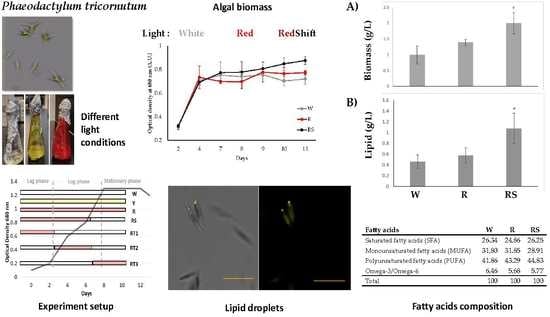
3.1. Effect of Light Colors on Biomass and Lipid Yield in Mixotrophic and Autotrophic Conditions
For the mixotrophic conditions, two different carbon sources were used by supplementing the standard L1 media with 1% glucose (W1, Y1, and R1) or 1% glycine (W2, Y2, and R2). The highest biomass was achieved under the red light condition compared to white or yellow light (
Figure 1A). Specifically, the level of biomass productivity obtained in autotrophic growth (R) was significantly higher by 1.16 and 1.05-fold compared to W and Y autotrophic conditions, respectively. Our results are in agreement with multiple studies that have reported an increased higher production rate of
Scenedesmus sp. under red and blue light [
25] and
Nannochloropsis sp. under red and yellow light [
26]. No additional biomass was obtained in R1 or R2 conditions compared to R, suggesting that mixotrophic conditions are less interesting for an industrial application.
The lipid production of
P. tricornutum was only different, and significantly higher, in red light autotrophic (R) or mixotrophic supplemented with glycine (R2) conditions (
Figure 1B). The latter was expected, as studies have reported an increase in biomass and lipids by multi-fold under mixotrophic conditions in different species [
27,
28]. Furthermore, many studies have shown a positive trend of red light with an increase in the specific growth rate and lipid production in
Chlorella,
Botryococcus, and
Nannochloropsis [
7,
26,
29]. Another study showed a positive trend of the combined impact of red and blue light on biomass productivity and fucoxanthin production in
P. tricornutum [
30]. However, contrasting results have been reported where they observed high productivity under yellow light as compared to red and blue light for
Chlamydomonas reinhardtii [
13]. However, these experiments were conducted in turbidostat controlled lab-scale panel photobioreactors and reported that in mass culture, productivity and biomass specific light absorption are inversely correlated [
13]. Furthermore, in the same study, they have reported that the supplementation of blue light to yellow light is better for stable cultivation.
The growth of microalgae in mixotrophic conditions is an interesting choice for enhancing the productivity of particular metabolites, which will vary according to the product of interest wanted [
31]. However, there are cost-effective and economic advantages to growth in mixotrophy if the carbon source is derived from cheap waste resources such as agricultural and municipal wastewater that are rich in micronutrients and carbon sources [
32]. There has been in-depth research conducted on the impact of varied carbon source such as glucose, fructose, and glycerol on enhancing the biomass of
P. tricornutum [
31,
33]. A relevant point to consider when comparing results from red light with those of red light combined with blue, or other wavelengths is that the monochromatic red light corresponds to the absence of the others. However, our results, combined with findings from several published studies, have shown the importance of monochromatic light or the mixing of wavelengths on the biomass and lipid production in different microalgae species as discussed above [
7,
13,
25,
26]. This motivated us to further optimize the photoautotrophic conditions.
3.2. Growth Curve Analysis of P. tricornutum during Red Light Shift
Studies have reported that light color and intensity bring changes in the photon flux density, which have a strong impact on growth and metabolic pathways [
32,
33]. Photoautotrophic conditions create certain limitations when grown in bioreactors because self-shading by microalgal cells in high-density cultures limits the light available for growth, which can be confirmed in lab-scale experiments by observing the growth curve. Therefore, to reduce the impact of either loss of light as heat or self-shading, the strategy of light shifting from one spectrum to another can trigger different photoreceptors to activate different light-signaling pathways, allowing a better conversion of light energy [
34,
35,
36]. Therefore, we investigated the effect of red light shifts on
P. tricornutum growth during different growth phases under batch cultivation for 11 days.
P. tricornutum grown in red light until the end of exponential phase (from day 1 to 7) and then shifted to white light (RS) showed the greatest growth compared to all other conditions tested (
Figure 2,
Appendix A—
Figure A1). In addition, RS is significantly better to promote growth compared to continuous white (W) or red light (R) (
Figure 2).
