Abstract
The observation of a much-improved fitness of wild-type plants over abscisic acid (ABA)-deficient mutants during drought has led researchers from all over to world to perform experiments aiming at a better understanding of how this hormone modulates the physiology of plants under water-limited conditions. More recently, several promising approaches manipulating ABA biosynthesis and signaling have been explored to improve water use efficiency and confer drought tolerance to major crop species. Here, we review recent progress made in the last decade on (i) ABA biosynthesis, (ii) the roles of ABA on plant-water relations and on primary and secondary metabolisms during drought, and (iii) the regulation of ABA levels and perception to improve water use efficiency and drought tolerance in crop species.
1. Introduction
The phytohormone abscisic acid (ABA) is a fundamental regulator of the morpho-physiological response of plants during drought stress. The central and best-known plant response to ABA during drought is the closure of stomata in seed plants [1,2]. However, this hormone regulates many other aspects of plant-water relations (Figure 1). For instance, many studies have demonstrated a pronounced inhibitory effect of increased levels of endogenous ABA on shoot growth in plants experiencing drought [3,4]. At the same time, high levels of ABA promote root elongation [5], while strongly inhibiting lateral root formation [6] as an adaptive response to increase the effectiveness of root water uptake under water deficit conditions. The effects of ABA on shoot and root growth, however, can be positive or negative depending on its concentration and the plant species [7]. Increased levels of endogenous ABA also play key roles in down-regulating leaf hydraulic conductance (which is proposed to further induce stomatal closure) [7,8,9] and mesophyll conductances [10,11,12], as well as up-regulating the cuticular wax formation (which results in a thicker, less permeable cuticle) [13,14]. Finally, ABA is a critical messenger produced in response to water deficit that promotes a variety of biochemical responses in different plant tissues [15]. Due to the ABA’s key roles in the physiology of plants during drought, several promising approaches to manipulate ABA biosynthesis and signaling (i.e., either genetically or through chemical intervention with agonists) have been explored aiming at conferring drought tolerance to major crop species and improving water use efficiency (WUE, i.e., the ratio of photosynthesis to water loss through foliar transpiration). In this review, we cover the progress made in the last decade in our understanding of how ABA signaling regulates plant physiology and biochemistry during drought. Special attention has been paid to (i) the ABA biosynthesis within the plant, (ii) the impacts of ABA on stomatal aperture and xylem embolism, (iii) the regulation of primary and secondary metabolisms by ABA, and (iv) the potential applications of approaches modulating ABA levels and perception to improve WUE and drought tolerance in crop species.
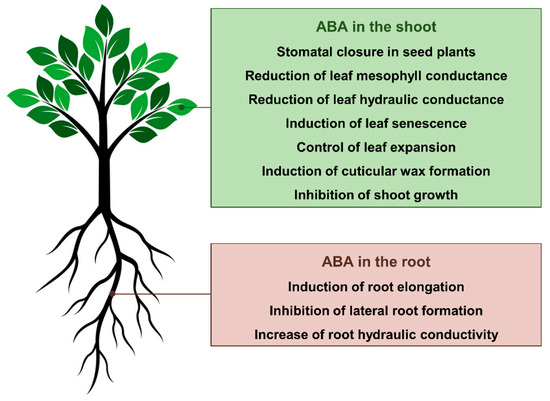
Figure 1.
Current understanding of how abscisic acid (ABA) regulates plant form and function during drought.
2. Drought-Induced Biosynthesis of ABA
Given the key role played by ABA in enhancing plant survival during drought, a comprehensive understanding of how the biosynthesis of ABA occurs, how it is regulated within the plant, and where it mostly takes place is of major importance. In recent years, the pathway by which ABA is synthesized in plants has been extensively investigated through the use of mutants deficient in the synthesis and perception of ABA [16,17,18,19,20]. It has been demonstrated that the biosynthetic pathway for ABA accumulation begins in the chloroplasts with the hydroxylation of β-carotene to zeaxanthin, which is then converted to violaxanthin through the xanthophyll cycle. Next, neoxanthin synthase converts violaxanthin to neoxanthin, which is then isomerized from 9-trans to 9-cis (neoxanthin). The last step of the biosynthetic pathway that occurs in the chloroplasts is the oxidative cleavage of 9-cis-neoxanthin and/or 9-cis-violaxanthin to xanthoxin, which is further catalyzed by 9-cis-epoxycarotenoid dioxygenase (NCED). NCED is known to be the key rate-limiting enzyme in the ABA biosynthesis and it is responsible for catalyzing the first non-reversible step in the pathway [21]. The formed xanthoxin then moves from the chloroplasts to the cytosol, where it is converted to abscisic aldehyde, and finally to ABA. ABA accumulation in plants occurs not only through activation of ABA biosynthesis but also by the hydrolysis of its glycosyl ester (i.e., ABA-GE) [22,23], which can be utilized to maintain the high levels of ABA in plants during drought [24,25,26].
Since the first studies linking ABA with stomatal closure under drought, a number of results have led to the critical assumption that the pool of ABA resulting in stomatal closure in leaves is mostly synthesized in the root tips [27,28]. However, this conventional view of root-to-shoot ABA signaling has recently been challenged by studies using a number of experimental approaches, including the reciprocal grafting between ABA biosynthetic mutant and wild-type plants and exogenous labeled ABA [29,30,31,32,33]. In fact, these studies clearly demonstrate that although multiple organs and tissues can accumulate small amounts of ABA, leaves are the predominant site for ABA biosynthesis, and they are even responsible to export ABA to the roots, maintaining normal root levels of ABA levels and determining root architecture and growth [30,32,33]. Inside leaves, phloem companion cells, guard cells, and mesophyll cells have been demonstrated to be capable of synthesizing ABA [34,35,36,37]. In addition, ABA biosynthesis has been recently observed to occur in parenchyma cells around the vascular tissues [38]. Regarding the ABA transport within plants, the recent identification of a number of transmembrane ABA transporters strongly suggests that the movement of this hormone is actively regulated in an intercellular network [15]. However, the fine regulation of ABA biosynthesis and transport among different plant tissues remains a matter of debate [15].
The major production of ABA in leaves, not roots, during drought seems considerably more advantageous for plants to avoid excessive declines in water potential as leaves represent the main organ exchanging (and mostly losing) water with the atmosphere. For instance, a de novo biosynthesis of ABA over an extremely short time-frame following leaf exposure to high vapor pressure deficit (VPD) has been demonstrated to allow an efficient stomatal closure in angiosperm species [39,40]. Declines in leaf water potential to a threshold leaf water potential results in major increases in foliar ABA [41,42] and such threshold water potential strongly coincides with bulk leaf turgor loss point [20,40,43,44]. Although the water potential at turgor loss is broadly accepted as the threshold for the major increment in foliar ABA biosynthesis, it is still uncertain whether changes in turgor itself or changes in cell volume are the main signal up-regulating the enzymes responsible for ABA production [20,45]. In addition, given the lower water potential in the guard cells and the mesophyll cells nearby stomata in transpiring leaves [46], small levels of ABA are expected to be produced in these cells as transpirational demand increases, resulting in stomatal closure even prior to leaf turgor loss point [40].
