1. Introduction
Yeast culture (YC) is a natural fermentation product produced by
Saccharomyces cerevisiae in anaerobic environments, and it has been widely used as an additive in ruminants [
1,
2,
3]. It has been found that YC plays a beneficial role in the growth performance of beef cattle, which may be attributed to the improvement of dietary neutral detergent fiber (NDF) and crude protein (CP) digestibility [
4]. Moreover, Chen et al. [
5] found that YC can improve the serum total antioxidant capacity (T-AOC) and catalase and glutathione peroxidase (GSH-Px) concentrations of lambs. Chen et al. [
5] and Li et al. [
6] found that the supplementation of YC to ruminants increased the levels of immunoglobulin A (IgA), immunoglobulin G (IgG), immunoglobulin M (IgM) and interferon-γ (IFN-γ) in blood, which means that YC can enhance the antioxidant capacity and immunity of ruminants. However, Zhang et al. [
7] found that the supplementation of YC had no effects on the immunity of cattle, which may be caused by the differences in effective components of different YC. Yeast culture includes the cells and metabolites of
Saccharomyces cerevisiae and culture medium components, including organic acids, mannan, β-glucan, active peptides, nucleotides, vitamins, oligosaccharides, etc. [
1], which may contribute to the various biological functions of YC. However, it is not clear how the specific metabolites in YC affect the growth performance and health of animals.
The enteric methane (CH
4) of ruminants is an important anthropogenic greenhouse gas emission source globally, which is accompanied by a large amount of energy loss of ruminants, which reduces the breeding efficiency [
8,
9]. Hydrogen (H
2) is produced during carbohydrate fermentation and is mainly utilized by methanogens in the dissolved form to produce CH
4 in the rumen [
10]. The remaining H
2 can be reduced to other reduction products, such as microbial protein (MCP) and propionate, or emitted in gaseous form of [
10,
11]. The amount of H
2 production is usually determined by the volatile fatty acids (VFA) production pattern in the rumen, and the formation of acetate and propionate leads to its production and consumption, respectively [
9]. Although previous studies have found that [
12,
13] active dried
Saccharomyces cerevisiae did not reduce CH
4 emissions in ruminants, the potential and mechanism of CH
4 emission reduction of YC need to be further explored. The supplementation of YC usually unalters the fermentation characteristics of ruminal VFA in ruminants, such as the molar percentages of propionate, acetate and butyrate [
2,
14], which means that YC supplementation may not reduce enteric CH
4 production by reducing the H
2 production. Methanogenic archaea dominate the synthesis of CH
4 in the rumen, and inhibiting its activity is also beneficial to CH
4 emission reduction [
15]. Therefore, it is necessary to further explore the CH
4 inhibition effects of YC by combining the VFA profile and the relative abundance of methanogenic archaea in the rumen.
Herein, our purpose is to explore the effects of YC on growth performance and health of beef cattle, and its potential to inhibit methanogenesis in the rumen. Thus, we analyzed the nutrient and metabolite components of two different YC, Diamond V XP (XP) and Keliben (KLB), and explored the effects of YC on the growth performance, dietary nutrients digestibility, ruminal fermentation characteristics and dissolved gases, ruminal bacterial composition and antioxidant and immune capacities of beef cattle.
2. Materials and Methods
2.1. Sources of YC
The samples of XP and KLB were purchased from Diamond V Bio-fermentation Technology Co., Ltd., Shenzhen, China and Angel yeast Co., Ltd., Yichang, China, respectively, and were stored in sealed bags at room temperature.
2.2. Animals, Experimental Design and Feeding
A total of 36 finishing male Simmental beef cattle with similar age (18.3 ± 0.70 months) and weight (460 ± 17.4 kg) were randomly divided into 3 groups: CON (the basal diet;
Table 1), XP (the basal diet supplemented with 50 g/day per cattle of XP) and KLB (the basal diet supplemented with 50 g/day per cattle of KLB), with 12 cattle in each group. The experiment was conducted on a beef cattle farm of Xindalinmu Science and Technology Development Co., Ltd., Yuanjiang, Yiyang, Hunan, China (29°07′ N, 112°34′ E). Cattle were kept in separate pens, fed with total mixed rations (TMR; mixed by a horizontal mixer) at 07:00 and 16:00 every day with an independent feeder, and were free to drink water. Yeast culture was evenly top-dressed on the TMR diet of each cattle. The experiment lasted for 70 days (including 10 days of a dietary adaptation period).
