A Pilot Longitudinal Evaluation of MicroRNAs for Monitoring the Cognitive Impairment in Pediatric Multiple Sclerosis

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Subjects and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Compston, A.; Coles, A.J. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Olsson, T.; Barcellos, L.F.; Alfredsson, L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 2017, 13, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Banwell, B. Multiple sclerosis in children. Handb. Clin. Neurol. 2014, 122, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; Beadnall, H.; Barnett, M.H.; Suter, C.M.; Buckland, M.E. Exosomal microRNA signatures in multiple sclerosis reflect disease status. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Liguori, M.; Nuzziello, N.; Licciulli, F.; Consiglio, A.; Simone, M.; Viterbo, R.G.; Creanza, T.M.; Ancona, N.; Tortorella, C.; Margari, L.; et al. Combined microRNA and mRNA expression analysis in pediatric multiple sclerosis: An integrated approach to uncover novel pathogenic mechanisms of the disease. Hum. Mol. Genet. 2018, 27, 66–79. [Google Scholar] [CrossRef]
- Nuzziello, N.; Vilardo, L.; Pelucchi, P.; Consiglio, A.; Liuni, S.; Trojano, M.; Liguori, M. Investigating the Role of MicroRNA and Transcription Factor Co-regulatory Networks in Multiple Sclerosis Pathogenesis. Int. J. Mol. Sci. 2018, 19, 3652. [Google Scholar] [CrossRef]
- Liguori, M.; Nuzziello, N.; Simone, M.; Amoroso, N.; Viterbo, R.G.; Tangaro, S.; Consiglio, A.; Giordano, P.; Bellotti, R.; Trojano, M. Association between miRNAs expression and cognitive performances of Pediatric Multiple Sclerosis patients: A pilot study. Brain Behav. 2019, 9, e01199. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444. [Google Scholar] [CrossRef]
- Amato, M.P.; Goretti, B.; Ghezzi, A.; Lori, S.; Zipoli, V.; Moiola, L.; Falautano, M.; De Caro, M.F.; Viterbo, R.; Patti, F.; et al. Cognitive and psychosocial features in childhood and juvenile MS: Two-year follow-up. Neurology 2010, 75, 1134–1140. [Google Scholar] [CrossRef]
- Chang, W.-S.; Wang, Y.-H.; Zhu, X.-T.; Wu, C.-J. Genome-Wide Profiling of miRNA and mRNA Expression in Alzheimer’s Disease. Med. Sci. Monit. 2017, 23, 2721–2731. [Google Scholar] [CrossRef]