3. ABA and Drought Tolerance: From Stomata Closure to the Modulation of Plant Metabolism
As previously mentioned, guard cells are not the only site of ABA signaling and action. Instead, ABA plays crucial roles in transducing the water stress signal to a variety of cell-types and tissues [47]. For instance, ABA accumulation activates seed dormancy in developing embryos [48,49,50], leaf osmotic adjustment inducing compatible solute accumulation (e.g., proline and sucrose [51,52,53]), and leaf senescence [54,55,56]. In addition, ABA has a protective effect against oxidative damage by enhancing the activity and expression of antioxidant enzymes and the levels of secondary metabolites in different plant organs [38,57,58,59]. Amongst the multiple processes controlled by ABA under drought, we describe here the regulation of gas exchange through the stomata and how it affects the water transport within the xylem. In addition, potential roles of ABA on the modulation of primary and secondary metabolites are also discussed.
3.1. Linking ABA with Stomatal Aperture and Xylem Embolism
Stomatal closure during drought is one of the first attempts of vascular plants to reduce transpiration, preventing tissue desiccation and damage [60,61]. The signaling pathway for ABA-induced stomatal closure in angiosperm species has been the subject of detailed investigations in recent years and it is now reasonably well characterized [62,63,64,65]. As angiosperms often exhibit stomatal conductances higher than the minimum amount sufficient to allow maximum rates of photosynthesis, declines in stomatal conductances during drought driven by increased foliar ABA levels often result in optimized WUE [66,67]. Aiming to increase WUE in economically important crop species, genetic and molecular tools have been applied to generate plants with increased ABA biosynthesis and signaling in recent years [66,68,69]. See topic 4.0 for further details.
Although increasing levels of foliar ABA has long been associated with stomatal closure and increases in WUE in angiosperm species during drought [1,38,67,70,71], it has recently been discovered that the stomata of ferns and lycophytes do not close in response to both endogenous and exogenous ABA [72,73,74]. Instead, in these early-diverging species, changes in leaf water status regulate stomatal aperture by an ABA-independent, hydropassive mechanism [72,75,76]. In a similar way, recent findings demonstrate a number of hornwort species that lack stomatal responses to ABA [77]. These findings indicate a gradualistic model of stomatal control, which has been the focus of considerable debate in the last decade [76,78,79,80,81,82,83]. For a recent and comprehensive review on ABA signaling pathway and function across the evolution of land plants, see McAdam and Sussmilch [84].
Early closure of stomata has been demonstrated to occur prior to leaf and stem xylem embolism in a number of species from ferns to angiosperms, supporting the idea of the water-saving function of stomata and xylem protection during drought [61,76,85,86,87]. This protective mechanism is of paramount importance given that leaf xylem embolism has been associated with foliar mortality across the canopy [88,89], potentially affecting plant growth and productivity. In addition, extensive embolism in the stem xylem and consequent hydraulic failure have been largely associated with drought-induced mortality of plants [90,91,92,93]. Curiously, ABA signaling has very recently been associated with the construction of conduits of stem xylem more prone to suffer embolism in a study comparing a transgenic line of tomato overproducing ABA with its wild-type [94]. The specific ABA over-expressing transgenic line utilized in this study, however, exhibits several differences from the wild-type plants regarding stem xylem anatomy traits that are closely linked to xylem resistance to embolism (i.e., xylem conduits are much less numerous and narrower in the transgenic line compared with the wild-type). Whether this represents that ABA signaling modulates xylem anatomy, or if this arises from pleiotropic effects is of major importance to confirm the association between ABA and xylem vulnerability to embolism. Besides, the transgenic line utilized in this study presents a very similar hydraulic resistance to other cultivars of tomato that do not overproduce ABA [95] (and personal observations), which might suggest that the wild-type utilized, rather than the transgenic line itself, presents a different xylem vulnerability from other cultivars potentially due to differences in the xylem anatomy. In any case, the idea of hydraulic vulnerable plants when ABA biosynthesis is increased might challenge the use of transgenic plants with increased ABA biosynthesis and signaling to improve crop production under drought and thus should be further investigated.
3.2. Primary and Secondary Metabolic Pathways Regulated by ABA
ABA signaling is involved in transcriptional and post-transcriptional regulation of genes encoded for enzymes related to carbohydrate metabolism (Table 1) [96,97,98]. ABA can regulate sucrose metabolism through the induction of vacuolar invertase activity and the up-regulation of vacuolar invertase gene expresion [97,99]. Furthermore, Thalmann et al. [100] showed that ABA promotes leaf starch degradation under osmotic stress by the synergistic action of β-AMYLASE1 (BAM1) and α-AMYLASE3 (AMY3). Both mechanisms lead to the release of hexoses (i.e., fructose, glucose, and maltose) that function as compatible solutes, preserving membrane integrity and maintaining leaf turgor [101]. The accumulation of soluble carbohydrates concomitantly with the up-regulation of ABA biosynthesis in stem of Populus nigra exposed to drought has been recently observed [38,102]. Stem soluble carbohydrates may help to preserve xylem hydraulic integrity and act as a buffer when photoassimilation decreases under drought, thus ensuring an adequate energy source for stress defense and viable seed production [103,104]. In addition, ABA has been demonstrated to regulate foliar proline accumulation under osmotic stress (Table 1), although a direct action of ABA on the transcriptional regulation of the key genes and enzymes involved in proline biosynthesis is still unclear [59,105,106,107,108,109]. Similar to other compatible osmolytes, significant levels of proline can accumulate in the cytoplasm and organelles allowing osmotic adjustment, stabilizing protein structure and cell membranes, and acting as a free radical scavenger [110]. Furthermore, proline may provide regulation of cytosolic acidity and act as a signaling molecule able to activate further stress responses [111].

Table 1.
Representative metabolites and enzymes associated with primary and secondary metabolisms that are affected by ABA.
Regarding the regulation of secondary metabolites under water deficit, several studies have shown the impact of exogenous application of ABA on flavonoid biosynthetic pathway (Table 1) [59,112,113,114]. In addition, many key genes of the flavonoid biosynthetic pathway were demonstrated to be up-regulated both in fruits and leaves in parallel to increases in the ABA content during drought [115,116,117]. However, whether these genes are activated by ABA or by the stress conditions remains under debate. The possible relationship between ABA and flavonoid could be important due to the ability of flavonoids to modulate the reactive oxygen species-signaling cascade and to buffer stress-induced alterations in metabolic homeostasis [118]. Future studies focusing on ABA-flavonoid interaction utilizing ABA-deficient mutants or any additional tools to manipulate the endogenous levels of ABA are necessary in order to confirm the role of ABA on flavonoid biosynthesis both at the transcriptional and post-transcriptional levels.
4. Regulation of ABA Levels and Perception in Crop Species
Ongoing climate change coupled with the projected increase in human population by over 2.4 billion until 2050 [119,120] represents a major challenge for the agricultural sector [121]. Over the next decades, climate change is expected to result in considerable rise in the intensity and frequency of drought events. In this water-limited scenario, achieving ‘more crop per drop’ (i.e., crop water productivity that is defined as the mass of agricultural produce per unit of water consumed) is a critical target for food production. Especially when considering that 70% of the available fresh water is used for crop production and that drought is expected to cause the largest yield reductions relative to any other abiotic and biotic stresses [122]. Therefore, enhancing crop WUE is still foreseen as a good means for saving water in agriculture [123] and has proven to be an accurate target in both genotype selection [124,125,126] and engineering [127].
