Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Material and Methods
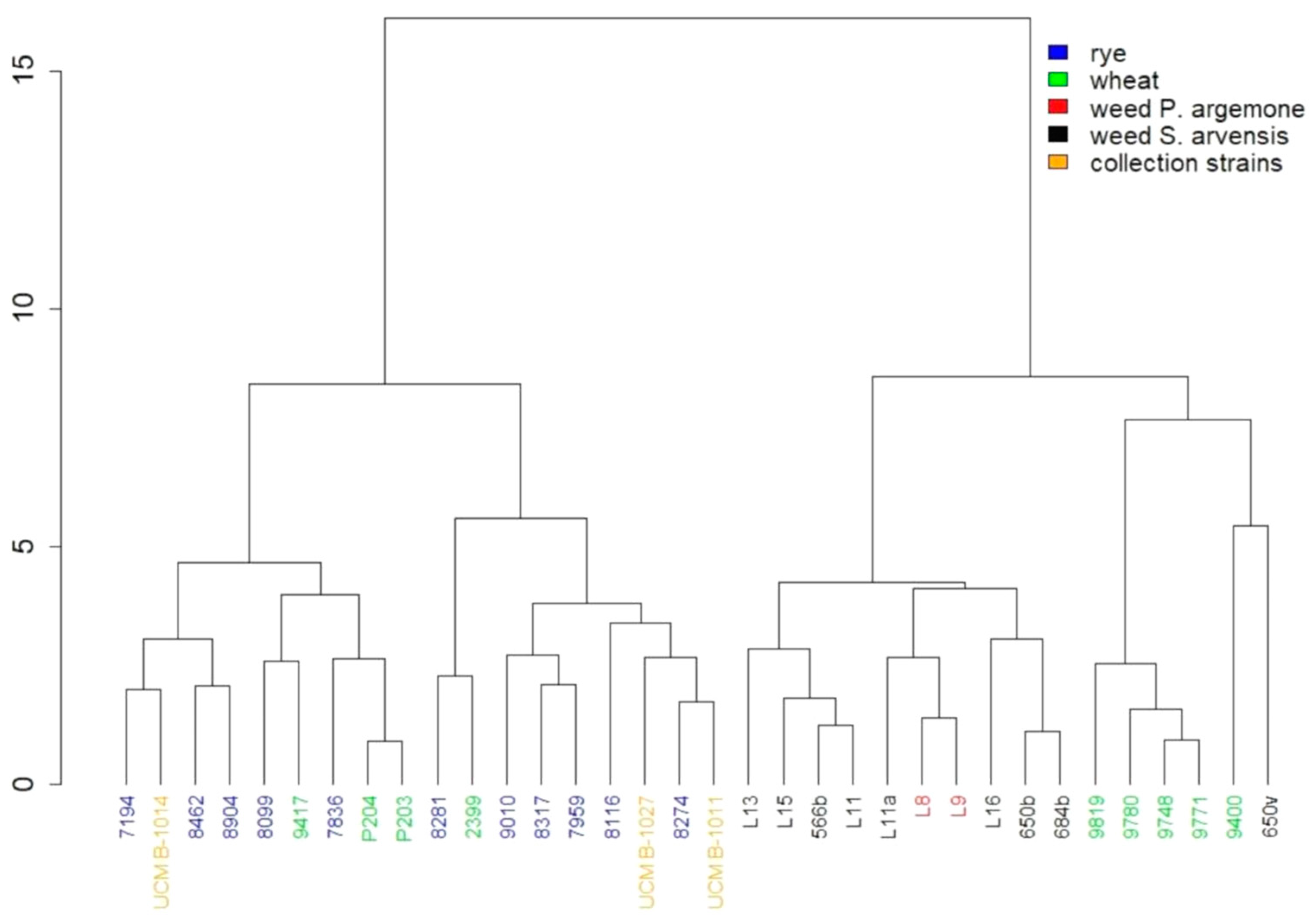
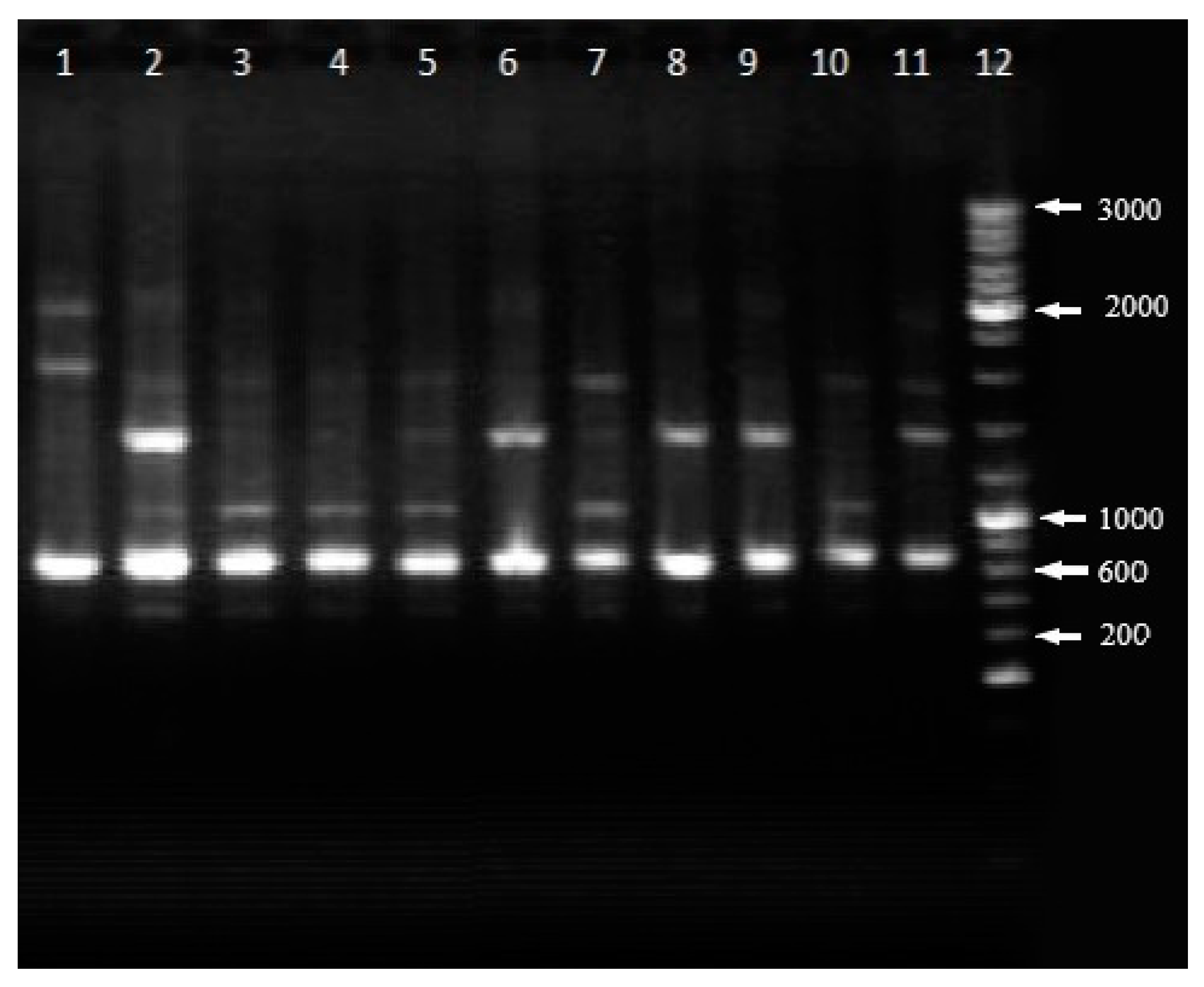
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tripathi, D. Bacterial pathogens in plants. J. Bacteriol. Mycol. 2017, 4, 38–39. [Google Scholar] [CrossRef][Green Version]
- Sundin, G.W.; Castiblanco, L.F.; Yuan, X.; Zeng, Q.; Yang, C.H. Bacterial disease management: Challenges, experience, innovation and future prospects: Challenges in Bacterial Molecular Plant Pathology. Mol. Plant Pathol. 2016, 17, 1506–1518. [Google Scholar] [CrossRef] [PubMed]
- Patyka, V.P. Phytopathogenic bacteria in contemporary agriculture. Microbiolohichnyi Zhurnal 2016, 78, 71–83. [Google Scholar] [CrossRef]
- Kolomiiets, Y.; Grygoryuk, I.; Likhanov, A.; Butsenko, L.; Blume, Y. Induction of Bacterial Canker Resistance in Tomato Plants Using Plant Growth Promoting Rhizobacteria. Open Agric. J. 2019, 13, 215–222. [Google Scholar] [CrossRef]
- Patyka, V.; Pasichnyk, L.; Butsenko, L.; Petrychenko, V.; Zubachev, S.; Dankevych, L.; Gnatiuk, Y.; Huliaiva, H.; Tokovenko, I.; Kalinichenko, A.; et al. Express Diagnostics of Phytopathogenic Bacteria and Phytoplasmas in Agrophytocenosis; Suszanowich, D., Patyka, V., Wyd-wo, I., Eds.; Drukarnia Swietego Krzyza: Opole, Poland, 2019; ISBN 978-83-7342-684-9. [Google Scholar]
- Maraite, H.; Bragard, C.; Duveiller, E. The status of resistance to bacterial diseases of wheat. In Wheat Production in Stressed Environments; Buck, H.T., Nisi, J.E., Salomon, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 37–49. [Google Scholar] [CrossRef]
