
Effects of Adding Bacillus coagulans BCH0 to the Diet on Growth Performance, Tissue Structures, and Gut Microbiota in Broilers
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of B. coagulans BCH0 Additive
2.2. Animals and Experimental Location
2.3. Animals and Trial Design
2.4. Sample Collection and Assay
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
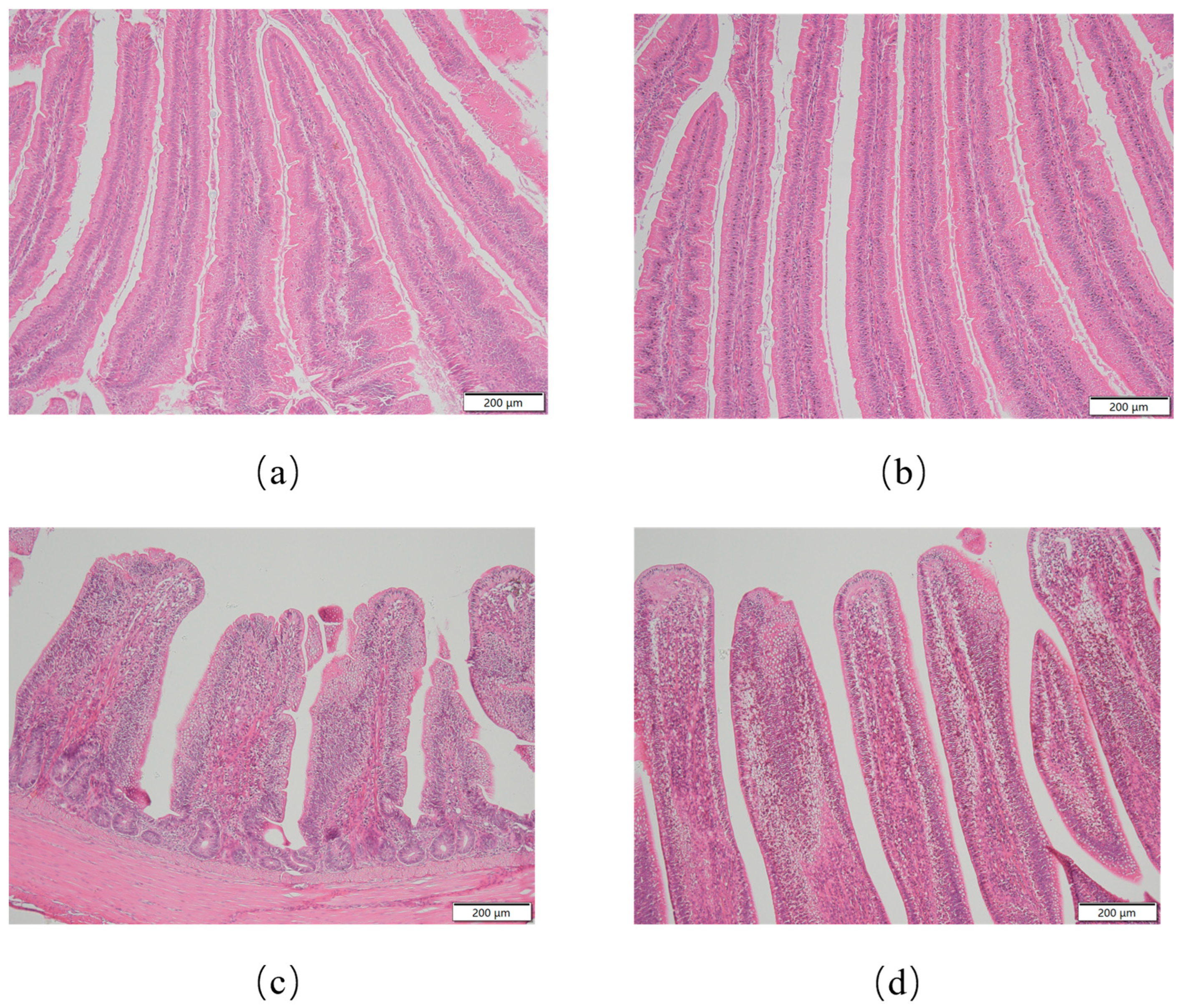
3.2. Intestinal Morphology
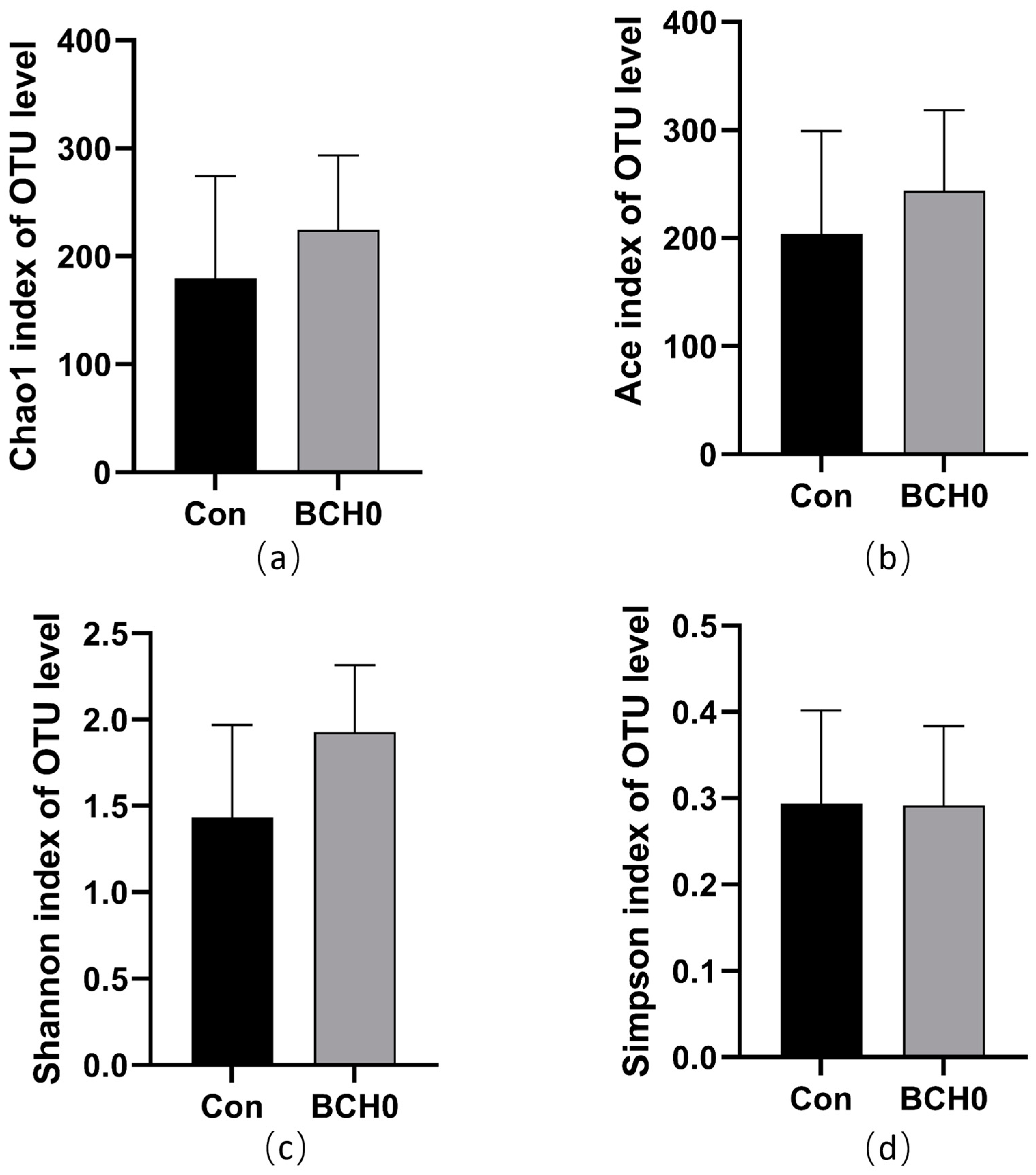
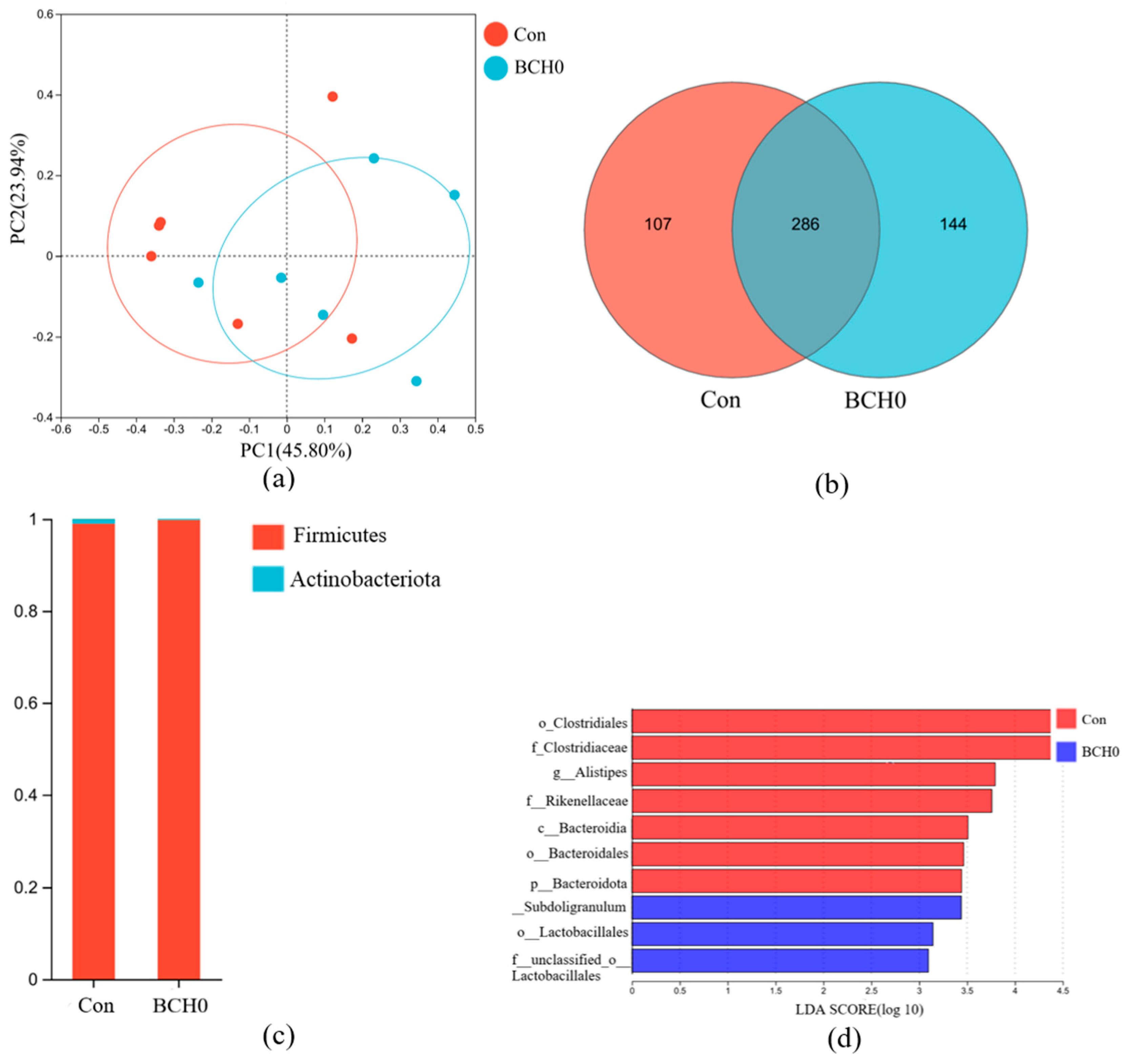
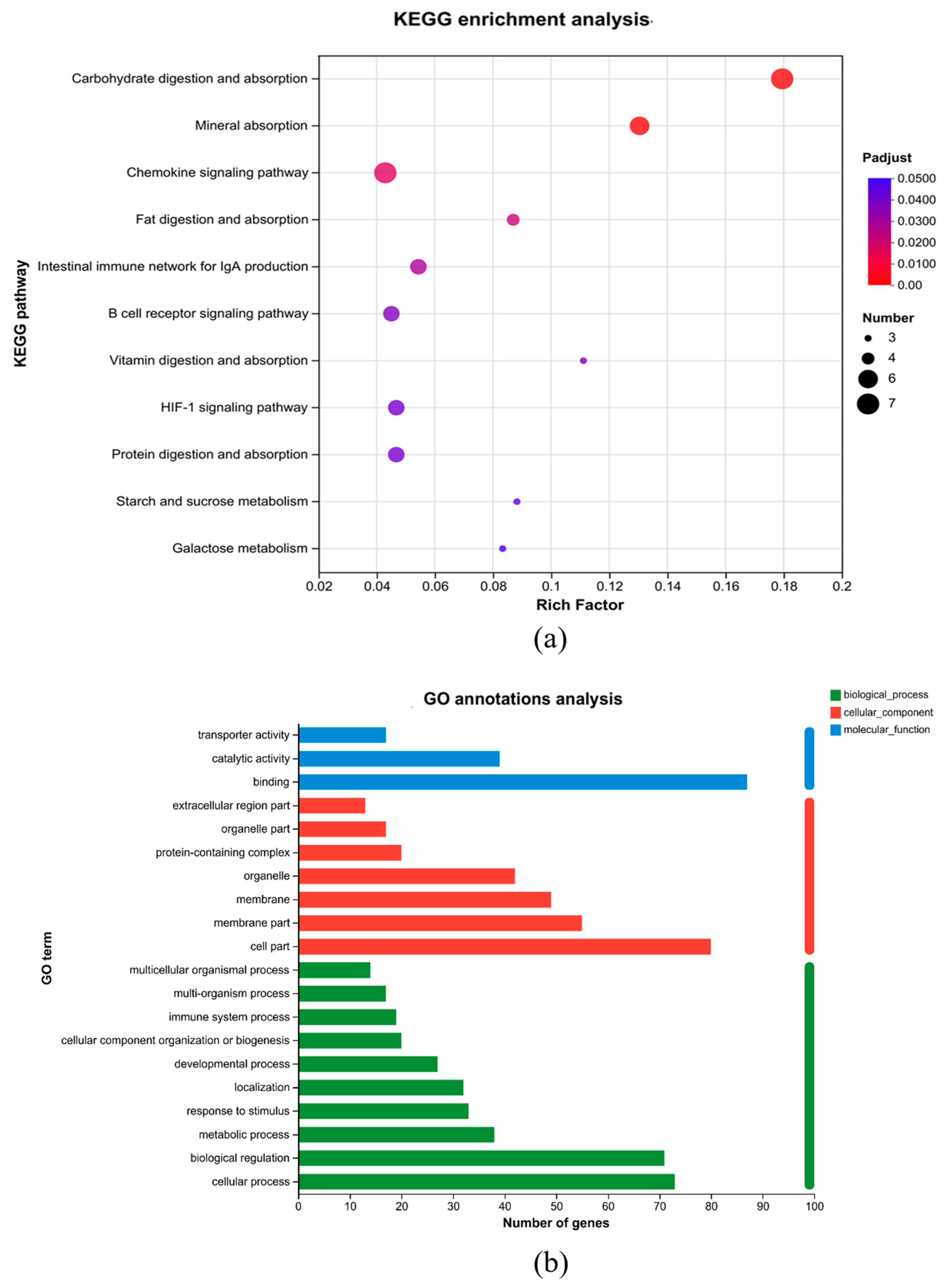
3.3. Gut Microbiota
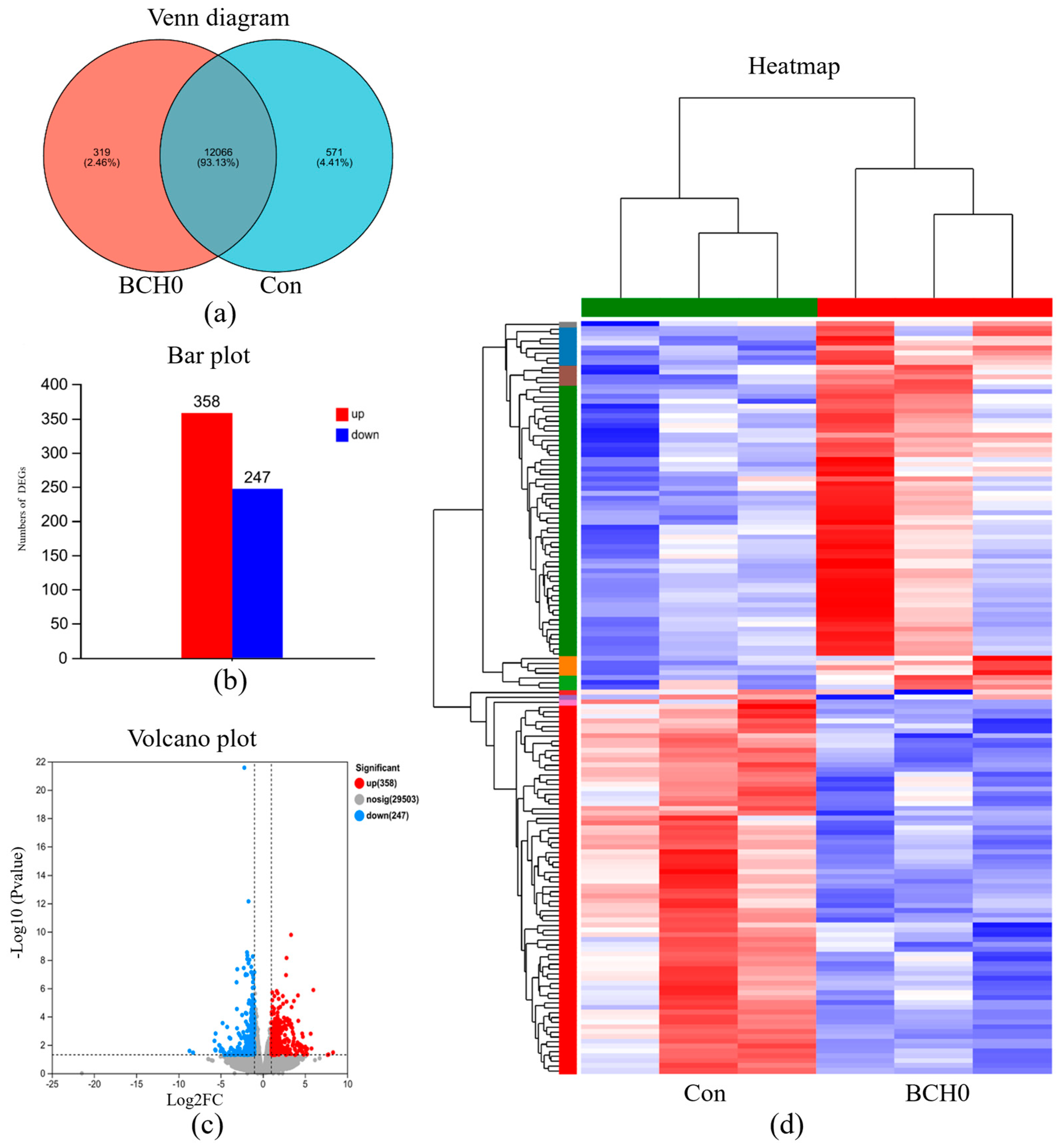
3.4. Intestinal Tissue Gene Transcription
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, V.K.; Rajendraprasad, S.; Ozkan, M.; Ramachandran, D.; Ahmad, S.; Bakken, J.S.; Laudanski, K.; Gajic, O.; Bauer, B.; Zec, S.; et al. Safety, Feasibility, and Impact on the Gut Microbiome of Kefir Administration in Critically Ill Adults. BMC Med. 2024, 22, 80. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Ma, X.K.; Tu, C.K.; Teng, H.; Shao, X.; Chen, J.; Hu, M.X. Lactobacillus reuteri Biofilms Formed on Porous Zein/Cellulose Scaffolds: Synbiotics to Regulate Intestinal Microbiota. Int. J. Biol. Macromol. 2024, 262 Pt 2, 130152. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, M.; Shaheen, M.; Sivakumar, A.; Furqan, A.; Kirankumar, B. Comparative Evaluation for Thermostability and Gastrointestinal Survival of Probiotic Bacillus coagulans MTCC 5856. Biosci. Biotechnol. Biochem. 2021, 85, 962–971. [Google Scholar]
- Galvão, A.M.; Rodrigues, S.; Fernandes, F.A. Probiotic Dried Apple Snacks: Development of Probiotic Coating and Shelf-Life Studies. J. Food Process. Preserv. 2020, 44, e14974. [Google Scholar] [CrossRef]
- Ma, N.; Chen, X.; Johnston, L.J.; Ma, X. Gut Microbiota-Stem Cell Niche Crosstalk: A New Territory for Maintaining Intestinal Homeostasis. iMeta 2022, 1, e54. [Google Scholar] [CrossRef]
- Song, B.; Li, P.; Xu, H.; Wang, Z.; Yuan, J.; Zhang, B.; Lv, Z.; Song, Z.; Guo, Y. Effects of Rearing System and Antibiotic Treatment on Immune Function, Gut Microbiota and Metabolites of Broiler Chickens. J. Anim. Sci. Biotechnol. 2023, 14, 827–852. [Google Scholar] [CrossRef]
