo, C. Alternatives to nitrite in processed meat: Up to date. Trends Food Sci. Technol. 2015, 45, 37–49. [Google Scholar] [CrossRef]

| Region | City | Encode | Average Temperature (°C) A | Average Relative Humidity (%) A | Altitude (m) |
|---|---|---|---|---|---|
| Basin | Suining | BA | 7~8 °C | 75~80% | <500 m |
| Leshan | BB | ||||
| Chengdu | BC | ||||
| Mountains | Yibin | MA | 9~10 °C | 75~80% | 500~2000 m |
| Ya’an | MB | ||||
| Xichang | MC | ||||
| Plateau | Kangding | PA | −1~0 °C | 50~55% | >2500 m |
| Ma’er kang | PB |
| Parameters | Groups | ||
|---|---|---|---|
| Basin (n = 14) | Mountains (n = 15) | Plateau (n = 10) | |
| Aw | |||
| Means ± SE | 0.814 ± 0.037 b | 0.855 ± 0.023 a | 0.879 ± 0.049 a |
| Range | 0.761~0.924 | 0.825~0.912 | 0.828~0.974 |
| pH | |||
| Means ± SE | 5.45 ± 0.36 b | 5.68 ± 0.30 ab | 5.92 ± 0.25 a |
| Range | 5.02~6.03 | 5.27~6.48 | 5.59~6.44 |
| Salt content A | |||
| Means ± SE | 3.37 ± 1.19 | 3.00 ± 0.86 | 2.807 ± 1.359 |
| Range | 1.88~5.24 | 1.87~4.95 | 1.01~5.53 |
| Nitrite content B | |||
| Means ± SE | 1.12 ± 0.80 b | 4.80 ± 3.46 a | 2.57 ± 2.15 b |
| Range | 0.21~2.39 | 0.56~13.34 | 0.35~8.06 |
| Peroxide value C | |||
| Means ± SE | 17.73 ± 17.22 | 13.59 ± 9.29 | 13.51 ± 6.70 |
| Range | 1.54~61.70 | 3.40~34.00 | 7.59~26.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, X.; Pan, W.; Liu, A.; Liu, S.; Yang, Y.; Zou, L. Evaluation of Bacterial Diversity and Quality Features of Traditional Sichuan Bacon from Different Geographical Region. Appl. Sci. 2021, 11, 9738. https://doi.org/10.3390/app11209738
Wang S, Wang X, Pan W, Liu A, Liu S, Yang Y, Zou L. Evaluation of Bacterial Diversity and Quality Features of Traditional Sichuan Bacon from Different Geographical Region. Applied Sciences. 2021; 11(20):9738. https://doi.org/10.3390/app11209738
Chicago/Turabian StyleWang, Song, Xingjie Wang, Wanshu Pan, Aiping Liu, Shuliang Liu, Yong Yang, and Likou Zou. 2021. "Evaluation of Bacterial Diversity and Quality Features of Traditional Sichuan Bacon from Different Geographical Region" Applied Sciences 11, no. 20: 9738. https://doi.org/10.3390/app11209738
APA StyleWang, S., Wang, X., Pan, W., Liu, A., Liu, S., Yang, Y., & Zou, L. (2021). Evaluation of Bacterial Diversity and Quality Features of Traditional Sichuan Bacon from Different Geographical Region. Applied Sciences, 11(20), 9738. https://doi.org/10.3390/app11209738