2.3. Apparent Total-Tract Digestibility, Nutrient Components and Growth Performance
During the 11–70th days of the experimental period, the dry matter intake (DMI) was calculated by recording the feed intake and refusals daily. During the 60–64th days of the experimental period, about 200 g fecal samples were collected from the rectum at 08:00 and 17:00 and divided into two subsamples on average. One subsample was added with 10 mL 10% (v/v) dilute sulfuric acid for nitrogen fixation to determine CP, and the other subsample was used to determine other nutrient components, and then immediately stored at −20 °C. After that, the fecal samples were dried and crushed at 65 °C, and the samples of the same cattle for 5 days were mixed in equal proportions with the DMI and then stored in a sealed bag. Feed samples were randomly collected every 7 days, mixed and dried at 65 °C, and kept in sealed bags for determination. On the 34th and 70th days of the experimental period, all cattle were weighed before the morning feeding, and the average daily gain (ADG) was calculated.
The dry matter (DM; method 934.01), organic matter (OM; method 942.05) and CP (method 990.03) contents were determined according to the AOAC [
17]. The gross energy (GE) was measured by an isothermal automatic calorimeter (5E-AC8018; Changsha kaide measurement and control instruments Co., Ltd., Changsha, China). The contents of NDF and acid detergent fiber (ADF) were determined according to the method previously described by Van Soest et al. [
18], and the heat-stable amylase and sodium sulfite were added when the NDF was determined. The starch content was determined following the method previously described by Kartchner et al. [
19]. The acid-insoluble ash content of feed and feces samples was determined in accordance with the GB/T 23,742 method [
20], and the apparent total-tract digestibility of nutrients was calculated according to the method described by Van Keulen et al. [
21] with the acid-insoluble ash contents.
Mannan content in YC was analyzed according to T/QBAA 001 method [
22] by high-performance liquid chromatography (HPLC; LC1290; Agilent Inc., Santa Clara, CA, USA). The contents of free amino acids, acid-soluble protein and total organic acids in YC were analyzed according to GB/T 18,246 [
23], NY/T 3801 [
24] and GB/T 12,456 [
25] methods. The metabolites content of YC was determined by Nuohezhiyuan Technology Co., Ltd., Beijing, China. The content of metabolites was analyzed by a liquid chromatography–mass spectrometer (LC-MS; Agilent Inc., Santa Clara, CA, USA) [
26,
27]. The original data of metabolite determination are preprocessed by Compound Discoverer 3.1 data processing software. Then, the metabolites were identified by comparing the high-resolution secondary spectrogram databases of mzCloud and mzVault with the primary database of MassList. Metabolites with a coefficient of variance less than 30% in the samples were retained as the final result [
28]. The chromatographic peaks detected in the samples are integrated, in which the peak area of each characteristic peak represents the relative quantitative value of a metabolite, and the peak area quantitative result of each characteristic peak is standardized by using the total peak area. The analysis of the nutrients and metabolites components was performed in triplicate and sextuplicate, respectively.
2.4. Collection and Determination of Ruminal Contents
During the 67–70th days of the experimental period, ruminal contents were taken from the middle of the rumen at 0 and 2.5 h after feeding in the morning through a rumen cannula. About 100 mL of ruminal contents collected at the beginning were discarded to avoid saliva pollution. The pH value of ruminal contents was measured immediately by a pH meter (Starter 300; Ohaus Instruments Co., Ltd., Shanghai, China). The dissolved hydrogen (dH
2), dissolved CH
4 (dCH
4) and VFA concentrations were analyzed as described by Wang et al. [
29], by a gas chromatograph (7890A; Agilent Inc., Santa Clara, CA, USA). Briefly, 35 mL of ruminal contents were transferred to a 50 mL-plastic syringe, and the dissolved gases in ruminal contents were injected into another connected 20 mL-syringe with 10 mL of nitrogen by a shaker (Huichen Biotech Co., Ltd., Wuhan, China), and the gases were collected into a sealed bottle for determination. About 2 mL of ruminal contents were centrifuged at 12,000×
g for 10 min at 4 °C, and the supernatants (1.5 mL) were acidified with 0.15 mL of 25% (
w/
v) metaphosphoric acid and stored at −20 °C for the determination of VFA. About 5 mL of ruminal contents were immediately frozen in liquid nitrogen and then stored at −20 °C for MCP content analysis according to the method described by Zinn et al. [
30]. In addition, about 2 mL of ruminal contents were immediately frozen in liquid nitrogen and then stored at −80 C for subsequent DNA extraction.