- Lavrentev, F.V.; Ashikhmina, M.S.; Ulasevich, S.A.; Morozova, O.V.; Orlova, O.Y.; Skorb, E.V.; Iakovchenko, N.V. Perspectives of Bacillus coagulans Mtcc 5856 in the Production of Fermented Dairy Products. LWT 2021, 148, 111623. [Google Scholar] [CrossRef]
- Zhuang, X.; Stephanie, C.; Nuria, A. Bigels-Oleocolloid Matrices-as Probiotic Protective Systems in Yogurt. J. Food Sci. 2021, 86, 4892–4900. [Google Scholar] [CrossRef]
- Roobab, U.; Batool, Z.; Manzoor, M.F.; Shabbir, M.A.; Khan, M.R.; Aadil, R.M. Sources, Formulations, Advanced Delivery and Health Benefits of Probiotics. Curr. Opin. Food Sci. 2020, 32, 17–28. [Google Scholar] [CrossRef]
- Gözde, K.A.; Zerrin, E. Identification and Characterization of Bacillus coagulans Strains for Probiotic Activity and Safety. LWT 2021, 151, 112233. [Google Scholar]
- Sui, L.; Zhu, X.; Wu, D.; Ma, T.; Tuo, Y.; Jiang, S.; Qian, F.; Mu, G. In Vitro Assessment of Probiotic and Functional Properties of Bacillus coagulans T242. Food Biosci. 2020, 36, 100675. [Google Scholar] [CrossRef]
- Aulitto, M.; Alfano, A.; Maresca, E.; Avolio, R.; Errico, M.E.; Gentile, G.; Cozzolino, F.; Monti, M.; Pirozzi, A.; Donsì, F.; et al. Thermophilic Biocatalysts for One-Step Conversion of Citrus Waste into Lactic Acid. Appl. Microbiol. Biotechnol. 2024, 108, 155. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.P.; Liu, D.M.; Zhao, S.; Huang, Y.Y.; Yu, J.J.; Zhou, Q.Y. Assessing the Safety and Probiotic Characteristics of Bacillus coagulans 13002 Based on Complete Genome and Phenotype Analysis. LWT 2022, 155, 112847. [Google Scholar] [CrossRef]
- Tan, P.; Wu, C.; Tang, Q.; Wang, T.; Zhou, C.; Ding, Y.; Fu, H.; Xu, S.; Feng, Y.; Zhang, Y.; et al. pH-Triggered Size-Transformable and Bioactivity-Switchable Self-Assembling Chimeric Peptide Nano-Assemblies for Combating Drug-Resistant Bacteria and Biofilms. Adv. Mater. 2023, 35, e2210766. [Google Scholar] [CrossRef]
- Tan, P.; Sun, Z.; Tang, Q.; Xu, S.; Wang, T.; Ding, Y.; Fu, H.; Zhou, C.; Zhang, Y.; Yue, Z.; et al. Manipulation of Hydrophobic Motifs and Optimization of Sequence Patterns to Design High Stability Peptides against Piglet Bacterial Infections. Nano Today 2023, 49, 101793. [Google Scholar] [CrossRef]
