1. Introduction
Globally, post-weaning diarrhea (PWD) is an economically important challenge in pig production. The post-weaning period involves several stressful situations for pigs, including a sudden change in the composition and physical form of the diet, separation from the sow, new social groups, and new environment adaption. During the two weeks following weaning, PWD can affect pigs and reduce feed intake, causing poor growth, diarrhea, and mortality [
1]. These factors contribute to the growth of pathogens in the gastrointestinal tract (GIT), primarily enterotoxigenic strains of
Escherichia coli (
E. coli), which are the main causative agents.
The use of colistin as an in-feed antibiotic (ABO) has been effective at alleviating weaning stresses and preventing PWD in piglets. However, although colistin use is prohibited in Europe, it is still used in several nations around the world, notably those in Asia. In 2017, Thailand restricted the long-term use of colistin for the prevention of Gram-negative bacterial infections in animal feed, but it is still permitted for short-term treatments. Consequently, in 2018, the Department of Livestock Development (DLD) restricted the use of colistin for disease prevention in livestock and replaced it with halquinol. Practically, the commercial diets for weaned pigs contain multiple in-feed ABOs, including 300, 300, and 120 ppm of chlortetracycline, amoxycillin, and halquinol, respectively, to treat and inhibit diseases. Nevertheless, the inappropriate and excessive use of ABOs contributes to the widespread emergence and spread of antimicrobial resistance in livestock farms.
Halquinol (trade name Quixalud
®) is an antimicrobial used as an in-feed ABO in poultry and swine. In swine, it is used as a growth promotant and for the treatment of intestinal infections caused by
E. coli and
Salmonella spp. The recommended concentration of halquinol ranges from 12% to 60%, which can be associated with various dosing regimens and a withdrawal period of 10 consecutive days [
2]. Researchers have demonstrated that 120 ppm of in-feed colistin led to a greater final body weight gain (BWG; 20.4 vs. 19.0 kg;
p < 0.001) and average daily gain (ADG; 344.1 vs. 304.9 g/d;
p < 0.05) in pigs than 120 ppm of halquinol [
3]. In addition, the introduction of halquinol into the diet was not sufficient to change the composition of the microbiota or cause changes in the structure of the small intestine [
4]. Nevertheless, halquinol can slow peristalsis in the gut [
5]; this decelerated passage rate of digesta in the GIT may facilitate improved absorption. Beyond this, halquinol was invented to minimize the antimicrobial resistance that is common for Gram-negative bacteria, including members of the
Enterobacteriaceae family [
6,
7].
Although many studies have been performed investigating the efficacy and performance of halquinol in weaned pigs, along with its elimination effect on pathogenic bacteria in GIT, to the best of our knowledge, there are limited data available comparing the effect of different concentrations of halquinol with colistin in the early weaning period in terms of digestion and microbiome modulation. In the present study, the hypothesis proposed that colistin could be replaced with an appropriate amount of halquinol supplementation, which would improve nutrient digestion, modulate the microbiome, and contribute to enhanced growth performance during the early weaning period. Therefore, the present study aimed to investigate the efficacy of colistin and halquinol, including assessments of performance, diarrhea incidence, coefficient of apparent total tract digestibility (CATTD), fecal volatile fatty acids (VFAs), and fecal microbiota.
3. Results
3.1. Growth Performance
The average performance results are presented (
Table 2).
The BW, ADG, and FCR were not influenced by the in-feed ABO (p > 0.05). Meanwhile, on D10, the FCR was lower in pigs fed the T3 and T4 diets than that in pigs fed the CON, T2, and T5 diets (p < 0.05). Piglets receiving 240 ppm of halquinol (T4) had the lowest ADFI (p < 0.01). Additionally, the in-feed ABO groups had lower ADFI (p < 0.01) than the CON group. Therefore, these results show the reduction in FCR in piglets receiving 180 ppm (T3) and 240 ppm (T4) halquinol (p < 0.01). There was no significant difference between the CLT and HAL groups (T2 vs. T3–T5, p > 0.05) in terms of the mean values of BW, ADG, ADFI, and FCR. However, when conducting comparisons between the different HAL groups (T3 vs. T4 vs. T5), ADFI (linear and quadratic, p < 0.05) values were lower for piglets fed T4 compared to piglets fed T3 and T5 from days 1 to 5, 6 to 10, and during the overall period (days 1 to 10).
