

Upper Critical Temperature of Iberian Pigs

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
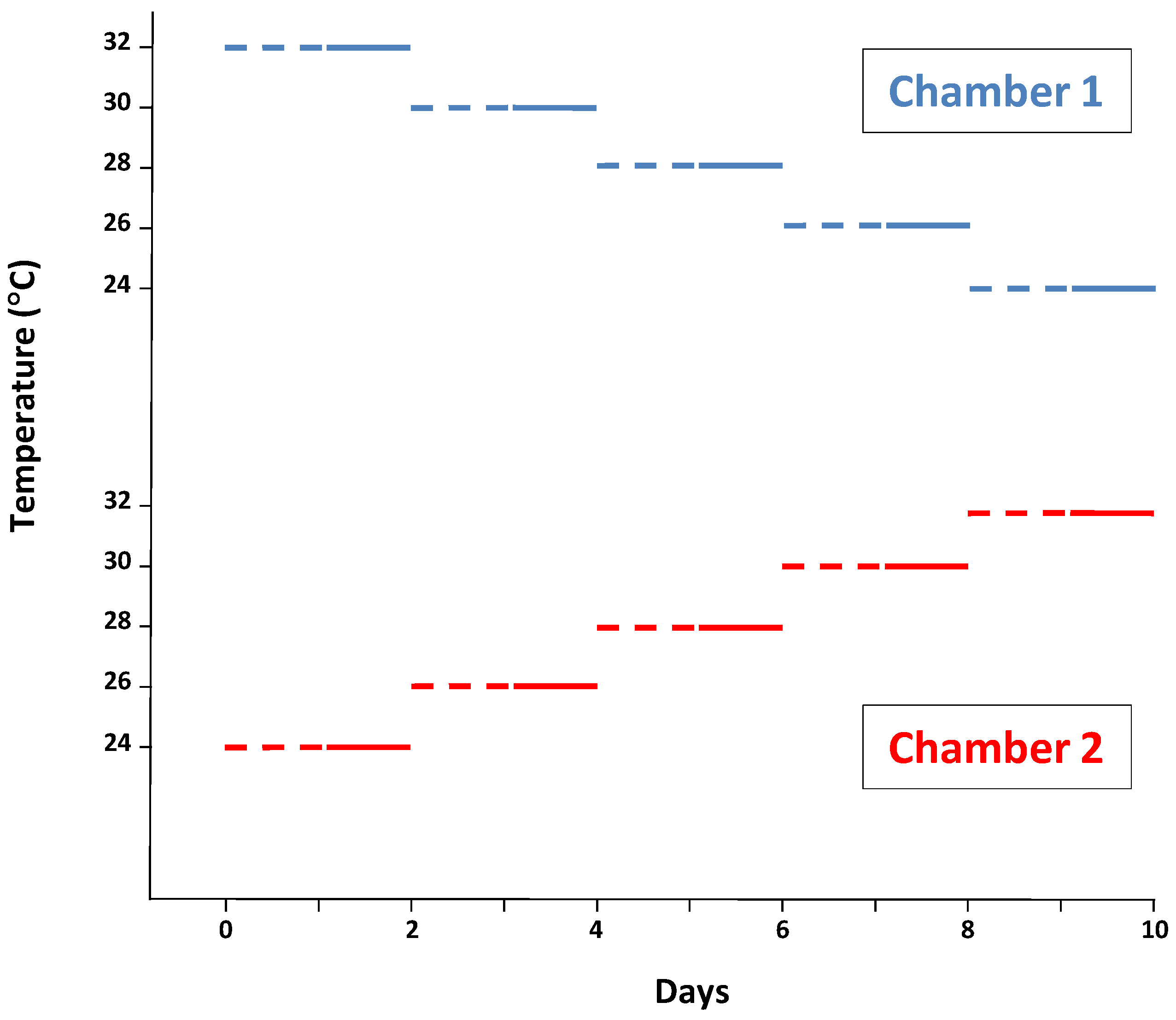
2.1. Animals and Diets
2.2. Heat Production (HP) Measurements
2.3. Physiological Measures
2.4. Chemical Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobao, M.T.; Rodrigañez, J.; Silio, L.; Toro, M.A. Iberian pig production in Spain. Pig News Inf. 1988, 9, 277–282. [Google Scholar]
- Ross, J.W.; Hale, B.J.; Gabler, N.K.; Rhoads, R.P.; Keating, A.F.; Baumgard, L.H. Physiological consequences of heat stress in pigs. Anim. Prod. Sci. 2015, 55, 1381–1390. [Google Scholar] [CrossRef]
- Pardo, Z.; Seiquer, I.; Lachica, M.; Nieto, R.; Lara, L.; Fernández-Fígares, I. Exposure of growing Iberian pigs to heat stress and effects of dietary betaine and zinc on heat tolerance. J. Therm. Biol. 2022, 106, 103230. [Google Scholar] [CrossRef]
- Renaudeau, D.; Hue, E.; Noblet, J. Acclimation to high ambient temperature in Large White and Caribbean Creole growing pigs. J. Anim. Sci. 2007, 85, 779–790. [Google Scholar] [CrossRef]
- Renaudeau, D.; Gourdine, J.L.; St-Pierre, N.R. A meta-analysis of the effects of high ambient temperature on growth performance of growing-finishing pigs. J. Anim. Sci. 2011, 89, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Sterrenburg, P.; van Ouwerkerk, E.N.J. Rekenmodel voor de Bepaling van de Thermische Behaaglijkheidszone van Varkens (BEZOVA) (Model to Determine the Thermal Comfort Zone in Pigs—BEZOVA); Rapport/Instituut voor Mechanisatie, Arbeid en Gebouwen, No. 78; I.M.A.G.: Wageningen, The Netherlands, 1986. [Google Scholar]