The WUE can be determined both at the plant and leaf levels. At the leaf level, the photosynthesis-to-stomatal conductance ratio is defined as the intrinsic water-use efficiency (WUEi). This parameter is typically determined in studies aimed at improving crop yield while saving soil water and mitigating yield limitations by water deficit [127,128]. In this context, the importance of ABA for WUEi improvement is well-known due to its central role in regulating plant-water relations, specially by modulating stomatal conductance in several crop species [66,129,130,131]. As outlined above, the direct influence of ABA on WUEi in angiosperms has already been demonstrated and several studies have concluded that plant responses driven by ABA depend on the severity and the duration of the drought events [67]. Particularly under short-term water deficit, ABA induces stomatal closure through its direct action on guard cells and modulates mesophyll conductance without restraining CO2 fixation [8,12,38]. During severe and prolonged drought conditions, ABA can trigger further alterations at the transcriptome level, including genes encoding for LEA (late embryogenesis abundant) proteins [132]. Besides, it can alter the levels of osmoproctectans [100] and cause morphological changes in stomatal size and density [133], leaf size, and shoot and root development [7,134]. All these alterations strongly affect plant water balance and WUE in the long-term [135]. In addition, ABA have been observed to regulate WUE through a tissue-specific action on hydraulic conductivity. Particularly in leaves, ABA has been found to decrease the hydraulic conductance by reducing the permeability of bundle sheath aquaporins [8,9,136] and also indirectly controlling mesophyll hydraulic conductances [11,137]. In the roots, ABA has been observed to increase hydraulic conductivity, facilitating water uptake under non-transpiring conditions [7,94]. Therefore, ABA signaling network can be modulated to reduce transpiration and increase crop water productivity under stressful conditions [67,138].
The ABA signal transduction pathway has recently been well documented, consisting of pyrabactin resistance/pyrabactin resistance-like/regulatory component of ABA receptors (PYR/PYL/RCAR), clade A type 2C protein phosphatases (PP2Cs), and SNFl related protein kinase 2 (SnRK2s) [139,140]. The first attempts to use ABA in large scale agriculture were constrained by the chemical instability of ABA (e.g., under UV-light), its relatively expensive production, and its rapid cellular catabolism [141]. The modulation of ABA signaling, however, can be efficiently achieved through genetically engineered plants with an overexpression of ABA receptors [54,69,127,142] and signal transduction components [143,144], as well as through chemical intervention with ABA agonists [145,146,147,148,149].
Over the last years, studies on ABA overexpression receptors have led to promising results both in the field [150,151] and under controlled conditions [127,142]. The overexpression of PYL/RCAR receptors induces a higher plant sensitivity to ABA, leading to a globally enhanced WUE and drought resistance, as previously demonstrated in rice, poplar, and wheat [69,142,152,153]. The actual challenge, however, is to balance the improvements in WUE against the negative effects on growth due to ABA oversensitivity. Recent studies on plants overexpressing PYL receptors have shown an opposite effect on growth and stress adaptation and this may be due to the specific receptor choice. Indeed, the overexpression of some specific subfamilies result in plants with less transpiration for a similar leaf area and biomass compared to the wild-types, thus leading to an increased WUE [138]. In particular, PYL12/RCAR6 and PYL4/RCAR10 (subfamily I and II) overexpressing lines in Arabidopsis possessed a balance of reduced water use with negligible effects on growth and a “water-saving” phenotype that resulted in a significant improvement in total biomass relative to water use gains [127]. Similar observations have been made in transgenic wheat and in rice, in which the overexpression of specific PYL receptors has led to a reduced transpiration and a concomitant increase in photosynthetic activity compared to the wild-type, improving grain production per liter of water and protecting productivity during water deficit [69,142,151].
A faster and a more manageable alternative to the expression of genes for increasing crop’s WUE and conferring drought resistance is the use of ABA agonists [147,154], which are small synthetic molecules binding ABA receptors and activating ABA signaling pathway [155]. Among the ABA agonists, the first synthetized molecule Pyrabactin has been identified in a chemical genetic screen for seed germination inhibitors and it has been instrumental in showing how ABA binds the PYR/PYL receptor family [140]. The direct application of this molecule is not practical for agricultural purposes since its major effects are in seeds rather than in vegetative tissues. However, the examination of the structure of Pyrobactin and its interaction with ABA receptors has provided a framework for the design of novel ABA agonists [154]. As a consequence, several ABA agonists have been developed during the last ten years, with promising results in increasing crop water productivity [68]. An example is represented by Quinabactin, which appeared to be an overall better ABA agonist than Pyrabactin, inducing ABA responses both in seeds and vegetative tissues. Application of Quinabactin in plants resulted in improved water use upon drought stress [145]. This molecule targets all of the dimeric subfamily III ABA receptors, affecting guard cell closure to prevent water loss from detached leaves and conferring drought tolerance in both Arabidopsis and soybean during drought [145]. In addition, Quinabactin has shown effect in rapeseed and tomato, inhibiting germination and eliciting drought stress responses [156,157,158]. Another example concerns a tetrafluoro derivative of Quinabactin, AMF4, which seems to be more effective than Quinabactin due to its higher stability and bioavailability, even targeting only a subset of ABA receptors [145,159]. Finally, a recent example is represented by the agonist B2, which was shown to improve drought tolerance in wheat by increasing root biomass and preventing leaf dehydration as well as amplifying antioxidant responses and enhancing the photosynthetic performances [149].
A recent innovative approach aims at combining the application of agrochemicals on genetically modified crops such as AMF4 on PYL2 overexpressed plants or mandipropamid on engineered PYR1 receptor (PYR1MANDI) [146]. The mandipropamid-PYR1MANDI system resulted in increased seedling survival during drought and transcriptional responses similar to those induced by ABA. Recently, Cao et al. [159] optimized the synthesis of new molecules based on the Quinabactin backbone and applied them to transgenic Arabidopsis and soybean plants with an abiotic stress-inducible AtPYL2 overexpression. This combined approach dramatically increased drought resistance in these plants, making this system a compelling alternative strategy to manipulate plant water use. Although the utilization of agrochemicals acting as ABA receptor agonists combined with transgenic approaches to increase ABA signaling holds great promise for the production of plants with enhanced WUE, the translation of these applications at field level into practical improvements in crop yield can be challenging [160,161]. For instance, as previously stated, a recent study conducted on tomato has highlighted a higher vulnerability to embolism of the vascular system of the transgenic line sp12 which overproduces ABA [94]. A reduced hydraulic safety margin can compromise the maintenance of hydraulic conductivity during periods of soil water deficit, with negative consequences not only for photosynthesis and productivity, but also for plant survival [162]. Given the importance of the potential use of plants with increased ABA biosynthesis and signaling to the future of agriculture in a world facing an ongoing climate change, it is of critical importance to confirm whether increased levels of ABA result in more vulnerable xylem and to understand the underlying mechanisms linking ABA signaling and xylem functioning. Important questions remain unanswered, especially those related to the extent of variation in embolism resistance among single crop species and their different varieties [163,164], including the relationship with genotypic variations in endogenous ABA content. Therefore, to optimize WUE strategies in the field, a complete understanding of the changes at the hydraulic and metabolic levels induced by the manipulation of ABA levels is needed. Timely investments in research at different levels will likely allow to improve crop resilience and yields in a water-limited scenario.
Author Contributions
A.A.C. wrote the abstract and Section 1 and Section 3.1; C.J.D.-S. wrote Section 2; A.G. wrote Section 4; C.B. wrote Section 3.2. All authors read and approved the final version of the review. All authors have read and agreed to the published version of the manuscript.