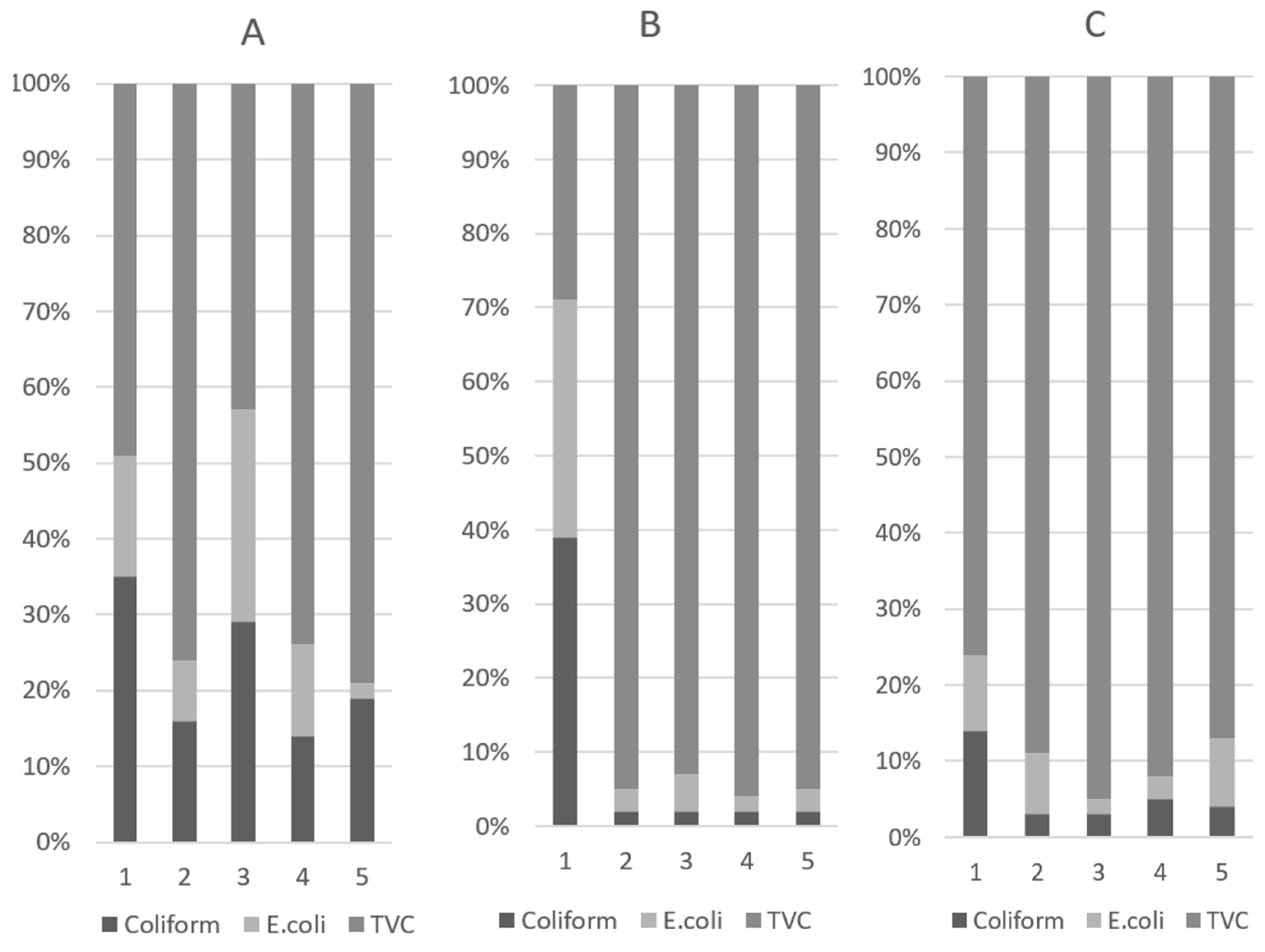
3.2. Fecal Microbial Populations
The results of the analysis of fecal microbial populations and their proportions are presented in
Table 3 and
Figure 2, respectively. On D10, the halquinol groups had lower coliform (
p < 0.01) and
E. coli (
p < 0.01) levels than the other treatment groups. In particular, adding 180 or 240 ppm of halquinol to the pigs’ diets significantly decreased the abundance of coliform (
p < 0.01) and
E. coli (
p < 0.01) compared to adding colistin. This indicated that piglets fed colistin and halquinol had lower amounts of pathogenic bacteria compared with those fed a basal diet with ABOs. On D10, in pigs treated with 120 ppm (T3) of in-feed colistin, the TVC revealed variations in microbial populations (
p < 0.001). In contrast, colistin-treated pigs also had higher coliform and
E. coli (
p < 0.01) proportions than pigs fed with 240 and 360 ppm halquinol.
The results for the total viable count (TVC) are also summarized in
Table 3. On D5, the TVCs of the in-feed ABO groups were higher than the TVC in the control group (
p < 0.01). On D10, the TVC of colistin was higher than that in the halquinol groups (
p < 0.01). Furthermore, comparisons within the HAL groups (T3 vs. T4 vs. T5) showed that increasing the halquinol concentration in the diet decreased fecal coliform (linear,
p < 0.05) and
E.coli (linear and quadratic,
p < 0.05) abundance and the TVC (linear,
p < 0.05) over the experimental period (day 1 to 10).
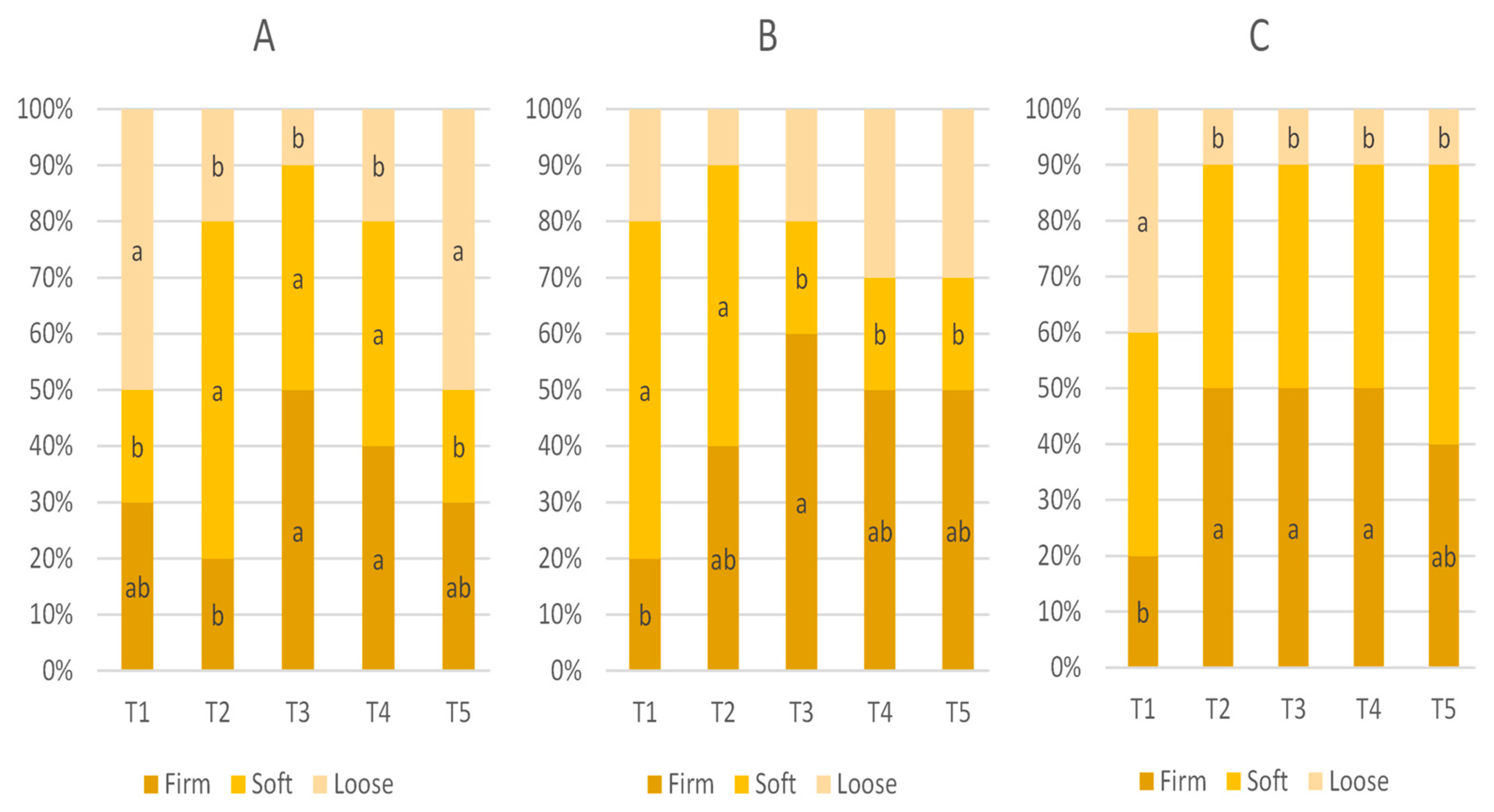
3.3. Fecal Score
The effect of in-feed ABO on fecal score is presented
Table 4 and
Figure 3. The mean fecal score of weaned pigs on D1, D5, and D10 was not statistically significant (
p > 0.05).
