Steroid Metabolites Support Evidence of Autism as a Spectrum

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Methods
2.4. Statistical Analysis
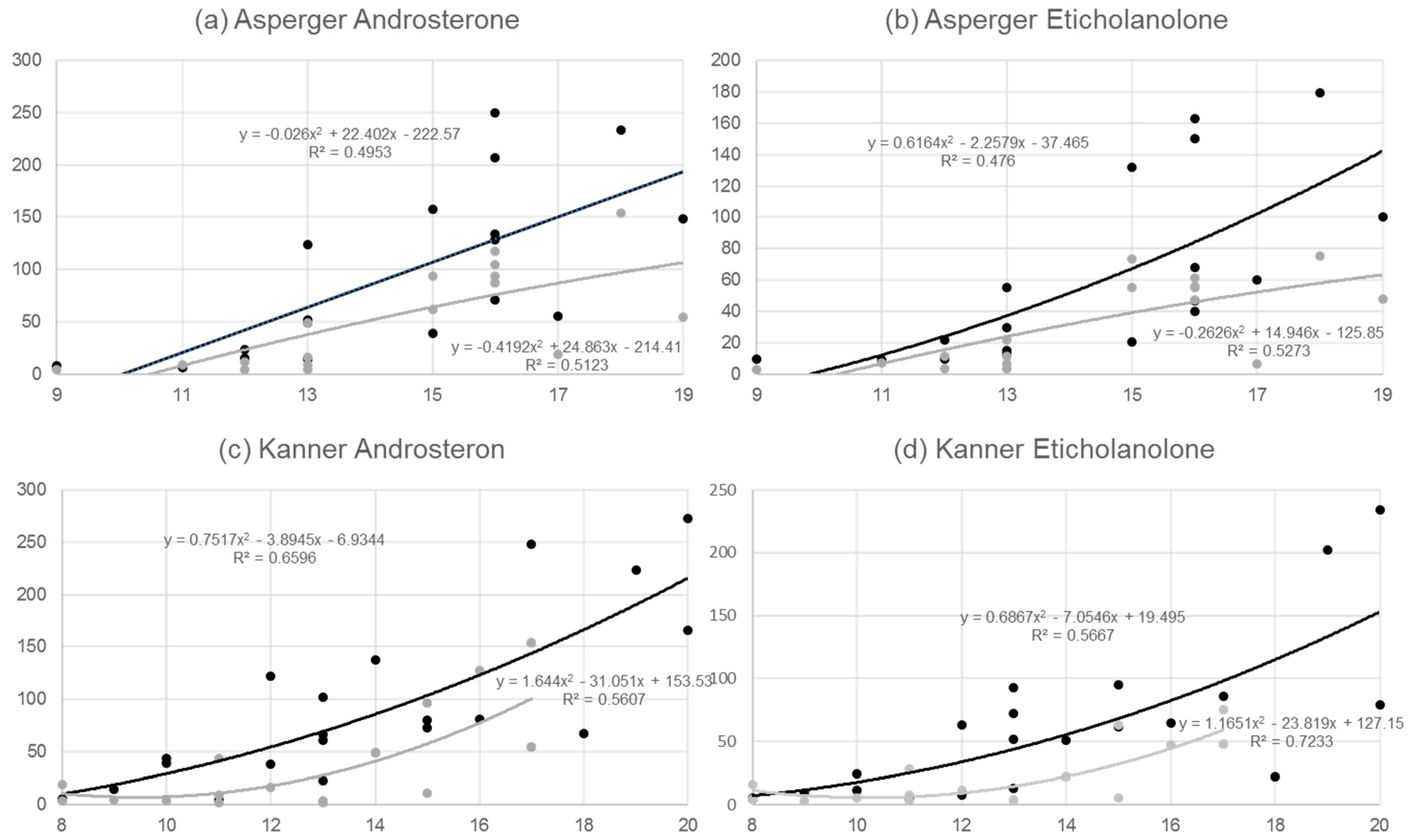
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gillberg, C.; Fernell, E.; Kočovská, E.; Minnis, H.; Bourgeron, T.; Thompson, L.; Allely, C. The role of cholesterol metabolism and various steroid abnormalities in autism spectrum disorders: A hypothesis paper. Autism Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Akshoomoff, N.; Pierce, K.; Courchesne, E. The neurobiological basis of autism from a developmental perspective. Dev. Psychopathol. 2002, 14, 613–634. [Google Scholar] [CrossRef]
- Dziobek, I.; Gold, S.M.; Wolf, O.T.; Convit, A. Hypercholesterolemia in Asperger syndrome: Independence from lifestyle, obsessive-compulsive behavior, and social anxiety. Psychiatry Res. 2007, 149, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Fangmeier, T. Patogenetische Modelle. In Das Asperger-Syndrom im Erwachsenenalter; Van Elst, L.T., Ed.; Medizinisch Wissenschaftliche Verlagsgesellschaft: Berlin, Germany, 2013. [Google Scholar]
- Lamb, J.A.; Moore, J.; Bailey, A.; Monaco, A.P. Autism: Recent molecular genetic advances. Hum. Mol. Genet. 2000, 9, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T. Genetic factors in Asperger syndrome. Nihon Rinsho 2007, 65, 443–448. [Google Scholar]
- Schwarz, E.; Guest, P.C.; Rahmoune, H.; Wang, L.; Levin, Y.; Ingudomnukul, E.; Ruta, L.; Kent, L.; Spain, M.; Baron-Cohen, S.; et al. Sex-specific serum biomarker patterns in adults with Asperger’s syndrome. Mol. Psychiatry 2011, 16, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Volkmar, F.R.; Lord, C.; Bailey, A.; Schultz, R.T.; Klin, A. Autism and pervasive developmental disorders. J. Child Psychol. Psychiatry 2004, 45, 135–170. [Google Scholar] [CrossRef] [PubMed]
- Volkmar, F.R.; Klin, A.; Pauls, D. Nosological and genetic aspects of Asperger syndrome. J. Autism Dev. Disord. 1998, 28, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.M.; Gutknecht, L.; Cohen, D.J.; Brailly-Tabard, S.; Cohen, J.H.; Ferrari, P.; Roubertoux, P.L.; Tordjman, S. Serotonin transporter promotor variants in autism: Functional effects and relationship to platelet hyperserotonemia. Mol. Psychiatry 2002, 7, 831–836. [Google Scholar] [CrossRef]