- Liu, C.; Ma, N.; Feng, Y.; Zhou, M.; Li, H.; Zhang, X.; Ma, X. From Probiotics to Postbiotics: Concepts and Applications. Anim. Res. One Health 2023, 1, 92–114. [Google Scholar] [CrossRef]
- Li, S.; Ke, W.; Zhang, Q.; Undersander, D.; Zhang, G. Effects of Bacillus coagulans and Lactobacillus plantarum on the Fermentation Quality, Aerobic Stability and Microbial Community of Triticale Silage. Chem. Biol. Technol. Agric. 2023, 10, 79. [Google Scholar] [CrossRef]
- Sudha, M.R.; Jayanthi, N.; Aasin, M.; Dhanashri, R.D.; Anirudh, T. Efficacy of Bacillus coagulans Unique Is2 in Treatment of Irritable Bowel Syndrome in Children: A Double Blind, Randomised Placebo Controlled Study. Benef. Microbes 2018, 9, 563–572. [Google Scholar] [CrossRef]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Randomized Clinical Trial: The Effect of Probiotic Bacillus coagulans Unique Is2 vs. Placebo on the Symptoms Management of Irritable Bowel Syndrome in Adults. Sci. Rep. 2019, 9, 12210. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, M.; Cui, X.; Chen, J.; Liu, C.; Zhang, X. Bacillus coagulans MZY531 Alleviates Intestinal Mucosal Injury in Immunosuppressive Mice Via Modulating Intestinal Barrier, Inflammatory Response, and Gut Microbiota. Sci. Rep. 2023, 13, 11181. [Google Scholar] [CrossRef]
- Zheng, W.; Zhao, Z.; Yang, Y.; Ding, L.; Yao, W. The Synbiotic Mixture of Lactulose and Bacillus coagulans Protects Intestinal Barrier Dysfunction and Apoptosis in Weaned Piglets Challenged with Lipopolysaccharide. J. Anim. Sci. Biotechnol. 2023, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Laura, F.; Wilfried, V.; Jürgen, Z.; Ronald, G.; EvaMaria, S. The Impact of Pre- and Probiotic Product Combinations on Ex Vivo Growth of Avian Pathogenic Escherichia coli and Salmonella Enteritidis. Microorganisms 2022, 10, 121. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Radebe, S.M.; Zhang, H.; Jia, J.; Xie, S.; Shi, M.; Yu, Q. Effect of Bacillus coagulans on Maintaining the Integrity Intestinal Mucosal Barrier in Broilers. Vet. Microbiol. 2022, 266, 109357. [Google Scholar] [CrossRef]