- Denk, J.; Oberhauser, F.; Kornhuber, J.; Wiltfang, J.; Fassbender, K.; Schroeter, M.L.; Volk, A.E.; Diehl-Schmid, J.; Prudlo, J.; Danek, A.; et al. Specific serum and CSF microRNA profiles distinguish sporadic behavioural variant of frontotemporal dementia compared with Alzheimer patients and cognitively healthy controls. PLoS ONE 2018, 13, e0197329. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Mora, C.; Artigas, M.S.; Garcia-Martínez, I.; Pagerols, M.; Rovira, P.; Richarte, V.; Corrales, M.; Fadeuilhe, C.; Padilla, N.; De La Cruz, X.; et al. Epigenetic signature for attention-deficit/hyperactivity disorder: Identification of miR-26b-5p, miR-185-5p, and miR-191-5p as potential biomarkers in peripheral blood mononuclear cells. Neuropsychopharmacology 2019, 44, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.A.; Benton, M.C.; Lea, R.A.; Maltby, V.E.; Agland, S.; Griffin, N.; Scott, R.J.; Tajouri, L.; Lechner-Scott, J. Next-generation sequencing reveals broad down-regulation of microRNAs in secondary progressive multiple sclerosis CD4+ T cells. Clin. Epigenet. 2016, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qi, Y.; Liu, C.-F.; Lu, J.-M.; Shi, J.; Shi, Y. MicroRNA expression data analysis to identify key miRNAs associated with Alzheimer’s disease. J. Gene Med. 2018, 20, e3014. [Google Scholar] [CrossRef] [PubMed]
- Liguori, M.; Nuzziello, N.; Introna, A.; Consiglio, A.; Licciulli, F.; D’Errico, E.; Scarafino, A.; Distaso, E.; Simone, I.L. Dysregulation of MicroRNAs and Target Genes Networks in Peripheral Blood of Patients With Sporadic Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2018, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Dursun, E.; Candaş, E.; Yılmazer, S.; Gezen-Ak, D. Amyloid Beta 1–42 Alters the Expression of miRNAs in Cortical Neurons. J. Mol. Neurosci. 2019, 67, 181–192. [Google Scholar] [CrossRef]
- Piscopo, P.; Grasso, M.; Puopolo, M.; D’Acunto, E.; Talarico, G.; Crestini, A.; Gasparini, M.; Campopiano, R.; Gambardella, S.; Castellano, A.E.; et al. Circulating miR-127-3p as a Potential Biomarker for Differential Diagnosis in Frontotemporal Dementia. J. Alzheimer’s Dis. 2018, 65, 455–464. [Google Scholar] [CrossRef]
- Boese, A.S.; Saba, R.; Campbell, K.; Majer, A.; Medina, S.; Burton, L.; Booth, T.F.; Chong, P.; Westmacott, G.R.; Dutta, S.M.; et al. MicroRNA abundance is altered in synaptoneurosomes during prion disease. Mol. Cell. Neurosci. 2016, 71, 13–24. [Google Scholar] [CrossRef]
- Regev, K.; Healy, B.C.; Khalid, F.; Paul, A.; Chu, R.; Tauhid, S.; Tummala, S.; Diaz-Cruz, C.; Raheja, R.; Mazzola, M.A.; et al. Association Between Serum MicroRNAs and Magnetic Resonance Imaging Measures of Multiple Sclerosis Severity. JAMA Neurol. 2017, 74, 275–285. [Google Scholar] [CrossRef]