Funding
A.A.C. and C.J.D.-S. acknowledge postdoctoral fellowships (Finance Code 001) granted by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mittelheuser, C.J.; Van Steveninck, R.F.M. Stomatal closure and inhibition of transpiration induced by (RS)-abscisic acid. Nature 1969, 221, 281–282. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.M. Evolution of the stomatal regulation of plant water content. Plant Physiol. 2017, 174, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Parent, B.; Simonneau, T. Control of leaf growth by abscisic acid: Hydraulic or non-hydraulic processes? Plant Cell Environ. 2010, 33, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Finlayson, S.A. Abscisic acid is a general negative regulator of Arabidopsis axillary bud growth. Plant Physiol. 2015, 169, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, J.; He, J.; Qin, Y.; Hua, D.; Duan, Y.; Chen, Z.; Gong, Z. ABA-mediated ROS in mitochondria regulate root meristem activity by controlling PLETHORA expression in Arabidopsis. PLoS Genet. 2014, 10, e1004791. [Google Scholar] [CrossRef] [PubMed]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.; Testerink, C. Roots withstanding their environment: Exploiting root system architecture responses to abiotic stress to improve crop tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef]
- Rosales, M.; Maurel, C.; Nacry, P. Abscisic acid coordinates dose-dependent developmental and hydraulic responses of roots to water deficit. Plant Physiol. 2019, 180, 2198–2211. [Google Scholar] [CrossRef]
- Pantin, F.; Monnet, F.; Jannaud, D.; Costa, J.M.; Renaud, J.; Muller, B.; Simonneau, T.; Genty, B. The dual effect of abscisic acid on stomata. New Phytol. 2013, 197, 65–72. [Google Scholar] [CrossRef]
- Coupel-Ledru, A.; Tyerman, S.D.; Masclef, D.; Lebon, E.; Christophe, A.; Edwards, E.J.; Simonneau, T. Abscisic acid down-regulates hydraulic conductance of grapevine leaves in isohydric genotypes only. Plant Physiol. 2017, 175, 1121–1134. [Google Scholar] [CrossRef]
- Mizokami, Y.; Noguchi, K.; Kojima, M.; Sakakibara, M.; Terashima, I. Effects of instantaneous and growth CO2 levels and abscisic acid on stomatal and mesophyll conductances. Plant Cell Environ. 2018, 42, 1257–1269. [Google Scholar] [CrossRef]
- Sade, N.; Galle, A.; Flexas, J.; Lerner, S.; Peleg, G.; Yaaran, A.; Moshelion, M. Differential tissue-specific expression of NtAQP1 in Arabidopsis thaliana reveals a role for this protein in stomatal and mesophyll conductance of CO2 under standard and salt-stress conditions. Planta 2014, 239, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, G.; Haworth, M.; Said Wahbi, T.M.; Zuomin, S.; Centritto, M. Abscisic acid induces rapid reductions in mesophyll conductance to carbon dioxide. PLoS ONE 2016, 11, e0148554. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Bourdenx, B.; Bernard, A.; Parsons, E.P.; Lü, S.; Joubès, J.; Jenks, M.A. The impact of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol. 2009, 151, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Xiong, L.; Li, W.; Zhu, J.-K.; Zhu, J. The plant cuticle is required for osmotic stress regulation of abscisic acid biosynthesis and osmotic stress tolerance in Arabidopsis. Plant Cell 2011, 23, 1971–1984. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA transport and plant water stress responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Taylor, I.B.; Sonneveld, T.; Bugg, T.D.H.; Thompson, A.J. Regulation and manipulation of the biosynthesis of abscisic acid, including the supply of xanthophyll precursors. J. Plant Growth Regul. 2005, 24, 253–273. [Google Scholar] [CrossRef]
- Neuman, H.; Galpaz, N.; Cunningham, F.X.J.; Zamir, D.; Hirschberg, J. The tomato mutation nxd1 reveals a gene necessary for neoxanthin biosynthesis and demonstrates that violaxanthin is a sufficient precursor for abscisic acid biosynthesis. Plant J. 2014, 78, 80–93. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Sussmilch, F.C.; Brodribb, T.J.; Ross, J.J. Molecular characterization of a mutation affecting abscisic acid biosynthesis and consequently stomatal responses to humidity in an agriculturally important species. AoB Plants 2015, 27, 7. [Google Scholar] [CrossRef]
- Sussmilch, F.C.; Brodribb, T.J.; McAdam, S.A.M. Up-regulation of NCED3 and ABA biosynthesis occur within minutes of a decrease in leaf turgor but AHK1 is not required. J. Exp. Bot. 2017, 68, 2913–2918. [Google Scholar] [CrossRef]
- Thompson, A.J.; Jackson, A.C.; Symonds, R.C.; Mulholland, B.J.; Dadswell, A.R.; Blake, P.S.; Burbidge, A.; Taylor, I.B. Ectopic expression of a tomato 9-cis-epoxycarotenoid dioxygenase gene causes overproduction of abscisic acid. Plant J. 2000, 23, 363–374. [Google Scholar] [CrossRef]
- Dietz, K.J.; Sauter, A.; Wichert, K.; Messdaghi, D.; Hartung, W. Extracellular β-glucosidase activity in barley involved in the hydrolysis of ABA glucose conjugate in leaves. J. Exp. Bot. 2000, 51, 937–944. [Google Scholar] [CrossRef]
- Seiler, C.; Harshavardhan, V.T.; Rajesh, K.; Reddy, P.S.; Strickert, M.; Rolletschek, H.; Scholz, U.; Wobus, U.; Sreenivasulu, N. ABA biosynthesis and degradation contributing to ABA homeostasis during barley seed development under control and terminal drought-stress conditions. J. Exp. Bot. 2011, 62, 2615–2632. [Google Scholar] [CrossRef]
- Lee, K.H.; Piao, H.L.; Kim, H.-Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.-J.; Hwang, I. Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhang, B.; Su, X.; Zhang, S.; Huang, M. Reference gene selection for quantitative real-time polymerase chain reaction in Populus. Anal. Biochem. 2011, 408, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Watanabe, S.; Shimada, H.; Sakamoto, A. Dynamics of the leaf endoplasmic reticulum modulate β-glucosidase-mediated stress-activated ABA production from its glucosyl ester. J. Exp. Bot. 2020, 71, 2058–2071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Schurr, U.; Davies, W.J. Control of stomatal behaviour by abscisic acid which apparently originates in the roots. J. Exp. Bot. 1987, 38, 1174–1181. [Google Scholar] [CrossRef]
- Zhang, J.; Davies, W.J. Abscisic acid produced in dehydrating roots may enable the plant tomeasure the water status of the soil. Plant Cell Environ. 1989, 12, 73–81. [Google Scholar] [CrossRef]
- Holbrook, N.M.; Shashidhar, V.R.; James, R.A.; Munns, R. Stomatal control in tomato with ABA-deficient roots: Response of grafted plants to soil drying. J. Exp. Bot. 2002, 53, 1503–1514. [Google Scholar]
- Manzi, M.; Lado, J.; Rodrigo, M.J.; Zacarías, L.; Arbona, V.; Gómez-Cadenas, A. Root ABA accumulation in long-term water-stressed plants is sustained by hormone transport from aerial organs. Plant Cell Physiol. 2015, 56, 2457–2466. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Brodribb, T.J.; Ross, J.J. Shoot-derived abscisic acid promotes root growth. Plant Cell Environ. 2016, 39, 652–659. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Manzi, M.; Ross, J.J.; Brodribb, T.J.; Gómez-Cadenas, A. Uprooting an abscisic acid paradigm: Shoots are the primary source. Plant Signal. Behav. 2016, 11, 11–12. [Google Scholar] [CrossRef]