- Guo, S.; Xi, Y.; Xia, Y.; Wu, T.; Zhao, D.; Zhang, Z.; Ding, B. Dietary Lactobacillus fermentum and Bacillus coagulans Supplementation Modulates Intestinal Immunity and Microbiota of Broiler Chickens Challenged by Clostridium perfringens. Front. Vet. Sci. 2021, 8, 680742. [Google Scholar] [CrossRef]
- Sjofjan, O.; Adli, D.N.; Harahap, R.P.; Jayanegara, A.; Utama, D.T.; Seruni, A.P. The Effects of Lactic Acid Bacteria and Yeasts as Probiotics on the Growth Performance, Relative Organ Weight, Blood Parameters, and Immune Responses of Broiler: A Meta-Analysis. F1000Research 2021, 10, 183. [Google Scholar] [CrossRef] [PubMed]
- Uzabaci, E.; Yibar, A. Effects of Probiotic Supplementation on Broiler Growth Performance: A Meta-Analysis of Randomised Controlled Trials. Anim. Prod. Sci. 2023, 63, 645–651. [Google Scholar] [CrossRef]
- Ji, L.; Zhang, L.; Liu, H.; Shen, J.; Zhang, Y.; Lu, L.; Zhang, X.; Ma, X. Bacillus subtilis M6 Improves Intestinal Barrier, Antioxidant Capacity and Gut Microbial Composition in Aa Broiler. Front. Nutr. 2022, 9, 965310. [Google Scholar] [CrossRef]
- Zhang, L.; Laila, B.S.; Nadège, H.; Séverine, Z.; Sory, D.M.; Ismail, F. Effects of Drinking Water Supplementation with Lactobacillus reuteri, and a Mixture of Reuterin and Microcin J25 on the Growth Performance, Caecal Microbiota and Selected Metabolites of Broiler Chickens. J. Anim. Sci. Biotechnol. 2022, 13, 34. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Yu, Y.; Zhang, R.; Wu, Y.; Yue, M.; Yang, C. Effects of Bacillus coagulans on Growth Performance, Antioxidant Capacity, Immunity Function, and Gut Health in Broilers. Poult. Sci. 2021, 100, 101168. [Google Scholar] [CrossRef]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans Supplementation on the Growth Performance and Gut Health of Broiler Chickens with Clostridium perfringens-Induced Necrotic Enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Criado-Mesas, L.; Abdelli, N.; Noce, A.; Farré, M.; Pérez, J.F.; Solà-Oriol, D.; Martin-Venegas, R.; Forouzandeh, A.; González-Solé, F.; Folch, J.M. Transversal Gene Expression Panel to Evaluate Intestinal Health in Broiler Chickens in Different Challenging Conditions. Sci. Rep. 2021, 11, 6315. [Google Scholar] [CrossRef]