- Qin, S.-B.; Peng, D.-Y.; Lu, J.-M.; Ke, Z.-P. MiR-182-5p inhibited oxidative stress and apoptosis triggered by oxidized low-density lipoprotein via targeting toll-like receptor 4. J. Cell. Physiol. 2018, 233, 6630–6637. [Google Scholar] [CrossRef]
- Raheja, R.; Regev, K.; Healy, B.C.; Mazzola, M.A.; Beynon, V.; Von Glehn, F.; Paul, A.; Diaz-Cruz, C.; Gholipour, T.; Glanz, B.I.; et al. Correlating serum micrornas and clinical parameters in amyotrophic lateral sclerosis. Muscle Nerve 2018, 58, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.; Islam, T.; Turanli, B.; Zaman, T.; Faruquee, H.M.; Rahman, M.; Mollah, N.H.; Nanda, R.K.; Arga, K.Y.; Gov, E.; et al. Network-based approach to identify molecular signatures and therapeutic agents in Alzheimer’s disease. Comput. Biol. Chem. 2019, 78, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Guo, T.; Peng, Y.; Wang, K.; Bai, K.; Huang, Y. Screening of schizophrenia associated miRNAs and the regulation of miR-320a-3p on integrin β1. Medicine 2019, 98, e14332. [Google Scholar] [CrossRef] [PubMed]
- McKeever, P.M.; Schneider, R.; Taghdiri, F.; Weichert, A.; Multani, N.; Brown, R.A.; Boxer, A.L.; Karydas, A.; Miller, B.; Robertson, J.; et al. MicroRNA Expression Levels Are Altered in the Cerebrospinal Fluid of Patients with Young-Onset Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 8826–8841. [Google Scholar] [CrossRef]
- Camkurt, M.A.; Acar Şenel, T.; Coşkun, S.; Gunes, M.L.; Güneş, S.; Yılmaz, M.F.; Görür, A.; Tamer, L. Comparison of plasma MicroRNA levels in drug naive, first episode depressed patients and healthy controls. J. Psychiatr. Res. 2015, 69, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Liu, Y.; Wang, X.; Wu, J.; Liu, K.; Zhou, J.; Liu, L.; Zhang, C. Identification of Differential MicroRNAs in Cerebrospinal Fluid and Serum of Patients with Major Depressive Disorder. PLoS ONE 2015, 10, e0121975. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Exploratory study on microRNA profiles from plasma-derived extracellular vesicles in Alzheimer’s disease and dementia with Lewy bodies. Transl. Neurodegener. 2019, 8, 1–17. [Google Scholar] [CrossRef]
- Hoss, A.G.; Bs, V.N.L.; Frank, S.; Hadzi, T.C.; Myers, R.H.; Dsc, J.C.L. Study of plasma-derived miRNAs mimic differences in Huntington’s disease brain. Mov. Disord. 2015, 30, 1961–1964. [Google Scholar] [CrossRef]