- Auranen, M.; Vanhala, R.; Varilo, T.; Ayers, K.; Kempas, E.; Ylisaukko-Oja, T.; Sinsheimer, J.S.; Peltonen, L.; Järvelä, I. A genomewide screen for autism-spectrum disorders: Evidence for a major susceptibility locus on chromosome 3q25-27. Am. J. Hum. Genet. 2002, 71, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.W.; Tierney, E. Hypothesis: The role of sterols in autism spectrum disorder. Autism Res. Treat. 2011, 653570. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Matsuzaki, H.; Miyachi, T.; Shimmura, C.; Suda, S.; Tsuchiya, K.J.; Matsumoto, K.; Suzuki, K.; Iwata, Y.; Nakamura, K.; et al. Investigation of the serum levels of anterior pituitary hormones in male children with autism. Mol. Autism 2011, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Ariella Ritvo, R.; Ritvo, E.R.; Guthrie, D.; Ritvo, M.J. Clinical evidence that Asperger’s disorder is a mild form of autism. Compr. Psychiatry 2008, 49, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hu, V.W.; Nguyen, A.; Kim, K.S.; Steinberg, M.E.; Sarachana, T.; Scully, M.A.; Soldin, S.J.; Luu, T.; Lee, N.H. Gene expression profiling of lymphoblasts from autistic and nonaffected sib pairs: Altered pathways in neuronal development and steroid biosynthesis. PLoS ONE 2009, 4, e5775. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; Corbett, B.A. A review of rhythm and responsiveness of cortisol in individuals with autism spectrum disorders. Psychoneuroendocrinology 2014, 49, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Bagnoli, F.; Mori, A.; Fommei, C.; Coriolani, G.; Badii, S.; Tomasini, B. ACTH and cortisol cord plasma concentrations in preterm and term infants. J. Perinatol. 2013, 33, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.T.; Hewedi, D.H.; Ismail, M.A. Basal and adrenocorticotropic hormone stimulated plasma cortisol levels among Egyptian autistic children: Relation to disease severity. Ital. J. Pediatr. 2010, 36, 71. [Google Scholar] [CrossRef]
- Brosnan, M.; Turner-Cobb, J.; Munro-Naan, Z.; Jessop, D. Absence of a normal cortisol awakening response (CAR) in adolescent males with Asperger syndrome (AS). Psychoneuroendocrinology 2009, 34, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Marinović-Curin, J.; Marinović-Terzić, I.; Bujas-Petković, Z.; Zekan, L.; Skrabić, V.; Dogas, Z.; Terzić, J. Slower cortisol response during ACTH stimulation test in autistic children. Eur. Child Adolesc. Psychiatry 2008, 17, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Tani, P.; Lindberg, N.; Matto, V.; Appelberg, B.; Nieminen-von Wendt, T.; von Wendt, L.; Porkka-Heiskanen, T. Higher plasma ACTH levels in adults with Asperger syndrome. Psychosom. Res. 2005, 58, 533–536. [Google Scholar] [CrossRef]