2.5. Collection and Determination of Blood Samples
On the 65–66th days of the experimental period, blood samples were collected through the jugular vein at 2 h before feeding in the morning using a 5 mL vacuum blood collection tube. The collected blood was immediately transferred to an incubator for incubation for 30 min, then centrifuged at 3000× g for 15 min at room temperature to separate the serum samples and stored at −20 °C.
Serum metabolites were determined by Roche automatic biochemical analyzer (cobas c311; Roche Inc., Basel, Switzerland). The serum antioxidant index and immune index were determined by unitized biochemical and ELISA kits (Nanjing jiancheng bioengineering institute, Nanjing, China), respectively.
2.6. Analysis of Ruminal Microbiota Composition
Samples of ruminal contents collected at 0 h and 2.5 h after feeding were mixed, and the total microbial DNA was extracted according to Ma et al. [
31]. The 16S rDNA sequencing was carried out by Biozeron Biotechnology Co., Ltd., Shanghai, China, on the MiSeq platform (Illumina Inc., San Diego, CA, USA), and the PCR amplification process was as described by Zakrzewski et al. [
32]. The universal primer of 341F: (5′-CCTAYGGGRBGCASCAG-3′) and 806R: (5′-GGACTACNNGGGTATCTAAT-3′) was used to amplify the V3-V4 region of bacteria. The operational taxonomic units were clustered with a 98.7% similarity cutoff by UPARSE software (version 7.1;
http://drive5.com/uparse/ (accessed on 6 July 2024)), and UCHIME was used to identify and remove the chimeric sequences. The phylogenetic affiliation of each 16S rRNA gene sequence was analyzed using RDP Classifier (
http://rdp.cme.msu.edu/ (accessed on 6 July 2024)) against the 16S rRNA database of silva (SSU132) with a 70% confidence threshold [
33]. The analysis of microbiota composition was performed in twelve duplicates.
2.7. Statistical Analysis
The data were analyzed using a linear mixed model by SPSS 26.0 software (SPSS Inc., Chicago, IL, USA), with the groups (CON, XP and KLB) as a fixed effect, and the animal as a random effect. When the sampling time is included, the groups and interaction between groups and sampling times were used as the fixed effects, the animal as a random effect, and the sampling time as a repeated measurement. The data of bacterial compositions were analyzed by nonparametric test models, and the differences were judged by the Kruskal–Wallis test. The statistical significance was set at p < 0.05, and p < 0.10 is considered to have a difference trend.
4. Discussion
Yeast culture has been reported to improve the growth performance, nutrient digestibility, antioxidant capacity and immune function of beef cattle [
4,
6]. These functions of YC may be related to its rich active substances, such as organic acids, mannan, β-glucan, active peptides, nucleotides and some undiscovered growth factors [
1]. In the present study, various metabolites, such as betaine, ascorbic acid, citric acid, succinic acid, malic acid, acetophenone, oleamide and indole-3-acrylic acid were found to be enriched in YC, which may also improve the health and then promote the growth performance of livestock.
Yeast culture can improve the nutrients digestibility, which is related to the improvement of the activities of beneficial microorganisms and digestive enzymes [
34,
35]. The supplementation of XP and KLB into the diet of beef cattle resulted in a significant increase in the digestibility of NDF and ADF. Consistent with the present study, Wang et al. [
2] supplemented YC (10–40 g/d/head) in the diet of fattening sheep and improved the digestibility of NDF and ADF. In addition, supplementation of XP increased the CP digestibility of beef cattle, while KLB supplementation did not, which may be related to the change in rumen microbial community.