P. tricornutum cultured cells were adapting to each condition in a similar manner during the adaptation and exponential phases (
Figure 2). It should be noted that there seems to be a crucial lag between day 4 and day 7 (exponential to stationary), where it is not clear whether cell growth continued before slowing down (
Figure 2). Additionally, the slight difference between R and RS at day 7 is unexpected, since they were under the same conditions. It is possible that these small differences are due to replicate variation and growth chamber light variations. Indeed, when we checked the light intensity using the flux meter, we observed a variation in light intensity of 1–2 μE m
−2 s
−1 between the different corners of the growth chamber, which could explain the variation between R and RS at day 7 and the width of the error bars. However, major differences in growth were observed at the end of the exponential phase and during the stationary phase, where RS clearly promoted growth compared to other treatments (
Figure 2;
Appendix A—
Figure A1). Indeed, at day 11, the RS growth was significantly higher with 22% and 14% more cells than continuous W and R light, respectively (
Figure 2). The cell growth observed under R and RS treatment can be explained, in part, by the presence of a wide range of photoreceptors with the ability to sense red/far-red wavelengths and their potential to prevent photoinhibition [
30,
32]. A similar study has reported the increasing growth of
P. tricornutum under defined and efficient combinations of red and blue lights [
30]. Another study has shown the positive impact of supplementing different light colors on
Chlamydomonas reinhardtii culture [
13]. Through various studies, it was suggested that different classes of photoreceptors have specific responses to specific light mediated by different light-signaling pathways that are yet to be completely elucidated [
2,
37,
38]. The other physiological phenomenon that can change the culture conditions is the rate of photoinhibition, which seems to be higher in W as compared to R. The energy of each photon is inversely proportional to the wavelength. For example, R (λ = 700 nm) is 1.8 eV, whereas blue light (λ = 450 nm) is 2.8 eV. The W or full spectrum (λ = 400–700 nm) tends to cause photoinhibition, inducing cell damage and reducing the overall time of stationary phase as compared to R (
Figure 2). Since the cumulative wavelength of red in R is lower than that in W, it may prevent photoinhibition and consequently lead to higher growth and overall biomass [
39]. Similar results have been reported for
Chlorella sp. [
39] and
Scenedesmus sp. [
25]. In all, the RS further increased growth, which can be attributed to the use of different photoreceptors and the reduction of photoinhibition.
Interestingly, the timing of light shift was important. Indeed, red light shifts during other growth phases (lag, exponential, and stationary phase (RT1, RT2, and RT3) were not efficient to increase biomass compared to RS (
Appendix A—
Figure A1). Future work should investigate other experimental strategies to study circadian rhythms and photosynthesis during light shift conditions.
3.3. Analysis of Total Biomass and Lipid Content under Red Light Shift Treatment
The biomass of
P. tricornutum was significantly greater with RS treatment by 2-fold and 1.4-fold compared to W and R conditions, respectively (
Figure 3A). This correlates with increased cell density (
Figure 2). In addition, our results confirm observations from another study on the impact of red light where biomass increased by 40% under enriched red light [
40]. Similarly, another study reported better growth and higher fucoxanthin content in
P. tricornutum under red and blue light [
30]. That study is similar to our treatment RS, where we have supplied a red wavelength for the first 7 days and then exposed the culture to the white light during the stationary phase. Furthermore, it has been reported that cultured
P. tricornutum under R contains a greater number of thylakoids, where light-dependent reactions occur, compared to
P. tricornutum cultured in W [
41]. On a wider scale, the impact of red light on the ultrastructure of chloroplasts and creating superstacks of thylakoid membrane in diatoms suggest that R will affect photosynthesis, cell division, and consequently promote biomass accumulation [
41]. Altogether, the data suggest that the use of red light and variations thereof improves the overall biomass of
P. tricornutum at the laboratory scale, offering a promising first stage, which should be pursued at a larger scale to be considered a useful strategy for industrial applications.
Total lipids were also analyzed in the same conditions, using dry weight estimation. Nile red assay was used for rapid screening of lipid and confocal microscopy for visual analysis, as described previously [
20].