- Ishimoto, H.; Ginzinger, D.G.; Jaffe, R.B. Adrenocorticotropin preferentially up-regulates angiopoietin 2 in the human fetal adrenal gland: Implications for coordinated adrenal organ growth and angiogenesis. J. Clin. Endocrinol. Metab. 2006, 91, 1909–1915. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, H.; Minegishi, K.; Higuchi, T.; Furuya, M.; Asai, S.; Kim, S.H.; Tanaka, M.; Yoshimura, Y.; Jaffe, R.B. The periphery of the human fetal adrenal gland is a site of angiogenesis: Zonal differential expression and regulation of angiogenic factors. J. Clin. Endocrinol. Metab. 2008, 93, 2402–2408. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hitoshi, I.; Robert, B. Development and Function of the Human Fetal Adrenal Cortex: A Key Component in the Feto-Placental Unit. Endocr. Rev. 2011, 32, 317–355. [Google Scholar]
- Ruta, L.; Ingudomnukul, E.; Taylor, K.; Chakrabarti, B.; Baron-Cohen, S. Increased serum androstenedione in adults with autism spectrum conditions. Psychoneuroendocrinology 2011, 36, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Marotta, R.; Di Cello, A.; Russo, T.; Falbo, A.; Orio, F.; Tolino, A.; Zullo, F.; Esposito, R.; La Sala, G.B. Pervasive developmental disorders in children of hyperandrogenic women with polycystic ovary syndrome: A longitudinal case-control study. Clin. Endocrinol. 2012, 77, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Engberg, H.; Butwicka, A.; Nordenström, A.; Hirschberg, A.L.; Falhammar, H.; Lichtenstein, P.; Nordenskjöld, A.; Frisén, L.; Landén, M. Congenital adrenal hyperplasia and risk for psychiatric disorders in girls and women born between 1915 and 2010: A total population study. Psychoneuroendocrinology 2015, 60, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Falhammar, H.; Butwicka, A.; Landén, M.; Lichtenstein, P.; Nordenskjöld, A.; Nordenström, A.; Frisén, L. Increased psychiatric morbidity in men with congenital adrenal hyperplasia due to 21-hydroxylase deficiency. J. Clin. Endocrinol. Metab. 2013, 99, E554–E560. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Zhang, H.F.; Shou, X.J.; Li, J.; Jing, W.L.; Zhou, Y.; Qian, Y.; Han, S.P.; Zhang, R.; Han, J.S. Prenatal hyperandrogenic environment induced autistic-like behavior in rat offspring. Physiol. Behav. 2015, 138, 13–20. [Google Scholar] [CrossRef]