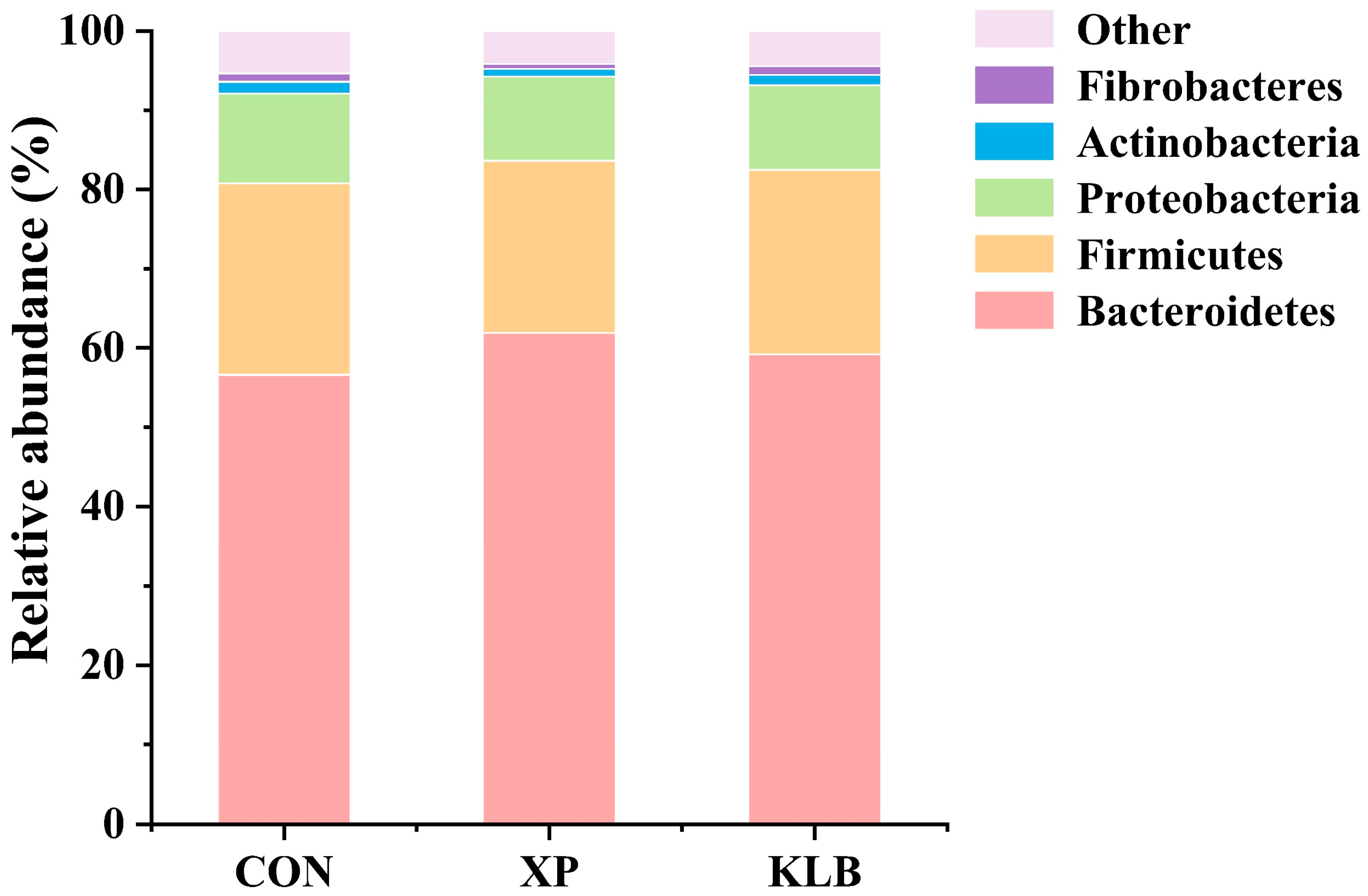
Prevotella is usually the dominant bacteria in the rumen and plays an important role in the degradation of protein [
36]. In the present study, the supplementation of XP increased the relative abundance (48.0%) of
Prevotella in the rumen compared with the CON (42.4%) and KLB (43.7%) groups, which helps uphold the improved CP digestibility. Thus, supplementing YC can promote the digestion of dietary nutrients, which may further improve the growth performance of beef cattle, as demonstrated by the increased ADG.
The total rumen VFA concentration is closely related to the digestion of dietary nutrients. However, the supplementation of XP and KLB did not increase the total VFA concentration as expected, XP even reduced the VFA concentration. One possible reason is that YC promoted the absorption of VFA by rumen epithelium [
2,
37]. We found that the supplementation of XP and KLB unaltered the molar percentages of propionate, acetate and butyrate and the acetate to propionate ratio. Our results are consistent with Wang et al. [
2], who reported that the supplementation of YC (10–40 g/d/head) to sheep did not change the rumen fermentation pathway. However, supplementing XP and KLB to beef cattle increased isobutyrate molar percentages before the morning feeding. Ruminal isobutyrate is mainly derived from the degradation of dietary amino acids, i.e., valine, isoleucine, leucine and proline [
38], and the higher contents of valine and proline in XP and KLB could contribute to the increased production of isobutyrate. Furthermore, the cattle fed with XP tended to have a higher MCP concentration compared with the CON group, which is consistent with the fact that XP supplementation enhanced amino acids availability for microbes, and the increased CP digestibility due to XP supplementation can also contribute to the enhanced MCP synthesis.