- Valencia-Botín, A.J.; Cisneros-López, M.E. A Review of the Studies and Interactions of Pseudomonas syringae Pathovars on Wheat. Intern. J. Agron. 2012, 2012, 692350. [Google Scholar] [CrossRef]
- Matveeva, Y.V.; Pekhtereva, E.S.; Polityko, V.A.; Ignatov, A.N.; Nikolaeva, E.V.; Schaad, N.W. Distribution and virulence of Pseudomonas syringae pv. atrofaciens, causal agent of basal glume rot, in Russia. In Pseudomonas Syringae and Related Pathogens; Iacobellis, N.S., Collmer, A., Hutcheson, S.W., Mansfield, J.W., Morris, C.E., Schaad, N.W., Stead, D.E., Surico, G., Ullrich, M.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 97–105. [Google Scholar] [CrossRef]
- Alexandrova, M.; Zaccardelli, M.; Stefani, E.; Bazzi, C. Testing for Pseudomonas syringae pv. atrofaciens and Xanthomonas campestris pathovars on cereals in Italy. EPPO Bull. 1995, 25, 437–448. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef]
- Kazempour, M.N.; Kheyrgoo, M.; Pedramfar, H.; Rahimian, H. Isolation and identification of bacterial glum blotch and leaf blight on wheat (Triticum aestivum L.) in Iran. Afr. J. Biotechnol. 2010, 9, 2866–2871. Available online: https://www.ajol.info/index.php/ajb/article/view/79941 (accessed on 3 November 2020).
- Sultanov, R.I.; Arapidi, G.P.; Vinogradova, S.V.; Govorun, V.M.; Luster, D.G.; Ignatov, A.N. Comprehensive analysis of draft genomes of two closely related pseudomonas syringae phylogroup 2b strains infecting mono- and dicotyledon host plants. BMC Genom. 2016, 17, 1010. [Google Scholar] [CrossRef]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balesta, G.M.; Spinelli, F. Pseudomonas syringae pv. actinidiae: Ecology, Infection Dynamics and Disease Epidemiology. Microb. Ecol. 2020, 80, 81–102. [Google Scholar] [CrossRef]