There was also no significant difference in the mean fecal score values when using CLT or HAL (T2 vs. T3–T5, p > 0.05); however, the scores were higher in pigs receiving the T2 and T4 diets (120 ppm of colistin and 240 ppm of halquinol, respectively).
3.4. Coefficient of Apparent Total Tract Digestibility
The effects of in-feed ABO on the CATTD were analyzed on D10 (
Table 5). No significant differences between any parameter of the CATTD were observed among treatments, except for EED in the T4 group (
p < 0.01). Moreover, there was also no significant difference between CLT and HAL treatments (T2 vs. T3–T5,
p > 0.05) for the mean CATTD values of GED, DMD, OMD, CPD, EED, and CHOD.
3.5. Fecal Volatile Fatty Acid Production
To analyze the impacts of in-feed ABO on fecal VFA production, fecal VFA assessments were conducted on D10 (
Table 6). When 240 ppm (T4) and 360 ppm (T5) of halquinol were added to the diet, the ratios of propionate and A:P changed significantly (
p < 0.01). In addition, there was also a significant difference between CLT and HAL (T2 vs. T3–T5,
p < 0.05) in terms of the mean propionate and A:P ratios (
p < 0.001). Supplementation with increasing halquinol concentrations linearly decreased the A:P ratio (
p < 0.05).
However, the total VFA, acetate, butyrate, isobutyrate, and valerate levels did not differ between the CLT and HAL groups (p > 0.05). In addition, there was no significant difference in the fecal concentrations of total VFAs, acetate, butyrate, isobutyrate, and valerate among the different groups (p > 0.05).
3.6. Overall Sequencing Data and Microbial Diversity of the Piglet Fecal Samples