- Mikako, S.; Tatsuyuki, T.; Takaharu, K.; Junji, S.; Yohei, K. Development of Active Jejunal Glucose Absorption in Broiler Chickens. Poult. Sci. 2023, 102, 102804. [Google Scholar]
- Wang, X.; Jian, H.; Zhao, W.; Li, J.; Zou, X.; Dong, X. Effects of Dietary Bacillus coagulans on the Productive Performance, Egg Quality, Serum Parameters, and Intestinal Morphology of Laying Hens During the Late Laying Period. Ital. J. Anim. Sci. 2023, 22, 95–105. [Google Scholar] [CrossRef]
- Zi, Q.; Zhu, S.; Li, P.; Liao, Y.; Chen, D.; He, C.; Guo, S.; Zou, X. Effects of Combined Bacillus coagulans and Yeast Fermentation Culture on Growth Performance, Plasma Biochemical Indices, Intestinal Morphology and Microbial of Broilers. J. Anim. Sci. 2025, skae325. [Google Scholar] [CrossRef]
- Pan, L.; Zhao, P.F.; Ma, X.K.; Shang, Q.H.; Xu, Y.T.; Long, S.F.; Wu, Y.; Yuan, F.M.; Piao, X.S. Probiotic Supplementation Protects Weaned Pigs Against Enterotoxigenic Escherichia coli K88 Challenge and Improves Performance Similar to Antibiotics. J. Anim. Sci. 2017, 95, 2627–2639. [Google Scholar]
- Fu, R.; Liang, C.; Chen, D.; Yan, H.; Tian, G.; Zheng, P.; He, J.; Yu, J.; Mao, X.; Huang, Z.; et al. Effects of Dietary Bacillus coagulans and Yeast Hydrolysate Supplementation on Growth Performance, Immune Response and Intestinal Barrier Function in Weaned Piglets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Sokale, A.O.; Menconi, A.; Mathis, G.F.; Lumpkins, B.; Sims, M.D.; Whelan, R.A.; Doranalli, K. Effect of Bacillus subtilis Dsm 32315 on the Intestinal Structural Integrity and Growth Performance of Broiler Chickens under Necrotic Enteritis Challenge. Poult. Sci. 2019, 98, 5392–5400. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, Y.; Lv, Y.; Li, P.; Yi, D.; Wang, L.; Zhao, D.; Chen, H.; Gong, J.; Hou, Y. Beneficial Impact and Molecular Mechanism of Bacillus coagulans on Piglets’ Intestine. Int. J. Mol. Sci. 2018, 19, 2084. [Google Scholar] [CrossRef]