- Taghavi, S.M.; Keshavarz, K. Identification of the causal agent of bacterial wheat blight in Fars and Kohgiluyeh Boyrahmad provinces and the reaction of certain wheat cultivars to them. J. Sci. Technol. Agric. Nat. Res. 2003, 6, 171–180. Available online: https://jstnar.iut.ac.ir/browse.php?a_code=A-10-2-146&slc_lang=en&sid=1 (accessed on 3 November 2020).
- Tarkowski, P.; Vereecke, D. Threats and opportunities of plant pathogenic bacteria. Biotechnol. Adv. 2014, 32, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.S.D.A.; Samson, R. Population dynamics of Pseudomonas savastanoi pv. phaseolicola in bean, throughout the epiphytic and pathogenic phases. Pesqui. AgropecuáRia Bras. 2016, 51, 623–630. [Google Scholar] [CrossRef]
- Dobrovol’skaya, T.G.; Khusnetdinova, K.A.; Manucharova, N.A.; Golovchenko, A.V. Structure of epiphytic bacterial communities of weeds. Microbiology 2017, 86, 257–263. [Google Scholar] [CrossRef]
- Klement, Z.; Rudolph, K.; Sands, D. Methods in Phytobacteriology; Academiai Kiado: Budapest, Hungary, 1990. [Google Scholar] [CrossRef]
- Pasichnyk, L.A.; Butsenko, L.M. Serological features of bacteria Pseudomonas syringae agroecosystems of cereal. Microbiolohichnyi Zhurnal 2018, 80, 41–54. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2018. Available online: https://www.R-project.org/ (accessed on 3 November 2020).