- Zhang, F.; Sussmilch, F.; Nichols, D.S.; Cardoso, A.A.; Brodribb, T.J.; McAdam, S.A.M. Leaves, not roots or floral tissue, are the main site of rapid, external pressure-induced ABA biosynthesis in angiosperms. J. Exp. Bot. 2018, 69, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Bauer, H.; Ache, P.; Lautner, S.; Fromm, J.; Hartung, W.; Al-Rasheid, K.A.S.; Sonnewald, S.; Sonnewald, U.; Kneitz, S.; Lachmann, N.; et al. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr. Biol. 2013, 23, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Intertissue signal transfer of abscisic acid from vascular cells to guard cells. Plant Physiol. 2014, 164, 587–1592. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Brodribb, T.J. Mesophyll cells are the main site of abscisic acid. Plant Physiol. 2018, 177, 911–917. [Google Scholar] [CrossRef]
- Merilo, E.; Yarmolinsky, D.; Jalakas, P.; Parik, H.; Tulva, I.; Rasulov, B.; Kilk, K.; Kollist, H. Stomatal VPD response: There is more to the story than ABA. Plant Physiol. 2018, 176, 851–864. [Google Scholar] [CrossRef]
- Brunetti, C.; Gori, A.; Marino, G.; Latini, P.; Sobolev, A.P.; Nardini, A.; Haworth, M.; Giovannelli, A.; Capitani, D.; Loreto, F.; et al. Dynamic changes in ABA content in water-stressed Populus nigra: Effects on carbon fixation and soluble carbohydrates. Ann. Bot. 2019, 124, 627–643. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Sussmilch, F.C.; Brodribb, T.J. Stomatal responses to vapour pressure deficit are regulated by high speed gene expression in angiosperms. Plant Cell Environ. 2016, 39, 485–491. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Brodribb, T.J.; Kane, C.N.; DaMatta, F.M.; McAdam, S.A.M. Osmotic adjustment and hormonal regulation of stomatal responses to vapour pressure deficit in sunflower. AoB Plants 2020, 12, plaa025. [Google Scholar] [CrossRef]
- Zabadal, T.J. A water potential threshold for the increase of abscisic acid in leaves. Plant Physiol. 1974, 53, 125–127. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Beardsell, M.F.; Cohen, D. Relationships between leaf water status, abscisic acid levels, and stomatal resistance in maize and sorghum. Plant Physiol. 1975, 56, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Pierce, M.; Raschke, K. Synthesis and metabolism of abscisic acid in detached leaves of Phaseolus vulgaris L. after loss and recovery of turgor. Planta 1981, 153, 156–165. [Google Scholar] [CrossRef] [PubMed]
- McAdam, S.A.M.; Brodribb, T.J. Linking turgor with ABA biosynthesis: Implications for stomatal responses to vapor pressure deficit across land plants. Plant Physiol. 2016, 171, 2008–2016. [Google Scholar] [CrossRef]
- Sack, L.; John, G.P.; Buckley, T.N. ABA accumulation in dehydrating leaves is associated with decline in cell volume not turgor pressure. Plant Physiol. 2018, 176, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Buckley, T.N.; John, G.P.; Scoffoni, C.; Sack, L. How does leaf anatomy influence water transport outside the xylem? Plant Physiol. 2015, 168, 1616–1635. [Google Scholar] [CrossRef]
- Jones, A.M. A new look at stress: Abscisic acid patterns and dynamics at high-resolution. New Phytol. 2016, 38–44. [Google Scholar] [CrossRef]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. Book 2013, 11, e0166. [Google Scholar] [CrossRef]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2015, 6, 8113. [Google Scholar] [CrossRef]
- Verslues, P.E.; Bray, E.A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. J. Exp. Bot. 2006, 57, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Rajasheker, G.; Jawahar, G.; Jalaja, N.; Kumar, S.A.; Kumari, P.H.; Punita, D.L.; Karumanchi, A.R.; Reddy, P.S.; Rathnagiri, P.; Sreenivasulu, N.; et al. Role and regulation of osmolytes and ABA interaction in salt and drought stress tolerance. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 417–436. [Google Scholar]
- Zhang, P.; Yuan, L.; Mengfei, L.; Jingfu, M.; Caixiang, W.; Junji, S.; Delong, Y. Abscisic acid associated with key enzymes and genes involving in dynamic flux of water soluble carbohydrates in wheat peduncle under terminal drought stress. Plant Physiol. Biochem. 2020, 151, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.C.; Hong, S.W.; Whang, S.S.; Lim, P.O.; Nam, H.G.; Koo, J.C. Age-dependent action of an ABA-inducible receptor kinase, RPK1, as a positive regulator of senescence in Arabidopsis leaves. Plant Cell Physiol. 2011, 52, 651–662. [Google Scholar] [CrossRef]
- Asad, M.A.U.; Zakari, S.A.; Zhao, Q.; Zhou, L.; Ye, Y.; Cheng, F. Abiotic stresses intervene with ABA signaling to induce destructive metabolic pathways leading to death: Premature leaf senescence in plants. Int. J. Mol. Sci. 2019, 20, 256. [Google Scholar] [CrossRef]
- Bano, A.; Ullah, F.; Nosheen, A. Role of abscisic acid and drought stress on the activities of antioxidant enzymes in wheat. Plant Soil Environ. 2012, 58, 181–185. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef]
- Murcia, G.; Fontana, A.; Pontin, M.; Baraldi, R.; Bertazza, G.; Piccoli, P.N. ABA and GA3 regulate the synthesis of primary and secondary metabolites related to alleviation from biotic and abiotic stresses in grapevine. Phytochemistry 2017, 135, 34–52. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Holbrook, N.M. Stomatal protection against hydraulic failure: A comparison of coexisting ferns and angiosperms. New Phytol. 2004, 162, 663–670. [Google Scholar] [CrossRef]
- Martin-StPaul, N.; Delzon, S.; Cochard, H. Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 2017, 20, 1437–1447. [Google Scholar] [CrossRef]
- Geiger, D.; Maierhofer, T.; Al-Rasheid, K.A.S.; Scherzer, S.; Mumm, P.; Liese, A.; Ache, P.; Wellmann, C.; Marten, I.; Grill, E.; et al. Stomatal closure by fast abscisic acid signaling is mediated by the guard cell anion channel SLAH3 and the receptor RCAR1. Sci. Signal. 2011, 4, ra32. [Google Scholar] [CrossRef] [PubMed]
- Wege, S.; De Angeli, A.; Droillard, M.-J.; Kroniewicz, L.; Merlot, S.; Cornu, D.; Gambale, F.; Martinoia, E.; Barbier-Brygoo, H.; Thomine, S.; et al. Phosphorylation of the vacuolar anion exchanger AtCLCa is required for the stomatal response to abscisic acid. Sci. Signal. 2014, 7, ra65. [Google Scholar] [CrossRef]