- Hara, N.; Kikuchi, M.; Miyashita, A.; Hatsuta, H.; Saito, Y.; Kasuga, K.; Murayama, S.; Ikeuchi, T.; Kuwano, R. Serum microRNA miR-501-3p as a potential biomarker related to the progression of Alzheimer’s disease. Acta Neuropathol. Commun. 2017, 5, 1–9. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, J.; Wang, Z.; Yao, X.; Wu, X.; Liu, F.; Zheng, W.; Li, Z.; Lin, A. MicroRNAs Correlate with Multiple Sclerosis and Neuromyelitis Optica Spectrum Disorder in a Chinese Population. Med. Sci. Monit. 2017, 23, 2565–2583. [Google Scholar] [CrossRef]
- Mansouri, L.; Lundwall, K.; Moshfegh, A.; Jacobson, S.H.; Lundahl, J.; Spaak, J. Vitamin D receptor activation reduces inflammatory cytokines and plasma MicroRNAs in moderate chronic kidney disease—A randomized trial. BMC Nephrol. 2017, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ander, B.P.; Barger, N.; Stamova, B.; Sharp, F.R.; Schumann, C.M. Atypical miRNA expression in temporal cortex associated with dysregulation of immune, cell cycle, and other pathways in autism spectrum disorders. Mol. Autism 2015, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Rhead, B.; Shao, X.; Graves, J.S.; Chitnis, T.; Waldman, A.T.; Lotze, T.; Schreiner, T.; Belman, A.; Krupp, L.; Greenberg, B.M.; et al. mi RNA contributions to pediatric-onset multiple sclerosis inferred from GWAS. Ann. Clin. Transl. Neurol. 2019, 6, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, M.; Rezaei, N.; Nematalahi, F.S.; Shaygannejad, V.; Fouladi, S.; Karimi, L.; Fathi, F.; Dehghani, L.; Mirmosayyeb, O.; Eskandari, N. MicroRNA-29b variants and MxA expression change during interferon beta therapy in patients with relapsing-remitting multiple sclerosis. Mult. Scler. Relat. Disord. 2019, 35, 241–245. [Google Scholar] [CrossRef]
- Benešová, Y.; Tvaroh, A. Cognition and fatigue in patients with relapsing multiple sclerosis treated by subcutaneous interferon β-1a: An observational study SKORE. Ther. Adv. Neurol. Disord. 2017, 10, 18–32. [Google Scholar] [CrossRef]
- Nunan-Saah, J.H.; Paulraj, S.R.; Waubant, E.; Krupp, L.B.; Gomez, R.G. Neuropsychological correlates of multiple sclerosis across the lifespan. Mult. Scler. J. 2015, 21, 1355–1364. [Google Scholar] [CrossRef]
- Sokolov, A.A.; Grivaz, P.; Bove, R. Cognitive Deficits in Multiple Sclerosis: Recent Advances in Treatment and Neurorehabilitation. Curr. Treat. Options Neurol. 2018, 20, 53. [Google Scholar] [CrossRef]
- Öztürk, Z.; Gucuyener, K.; Acar, A.; Şebnem, S.; Konuşkan, G.D.; Konuşkan, B.; Dikmen, A.U.; Anlar, B. Cognitive functions in pediatric multiple sclerosis: 2-years follow-up. Neurol. Res. 2020, 42, 159–163. [Google Scholar] [CrossRef]