- Fu, T.; Kalbacher, H.; Hoffmann, W. TFF1 is Differentially Expressed in Stationary and Migratory Rat Gastric Epithelial Cells (Rgm-1) after in Vitro Wounding: Influence of Tff1 Rna Interference on Cell Migration. Cell. Physiol. Biochem. 2013, 32, 997–1010. [Google Scholar] [CrossRef]
- Hung, A.T.; Lin, S.-Y.; Yang, T.-Y.; Chou, C.-K.; Liu, H.-C.; Lu, J.-J.; Wang, B.; Chen, S.-Y.; Lien, T.-F. Effects of Bacillus coagulans Atcc 7050 on Growth Performance, Intestinal Morphology, and Microflora Composition in Broiler Chickens. Anim. Prod. Sci. 2012, 52, 874–879. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Yue, Z.; Tan, P.; Sun, M.; Ji, L.; Bai, Y.; Ma, X. Wickerhamomyces Anomalus Relieves Weaning Diarrhea Via Improving Gut Microbiota and Redox Homeostasis Using a Piglet Model. Food Funct. 2022, 13, 11223–11235. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Li, M.; Qian, M.; Yang, Z.; Han, X. Co-Cultures of Lactobacillus acidophilus and Bacillus subtilis Enhance Mucosal Barrier by Modulating Gut Microbiota-Derived Short-Chain Fatty Acids. Nutrients 2022, 14, 4475. [Google Scholar] [CrossRef]
- Zhu, X.; Yang, B.; Hao, Z.; Li, H.; Cong, W.; Kang, Y. Dietary Supplementation with Weissella Cibaria C-10 and Bacillus amyloliquefaciens T-5 Enhance Immunity against Aeromonas veronii Infection in Crucian Carp (Carassiu auratus). Microb. Pathog. 2022, 167, 105559. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, S.; Chen, X.; Yang, S.; Deng, X.; Tu, M.; Tao, Y.; Xiang, W.; Rao, Y. Metabolic Profiles of Lactobacillus paraplantarum in Biofilm and Planktonic States and Investigation of Its Intestinal Modulation and Immunoregulation in Dogs. Food Funct. 2021, 12, 5317–5332. [Google Scholar] [CrossRef]
- Li, F.; Lv, B.; Zuo, J.; Nawaz, S.; Wang, Z.; Lian, L.; Yin, H.; Chen, S.; Han, X.; Wang, H. Effect of Solid-State Fermentation Products of Lactobacillus plantarum, Candida utilis, and Bacillus coagulans on Growth Performance of Broilers and Prevention of Avian Colibacillosis. Vet. Sci. 2024, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, X.; Dong, Y.; Li, R.; Shen, M.; Yi, D.; Wu, T.; Wang, L.; Zhao, D.; Hou, Y. Bacillus coagulans Prevents the Decline in Average Daily Feed Intake in Young Piglets Infected with Enterotoxigenic Escherichia coli K88 by Reducing Intestinal Injury and Regulating the Gut Microbiota. Front. Cell. Infect. Microbiol. 