- Bradstreet, J.J.; Smith, S.; Granpeesheh, D.; El-Dahr, J.M.; Rossignol, D. Spironolactone might be a desirable immunologic and hormonal intervention in autism spectrum disorders. Med. Hypotheses 2006, 68, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Escobar, J.C.; Patel, S.S.; Beshay, V.E.; Suzuki, T.; Carr, B.R. The human placenta expresses CYP17 and generates androgens de novo. J. Clin. Endocrinol. Metab. 2011, 96, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.; Schmidt, R.J.; Hertz-Picciotto, I. Maternal lifestyle and environmental risk factors for autism spectrum disorders. Int. J. Epidemiol. 2014, 43, 443–464. [Google Scholar] [CrossRef]
- Majewska, M.R.; Hill, M.; Urbanowicz, E.; Rok-Bujko, R.; Bien’kowski, P.; Namysłowska, I.; Mierzejewski, P. Marked elevation of adrenal steroids, especially androgens, in saliva of prepubertal autistic children. Eur. Child Adolesc. Psychiatry 2014, 23, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Baron-Cohen, S.; Auyeung, B.; Nørgaard-Pedersen, B.; Hougaard, D.M.; Abdallah, M.W.; Melgaard, L.; Cohen, A.S.; Chakrabarti, B.; Ruta, L.; Lombardo, M.V. Elevated fetal steroidogenic activity in autism. Mol. Psychiatry 2015, 20, 369–376. [Google Scholar] [CrossRef]
- Saenz, J.; Alexander, G.M. Postnatal testosterone levels and disorder relevant behavior in the second year of life. Biol. Psychol. 2013, 94, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.V.; Ashwin, E.; Auyeung, B.; Chakrabarti, B.; Taylor, K.; Hackett, G.; Bullmore, E.T.; Baron-Cohen, S. Fetal testosterone influences sexually dimorphic gray matter in the human brain. J. Neurosci. 2012, 32, 674–680. [Google Scholar] [CrossRef]
- Popper, K.R. Logik der Forschung; Mohr Siebeck: Tübingen, Germany, 1969. [Google Scholar]
- Bennett, T.; Szatmari, P.; Bryson, S.; Volden, J.; Zwaigenbaum, L.; Vaccarella, L.; Duku, E.; Boyle, M. Differentiating autism and Asperger syndrome on the basis of language delay or impairment. J. Autism Dev. Disord. 2008, 38, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Shackleton, C.H.L. Profiling steroid hormones and urinary steroids. J. Chromatogr. 1986, 379, 91–156. [Google Scholar] [CrossRef]
- Shackleton, C.H.L. Role of a Disordered Steroid Metabolome in the Elucidation of Sterol and Steroid Biosynthesis. Lipids 2012, 47, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vogt, B.; Dick, B.; N’Gankam, V.; Frey, F.J.; Frey, B.M. Reduced 11B-hydroxysteroid dehydrogenase activity in patients with the nephrotic syndrome. J. Clin. Endocrinol. Metab. 1999, 84, 811–814. [Google Scholar]