- O’Roak, B.J.; Vives, L.; Fu, W.; Egertson, J.D.; Stanaway, I.B.; Phelps, I.G.; Carvill, G.; Kumar, A.; Lee, C.; Ankenman, K.; et al. Multiplex Targeted Sequencing Identifies Recurrently Mutated Genes in Autism Spectrum Disorders. Science 2012, 338, 1619–1622. [Google Scholar] [CrossRef]
- Yamamoto, T.; Imaizumi, T.; Yamamoto-Shimojima, K.; Lu, Y.; Yanagishita, T.; Shimada, S.; Chong, P.F.; Kira, R.; Ueda, R.; Ishiyama, A.; et al. Genomic backgrounds of Japanese patients with undiagnosed neurodevelopmental disorders. Brain Dev. 2019, 41, 776–782. [Google Scholar] [CrossRef]
- Vardarajan, B.N.; Tosto, G.; Lefort, R.; Yu, L.; Bennett, D.A.; De Jager, P.L.; Barral, S.; Reyes-Dumeyer, D.; Nagy, P.L.; Lee, J.H.; et al. Ultra-rare mutations in SRCAP segregate in Caribbean Hispanic families with Alzheimer disease. Neurol. Genet. 2017, 3, e178. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Fong, T.; Chen, X.; Chen, C.; Luo, P.; Xie, H. Glia maturation factor-β: A potential therapeutic target in neurodegeneration and neuroinflammation. Neuropsychiatr. Dis. Treat. 2018, 14, 495–504. [Google Scholar] [CrossRef] [PubMed]


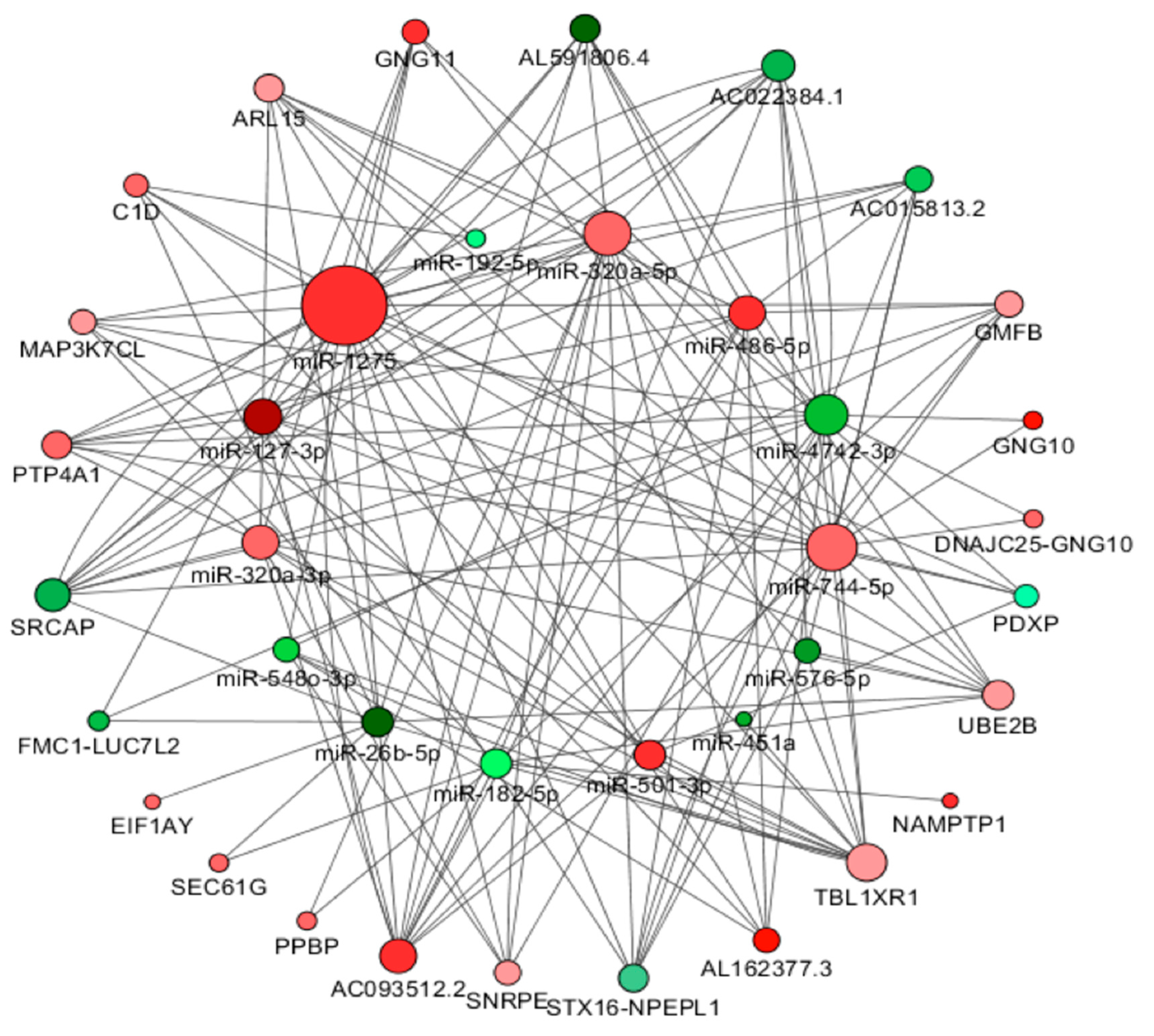{kind=link}
| A. | |||
| PedMS (No. 9) | Mean ± SD | ||
| Age at onset (years) | 14.1 ± 2.1 | ||
| Disease duration (years) | 2.2 ± 2.0 | ||
| EDSS | 2.7 ± 0.9 (range 1.5–4.5) | ||
| FSS | 3.1 ± 2.0 | ||
| CDI | 5.4 ± 1.8 | ||
| Schooling (years) | 10.2 ± 2.0 | ||
| B. | |||
| Cognitive Domains and Tests | Baseline (Mean ± SD) | Follow-Up (Mean ± SD) | |
| Verbal memory | |||
| SRT-LTS | 30.4 ± 12 | 30.8 ± 10.8 | |
| SRT-CLTR | 22 ± 12.6 | 23.6 ± 11.2 | |
| SRT-D | 6.7 ± 2.2 | 6.8 ± 2.1 | |
| Visual-spatial memory | |||
| SPART | 21 ± 4.9 | 20.6 ± 3.2 | |
| SPART-D | 7.3 ± 2.2 | 7.1 ± 1.6 | |
| Attention, IPS | |||
| SDMT | 40.9 ± 7.9 | 42.9 ± 8.9 | |
| TMT-A | 41.8 ± 12.7 | 41.8 ± 11.8 | |
| TMT-B | 97.4 ± 46.3 | 93.6 ± 45.7 | |
| Executive functioning | |||
| TOL | 25.4 ± 5.2 | 25 ± 4.7 | |
| Expressive language | |||
| SVFT | 24.6 ± 10.6 | 22.6 ± 8.6 | |
| PVFT | 15.6 ± 5.7 | 17.4 ± 4.9 | |
| TRANSCRIPT ID | LogFC | Adjusted p-Value | Reported Associations (References, See Supplementary Files) |
|---|---|---|---|
| hsa-miR-26b-5p | 2.254118 | 0.009 | Upregulated in AD [10] Upregulated in AD vs. FTD vs. HC [11] Upregulated in ADHD [12] Downregulated in SPMS [13] Downregulated in AD [14] Downregulated in ALS [15] Related to A-beta expression in cortical neurons animal model [16] |