Weaned pig fecal samples that were subjected to 16S rRNA amplicon sequencing generated 1,111,765 raw sequences (n = 6), ranging from 65,625 to 133,772 raw sequences per sample, after quality control and filtering steps. The Good’s coverage for all samples was always greater than 0.99. The summary of sequence counts and OTUs that passed the steps of filtering, clean-up, and normalization were grouped into 621 OTUs present in at least six samples. A total of 7 phyla, 37 families, and 83 genera were identified in the fecal samples from weaned pigs across all treatment and sampling points, with Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria being the predominant phyla.
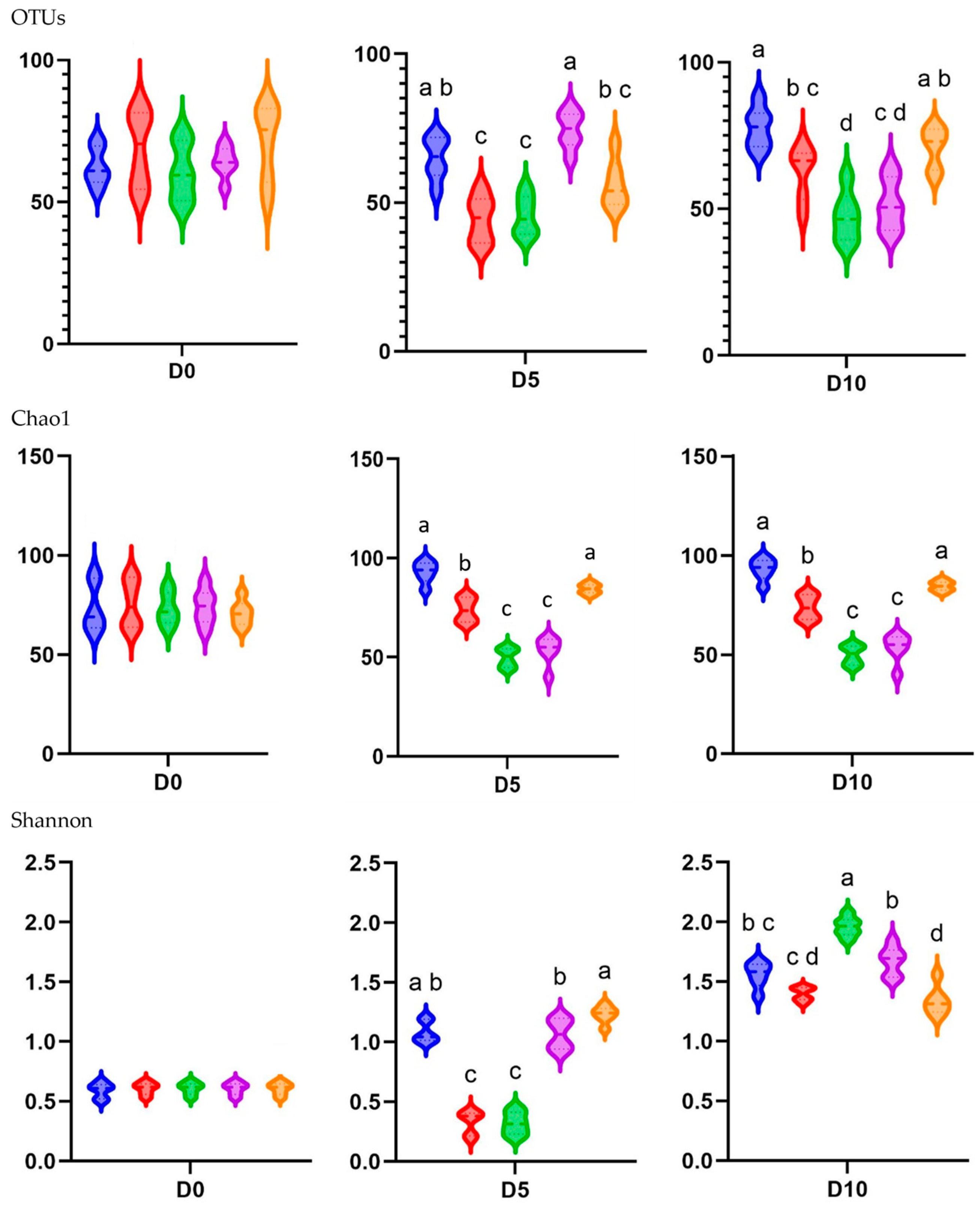
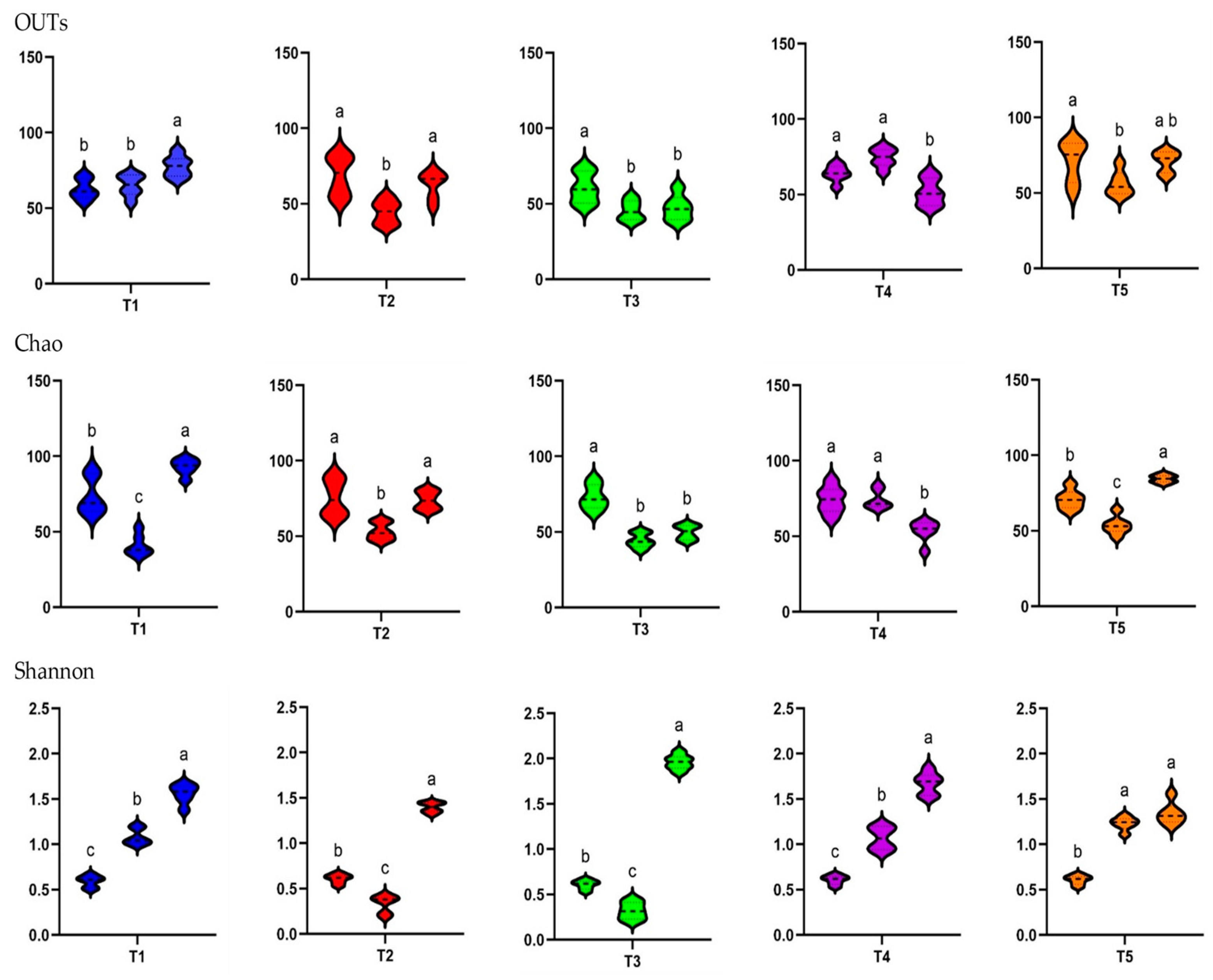
3.7. Alpha Diversity Patterns Between Treatment Groups and Sampling Times
In the alpha diversity analysis, which evaluated the diversity within piglet fecal microbiota, the observed OTUs and Chao1 and Shannon indices were calculated to assess the diversity and richness of the fecal microbiota in in-feed ABO and control piglets (
Figure 4).
In terms of the different treatments, the observed OTUs and Shannon alpha diversity index were higher for the T1 and T4 groups than for the T2 and T3 groups at D5 (p < 0.05). The Chao1 diversity index was higher for the T1 and T5 groups than for the T2, T3, and T4 groups at D5 (p < 0.05). The observed species, Chao1, and Shannon indices indicated significantly lower diversity on both D5 and D10 when dietary supplementation with in-feed ABOs was used (p < 0.05) compared with the control diet. The exception was the Shannon index on D10, where 180 ppm of halquinol (T3) led to a higher diversity than that in the CON group (T1).