- Grondin, A.; Rodrigues, O.; Verdoucq, L.; Merlot, S.; Leonhardt, N.; Maurel, C. Aquaporins contribute to ABA-triggered stomatal closure through OST1-mediated phosphorylation. Plant Cell 2015, 27, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Eisenach, C.; Baetz, U.; Huck, N.V.; Zhang, J.; De Angeli, A.; Beckers, G.J.; Martinoia, E. ABA-induced stomatal closure involves ALMT4, a phosphorylation-dependent vacuolar anion channel of Arabidopsis. Plant Cell 2017, 29, 2552–2569. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Andrews, J.; Mulholland, B.J.; McKee, J.M.T.; Hilton, H.W.; Horridge, J.S.; Farquhar, G.D.; Smeeton, R.C.; Smillie, I.R.A.; Black, C.R.; et al. Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol. 2007, 143, 1905–1917. [Google Scholar] [CrossRef]
- Negin, B.; Moshelion, M. The evolution of the role of ABA in the regulation of water-use efficiency: From biochemical mechanisms to stomatal conductance. Plant Sci. 2016, 251, 82–89. [Google Scholar] [CrossRef]
- Helander, J.D.M.; Vaidya, A.S.; Cutler, S.R. Chemical manipulation of plant water use. Bioorg. Med. Chem. 2016, 24, 493–500. [Google Scholar] [CrossRef]
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef]
- De Silva, D.L.R.; Cox, R.C.; Hetherington, A.M.; Mansfield, T.A. The role of abscisic acid and calcium in determining the behaviour of adaxial and abaxial stomata. New Phytol. 1986, 104, 41–51. [Google Scholar] [CrossRef]
- Tardieu, F.; Lafarge, T.; Simonneau, T. Stomatal control by fed or endogenous xylem ABA in sunflower: Interpretation of correlations between leaf water potential and stomatal conductance in anisohydric species. Plant Cell Environ. 1996, 19, 75–84. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.M. Passive origins of stomatal control. Science 2011, 582–585. [Google Scholar] [CrossRef] [PubMed]
- McAdam, S.A.M.; Brodribb, T.J. Fern and lycophyte guard cells do not respond to endogenous abscisic acid. Plant Cell 2012, 24, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.A.; Visel, D.; Kane, C.N.; Batz, T.A.; García, C.; Kaack, L.; Lamarque, L.J.; Wagner, Y.; King, A.; Torres-Ruiz, J.M.; et al. Drought-induced lacuna formation in the stem causes hydraulic conductance to decline before xylem embolism in Selaginella. New Phytol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Deans, R.M.; Brodribb, T.J.; Mcadam, S.A.M. An integrated hydraulic-hormonal model of conifer stomata predicts water stress dynamics. Plant Physiol. 2017, 174, 478–486. [Google Scholar] [CrossRef][Green Version]
- Cardoso, A.A.; Randall, J.M.; McAdam, S.A.M. Hydraulics regulate stomatal responses to changes in leaf water status in the fern Athyrium filix-femina. Plant Physiol. 2019, 179, 533–543. [Google Scholar] [CrossRef]
- Pressel, S.; Renzaglia, K.S.; Clymo, R.S.D.; Duckett, J.G. Hornwort stomata do not respond actively to exogenous and environmental cues. Ann. Bot. 2018, 122, 45–57. [Google Scholar] [CrossRef]
- Chater, C.; Gray, J.E.; Beerling, D.J. Early evolutionary acquisition of stomatal control and development gene signalling networks. Curr. Opin. Plant Biol. 2013, 16, 638–646. [Google Scholar] [CrossRef]
- Chater, C.; Kamisugi, Y.; Movahedi, M.; Fleming, A.; Cuming, A.C.; Gray, J.E.; Beerling, D.J. Regulatory mechanism controlling stomatal behavior conserved across 400 million years of land plant evolution. Curr. Biol. 2011, 21, 1025–1029. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Brodribb, T.J. The evolution of mechanisms driving the stomatal response to vapor pressure deficit. Plant Physiol. 2015, 167, 833–843. [Google Scholar] [CrossRef]
- Cai, S.; Chen, G.; Wang, Y.; Huang, Y.; Marchant, D.B.; Wang, Y.; Yang, Q.; Dai, F.; Hills, A.; Franks, P.J.; et al. Evolutionary conservation of ABA signaling for stomatal closure. Plant Physiol. 2017, 174, 732–747. [Google Scholar] [CrossRef]
- Hõrak, H.; Kollist, H.; Merilo, E. Fern stomatal responses to ABA and CO2 depend on species and growth conditions. Plant Physiol. 2017, 174, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.A.; McAdam, S.A.M. Misleading conclusions from exogenous ABA application: A cautionary tale about the evolution of stomatal responses to changes in leaf water status. Plant Signal. Behav. 2019, 14, 1610307. [Google Scholar] [CrossRef] [PubMed]
- McAdam, S.A.M.; Sussmilch, F.C. The evolving role of abscisic acid in cell function and plant development over geological time. Semin. Cell Dev. Biol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Windt, C.W.; Ponomarenko, A.; Zhang, Y.-J.; Gersony, J.; Rockwell, F.E.; Holbrook, N.M. Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiol. 2017, 174, 764–775. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Brodribb, T.J.; Lucani, C.J.; DaMatta, F.M.; McAdam, S.A.M. Coordinated plasticity maintains hydraulic safety in sunflower leaves. Plant Cell Environ. 2018, 41, 2567–2576. [Google Scholar] [CrossRef] [PubMed]
- Creek, D.; Lamarque, L.J.; Torres-Ruiz, J.M.; Parise, C.; Burlett, R.; Tissue, D.T.; Delzon, S. Xylem embolism in leaves does not occur with open stomata: Evidence from direct observations using the optical visualisation technique. J. Exp. Bot. 2019. [Google Scholar] [CrossRef]
- Davis, S.D.; Ewers, F.W.; Sperry, J.S.; Portwood, K.A.; Crocker, M.C.; Adams, G.C. Shoot dieback during prolonged drought in Ceanothus (Rhamnaceae) chaparral of California: A possible case of hydraulic failure. Am. J. Bot. 2002, 89, 820–828. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Batz, T.A.; McAdam, S.A.M. Xylem embolism resistance determines leaf mortality during drought in Persea americana. Plant Physiol. 2020, 182, 547–554. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Cochard, H. Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol. 2009, 149, 575–584. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Hammond, W.M.; Yu, K.; Wilson, L.A.; Will, R.E.; Anderegg, W.R.L.; Adams, H.D. Dead or dying? Quantifying the point of no return from hydraulic failure in drought-induced tree mortality. New Phytol. 2019, 223, 1834–1843. [Google Scholar] [CrossRef] [PubMed]
- Lamarque, L.; Delzon, S.; Toups, H.; Gravel, A.-I.; Corso, D.; Badel, E.; Burllet, R.; Charrier, G.; Cochard, H.; Jansen, S.; et al. Over-accumulation of abscisic acid in transgenic tomato plants increases the risk of hydraulic failure. Plant Cell Environ. 2020, 43, 548–562. [Google Scholar] [CrossRef]
- Skelton, R.P.; Brodribb, T.J.; Choat, B. Casting light on xylem vulnerability in an herbaceous species reveals a lack of segmentation. New Phytol. 2017, 214, 561–569. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Ogata, Y.; Morishita, Y.; Takeda, M.; Sakurai, N.; Suzuki, H.; Saito, K.; Shibata, D.; Kobayashi, M.; et al. Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 57, 1065–1078. [Google Scholar] [CrossRef]
- Kempa, S.; Krasensky, J.; Dal Santo, S.; Kopka, J.; Jonak, C. A central role of abscisic acid in stress-regulated carbohydrate metabolism. PLoS ONE 2008, 3, e3935. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef]
- Trouverie, J.; Thévenot, C.; Rocher, J.P.; Sotta, B.; Prioul, J.L. The role of abscisic acid in the response of a specific vacuolar invertase to water stress in the adult maize leaf. J. Exp. Bot. 2003, 54, 2177–2186. [Google Scholar] [CrossRef]