P. tricornutum produces lipids that are potentially significant for several industrial applications. It has been reported that the crude lipids extracted from
P. tricornutum are around 321.89 mg/g dry weight [
42] or 34% of dry weight [
43] and are comparable to the lipid extracted in W in our study, which is 0.4 g/g dry weight in control white light. Furthermore, the lipid dry weight in RS was significantly increased to 2.3-fold and 1.8-fold as compared to W and R, respectively (
Figure 3B). It is interesting to note the 1.8 fold increase in lipid from R to RS, which suggests that shifting from red light to full spectrum during the stationary phase has an impact on lipid accumulation. It is known that the usual change in the lipid composition is influenced by the cellular C/N ratio, light intensity, nutrient depletion, and other possible inducible biotic/abiotic stress conditions [
28,
29,
30]. An increase in C/N ratio, lipid, and carbohydrate in red light acclimated
P. tricornutum culture has been reported [
11]. The same study reported an increase in the lipid content from red light to blue light shift, whereas the reverse shift, from blue to red light, promoted an increase in carbohydrates. In addition, lipid accumulation under red light has been observed in
Botryococcus sp. [
7] and
Chlorella sp. [
29], which can be linked with effective nitrogen consumption rate and a doubling of cell numbers in red light, which aids in increasing the total lipid content. In addition, high light intensity has been shown to affect lipid biosynthesis in
Nannocloropsis gaditana, which has been correlated with the regulation of cytosolic fatty acid synthase of type 1 (
FAS1) [
44] and polyketide synthase (
PKS) and the down-regulation of the chloroplast fatty acid synthase of type 2 (
FAS2) [
45]. One of the studies has reported a significant increase in lipids in
P. tricornutum at low light, which is in the range of (30 μmol photons m
−2 s
−1) and medium light (300 μmol photons m
−2 s
−1) [
46]. Therefore, lower intensities or light shift treatment may also affect the expression levels of the cytosolic and chloroplastic fatty acid synthases, which in turn would regulate lipid content. Various studies have shown the relation between wavelength, specific wavelength, and wavelength shift on the accumulation of lipids, which can be attributed to enzymes involved in carbon cycle and lipid synthesis [
11,
40,
47]. Therefore, it appears that the combined effect of red-light acclimation and lower light intensity, observed by the red sheet, which reduces light intensity by 10 μE m
−2 s
−1, may be acting together to increase the total lipids in RS (
Figure 3B).
3.4. Visual Analysis of P. tricornutum Cell Morphology and Lipid Droplets
We next examined
P. tricornutum lipid droplets using confocal microscopy (
Figure 4). Our initial expectation based on a gravimetric analysis of lipids was to observe a change in the lipid droplet size. The results showed that the selected light conditions were not responsible for any changes in the morphology, size, or number of lipid droplets of
P. tricornutum cultures grown in the studied light conditions (
Figure 4a,b). The morphological changes in cells are important for microbes to respond to various environmental conditions such as the state of media, the temperature, or the salinity. Previous studies have reported the presence of three morphotypes in
P. tricornutum culture: oval, fusiform, and triradiate [
48]. It is also known that light absorption is the function of the cell shape, size, and pigments [
49]. Microscopic analyses (
Figure 4c,d) revealed a high occurrence of fusiform cells, a limited number of triradiate shapes, and no appearance of the oval form in all of our growth conditions. It has been reported that the fusiform type is dominant in suspension culture over the other forms [
43,
50], which is in agreement with our tested conditions. It has also been reported that the triradiate
P. tricornutum cells are rarely found in laboratory growth conditions [
1,
43]. However, our study detected the presence of a limited number of triradiate cells (
Appendix A—
Figure A2).
P. tricornutum can be grown in both silicon or silicon-free medium; we studied silicon-free media, as it showed better growth for our strain, and it has been reported that fusiform cells have a greater lipid percentage as compared to other forms [
43,
51].
3.5. Fatty Acid Methyl Esters Analysis (FAMEs)
Polyunsaturated fatty acids (PUFA) were present in greatest abundance, followed by monounsaturated fatty acids (MUFA) and lower amounts of saturated fatty acids (SFA), which constituted 41.86%, 31.80%, and 26.34%, respectively (
Table 3). MUFA levels were lower in RS (28.91%) compared to W (31.80%) or R (31.85%) (
Table 3). In addition, the
P. tricornutum PUFA ratio of omega-3/omega-6 was lower in R and RS compared to control W (
Table 3). The evidence suggests that a high ratio of dietary omega-3/omega-6 PUFAs reduces the risk of several diseases [
52]. Therefore,
P. tricornutum cultured in W is a potentially more valuable source of functional food and animal feed components than
P. tricornutum cultivated under R or RS conditions. These results suggest that the RS condition regulated the fatty acid content in
P. tricornutum, specifically lowering MUFAs and increasing PUFAs. It has been reported that stress conditions such as nitrogen depletion, high and low temperature, and continuous high light intensity impacts fatty acid composition, specifically PUFA in
P. tricornutum [
53,
54,
55].