There was a consistent increase in alpha diversity as piglet age increased, as indicated by the observed OTUs and Shannon index. The observed OTUs, indicative of species richness, dramatically increased from day 0 to day 10 (
p < 0.05). The Shannon index, indicative of species richness and evenness, exhibited a pronounced increase from D1 to D10, demonstrating a significant difference between these two time points (
p < 0.05). Overall, as illustrated in
Figure 5, weaning age significantly affected (
p < 0.05) the observed species, Chao1, and Shannon indices across the treatments.
3.8. Beta Diversity
As shown in
Figure 6, a PCoA was conducted based on phylum.
Each point in the PCoA represents the microbial ecology of a piglet; points closer together have more comparable microbial ecology. PCoA based on Bray–Curtis and weighted UniFrac distances revealed clear separation of the piglets’ fecal microbiota in the T3 and T4 diets at D10; however, the other microbiota mixed together from D1 to D10. Conversely, unweighted UniFrac distances revealed a significant alteration in the fecal microbiota composition of piglets from D1 to D10 across all diets.
When PCoA was plotted on the weighted UniFrac distances, T3 and T4 exhibited a significant difference in microbiota structure, but the administration of antibiotics changed the gut microbiota structure of piglets in the in-feed ABO group. The noteworthy alterations in the fecal microbiota structure from D1 to 10 were also verified by dramatic intra-individual variations in alpha diversity from D1 to D5 and D5 to D10, which were also significantly affected by age and diet supplementation. Such inter-individual and intra-individual structural variations were also supported by weighted UniFrac distance.
3.9. Effects of In-Feed ABO on Bacterial Composition in Fecal Samples
Across all the samples,
Proteobacteria (59%),
Firmicutes (17%),
Bacteroidetes (15%),
Actinobacteria (6%), and
Fusobacteria (2%) were the most abundant phyla (
Figure 7). At the genus level, there were nine taxa whose abundance exceeded 1% of the total sequences.
Escherichia (33%),
Acinetobacter (14%),
Enterococcus (14%),
Bacteroides (9%), and
Klebsiella (7%) were the most predominant genera in the fecal microbiota.
Phocaeicola (4%) and
Parabacteroides (1%) were the two dominant genera in
Bacteroidetes.
Erysipelatoclostridium (2%) was the main genus in
Firmicutes. The genus
Fusobacterium, classified under
Fusobacteria, constituted 2% of the total sequences.
The phylum-level composition of the gut microbiota in each sample revealed significant variations in bacterial community structure among piglets of different ages (D1 vs. D5 vs. D10). Proteobacteria, the most prevalent phylum, had notable reductions in relative abundance on day 5 compared to day 10, but Bacteroides demonstrated considerable increases in relative abundance. The phylum-level results showed that weaning on both day 5 and 10 did not affect (p > 0.05) the abundance of Firmicutes, Bacteroidetes, and Actinobacteria. Other phyla (Spirochaetes, Fusobacteria, and Synergistetes) were found at minimal relative abundances, with no significant differences observed (p > 0.05).
At the genus level, there was significant heterogeneity in the gut microbiota composition dependent on piglet age (D1 vs. D5 vs. D10).
Escherichia, the most prevalent phylum, had a notable decline in relative abundance on day 5 compared to day 10, but Bacteroides demonstrated a large increase in relative abundance. The relative abundances of Acinetobacter, Klebsiella, Enterococcus, Proteus, Fusobacterium, and Erysipelatoclostridium, representing the main phylum in the analysis, did not vary significantly with age.
We also conducted a genus-level comparison of the microbial population by dietary treatment. In-feed ABO supplementation led to a decrease in Escherichia abundance compared to the CON group on D5 and a significant decrease in Escherichia on D10 (p < 0.01). Bacteroidetes was present at significantly higher levels in the Hal group (16.02 to 20.94%; p < 0.01) than in the CON and HAL groups (9.86 and 7.82%, respectively), despite the fact that the CON and HAL groups did not differ much. The relative abundance of Acinetobacter in the COL group on D5 and D10 (24.04 and 21.01%, respectively) was significantly higher than that in the CON and HAL group (p < 0.05). The relative abundance of Klebsiella was significantly higher in the CLT group on D10 (17.68%), while it was significantly lower in the HAL group on D10 (2.74%; p < 0.01). Enterococcus was present at significantly higher levels in the CLT and HAL groups (17.93% and 15.05 to 21.21%; p < 0.01) than in the CON group (11.04%) on D10.