- Sazakli, E.; Leotsinidis, M.; Vantarakis, A.; Papapetropoulou, M. Comparative typing of Pseudomonas species isolated from the aquatic environment in Greece by SDS-PAGE and RAPD analysis. J. Appl. Microbiol. 2005, 99, 1191–1203. [Google Scholar] [CrossRef]
- Butsenko, L.M.; Savenko, O.A.; Pasichnyk, L.A.; Shcherbyna, T.M.; Patyka, V.P. Fatty Acid Composition of Cellular Lipids Pseudomonas syringae, Isolated from Cereal Agrophytocenosis. Microbiolohichnyi Zhurnal 2017, 79, 56–64. [Google Scholar] [CrossRef]
- Butsenko, L.M.; Pasichnyk, L.A. Genetic Heterogenicity of Pseudomonas syringae pv. atrofaciens Strains Based on RAPD-PCR Analyze. Microbiolohichnyi Zhurnal 2018, 80, 48–62. [Google Scholar] [CrossRef]
- Young, J.M. Taxonomy of Pseudomonas syringae. J. Plant Pathol. 2010, 92, S1.5–S1.14. [Google Scholar] [CrossRef]
- Gutiérrez-Barranquero, J.A.; Carrión, V.J.; Murillo, J.; Arrebola, E.; Arnold, D.L.; Cazorla, F.M.; De Vicente, A. A Pseudomonas syringae diversity survey reveals a differentiated phylotype of the pathovar syringae associated with the mango host and mangotoxin production. Phytopathology 2013, 103, 1115–1129. [Google Scholar] [CrossRef]
- Barta, T.M.; Willis, D.K. Biological and Molecular Evidence that Pseudomonas syringae pathovars coronafaciens, striafaciens and garcae are likely the same pathovar. J. Phytopathol. 2005, 153, 492–499. [Google Scholar] [CrossRef]
- Lindström, K.; Martínez-Romero, M.E. International Committee on Systematics of Prokaryotes; Subcommittee on the taxonomy of Agrobacterium and Rhizobium. Minutes of the meeting, 26 July 2004, Toulouse, France. Int. J. Syst. Evol. Microbiol. 2005, 55, 1383. [Google Scholar] [CrossRef]
- Peix, A.; Ramírez-Bahena, M.-H.; Velázquez, E. The current status on the taxonomy of Pseudomonas revisited: An update. Infect. Genet. Evol. 2018, 57, 106–116. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, H.E.; Thakur, S.; Guttman, D.S. Evolution of plant pathogenesis in Pseudomonas syringae: A genomics perspective. Annu. Rev. Phytopathol. 2011, 49, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Takeuchi, T.; Matsuda, I. Comparative analysis of Pseudomonas syringae pv. actinidiae and pv. phaseolicola based on phaseolotoxin-resistant ornithine carbamoyltransferase gene (argK) and 16S-23S rRNA intergenic spacer sequences. Appl. Environ. Microbiol. 1997, 63, 282–288. [Google Scholar] [CrossRef]
- Maraite, H.; Weyns, J. Pseudomonas syringae pv. aptata and pv. atrofaciens, Specific Pathovars or Members of pv. syringae? In Pseudomonas Syringae Pathovars and Related Pathogens; Rudolph, K., Burr, T.J., Mansfield, J.W., Stead, D., Vivian, A., von Kietzell, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 515–520. [Google Scholar] [CrossRef]
- Xin, X.F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Stead, D.E.; Hennessy, J.; Elphinstone, J.G.; Wilson, J.K. Modern methods for classification of plant pathogenic bacteria including Pseudomonas syringae. Dev. Plant Pathol. 1997, 9, 427–434. [Google Scholar] [CrossRef]
- Iličić, R.; Balaž, J.; Stojšin, V.; Jošić, D. Characterization of Pseudomonas syringae pathovars from different sweet cherry cultivars by RAPD analysis. Genetika 2016, 48, 285–295. [Google Scholar] [CrossRef]
- Butsenko, L.; Pasichnyk, L.; Kolomiiets, Y.; Kalinichenko, T. The Effect of Pesticides on the Tomato Bacterial Speck Disease Pathogen Pseudomonas Syringae pv. Tomato. Appl. Sci. 2020, 10, 3263. [Google Scholar] [CrossRef]
- Kolomiiets, Y.; Grygoryluk, I.; Butsenko, L.; Kalinichenko, A. Biotechnological control methods against phytopathogenic bacteria in tomatoes. Appl. Ecol. Environ. Res. 2019, 17, 3215–3230. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Strains, Isolated from P. argemone | Strains, Isolated from S. arvensis | P. syringae pv. atrofaciens | P. syringae pv. syringae | |