- Asperger, H. Die “Autistischen Psychopathen” im Kindesalter. Archiv für Psychiatrie und Nervenkrankheiten 1944, 117, 76–136. [Google Scholar] [CrossRef]
- Kanner, L. Autistic disturbances of affective contact. Nerv. Child 1943, 2, 217–250. [Google Scholar]
- Gorinaa, A.S.; Kolesnichenkob, L.S.; Mikhnovichc, V.I. Catecholamine Metabolism in Children with Asperger’s and Kanner’s Syndromes. Biomed. Chem. 2011, 5, 397–401. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr. Rev. 2011, 32, 579. [Google Scholar] [CrossRef] [PubMed]
- Ueshiba, H.; Takeda, S.; Segawa, M.; Ueshiba, H.; Miyachi, Y. Serum androsterone levels in children. Horm. Metab. Res. 1996, 28, 190–192. [Google Scholar] [CrossRef] [PubMed]
- Margolese, M.S.; Janiger, O. Androsterone/Etiocholanolone Ratios in Male Homosexuals. Br. Med. J. 1973, 3, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Bocian-Sobkowska, J. Morphometric study of the human suprarenal gland in the first postnatal year. Folia Morphol. (Warsz) 2000, 58, 275–284. [Google Scholar] [PubMed]
- Sarkar, P.; Bergman, K.; O’Connor, T.G.; Glover, V. Maternal antenatal anxiety and amniotic fluid cortisol and testosterone: Possible implications for foetal programming. J. Neuroendocrinol. 2008, 20, 489–496. [Google Scholar] [CrossRef]
- Chakrabarti, B.; Dudbridge, F.; Kent, L.; Wheelwright, S.; Hill-Cawthorne, G.; Allison, C.; Banerjee-Basu, S.; Baron-Cohen, S. Genes related to sex steroids, neural growth, and social-emotional behavior are associated with autistic traits, empathy, and Asperger syndrome. Autism Res. 2009, 2, 157–177. [Google Scholar] [CrossRef]
- Mills, J.L.; Hediger, M.L.; Molloy, C.A.; Chrousos, G.P.; Manning-Courtney, P.; Yu, K.F.; Brasington, M.; England, L.J. Elevated levels of growth-related hormones in autism and autism spectrum disorder. Clin. Endocrinol. 2007, 67, 230–237. [Google Scholar] [CrossRef]


{kind=link}
| Asperger | p-Value | Kanner | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Boys | Controls | Boys | Controls | |||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |||
| Androgen Metabolites | ||||||||||
| Androsterone | 95.43 | 18.36 | 0.008 | 57.7 | 9.79 | 89.29 | 17.4 | 0.001 | 61.45 | 13.67 |
| Etiocholanolone | 62.45 | 13.21 | 0.043 | 38.92 | 6.16 | 59.78 | 13.45 | 0.009 | 29.23 | 6.24 |