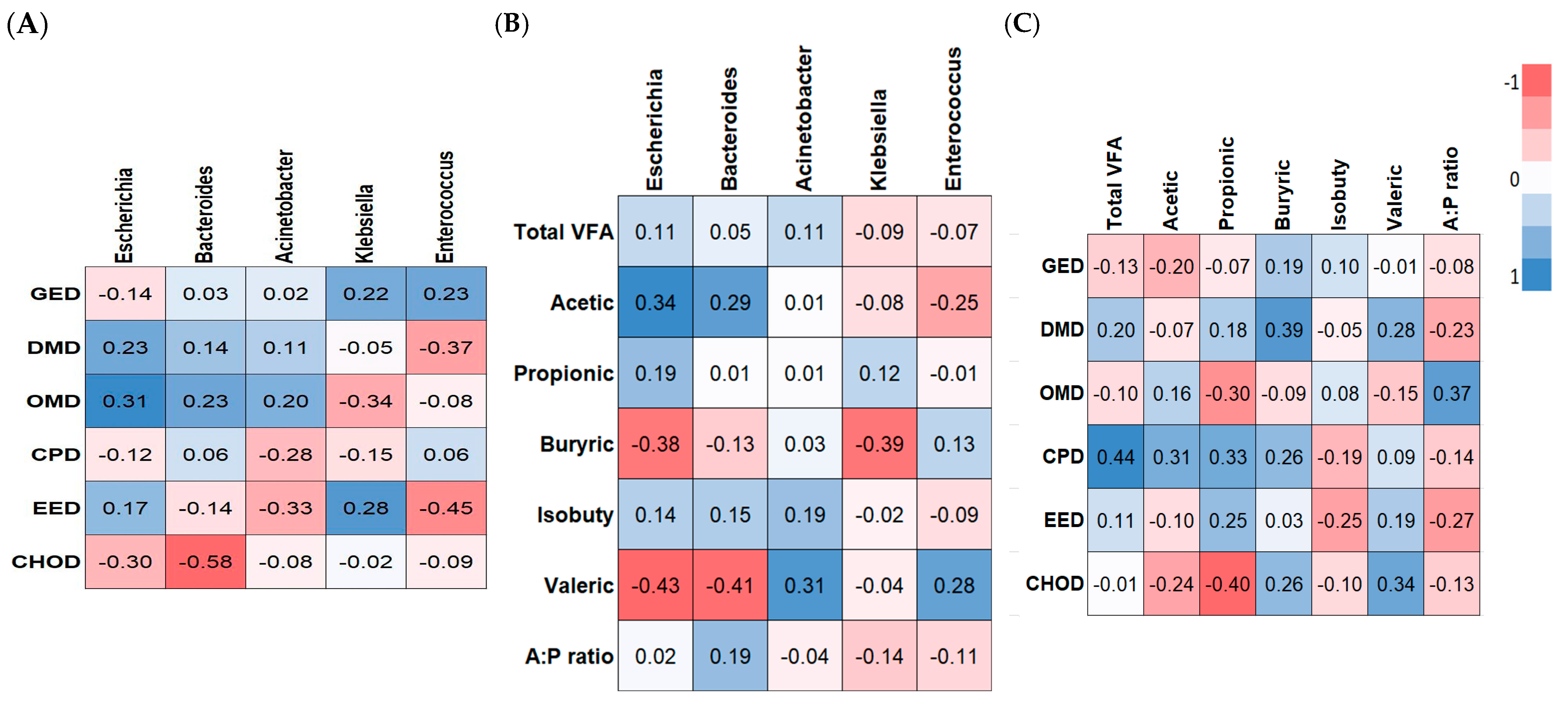
3.10. Correlation Analysis
The possible relationships between the changes in the composition of fecal microbiota and ATTD (
Figure 8A), VFAs (
Figure 8B), and ATTD vs. VFAs (
Figure 8C) were assessed using Pearson’s correlation analysis. The genus Bacteroides was negatively correlated with CHOD levels (R = −0.58,
p < 0.01). The genus
Enterococcus was negatively correlated with EED (R = −0.45,
p < 0.05). The genera Acinetobacter, Klebsiella, and Enterococcus trended toward being negatively correlated with EED, OMD, and DMD, respectively (R = −0.30,
p < 0.10). Only Escherichia trended toward being positively correlated with OMD (R = 0.31,
p < 0.10).
The genus Escherichia trended toward being positively correlated with levels of acetic acid (R = 0.36, p < 0.10), while it was significantly negatively correlated with the levels of butyric and valeric acids (R = 0.38 and R = 0.43, p < 0.05). The genus Bacteroides was negatively correlated with the levels of valeric acid (R = 0.41, p < 0.05). The genus Klebsiella was negatively correlated with butyric acid levels (R = 0.39, p < 0.05). The genus Acinetobacter trended toward being positively correlated with the levels of valeric acid (R = 0.31, p < 0.10). The concentration of propionic acid was negatively correlated with CHOD levels (R = 0.40, p < 0.05).
4. Discussion
The impact of the substitution of in-feed ABO in swine production was evaluated. According to our hypothesis, these antibiotics can influence the commensal microbial community and could promote a healthy GIT in the early weaning period. This study examined the composition and functional aspects of the fecal microbiota in the weaning period via fecal sample analysis.
Our findings indicate that in-feed ABOs did not positively influence BWG and ADG but significantly affected ADFI and FCR, especially in the group receiving 240 ppm halquinol (T4). This was verified by Barbosa et al. [
16] and Grecco et al. [
17], whose experiments used halquinol levels ranging from 60 to 200 ppm. Thus, in-feed ABO supplementation could not only control PWD but also optimize growth performance. Comparative studies on the use of in-feed ABO, such tilmicosin, amoxicillin, and doxycycline, in the weaning period concluded that growth performance was not influenced by these in-feed ABOs, despite the fact that pigs receiving ABO supplementation reached slaughter 2 kg heavier than those that did not [
18]. These results support our hypothesis that the inclusion of 240 to 360 ppm halquinol in a pig’s diet may contribute to a balanced microbial composition, a healthy gastrointestinal environment, and the preservation of intestinal structure, which contributes to growth in later development stages. Tummarak et al. [
3] added 120 ppm of halquinol to pig diets, which was insufficient to improve piglet performance metrics compared to 120 ppm of colistin. The discrepancy could be explained by the mechanisms of colistin. Colistin targets Gram-negative bacteria, including
Salmonella,
Escherichia coli,
Klebsiella, and
Pseudomonas. It binds to phospholipids and lipopolysaccharides, disrupting bacterial cell membranes and inhibiting the release of endotoxins from dead bacteria.
Based on our review of the literature, concentrations of 180 to 360 ppm of halquinol were selected and validated for our experiment as in-feed ABO replacements to improve the efficiency of feed utilization, reduce harmful microorganisms, and promote beneficial microbial growth in the GIT without negative consequences for the piglet [
3,
16,
17].
This study showed that piglets fed 240 ppm halquinol had significantly lower ADFI during the first 10 days after weaning, resulting in not only lower FCR but also lower diarrhea incidence. The association between pathogenic bacteria and ADFI could be explained by the intestinal environments of newly weaned pigs being susceptible to invasion by pathogenic bacteria, which impairs the digestive system. The consequence of this is damage to the GIT, which affects the pigs’ ability to digest and absorb nutrients; therefore, their food intake might need to increase significantly just to reach their minimum nutritional requirements. Additionally, undigested materials may be available for utilization by, e.g., enterotoxigenic
E. coli, resulting in infectious diarrhea [
19]. Therefore, the low ADFI in piglets fed 240 ppm halquinol may actually indicate the effectiveness of the in-feed ABO in enhancing nutrient utilization, given the low concentration of undigested and unabsorbed contents in the intestinal lumen [
20]. Moreover, the presence of quadratic contrast affected piglets’ ADFI over the entire period. This may explain why high levels of halquinol supplementation (T5; 360 ppm of halquinol) were found to suppress growth performance and reduce antimicrobial effectiveness.