Ruminal H
2 is produced during the carbohydrates fermentation process and exists as a gaseous or dissolved form, and only the dH
2 can be utilized by the microbes. The dH
2 concentration is determined by the balance between H
2 production pathways (e.g., acetate production from carbohydrate fermentation) and consumption pathways (e.g., CH
4, propionate and MCP formation) in the rumen, and methanogenesis makes the greatest contribution to the H
2 consumption [
9,
10,
11]. Interestingly, we found that the concentration of dCH
4 in the rumen decreased after XP supplementation, which seems to indicate that methanogenesis was inhibited. Moreover, Euryarchaeota, as the main methanogenic archaea in the rumen [
39], was found to be significantly decreased after XP supplementation, which is consistent with the decreased dCH
4 concentration. However, the dH
2 concentration was not increased after methanogenesis inhibition. These results may indicate that the supplementation of XP inhibited methanogenesis, which may lead to more H
2 being redirected to other reduction products, such as the MCP synthesis, which was likely demonstrated by the increased MCP concentration. Therefore, we speculated that XP supplementation inhibited the activity of methanogens, which may lead to more H
2 being utilized to form other reduction products (e.g., MCP), rather than CH
4.
Serum metabolites are an important indicator of nutrient digestion and body metabolism [
40]. In the present study, the supplementation of XP and KLB increased serum glucose concentration, which was consistent with the higher digestibility of fiber and starch. In addition, XP and KLB supplementation had no effects on total protein, total cholesterol, blood ammonia, total cholesterol, triglyceride, LDL-C and HDL-C concentrations in serum compared with the CON group, which means that YC had no negative impacts on body metabolism. Although KLB supplementation increased the serum contents of albumin, creatinine and ALT, it was still within the normal physiological range [
41].
The animal organism continues to carry out oxidation reactions to meet the needs of normal metabolism of the substances of life. However, when the level of reactive oxygen free radicals exceeds the upper limit of degradation ability (often caused by feed, environment and social factors), it can lead to oxidative stress, endangering animal health and having a negative impact on production efficiency [
42]. Serum SOD, GSH-Px, catalase and T-AOC are important indexes to measure the antioxidant performance of animals, while serum MAD concentration can reflect the degree of oxidative damage to the animals [
42]. In the present study, the supplementation of XP increased the concentration of GSH-Px, catalase and T-AOC, and decreased the concentration of MAD, indicating that XP supplementation can enhance the antioxidant capacity of beef cattle. Acetophenone, which can improve antioxidant activity, was found to be abundant in XP rather than KLB [
43], which may contribute to the improvement of antioxidant capacity in beef cattle.
Serum IgA, IgG and IgM play a pivotal role in both the acquired response and the innate immune systems [
44], and IL-2, IL-10, IFN-γ, C3 and C4 play an important role in maintaining physiological balance and innate immunity, regulating inflammation, preventing immune hyperresponsiveness, promoting T cell proliferation and differentiation, and resisting virus invasion [
45,
46,
47,
48]. In the present study, we found that supplementation with XP increased the concentrations of IgA, IgG, IL-2, IL-10, IFN-γ and C4 in the serum compared with the CON group, while KLB supplementation increased the concentrations of IgA, IgG, IgM, IL-10, IFN-γ, C3 and C4 compared with the CON and XP groups, and increased IL-2 concentration compared with the CON group. These results indicated that YC enhances the immune response of cattle, and KLB is more effective than XP. The metabolites, such as betaine, ascorbic acid, citric acid, succinic acid and malic acid, were abundant in XP and KLB, and have the functions of improving immunity [
49,
50,
51,
52,
53], which may be the contributing factor to the enhanced immunity observed in cattle. Mannan, oleamide and indole-3-acrylic acid, abundant in KLB rather than XP, have been found to enhance the immunity in animals [
54,
55,
56], which may promote KLB to be more effective than XP in improving immunity. In addition,
Paraprevotella in the rumen has been found to be related to immune function [
57,
58], thus the greater relative abundance of ruminal
Paraprevotella by KLB supplementation can help explain the improved immunity of beef cattle.


 ,
, 



{kind=link}
{kind=link}
{kind=link}