| Androstenediol | 3.35 | 0.66 | 0.056 | 2.2 | 0.41 | 3.58 | 0.7 | 0.087 | 2.88 | 0.6 |
| 11-Oxoetiocholanolone | 26.53 | 3.48 | 0.667 | 24.31 | 2.86 | 30.93 | 6.96 | 0.119 | 19.45 | 2.05 |
| 11β-Hydroxyandrosterone | 32.84 | 3.57 | 0.07 | 26.2 | 3.15 | 46.78 | 7.57 | 0.033 | 30.72 | 4.41 |
| 11β-Hydroxyetiocholanolone | 18.76 | 3.15 | 0.913 | 19.3 | 2.78 | 23.1 | 4.57 | 0,08 | 14.5 | 1.91 |
| Dehydroepiandrosterone | 6.02 | 2.65 | 0.72 | 4.92 | 1.31 | 36.82 | 19.5 | 0.129 | 7.57 | 2.03 |
| 5-Androstene-3β,17β-diol | 3.67 | 0.57 | 0.538 | 3.19 | 0.55 | 7.13 | 2.67 | 0.191 | 3.85 | 0.73 |
| 16α-Hydroxydehydroepiandrosterone | 11.05 | 3.31 | 0.311 | 20.16 | 7.29 | 16.32 | 4.42 | 0.506 | 21.26 | 6.88 |
| 5-Androstene-3β,16α,17β-triol | 18.73 | 4.53 | 0.537 | 23.24 | 5.44 | 19.31 | 4.06 | 0.504 | 23.23 | 5.18 |
| 5-Pregnene-3β, 16α,17β-triol | 6.41 | 2.21 | 0.739 | 7.21 | 1.22 | 10.55 | 5.2 | 0.696 | 8.66 | 1.33 |
| Testosterone | 2.93 | 0.71 | 0.056 | 1.46 | 0.32 | 3.01 | 1 | 0.134 | 1.62 | 0.34 |
| 5α-Dihydrotestosterone | 1.29 | 0.17 | 0.051 | 0.96 | 0.12 | 1.66 | 0.24 | 0.035 | 1.16 | 0.15 |
| Oestrogen metabolites | ||||||||||
| Estriol | 0.14 | 0.02 | 0.859 | 0.14 | 0.03 | 0.18 | 0.04 | 0.288 | 0.16 | 0.02 |
| 17β-Estradiol | 0.1 | 0.02 | 0.043 | 0.05 | 0.01 | 0.09 | 0.02 | 0.505 | 0.08 | 0.01 |
| Progesterone metabolites | ||||||||||
| 17-Hydroxypregnanolone | 5.43 | 0.86 | 0.068 | 3.32 | 0.76 | 7.34 | 1.32 | 0.005 | 3.7 | 0.46 |
| Pregnanediol | 10.31 | 0.95 | 0.37 | 8.91 | 2.05 | 13.45 | 1.61 | 0.019 | 9.41 | 1.16 |
| Pregnanetriol | 32.62 | 4.95 | 0.043 | 22.17 | 5.1 | 35.15 | 5.09 | 0.001 | 16.48 | 1.98 |
| 11-Oxopregnanetriol | 1.41 | 0.29 | 0.482 | 1.1 | 0.25 | 2.01 | 0.57 | 0.108 | 0.99 | 0.16 |
| 11-Deoxycortisol Metabolite | ||||||||||
| Tetrahydrodeoxycortisol | 0.48 | 0.04 | 0.588 | 0.44 | 0.07 | 0.68 | 0.09 | 0.031 | 0.47 | 0.03 |
| Corticosterone metabolites | ||||||||||
| Tetrahydrodihydrocorticosterone | 12.83 | 1.42 | 0.093 | 9.38 | 1.26 | 14.72 | 1.86 | 0.014 | 9.64 | 0.63 |
| Tetrahydrocorticosterone | 11.77 | 1.22 | 0.193 | 9.57 | 1.14 | 15.6 | 1.57 | 0.005 | 10.22 | 0.85 |
| 5α-Tetrahydrocorticosterone | 26.8 | 3.73 | 0.096 | 19.67 | 2.13 | 47.71 | 8.2 | 0.015 | 24.66 | 2.2 |
| 18-Hydroxytetrahydrodihydrocorticosterone | 4.53 | 0.51 | 0.364 | 3.34 | 0.58 | 6.24 | 1 | 0.073 | 3.28 | 0.67 |
| Tetrahydroaldosterone | 2.38 | 0.33 | 0.002 | 1.24 | 0.1 | 1.96 | 0.23 | 0.096 | 1.48 | 0.15 |