During weaning, the intestinal flora underwent a dramatic change in diversity and composition, and in-feed ABO supplementation may have changed the number of bacteria in each species [
20,
21]. In the present study, the fecal
E.coli levels and TVC were significantly decreased in halquinol groups compared to the colistin group at D10 (T2 vs. T3 to T5). This suggests that halquinol has the potential to significantly change the microbial structure of the host’s gut by boosting the abundance of beneficial bacteria and lowering the prevalence of harmful bacteria [
22].
Surprisingly, 240 ppm of halquinol can enhance fat digestion, leading to energy storage. This treatment, with its associated enhanced feed efficiency (ADFI and FCR) and reduced ADFI, FCR, and passage of food in the gut, may help enhance absorption in the first three weeks after weaning [
5]. Although the biological relevance of lingual lipase diminishes with pig age, lingual and pancreatic lipase activities can be comparable to or higher than stomach lipases in neonatal piglets [
23]. The interaction between halquinol and fat digestion is thought to enhance digestion capacity, which subsequently leads to a deceleration of the passage rate process. This enhances the digestion of lipid fat, which may require a longer duration for digestion compared to other nutrients.
Subsequently, we determined that the propionate concentrations in feces were significantly higher in the halquinol group than in the control and colistin groups. The results indicated that undigested CHO could be a substance the allows beneficial microbes to produce propionate, as confirmed in the correlation study. Propionate is known to regulate cellular function and the immune response [
24]. Collectively, supplementation with halquinol increased propionic acid levels and fat digestion, both of which are related to improved feed efficiency. Although improvements in feed efficiency, reduced diarrhea occurrence, and increased VFA production were observed in animals that were fed halquinol, none of the dietary treatments positively affected piglet performance. Halquinol’s mechanisms of action are described by Hall et al. [
2]; it primarily functions as an antimicrobial, anti-inflammatory, and gut health modulator through stabilizing gut pH, disrupting bacterial DNA and enzyme systems, reducing pathogenic bacteria, and enhancing nutrient absorption by maintaining gut integrity.
The diversity and richness indices are frequently used to measure the gut microbiota’s functional stability [
20,
21]. Our study demonstrated that the abundance and diversity (OTUs and Shannon index) of fecal microbiota increase as weaned piglets age. Our findings are in accordance with Feehan et al. [
25], who asserted that alpha diversity undergoes substantial alterations during this transition period. The present study revealed that the alpha diversity of the fecal bacteriome remained stable from day 0 to day 5 and subsequently grew from day 5 to day 10. Additionally, in-feed ABOs can diminish alpha diversity and induce alterations in gut microbiota due to their broad-spectrum activity and capacity to eliminate or inhibit the proliferation of both pathogenic and beneficial microorganisms [
26].
Pig fed in-feed colistin and halquinol had decreased fecal microbiota diversity (OTUs and Chao1). This result was consistent the findings of from Abeles et al. [
27] and Chen et al. [
28], who found that alpha diversity was reduced with in-feed ABO supplementation. Thus, the less diverse and more dynamic microbial community in the guts of pigs in the early weaned period may be more vulnerable than that in weaning pigs and thus can be easily influenced by environmental factors, for instance, the antibiotic exposure in the current study. In fact, the authors of [
29] found that more efficient microbiomes are prone to reduced diversity and result in a smaller variation in metabolites. Nevertheless, within that diminished pool of metabolites, there could be a greater concentration of biologically valuable metabolites that are more readily accessible for utilization by the host.
In contrast, increases in microbial diversity are often positively associated with the resilience of microflora against disease invasion. Similarly, the improvement of alpha and beta diversity indicated that the relative abundance of beneficial bacteria had increased [
28]. The contribution of diversity was mainly reflected in the modification, structure, and abundance of the microbiota and the maintenance of intestinal homeostasis. Furthermore, the antibiotics significantly influenced microbial alterations in the foregut but not in the hindgut. Additionally, early exposure to antibiotics does not substantially alter the intestinal ecological environment.
The alpha diversity of the fecal microbiome appears to follow a linear trend, likely due to the strong impact of halquinol toward restoring diversity. Overall, piglets receiving 360 ppm halquinol had a lower fecal bacterial biodiversity than the other groups. Supplementation with 180 and 240 ppm halquinol led to significant differences in fecal microbiome composition, reflecting a richness in alpha diversity. Similarly, it has been shown that pigs fed antibiotics have enhanced expression of functional genes that affect their microbiome and are associated with energy production and feed efficiency, suggesting that consumption of antibiotics leads to a more effective utilization of energy from feed [
30]. Moreover, the alpha diversity results, except for the Shannon index, indicate there are age-related differences, with diversity gradually increasing in the piglets as they age from 0 to 10 days after weaning. Alternatively, Huaman et al. [
31] demonstrated that weaned pigs receiving 200 ppm of halquinol had lower Chao1 alpha diversity and observed OTUs than a group receiving synbiotics. Living microorganisms, such as probiotics or synbiotics, often increase microbial richness and stability, which increases alpha diversity in weanling pigs, while biological and environmental factors can affect the outcome [
31].
Beta diversity analysis can corroborate this finding. Halquinol influenced the composition of fecal microbiota in weaned pigs, altering the distance between control group and in-feed ABO group samples in PCoA analysis. However, the beta diversity between all three age points was different, indicating that both dietary treatment and age affected it. Although there was a significant increase in Shannon diversity on D10, taxonomic changes occurred and were the primary driver of diversity changes.