- Thalmann, M.; Pazmino, D.; Seung, D.; Horrer, D.; Nigro, A.; Meier, T.; Kölling, K.; Pfeifhofer, H.W.; Zeeman, S.C.; Santelia, D. Regulation of leaf starch degradation by abscisic acid is important for osmotic stress tolerance in plants. Plant Cell 2016, 28, 1860–1878. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Brunetti, C.; Savi, T.; Nardini, A.; Loreto, F.; Gori, A.; Centritto, M. Changes in abscisic acid content during and after drought are related to carbohydrate mobilization and hydraulic recovery in poplar stems. Tree Physiol. 2020, 40, 1043–1057. [Google Scholar] [CrossRef]
- Blum, A.; Sullivan, C.Y.; Nguyen, H. The effect of plant size on wheat response to agents of drought stress. II. Water deficit, heat and ABA. Funct. Plant Biol. 1997, 24, 43–48. [Google Scholar] [CrossRef]
- Slewinski, T.L. Non-structural carbohydrate partitioning in grass stems: A target to increase yield stability, stress tolerance, and biofuel production. J. Exp. Bot. 2012, 63, 4647–4670. [Google Scholar] [CrossRef] [PubMed]
- Savoure, A.; Jaoua, S.; Hua, X.J.; Ardiles, W.; Vanmontagu, M.; Verbruggen, N. Isolation, characterization and chromosomal location of a gene encoding the Δ1-pyrroline-5-carboxylate synthetase in Arabidopsis thaliana. FEBS Lett. 1995, 372, 13–19. [Google Scholar] [CrossRef]
- Strizhov, N.; Abrahám, E.; Okrész, L.; Blickling, S.; Zilberstein, A.; Schell, J.; Koncz, C.; Szabados, L. Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. Plant J. 1997, 12, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.; Rigo, G.; Szekely, G.; Nagy, R.; Koncz, C.; Szabados, L. Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol. Biol. 2003, 51, 363–372. [Google Scholar] [CrossRef]
- Sharma, S.; Verslues, P.E. Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant Cell Environ. 2011, 33, 1838–1851. [Google Scholar] [CrossRef]
- Bhaskara, G.B.; Yang, T.H.; Verslues, P.E. Dynamic proline metabolism: Importance and regulation in water limited environments. Front. Plant Sci. 2015, 6, 484. [Google Scholar] [CrossRef]
- Bandurska, H.; Niedziela, J.; Pietrowska-Borek, M.; Nuc, K.; Chadzinikolau, T.; Radzikowska, D. Regulation of proline biosynthesis and resistance to drought stress in two barley (Hordeum vulgare L.) genotypes of different origin. Plant Physiol. Biochem. 2017, 118, 427–437. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: Multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Gai, Z.; Wang, Y.; Ding, Y.; Qian, W.; Qiu, C.; Xie, H.; Sun, L.; Jiang, Z.; Ma, Q.; Wang, L.; et al. Exogenous abscisic acid induces the lipid and flavonoid metabolism of tea plants under drought stress. Sci. Rep. 2020, 10, 12275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Gao, G.; Liu, J.; Yang, G.; Lv, Z.; Zhang, J.; He, C. Transcripts and ABA dependent signaling in response to drought stress in Hippophae rhamnoides L. Trees 2020, 34, 1033–1045. [Google Scholar] [CrossRef]
- Perin, E.C.; da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-dependent salt and drought stress improve strawberry fruit quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Villalobos-González, L.; Peña-Neira, A.; Ibáñez, F.; Pastenes, C. Long-term effects of abscisic acid (ABA) on the grape berry phenylpropanoid pathway: Gene expression and metabolite content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Li, D.; Luo, Z.; Mao, L.; Ying, T. Transcriptomic analysis reveals possible influences of ABA on secondary metabolism of pigments, flavonoids and antioxidants in tomato fruit during ripening. PLoS ONE 2015, 10, e0129598. [Google Scholar] [CrossRef]
- Brunetti, C.; Fini, A.; Sebastiani, F.; Gori, A.; Tattini, M. Modulation of phytohormone signaling: A primary function of flavonoids in plant–environment interactions. Front. Plant Sci. 2018, 9, 1042. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Sunderland, T.C.; Rowland, D. Forests, land use, and challenges to climate stability and food security. In Sustainable Food and Agriculture; Campanhola, C., Pandey, S., Eds.; The Food and Agriculture Organization of the United Nations and Academic Press: Cambridge, MA, USA, 2019; pp. 95–116. [Google Scholar]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulías, J.; Pou, A.; Ribas-Carbo, M.; Tomàs, M.; Medrano, H. Improving water use efficiency in grapevines: Potential physiological targets for biotechnological improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Bunce, J.A. Variation among soybean cultivars in mesophyll conductance and leaf water use efficiency. Plants 2016, 5, 44. [Google Scholar] [CrossRef]
- Li, C.; Jackson, P.; Lu, X.; Xu, C.; Cai, Q.; Basnayake, J.; Lakshmanan, P.; Ghannoum, O.; Fan, Y. Genotypic variation in transpiration efficiency due to differences in photosynthetic capacity among sugarcane-related clones. J. Exp. Bot. 2017, 68, 2377–2385. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, Z.; Liu, J.; Tischer, S.V.; Christmann, A.; Windisch, W.; Schnyder, H.; Grill, E. Leveraging abscisic acid receptors for efficient water use in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6791–6796. [Google Scholar] [CrossRef] [PubMed]
- Nadal, M.; Flexas, J. Variation in photosynthetic characteristics with growth form in a water-limited scenario: Implications for assimilation rates and water use efficiency in crops. Agric. Water Manag. 2019, 216, 457–472. [Google Scholar] [CrossRef]
- Tardieu, F.; Zhang, J.; Katerji, N.; Bethenod, O.; Palmer, S.; Davies, W.J. Xylem ABA controls the stomatal conductance of field-grown maize subjected to soil compaction or soil drying. Plant Cell Environ. 1992, 15, 193–197. [Google Scholar] [CrossRef]
- Saradadevi, R.; Bramley, H.; Siddique, K.H.; Edwards, E.; Palta, J.A. Contrasting stomatal regulation and leaf ABA concentrations in wheat genotypes when split root systems were exposed to terminal drought. Field Crop. Res. 2014, 165, 5–14. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Todaka, D.; Takahashi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. ABA-responsive gene expression in response to drought stress: Cellular regulation and long-distance signaling. Adv. Bot. Res. 2019, 92, 83–113. [Google Scholar] [CrossRef]
- Chater, C.C.C.; Oliver, J.; Casson, S.; Gray, J.E. Putting the brakes on: Abscisic acid as a central environmental regulator of stomatal development. New Phytol. 2014, 202, 376–391. [Google Scholar] [CrossRef]
- Arend, M.; Schnitzler, J.-P.; Ehlting, B.; Hänsch, R.; Lange, T.; Rennenberg, H.; Himmelbach, A.; Grill, E.; Fromm, J. Expression of the Arabidopsis mutant ABI1 gene alters abscisic acid sensitivity: Stomatal development, and growth morphology in gray poplars. Plant Physiol. 2009, 151, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [PubMed]
