
AP2X-8 Is Important for Tachyzoite Growth and Bradyzoite Differentiation of Toxoplasma gondii

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasite Culture and Transfection
2.2. Generation of Endogenously Tagged AP2X-8
2.3. Construction of ap2X-8 Knockout Strain
2.4. Immunofluorescence Assay (IFA)
2.5. Plaque Assay
2.6. Invasion Assay
2.7. Intracellular Replication Assay
2.8. Egress Assay
2.9. Bradyzoite Differentiation Assay
2.10. Mice and Virulence Assay
3. Results
3.1. AP2X-8 Is Constitutively Expressed in the Parasite Nucleus
3.2. Construction of the ap2X-8 Knockout Strain by CRISPR-Cas9
3.3. AP2X-8 Is Essential for Tachyzoite Replication In Vitro
3.4. Loss of ap2X-8 Induces Bradyzoite Differentiation
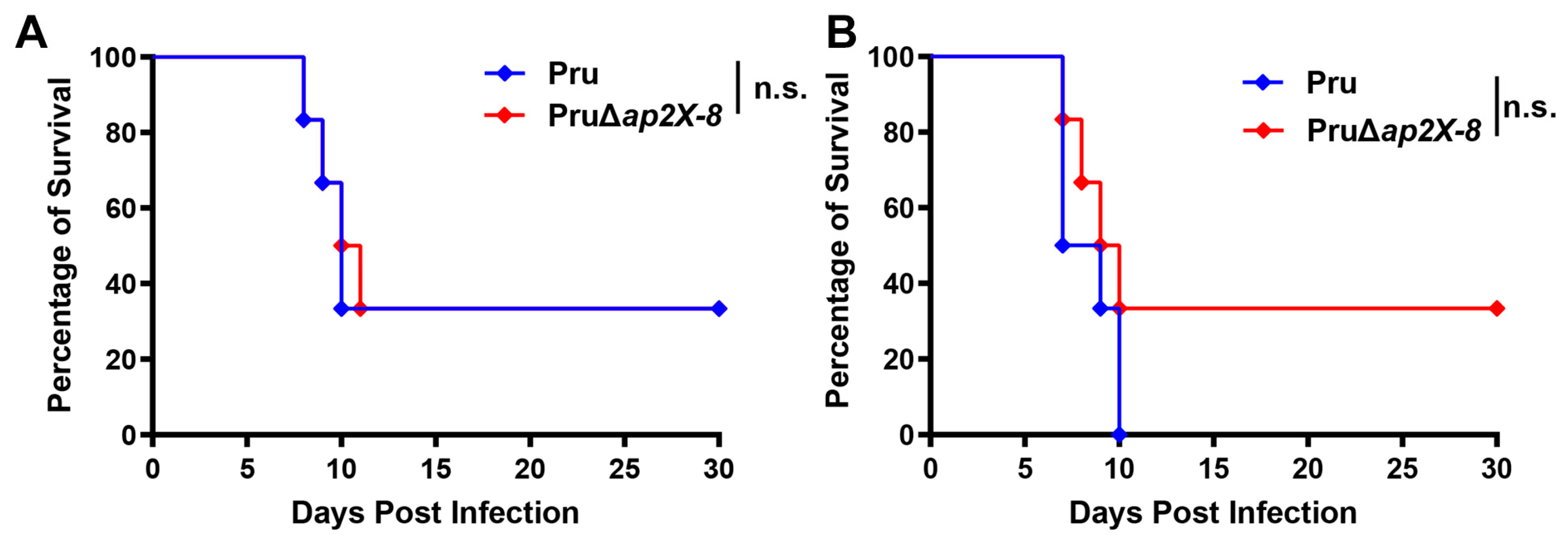
3.5. Deletion of ap2X-8 Does Not Affect the Virulence of T. gondii in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Djurković-Djaković, O.; Dupouy-Camet, J.; Van der Giessen, J.; Dubey, J.P. Toxoplasmosis: Overview from a one health perspective. Food Waterborne Parasitol. 2019, 15, e00054. [Google Scholar] [CrossRef]
- Elsheikha, H.M.; Marra, C.M.; Zhu, X.Q. Epidemiology, pathophysiology, diagnosis, and management of cerebral toxoplasmosis. Clin. Microbiol. Rev. 2021, 34, e00115-19. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Miller, N.L.; Frenkel, J.K. Toxoplasma gondii life cycle in cats. J. Am. Vet. Med. Assoc. 1970, 157, 1767–1770. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.R.; Guerini, M.N.; Jerome, M.; White, M.W. A change in the premitotic period of the cell cycle is associated with bradyzoite differentiation in Toxoplasma gondii. Mol. Biochem. Parasitol. 2003, 131, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Jerome, M.E.; Radke, J.R.; Bohne, W.; Roos, D.S.; White, M.W. Toxoplasma gondii bradyzoites form spontaneously during sporozoite-initiated development. Infect. Immun. 1998, 66, 4838–4844. [Google Scholar] [CrossRef]
- Buchholz, K.R.; Fritz, H.M.; Chen, X.; Durbin-Johnson, B.; Rocke, D.M.; Ferguson, D.J.; Conrad, P.A.; Boothroyd, J.C. Identification of tissue cyst wall components by transcriptome analysis of in vivo and in vitro Toxoplasma gondii bradyzoites. Eukaryot. Cell 2011, 10, 1637–1647. [Google Scholar] [CrossRef]
- Elsheikha, H.M. Congenital toxoplasmosis: Priorities for further health promotion action. Public Health 2008, 122, 335–353. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Dubey, J.P.; Hill, D.E.; Jones, J.L.; Hightower, A.W.; Kirkland, E.; Roberts, J.M.; Marcet, P.L.; Lehmann, T.; Vianna, M.C.; Miska, K.; et al. Prevalence of viable Toxoplasma gondii in beef, chicken, and pork from retail meat stores in the United States: Risk assessment to consumers. J. Parasitol. 2005, 91, 1082–1093. [Google Scholar] [CrossRef]
- Hong, D.P.; Radke, J.B.; White, M.W. Opposing transcriptional mechanisms regulate Toxoplasma development. mSphere 2017, 2, e00347-16. [Google Scholar] [CrossRef]