Furthermore, altered microbial composition has been associated with the production and composition of VFAs in the colon. The current study revealed that the fecal concentrations of VFAs, specifically propionic acid, dramatically increased in piglets whose diets were supplemented with 240 or 360 ppm halquinol; propionic acid is an essential metabolite of gut microbiota and may promote energy homeostasis and alleviate inflammation and metabolic disorders in the colon [
32]. In-feed ABO consumption was not the only factor influencing microbiota diversity; the PWD caused by ETEC also affected microbiota imbalances via the increased growth of species of pathogenic bacteria [
33].
Our findings reveal that
Proteobacteria (59%),
Firmicutes (17%), and
Bacteroidetes (15%) were the predominant phyla among the five groups of weaned piglets, supporting earlier studies in pigs [
33,
34]; however, the proportion of
Proteobacteria was indicative of the high frequency of PWD. Elevated levels of
Helicobacteraceae, a family within
Proteobacteria, have been documented to weaken the protective mucous layer in the gut, whereas
Prevotellaceae, a member of the
Bacteroidetes phylum, has been linked to fortifying the intestinal mucosa of healthy pigs fed plant-based diets [
35,
36]. Moreover, the high percentage of rice bran in the diet might be a reason for the predominance of
Proteobacteria, as this ingredient contains phytic acid and a trypsin inhibitor [
37], which negatively affect the digestive system.
The current study shows that in the early weaning period (first 5 days post-weaning), the fecal microbiota structures were similar among the five dietary treatments, except that the relative abundance of
Enterococcus was higher in pigs fed 240 to 360 ppm halquinol than in pigs in the other groups. In particular, halquinol had no effect on the bacteria that are the most common in feces. Compared to the control groups, the types of bacteria that were most common in the halquinol groups were
Escherichia (24%),
Bacteroides (18%), and
Enterococcus (17%). The rise in beneficial
Enterococcus bacteria belonging to the phylum
Firmicutes could be positively correlated with metabolites associated with inducing oxidative stress [
38].
A pig’s gut microbiota colonization begins at birth but continues to develop during the weaning process [
39]. Even though
E. coli is among the first bacteria to colonize in piglets’ intestines [
40], it is still dominant after weaning. The presence of ETEC infection or heightened levels of
E. coli during the post-weaning phase may influence the gut microbiota. ETEC K88 infection decreased microbial diversity and suppressed
Bacteroidetes and
Firmicutes proliferation in the feces and jejunum of weaned pigs [
41]. The proliferation of
Firmicutes and
Bacteroidetes in pigs’ gastrointestinal tracts contributes to the animal’s general health in numerous ways. In this study, pigs fed with 360 ppm halquinol (T5) had higher abundances of
Enterococcus and
Erysipelatoclostridium, which belong to phylum
Firmicutes, as well as higher
Bacteroides abundance compared to control and colistin diets.
Firmicutes are present in the GIT of pigs and are responsible for the body’s energy balance and utilization of carbohydrates in the gut [
42].
Bacteroides are crucial for health, as they generate butyrate, which activates T cell-mediated immune responses that minimize pathogenic bacteria’s colonization of the gastrointestinal tract [
43].
The highest abundances of
Bacteroidetes were mostly found in pigs fed with halquinol.
Bacteroides and
Parabacteroides, which are present in the early stages of life, have been documented to synthesize gamma-aminobutyric acid (GABA), which is linked to growth [
44]. Moreover, post-weaning diarrhea has been linked to the low abundance of the genus
Bacteroides, which is also thought to be a mucin glycan degrader [
45]. Microbial glycan degradation results in the generation of metabolites that help to modulate mucosal health.
The phylum
Bacteroidetes has a unique ability to digest carbohydrates, including simple and complex carbohydrates as well as polysaccharides [
46]. Conversely, an elevation in the prevalence of
Bacteroides is typically observed in cases of ulcerative colitis, colorectal cancer, and functional gastrointestinal problems [
47].
Furthermore, it was observed that
Fusobacteria were only present in pigs fed diets supplemented with halquinol. This observation may be attributed to a decrease in the proportion of beneficial bacteria and an increase in the prevalence of harmful bacteria. Generally, based on this study’s control group, the abundance of
Fusobacterium decreased by more than 70% from D1 to D10. As a result, newly weaned pigs’ gut microbial composition may be readily disrupted by a decrease in diversity, making pigs more susceptible to pathogenic microbes. On the other hand, halquinol fails to modulate and control the invasion of
Fusobacterium. Infections with
Fusobacterium spp. have been linked to a wide range of pathologies. Relevance networking also revealed that
Fusobacterium was negatively associated with α-diversity, as shown previously [
48,
49].
Fusobacterium is emerging as a pathogen that is associated with gastrointestinal diseases such as ulcerative colitis [
50].
Enterococcus faecium was found in both the control group and the in-feed ABO groups. This species is indicative of normal gut flora and is the most common type of bacteria in piglets when they are young. A high abundance of the
Enterococcus genera may be essential for post-weaning piglets to digest plant-based diets.
Enterococcus can form biofilm microcolonies throughout the gastrointestinal tract, increase immunoreactivity, and reduce instances of diarrhea [
51]. Researchers have conducted numerous studies to evaluate the probiotic characteristics of
Enterococcus strains, primarily
E. faecium. Feeding pigs with the probiotic
Enterococcus was found to reduce intestinal pathogens, improve the gut microbiota balance, and improve the growth and feed conversion of piglets [
52]. Thus, in the current study, the lack of relative abundance data for various genera in the feces of piglets may be explained by the short experiment duration, which might have been insufficient to fully impact the microbiota diversity.
Finally, our results indicate that excessive
Escherichia abundance and insufficient
Bacteroides abundance serve as indicators of digestive issues in the early weaning period (D0 to D5), because changes in their abundance have been correlated with impaired nutrient digestion and various disorders resulting from PWD [
53]. At weaning, the intestinal microbiota is unstable and the animals’ immune and digestive systems are not fully developed, which makes the piglets highly susceptible to PWD [
39]. The halquinol-mediated alterations in beneficial microbiota may be attributed to fat utilization and VFA regulation, which might be key modulators in the maintenance of intestinal homeostasis after weaning.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}