The distribution of fatty acids was characterized (
Appendix A;
Table A1), and various lipid classes of
P. tricornutum are summarized in
Table 4. The main lipid components were mid-chain FAs, 14:0, 16:0, 16:1n-7, and 20:5n-3, which together represented 76.46% of total fatty acids (
Table 4). Our results are similar to those reported by [
42]. Palmitoleic acid (16:1n-7), which constituted 28.20% of the total FAs, was predominant in
P. tricornutum control (W) but was reduced to 24.29% in RS condition (
Appendix A;
Table A1). Interestingly, hexadecatrienoic acid (16:3n-4) was increased in R and RS compared to W (
Appendix A;
Table A1). There was a comparable amount of the omega-3 fatty acid, eicosapentaenoic acid (20:5 n-3), in all light conditions and it constituted almost 22% of total acids, which was the second most predominant FA (
Table 4). In other studies, it was the PUFA that was present in highest mass proportions [
53,
54,
55]. Humans do not produce sufficient long-chain omega-3 fatty acids to meet their physiological needs; therefore, it is essential to obtain them from external sources. Omega-3 fatty acids have been associated with proper fetal development, including neuronal, retinal, and immune function [
56]. They have also shown positive effects on coronary disease and inflammation, and they have been approved for the treatment of patients with severe hypertriglyceridemia [
33]. These fatty acids are predominantly found in cold water fish, such as salmon. However, our work suggests that microalgae and diatoms can be considered as a sustainable and vegetarian source of omega-3 fatty acids.
The structural features (chain length, unsaturation, and branching) of fatty acids determine the properties of algae-derived biodiesel, including ignition quality, heat of combustion, cold flow, oxidative stability, exhaust emissions, viscosity, and lubricity [
57]. Thus, the major fractions of fatty acid composition, which was accepted as input in the Biodiesel Analyzer, were studied for properties of biodiesel [
23] (
Table 5). For biofuel of an industrial quality, it needs to have similar physical properties as diesel, which can be used efficiently in combustion engines. According to a study, fatty acids with chain lengths ranging from C16 to C18 should be high in potential feedstock for suitable biodiesel production [
58]. It is suggested that FAME with a high percentage of monounsaturation (C16–C18) is the most desirable compromise between cold flow and oxidative stability [
59]. The W and R induced the production of monounsaturated fatty acids as compared to RS. The total percent of saturation and unsaturation—especially the amount of long-chain polyunsaturated fatty acids such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)—will have an impact on the biodiesel fuel properties [
60]. The results showed that the cetane number (CN), which determines the ignition quality of the fuel, is higher in
P. tricornutum cultured in R and RS as compared to W (
Table 5). It is known that the higher the CN, the better the ignition of the fuel and vice versa [
57]. Literature studies have also mentioned that increases in the degree of unsaturation lead to decreases in CN viscosity, which is a useful measure for monitoring oxidation progression. As reported, fuels with high viscosity meeting the international standards have better lubricating quality, which increases the life of the engine [
61]. In the same study, there is a relationship between hydrocarbon (HC) emission and viscosity. The HC emissions increase with decreasing viscosity [
61]. In our study, the viscosity is lower in all the conditions comparing the standard requirement of EN 14214:2008, but it meets the requirement of ASTM D6751.
Since this study presents the importance of light as a crucial factor for affecting biomass and lipid content in
P. tricornutum, it supports its importance in metabolic processes of microalgae in the biotechnology field. However, the specific behavior of each species may be unique with respect to the varied light conditions, as supported by the following evidence studied in an interdisciplinary approach. For example, Orefice et al. [
62] showed that the fluctuation of red light on blue light spectral distribution affected the metabolic state of cells via increase in non-photochemical quenching and ultimately promoting an increase in carbohydrates, glycolipids, and saturated fatty acids in
Skeletonema marinoi [
62]. In another study, it was observed that different doses of blue wavelengths cause significant alternations in the growth cycle of
Skeletonema marinoi [
63]. The change in the growth pattern and biochemical composition can be referred to the ability of diatoms and microalgae to finely balance the light-harvesting and photoprotective capacity [
64,
65,
66,
67]. The ability of diatoms to respond favorably to red light is given to the presence of various photoreceptors such as phytochromes and exclusive aureochromes that are involved in the RL signaling process in
P. tricornutum [
68,
69,
70]. In a different approach using spectroscopy, the occurrence of red-shifted fucoxanthin-chlorophyll protein (FCP) showing the fluorescence peak at 714 nm was observed [
71]. It is considered that Phaeodactylum red-shifted FCP is involved in enhancing the light-harvesting ability of cells by the prolonged excited state lifetimes and the absorption spectrum extended to wavelengths longer than 700 nm [
71]. In general, the clarity in the relationship between diatoms and light is still an open topic for researchers to study both fundamental as well as industrial applications.
For the conclusion of this study, an effect of the specific RS light shift condition in the fatty acid composition was observed that can be important in applications in both human nutrition and renewable energy. Overall, it is interesting to note that changes in the specific wavelength during the stationary phase can impact the total accumulation of lipid and fatty acid composition.

 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