- Kim, K. The epigenome, cell cycle, and development in Toxoplasma. Annu. Rev. Microbiol. 2018, 72, 479–499. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Babu, M.M.; Iyer, L.M.; Aravind, L. Discovery of the principal specific transcription factors of Apicomplexa and their implication for the evolution of the AP2-integrase DNA binding domains. Nucleic Acids Res. 2005, 33, 3994–4006. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.B.; Lucas, O.; De Silva, E.K.; Ma, Y.; Sullivan, W.J., Jr.; Weiss, L.M.; Llinas, M.; White, M.W. ApiAP2 transcription factor restricts development of the Toxoplasma tissue cyst. Proc. Natl. Acad. Sci. USA 2013, 110, 6871–6876. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.B.; Worth, D.; Hong, D.; Huang, S.; Sullivan, W.J., Jr.; Wilson, E.H.; White, M.W. Transcriptional repression by ApiAP2 factors is central to chronic toxoplasmosis. PLoS Pathog. 2018, 14, e1007035. [Google Scholar] [CrossRef]
- Huang, S.; Holmes, M.J.; Radke, J.B.; Hong, D.P.; Liu, T.K.; White, M.W.; Sullivan, W.J., Jr. Toxoplasma gondii AP2IX-4 regulates gene expression during bradyzoite development. mSphere 2017, 2, e00054-17. [Google Scholar] [CrossRef]
- Wang, J.; Dixon, S.E.; Ting, L.M.; Liu, T.K.; Jeffers, V.; Croken, M.M.; Calloway, M.; Cannella, D.; Hakimi, M.A.; Kim, K.; et al. Lysine acetyltransferase GCN5b interacts with AP2 factors and is required for Toxoplasma gondii proliferation. PLoS Pathog. 2014, 10, e1003830. [Google Scholar] [CrossRef]
- Roos, D.S.; Donald, R.G.; Morrissette, N.S.; Moulton, A.L. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods Cell Biol. 1994, 45, 27–63. [Google Scholar]
- Wang, J.L.; Li, T.T.; Zhang, N.Z.; Wang, M.; Sun, L.X.; Zhang, Z.W.; Fu, B.Q.; Elsheikha, H.M.; Zhu, X.Q. The transcription factor AP2XI-2 is a key negative regulator of Toxoplasma gondii merogony. Nat. Commun. 2024, 15, 793. [Google Scholar] [CrossRef]
- Huynh, M.H.; Rabenau, K.E.; Harper, J.M.; Beatty, W.L.; Sibley, L.D.; Carruthers, V.B. Rapid invasion of host cells by Toxoplasma requires secretion of the MIC2-M2AP adhesive protein complex. EMBO J. 2003, 22, 2082–2090. [Google Scholar] [CrossRef]
- Graindorge, A.; Frénal, K.; Jacot, D.; Salamun, J.; Marq, J.B.; Soldati-Favre, D. The conoid associated motor MyoH is indispensable for Toxoplasma gondii entry and exit from host cells. PLoS Pathog. 2016, 12, e1005388. [Google Scholar] [CrossRef]
- Brown, K.M.; Sibley, L.D. Essential cGMP signaling in Toxoplasma is initiated by a hybrid P-type ATPase-guanylate cyclase. Cell Host Microbe 2018, 24, 804–816.e6. [Google Scholar] [CrossRef] [PubMed]
- Bohne, W.; Hunter, C.A.; White, M.W.; Ferguson, D.J.; Gross, U.; Roos, D.S. Targeted disruption of the bradyzoite-specific gene BAG1 does not prevent tissue cyst formation in Toxoplasma gondii. Mol. Biochem. Parasitol. 1998, 92, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M.; Long, S.; Sibley, L.D. Plasma membrane association by N-Acylation governs PKG function in Toxoplasma gondii. mBio 2017, 8, e00375-17. [Google Scholar] [CrossRef]