- Prado, K.; Maurel, C. Regulation of leaf hydraulics: From molecular to whole plant levels. Front. Plant Sci. 2013, 4, 255. [Google Scholar] [CrossRef] [PubMed]
- Shatil-Cohen, A.; Attia, Z.; Moshelion, M. Bundle-sheath cell regulation of xylem-mesophyll water transport via aquaporins under drought stress: A target of xylem-borne ABA? Plant J. 2011, 67, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Dejonghe, W.; Cutler, S.R. Abscisic acid as a gateway for the crops of tomorrow. Adv. Bot. Res. 2019, 92, 341–370. [Google Scholar]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Park, S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.-F.F.; et al. Abscisic acid inhibits Type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, J.; Poree, F.; Schaeufele, R.; Helmke, H.; Frackenpohl, J.; Lehr, S.; von Koskull-Döring, P.; Christmann, A.; Schnyder, H.; et al. Abscisic acid receptors and coreceptors modulate plant water use efficiency and water productivity. Plant Physiol. 2019, 180, 1066–1080. [Google Scholar] [CrossRef]
- Mega, R.; Tsujimoto, H.; Okamoto, M. Genetic manipulation of abscisic acid receptors enables modulation of water use efficiency. Plant Signal. Behav. 2019, 14, e1642039. [Google Scholar] [CrossRef]
- Oneto, C.D.; Otegui, M.E.; Baroli, I.; Beznec, A.; Faccio, P.; Bossio, E.; Blumwald, E.; Lewi, D. Water deficit stress tolerance in maize conferred by expression of an isopentenyltransferase (IPT) gene driven by a stress- and maturation-induced promoter. J. Biotechnol. 2016, 220, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M.; Kidokoro, S.; Yoshida, T.; Mizoi, J.; Kojima, M.; Takebayashi, T.; Sakakibara, H.; Fernie, A.R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A gene-stacking approach to overcome the trade-off between drought stress tolerance and growth in Arabidopsis. Plant J. 2019, 97, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Peterson, F.C.; Defries, A.; Park, S.Y.; Endo, A.; Nambara, E.; Volkman, B.F.; Cutler, S.R. Activation of dimeric ABA receptors elicits guard cell closure, ABA-regulated gene expression, and drought tolerance. Proc. Natl. Acad. Sci. USA 2013, 110, 12132–12137. [Google Scholar] [CrossRef]
- Park, S.Y.; Peterson, F.C.; Mosquna, A.; Yao, J.; Volkman, B.F.; Cutler, S.R. Agrochemical control of plant water use using engineered abscisic acid receptors. Nature 2015, 520, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Liu, X.; Zhang, Y.; Xue, X.; Zhou, X.E.; Melcher, K.; Gao, P.; Wang, F.; Zeng, L.; Zhao, Y.; et al. An ABA-mimicking ligand that reduces water loss and promotes drought resistance in plants. Cell Res. 2013, 23, 1043–1054. [Google Scholar] [CrossRef]
- Vaidya, A.S.; Helander, J.D.; Peterson, F.C.; Elzinga, D.; Dejonghe, W.; Kaundal, A.; Park, S.Y.; Xing, Z.; Mega, R.; Takeuchi, J.; et al. Dynamic control of plant water use using designed ABA receptor agonists. Science 2019, 366, eaaw8848. [Google Scholar] [CrossRef]
- Zhou, Y.; He, R.; Guo, Y.; Liu, K.; Huang, G.; Peng, C.; Liu, Y.; Zhang, M.; Li, Z.; Duan, L. A novel ABA functional analogue B2 enhances drought tolerance in wheat. Sci. Rep. 2019, 9, 2887. [Google Scholar] [CrossRef]
- Chang, Y.; Nguyen, B.H.; Xie, Y.; Xiao, B.; Tang, N.; Zhu, W.; Mou, T.; Xiong, L. Co-overexpression of the constitutively active form of OsbZIP46 and ABA-activated protein kinase SAPK6 improves drought and temperature stress resistance in rice. Front. Plant Sci. 2017, 8, 1102. [Google Scholar] [CrossRef]
- Verma, R.K.; Santosh Kumar, V.V.; Yadav, S.K.; Pushkar, S.; Rao, M.V.; Chinnusamy, V. Overexpression of ABA Receptor PYL10 gene confers drought and cold tolerance to indica rice. Front. Plant Sci. 2019, 10, 1488. [Google Scholar] [CrossRef]
- Kim, D.H. Mechanism of ABA signal transduction: Agricultural highlights for improving drought tolerance. J. Plant Biol. 2014, 57, 1–8. [Google Scholar] [CrossRef]
- Yu, J.; Ge, H.; Wang, X.; Tang, R.; Wang, Y.; Zhao, F.; Lan, W.; Luan, S.; Yang, L. Overexpression of pyrabactin resistance-like abscisic acid receptors enhances drought, osmotic, and cold tolerance in transgenic poplars. Front. Plant Sci. 2017, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Yin, P.; Yan, C.; Yuan, X.; Li, W.; Zhang, Z.; Liu, L.; Wang, J.; Yan, N. Functional mechanism of the abscisic acid agonist pyrabactin. J. Biol. Chem. 2010, 285, 28946–28952. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.L.; Lozano-Juste, J. Unnatural agrochemical ligands for engineered abscisic acid receptors. Trends Plant Sci. 2015, 20, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.S.; Dai, L.; Ahmad, F.; Ahmad, A.; Li, J.; Zhang, C. AM1 is a potential ABA substitute for drought tolerance as revealed by physiological and ultra-structural responses of oilseed rape. Acta Physiol. Plant. 2016, 38, 183. [Google Scholar] [CrossRef]
- Xiong, J.L.; Dai, L.L.; Ma, N.; Zhang, C.L. Transcriptome and physiological analyses reveal that AM1 as an ABA-mimicking ligand improves drought resistance in Brassica napus. Plant Growth Regul. 2018, 85, 73–90. [Google Scholar] [CrossRef]
- González-Guzmán, M.; Rodríguez, L.; Lorenzo-Orts, L.; Pons, C.; Sarrión-Perdigones, A.; Fernández, M.A.; Peirats-Llobet, M.; Forment, J.; Moreno-Alvero, M.; Cutler, S.R.; et al. Tomato PYR/PYL/RCAR abscisic acid receptors show high expression in root, differential sensitivity to the abscisic acid agonist quinabactin, and the capability to enhance plant drought resistance. J. Exp. Bot. 2014, 65, 4451–4464. [Google Scholar] [CrossRef]
- Cao, M.J.; Zhang, Y.L.; Liu, X.; Huang, H.; Zhou, X.E.; Wang, W.L.; Zeng, A.; Zhao, C.-Z.; Si, T.; Du, J.; et al. Combining chemical and genetic approaches to increase drought resistance in plants. Nat. Commun. 2017, 8, 1183. [Google Scholar] [CrossRef]
- Blum, A. Towards a conceptual ABA ideotype in plant breeding for water limited environments. Funct. Plant Biol. 2015, 42, 502–513. [Google Scholar] [CrossRef]
- Nuccio, M.L.; Paul, M.; Bate, N.J.; Cohn, J.; Cutler, S.R. Where are the drought tolerant crops? An assessment of more than two decades of plant biotechnology effort in crop improvement. Plant Sci. 2018, 273, 110–119. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Field, T.S. Stem hydraulic supply is linked to leaf photosynthetic capacity: Evidence from New Caledonian and Tasmanian rainforests. Plant Cell Environ. 2000, 23, 1381–1388. [Google Scholar] [CrossRef]
- Ahmad, H.B.; Lens, F.; Capdeville, G.; Burlett, R.; Lamarque, L.J.; Delzon, S. Intraspecific variation in embolism resistance and stem anatomy across four sunflowers. Physiol. Plant. 2017, 163, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, H.S.; Grantz, D.A.; Meinzer, F.C.; Goldstein, G.; Crisosto, G.M.; Crisosto, C. Genotypic variability in vulnerability of leaf xylem to cavitation in water-stressed and well-irrigated sugarcane. Plant Physiol. 1992, 100, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).