| Cortisone Metabolites | ||||||||||
| Cortisone | 10.8 | 1.34 | 0.592 | 11.86 | 1.18 | 13.74 | 1.91 | 0.884 | 13.42 | 1.46 |
| Tetrahydrocortisone | 245.62 | 21.04 | 0.005 | 171.72 | 12.04 | 278.77 | 27.31 | 0.007 | 185.14 | 16.29 |
| α-Cortolone | 84.51 | 6.91 | 0.002 | 59.54 | 3.65 | 99.76 | 16.05 | 0.031 | 63.1 | 4.78 |
| β-Cortolone | 0.69 | 0.07 | 0.001 | 110.39 | 18.11 | 0.73 | 0.1 | 0 | 137.5 | 25.01 |
| 20α-Dihydrocortisone | 1.3 | 0.19 | 0.989 | 1.29 | 0.13 | 1.64 | 0.28 | 0.784 | 1.55 | 0.19 |
| 20β-Dihydrocortisone | 3.79 | 0.49 | 0.849 | 3.92 | 0.39 | 4.88 | 0.79 | 0.932 | 4.96 | 0.61 |
| Cortisol metabolites | ||||||||||
| Cortisol | 8.72 | 1.4 | 0.209 | 6.33 | 0.65 | 12.72 | 2.22 | 0.044 | 7.36 | 1.02 |
| Tetrahydrocortisol | 76.84 | 6.13 | 0.016 | 56 | 4.02 | 106.23 | 9.53 | 0.001 | 63.06 | 6.38 |
| 5α-Tetrahydrocortisol | 102.92 | 14.16 | 0.017 | 60.91 | 4.07 | 146 | 20.78 | 0.005 | 79.77 | 6.97 |
| α-Cortol | 16.02 | 1.23 | 0.002 | 10.82 | 0.69 | 22.09 | 2.31 | 0.001 | 12.34 | 0.9 |
| β-Cortol | 21.11 | 1.61 | 0.125 | 16.93 | 2.11 | 29.17 | 2.97 | 0.009 | 17.18 | 2.35 |
| 20α-Dihydrocortisol | 3.28 | 0.31 | 0.556 | 3.71 | 0.55 | 5.91 | 1.15 | 0.297 | 4.55 | 0.7 |
| 6β-Hydroxycortisol | 9.68 | 0.85 | 0.754 | 9.13 | 1.39 | 19.58 | 6.07 | 0.095 | 9.51 | 1.47 |
| 18-Hydroxycortisol | 26.34 | 1.95 | 0.54 | 23.3 | 124 | 53.56 | 15.9 | 0.245 | 32.7 | 6.02 |
| Asperger Boys | p< | Control Boys | Kanner Boys | p< | Control Boys | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| 21-Hydroxylase Deficit | ||||||||||
| 17HP/(THE + THF + 5aTHF) | 0.014 | 0.01 | 0.684 | 0.013 | 0.01 | 0.004 | 0.021 | 0.711 | 0.014 | 0.02 |
| PT/(THE + THF + 5aTHF) | 0.009 | 0.007 | 0.17 | 0.007 | 0.005 | 0.016 | 0.108 | 0.00 | 0.005 | 0.003 |
| 100 × PT’ONE/(THE + THF + 5aTHF) | 0.003 | 0.004 | 0.000 | 0.357 | 0.377 | 0.002 | 0.001 | 0.00 | 0.532 | 0.481 |
| 17-Hydroxylase Deficit | ||||||||||
| (THA + THB + 5aTHB)/(THE + THF + 5aTHF) | 0.121 | 0.035 | 0.000 | 0.238 | 0.098 | 0.085 | 0.159 | 0.00 | 0.259 | 0.085 |
| 100 × THS/(THE + THF + 5aTHF) | 0.016 | 0.006 | 0.000 | 1.00 | 0.673 | 0.012 | 0.018 | 0.00 | 1.08 | 0.478 |
| 11-Hydroxylase Deficit | ||||||||||
| 100 × THDOC/(THE + THF + 5aTHF) | 0.001 | 0.000 | 0.000 | 7.147 | 5.212 | 0.042 | 0.158 | 0.00 | 7.241 | 3.54 |
| 11-Beta HSD Deficit | ||||||||||
| F/E | 0.786 | 0.203 | 0.000 | 0.386 | 0.197 | 0.175 | 0.793 | 0.009 | 0.339 | 0.088 |