- Harris, M.T.; Jeffers, V.; Martynowicz, J.; True, J.D.; Mosley, A.L.; Sullivan, W.J., Jr. A novel GCN5b lysine acetyltransferase complex associates with distinct transcription factors in the protozoan parasite Toxoplasma gondii. Mol. Biochem. Parasitol. 2019, 232, 111203. [Google Scholar] [CrossRef]
- Khelifa, A.S.; Guillen Sanchez, C.; Lesage, K.M.; Huot, L.; Mouveaux, T.; Pericard, P.; Barois, N.; Touzet, H.; Marot, G.; Roger, E.; et al. TgAP2IX-5 is a key transcriptional regulator of the asexual cell cycle division in Toxoplasma gondii. Nat. Commun. 2021, 12, 116. [Google Scholar] [CrossRef]
- Wang, C.; Hu, D.; Tang, X.; Song, X.; Wang, S.; Zhang, S.; Duan, C.; Sun, P.; Suo, J.; Ma, H.; et al. Internal daughter formation of Toxoplasma gondii tachyzoites is coordinated by transcription factor TgAP2IX-5. Cell Microbiol. 2021, 23, e13291. [Google Scholar] [CrossRef]
- Shi, Y.; Li, X.; Xue, Y.; Hu, D.; Song, X. Cell cycle-regulated transcription factor AP2XII-9 is a key activator for asexual division and apicoplast inheritance in Toxoplasma gondii tachyzoite. mBio 2024, 15, e0133624. [Google Scholar] [CrossRef]
- Ying, Z.; Yin, M.; Zhu, Z.; Shang, Z.; Pei, Y.; Liu, J.; Liu, Q. Iron stress affects the growth and differentiation of Toxoplasma gondii. Int. J. Mol. Sci. 2024, 25, 2493. [Google Scholar] [CrossRef]
- Waldman, B.S.; Schwarz, D.; Wadsworth, M.H.; Saeij, J.P., 2nd; Shalek, A.K.; Lourido, S. Identification of a master regulator of differentiation in Toxoplasma. Cell 2020, 180, 359–372.e16. [Google Scholar] [CrossRef]
- Licon, M.H.; Giuliano, C.J.; Chan, A.W.; Chakladar, S.; Eberhard, J.N.; Shallberg, L.A.; Chandrasekaran, S.; Waldman, B.S.; Koshy, A.A.; Hunter, C.A.; et al. A positive feedback loop controls Toxoplasma chronic differentiation. Nat. Microbiol. 2023, 8, 889–904. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.-X.; Wang, M.; Elsheikha, H.M.; Xie, S.-C.; Fu, B.-Q.; Zhu, X.-Q.; Liu, G.-H. AP2X-8 Is Important for Tachyzoite Growth and Bradyzoite Differentiation of Toxoplasma gondii. Animals 2025, 15, 1349. https://doi.org/10.3390/ani15091349
Sun L-X, Wang M, Elsheikha HM, Xie S-C, Fu B-Q, Zhu X-Q, Liu G-H. AP2X-8 Is Important for Tachyzoite Growth and Bradyzoite Differentiation of Toxoplasma gondii. Animals. 2025; 15(9):1349. https://doi.org/10.3390/ani15091349
Chicago/Turabian StyleSun, Li-Xiu, Meng Wang, Hany M. Elsheikha, Shi-Chen Xie, Bao-Quan Fu, Xing-Quan Zhu, and Guo-Hua Liu. 2025. "AP2X-8 Is Important for Tachyzoite Growth and Bradyzoite Differentiation of Toxoplasma gondii" Animals 15, no. 9: 1349. https://doi.org/10.3390/ani15091349
APA StyleSun, L.-X., Wang, M., Elsheikha, H. M., Xie, S.-C., Fu, B.-Q., Zhu, X.-Q., & Liu, G.-H. (2025). AP2X-8 Is Important for Tachyzoite Growth and Bradyzoite Differentiation of Toxoplasma gondii. Animals, 15(9), 1349. https://doi.org/10.3390/ani15091349