| hsa-miR-127-3p | −6.144506 | <0.0001 | Downregulated in FTD [17] Upregulated in PPMS exosomes [4] |
| hsa-miR-182-5p | 1.341916 | 0.01 | Upregulated in PedMS [5] Downregulated in prion disease and AD [18] Downregulated in ALS [19] Inhibition of oxidative stress and apoptosis in inflammatory disease [20] |
| hsa-miR-192-5p | 1.14595 | 0.03 | Upregulated in ALS vs. MS [21] Regulatory factor in AD [22] Related to A-beta expression in cortical neurons animal model [16] |
| hsa-miR-320a-3p | −0.862674 | 0.03 | Downregulated in schizophrenia [23] |
| hsa-miR-451a | 1.435193 | 0.02 | Upregulated in RR [4] Upregulated in YOAD [24] Upregulated in depression [25] Downregulated in ALS [15] Downregulated in MDD [26] Downregulated in AD [27] |
| hsa-miR-486-5p | −1.332208 | 0.03 | Upregulated (NC) in MS with low BPV [19] Upregulated in HD [28] |
| hsa-miR-501-3p | −1.562031 | 0.002 | Downregulated in sera AD [29] Upregulated in brain tissue—progression [29] |
| hsa-miR-576-5p | 1.74505 | 0.004 | Upregulated in Relapse MS and ON vs. HC [30] Downregulated in NMOSD [30] Downregulated in active inflammation [31] |
| hsa-miR-744-5p | −0.817595 | 0.04 | |
| hsa-miR-1275 | −1.241061 | 0.01 | |
| hsa-miR-4742-3p | 1.418434 | 0.03 | Downregulated in ASD [32] |
| hsa-miR-548o-3p | 1.391054 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuzziello, N.; Consiglio, A.; Viterbo, R.G.; Licciulli, F.; Liuni, S.; Trojano, M.; Liguori, M. A Pilot Longitudinal Evaluation of MicroRNAs for Monitoring the Cognitive Impairment in Pediatric Multiple Sclerosis. Appl. Sci. 2020, 10, 8274. https://doi.org/10.3390/app10228274
Nuzziello N, Consiglio A, Viterbo RG, Licciulli F, Liuni S, Trojano M, Liguori M. A Pilot Longitudinal Evaluation of MicroRNAs for Monitoring the Cognitive Impairment in Pediatric Multiple Sclerosis. Applied Sciences. 2020; 10(22):8274. https://doi.org/10.3390/app10228274
Chicago/Turabian StyleNuzziello, Nicoletta, Arianna Consiglio, Rosa Gemma Viterbo, Flavio Licciulli, Sabino Liuni, Maria Trojano, and Maria Liguori. 2020. "A Pilot Longitudinal Evaluation of MicroRNAs for Monitoring the Cognitive Impairment in Pediatric Multiple Sclerosis" Applied Sciences 10, no. 22: 8274. https://doi.org/10.3390/app10228274
APA StyleNuzziello, N., Consiglio, A., Viterbo, R. G., Licciulli, F., Liuni, S., Trojano, M., & Liguori, M. (2020). A Pilot Longitudinal Evaluation of MicroRNAs for Monitoring the Cognitive Impairment in Pediatric Multiple Sclerosis. Applied Sciences, 10(22), 8274. https://doi.org/10.3390/app10228274