2023, 13, 1284166. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes Robert, J.; Moore Robert, J. Microbiota of the Chicken Gastrointestinal Tract: Influence on Health, Productivity and Disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Roto, S.M.; Rubinelli, P.M.; Ricke, S.C. An Introduction to the Avian Gut Microbiota and the Effects of Yeast-Based Prebiotic-Type Compounds as Potential Feed Additives. Front. Vet. Sci. 2015, 2, 28. [Google Scholar] [CrossRef]
- Yan, L.; Lv, Z.Z.; An, S.; Xing, K.; Wang, Z.G.; Lv, M.B.; Choct, M.; Guo, Y.M.; Zhou, G.L. Effects of Rearing System and Narasin on Growth Performance, Gastro-Intestinal Development and Gut Microbiota of Broilers. Poult. Sci. 2020, 100, 100840. [Google Scholar] [CrossRef]
- Chang, X.; Kang, M.; Shen, Y.; Yun, L.; Yang, G.; Zhu, L.; Meng, X.; Zhang, J.; Su, X. Bacillus coagulans Scc-19 Maintains Intestinal Health in Cadmium-Exposed Common Carp (Cyprinus carpio L.) by Strengthening the Gut Barriers, Relieving Oxidative Stress and Modulating the Intestinal Microflora. Ecotoxicol. Environ. Saf. 2021, 228, 112977. [Google Scholar] [CrossRef]
- Borja, S.; Susana, D.; Aitor, B.-M.; Anália, L.; Miguel, G.; Abelardo, M. Probiotics, Gut Microbiota, and Their Influence on Host Health and Disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar]
- Michelangelo, C.; Hsien, T.C.; Valentina, P.; Chih, H.P.; Claudio, M. Lactate Modulation of Immune Responses in Inflammatory Versus Tumour Microenvironments. Nat. Rev. Immunol. 2020, 21, 151–161. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content | |
|---|---|---|
| 0–21 d | 22–42 d | |
| Corn | 58.04 | 64.06 |
| Soybean meal | 32.00 | 26.43 |
| Fish meal | 2.00 | 2.00 |
| Soybean oil | 3.40 | 3.40 |
| CaHPO4 | 1.45 | 1.04 |
| Limestone | 1.30 | 1.35 |
| NaCl | 0.30 | 0.30 |
| Premix 1 | 1.00 | 1.00 |
| L-lysine HCl | 0.36 | 0.33 |
| DL-methionine | 0.15 | 0.09 |
| Total | 100 | 100 |
| Nutritional components 2 | ||
| Metabolizable energy (MJ/Kg) | 12.34 | 12.60 |