|---|---|---|---|---|---|
| Strains | L8, L9 | L11, L11a, L16 | L13, L15, | 9780, UCM B-1011, UCM B-1014 | UCM B-1027 |
| Gram reaction | − | − | − | − | − |
| Shape | rod | rod | rod | rod | rod |
| Spore-forming | − | − | − | − | − |
| Motility | + | + | + | + | + |
| Fluorescence | + | + | + | + | + |
| OF-test | O | O | O | O | O |
| Levan * | + | + | + | + | + |
| Oxidase * | − | − | − | − | − |
| Potato rot * | − | − | − | − | − |
| Arginine dihydrolase * | − | − | − | − | − |
| Tobacco hypersensitivity * | + | + | + | + | + |
| NEFERMtest24 (MikroLaTEST®, ErbaLachema) | |||||
| Urease | − | − | − | − | − |
| Ornithine decarboxylase | − | − | − | − | − |
| Lysine decarboxylase | − | − | − | − | − |
| Acetamide | − | w | w | − | − |
| β-glucosidase | + | + | + | + | + |
| N-Acetyl-β-D-Glucosaminidase | − | − | − | − | − |
| Utilization of citrate | + | + | + | + | + |
| Lactose | − | − | − | − | − |
| Mannitol | + | + | + | + | + |
| Trehalose | − | − | − | − | − |
| Xylose | + | + | + | + | + |
| Arabinose | + | + | + | + | + |
| α-galactosidase | − | − | − | − | − |
| Malonate | − | − | − | − | − |
| Galactose | + | + | + | + | + |
| Maltose | − | − | − | − | − |
| Cellobiose | − | − | − | − | − |
| Saccharose | + | + | + | + | + |
| Inositol | w | w | − | − | − |
| γ-glutamyl transferase | − | − | − | − | − |
| Phosphatase | + | + | + | + | + |
| Aesculin hydrolysis | + | + | + | + | + |
| Strain | Host Plant | Source | Year of Isolation | Sero- Group | Aggressiveness | |
|---|---|---|---|---|---|---|
| for Wheat | for Weeds | |||||
| L8 | Papaver argemone L. | wheat field, Kyiv region | 2019 | 2 | 2 | 1 * |
| L9 | Papaver argemone L. | wheat field, Kyiv region | 2019 | 2 | 2 | 1 * |
| L11 | Sonchus arvensis L. | wheat field, Kyiv region | 2019 | 2 | 1 | 2 ** |
| L11a | Sonchus arvensis L. | wheat field, Kyiv region | 2019 | 2 | 2 | 2 ** |
| L13 | Sonchus arvensis L. | wheat field, Kyiv region | 2019 | 6 | 2 | 2 ** |
| L15 | Sonchus arvensis L. | wheat field, Kyiv region | 2019 | 6 | 3 | 2 ** |
| L16 | Sonchus arvensis L. | wheat field, Kyiv region | 2019 | 6 | 3 | 3 ** |
| P. syringae pv. atrofaciens 9780 | Triticum aestivum L. | Collection of microorganisms of the Department of phytopathogenic bacteria of the Zabolotny Institute of Microbiology and Virology of NASU | 2012 | 2 | 1–2 | 2 ** |
| P. syringae pv. atrofaciens UCM B-1014 | Triticum aestivum L. | 1972 | 2 | 4 | 2 ** | |
| P. syringae pv. atrofaciens UCM B-1011 | Triticum sp. | PDDCC 4394 | 4 | 4 | N/D | |
| P. syringae pv. syringae UCM B-1027 | Syringa vulgaris L. | NCPPB 281 | 1 | 4 | N/D | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butsenko, L.; Pasichnyk, L.; Kolomiiets, Y.; Kalinichenko, A.; Suszanowicz, D.; Sporek, M.; Patyka, V. Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field. Appl. Sci. 2021, 11, 286. https://doi.org/10.3390/app11010286
Butsenko L, Pasichnyk L, Kolomiiets Y, Kalinichenko A, Suszanowicz D, Sporek M, Patyka V. Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field. Applied Sciences. 2021; 11(1):286. https://doi.org/10.3390/app11010286
Chicago/Turabian StyleButsenko, Liudmyla, Lidiia Pasichnyk, Yuliia Kolomiiets, Antonina Kalinichenko, Dariusz Suszanowicz, Monika Sporek, and Volodymyr Patyka. 2021. "Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field" Applied Sciences 11, no. 1: 286. https://doi.org/10.3390/app11010286
APA StyleButsenko, L., Pasichnyk, L., Kolomiiets, Y., Kalinichenko, A., Suszanowicz, D., Sporek, M., & Patyka, V. (2021). Characteristic of Pseudomonas syringae pv. atrofaciens Isolated from Weeds of Wheat Field. Applied Sciences, 11(1), 286. https://doi.org/10.3390/app11010286