| (THF + 5aTHF)/THE | 21.517 | 14.141 | 0.000 | 2.821 | 4.513 | 19.736 | 9.452 | 0.00 | 1.362 | 0.471 |
| (F + E)/THE + THF + 5aTHF | 0.048 | 0.028 | 0.000 | 1.57 | 0.658 | 0.035 | 0.079 | 0.00 | 1.797 | 0.563 |
| ANDRO | ETIO | PT | THE | THF | 5ATHF | |
|---|---|---|---|---|---|---|
| (a) Asperger | ||||||
| ANDRO | 1.00 | 0.81 | 0.81 | 0.14 | 0.36 | 0.32 |
| ETIO | 0.81 | 1.00 | 0.71 | 0.39 | 0.39 | 0.39 |
| PT | 0.81 | 0.71 | 1.00 | 0.39 | 0.40 | 0.39 |
| THE- | 0.14 | 0.39 | 0.39 | 1.00 | 0.90 | 0.39 |
| THF | 0.36 | 0.39 | 0.40 | 0.90 | 1.00 | 0.59 |
| 5ATHF | 0.32 | 0.39 | 0.39 | 0.39 | 0.59 | 1.00 |
| (b) Kanner | ||||||
| ANDRO | 1.00 | 0.98 | 0.96 | −0.34 | −0.43 | 0.20 |
| ETIO | 0.98 | 1.00 | 0.95 | −0.33 | −0.33 | −0.33 |
| PT | 0.96 | 0.95 | 1.00 | −0.42 | −0.38 | −0.41 |
| THE | −0.34 | −0.33 | −0.42 | 1.00 | 0.91 | 0.64 |
| THF | −0.43 | −0.33 | −0.38 | 0.91 | 1.00 | 0.72 |
| 5ATHF | 0.20 | −0.33 | −0.41 | 0.64 | 0.72 | 1.00 |
| (c) Asp. Cont. | ||||||
| ANDRO | 1.00 | 0.91 | 0.84 | −0.32 | −0.30 | −0.11 |
| ETIO | 0.91 | 1.00 | 0.92 | −0.23 | −0.23 | −0.23 |
| PT | 0.84 | 0.92 | 1.00 | −0.12 | −0.15 | 0.91 |
| THE- | −0.32 | −0.23 | −0.12 | 1.00 | 0.91 | 0.79 |
| THF | −0.30 | −0.23 | −0.15 | 0.91 | 1.00 | 0.72 |
| 5ATHF | −0.11 | −0.23 | 0.91 | 0.79 | 0.72 | 1.00 |
| (d) Kan. Cont. | ||||||
| ANDRO | 1.00 | 0.93 | 0.82 | −0.27 | −0.35 | −0.03 |
| ETIO | 0.93 | 1.00 | 0.93 | −0.33 | −0.42 | −0.15 |
| PT | 0.82 | 0.93 | 1.00 | −0.22 | −0.15 | −0.19 |
| THE- | −0.27 | −0.33 | −0.22 | 1.00 | 0.90 | 0.52 |
| THF | −0.35 | −0.42 | −0.15 | 0.90 | 1.00 | 0.54 |
| 5ATHF | −0.03 | −0.15 | −0.19 | 0.52 | 0.54 | 1.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasser, B.A.; Kurz, J.; Dick, B.; Mohaupt, M.G. Steroid Metabolites Support Evidence of Autism as a Spectrum. Behav. Sci. 2019, 9, 52. https://doi.org/10.3390/bs9050052
Gasser BA, Kurz J, Dick B, Mohaupt MG. Steroid Metabolites Support Evidence of Autism as a Spectrum. Behavioral Sciences. 2019; 9(5):52. https://doi.org/10.3390/bs9050052
Chicago/Turabian StyleGasser, Benedikt Andreas, Johann Kurz, Bernhard Dick, and Markus Georg Mohaupt. 2019. "Steroid Metabolites Support Evidence of Autism as a Spectrum" Behavioral Sciences 9, no. 5: 52. https://doi.org/10.3390/bs9050052
APA StyleGasser, B. A., Kurz, J., Dick, B., & Mohaupt, M. G. (2019). Steroid Metabolites Support Evidence of Autism as a Spectrum. Behavioral Sciences, 9(5), 52. https://doi.org/10.3390/bs9050052