| Crude protein (%) | 20.12 | 18.02 |
| Ca (%) | 1.00 | 0.79 |
| P (%) | 0.46 | 0.34 |
| Lysine (%) | 1.14 | 0.95 |
| Methionine (%) | 0.46 | 0.34 |
| Items | Days | Con | BCH0 | SEM | p-Value |
|---|---|---|---|---|---|
| BW (g) | 1 | 46.90 | 46.55 | 0.41 | 0.5401 |
| 21 | 857.23 a | 925.06 b | 16.20 | 0.0001 | |
| 42 | 2405.96 a | 2531.02 b | 59.01 | 0.0400 | |
| ADG(g/d) | 1–21 | 38.59 a | 41.83 b | 0.78 | 0.0010 |
| 1–42 | 56.17 a | 59.15 b | 3.40 | 0.0470 | |
| F/G | 1–21 | 2.06 b | 1.83 a | 1.40 | 0.0480 |
| 1–42 | 2.24 b | 1.95 a | 0.12 | 0.0214 |
| Items | Segment | Con | BCH0 | SEM | p-Value |
|---|---|---|---|---|---|
| Duodenum | villus height (μm) | 1420.81 a | 1660.18 b | 35.25 | 0.0001 |
| crypt depth (μm) | 273.99 | 256.91 | 29.92 | 0.4641 | |
| V/C | 5.18 b | 6.49 a | 0.19 | 0.0001 | |
| Ileum | villus height (μm) | 750.44 a | 959.64 b | 27.85 | 0.0001 |
| crypt depth (μm) | 157.27 a | 173.69 b | 5.84 | 0.0100 | |
| V/C | 4.81 a | 5.53 b | 0.24 | 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Z.; Ji, L.; Zhang, Y.; Chen, Z.; Shen, J.; Men, Z.; Zhou, C.; Tan, P.; Ma, X. Effects of Adding Bacillus coagulans BCH0 to the Diet on Growth Performance, Tissue Structures, and Gut Microbiota in Broilers. Animals 2025, 15, 1243. https://doi.org/10.3390/ani15091243
Niu Z, Ji L, Zhang Y, Chen Z, Shen J, Men Z, Zhou C, Tan P, Ma X. Effects of Adding Bacillus coagulans BCH0 to the Diet on Growth Performance, Tissue Structures, and Gut Microbiota in Broilers. Animals. 2025; 15(9):1243. https://doi.org/10.3390/ani15091243
Chicago/Turabian StyleNiu, Zhili, Linbao Ji, Yucheng Zhang, Zeyi Chen, Jiakun Shen, Zhaoyue Men, Chenlong Zhou, Peng Tan, and Xi Ma. 2025. "Effects of Adding Bacillus coagulans BCH0 to the Diet on Growth Performance, Tissue Structures, and Gut Microbiota in Broilers" Animals 15, no. 9: 1243. https://doi.org/10.3390/ani15091243
APA StyleNiu, Z., Ji, L., Zhang, Y., Chen, Z., Shen, J., Men, Z., Zhou, C., Tan, P., & Ma, X. (2025). Effects of Adding Bacillus coagulans BCH0 to the Diet on Growth Performance, Tissue Structures, and Gut Microbiota in Broilers. Animals, 15(9), 1243. https://doi.org/10.3390/ani15091243