- Renaudeau, D.; Leclercq-Smekens, M.; Herin, M. Differences in skin characteristics in European (Large White) and Caribbean (Creole) growing pigs with reference to thermoregulation. Anim. Res. 2006, 55, 209–217. [Google Scholar] [CrossRef]
- Lopez-Bote, C.J. Sustained utilization of the Iberian pig breed. Meat Sci. 1998, 49, S17–S27. [Google Scholar] [CrossRef]
- Barea, R.; Nieto, R.; Lara, L.; García, M.A.; Vílchez, M.A.; Aguilera, J.F. Effects of dietary protein content and feeding level on carcass characteristics and organ weights of Iberian pigs growing between 50 and 100 kg live weight. Anim. Sci. 2006, 82, 405–413. [Google Scholar] [CrossRef]
- Fernández-Fígares, I.; Lachica, M.; Nieto, R.; Rivera-Ferre, M.G.; Aguilera, J. Serum profile of metabolites and hormones in obese (Iberian) and lean (Landrace) growing gilts fed balanced or lysine deficient diets. Livest. Sci. 2007, 110, 73–81. [Google Scholar] [CrossRef]
- Nieto, R.; Lara, L.; Barea, R.; García-Valverde, R.; Aguinaga, M.A.; Conde-Aguilera, J.A.; Aguilera, J.F. Response analysis of the Iberian pig growing from birth to 150 kg body weight to changes in protein and energy supply. J. Anim. Sci. 2012, 90, 3809–3820. [Google Scholar] [CrossRef]
- Bloemhof, S.; Kause, A.; Knol, E.F.; Van Arendonk, J.A.; Misztal, I. Heat stress effects on farrowing rate in sows: Genetic parameter estimation using within-line and crossbred models. J. Anim. Sci. 2012, 90, 2109–2119. [Google Scholar] [CrossRef] [PubMed]
- AAFCO. Association of American Feed Control Officials: Official Publication; Association of American Feed Control Officials: Atlanta, GA, USA, 1997. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Quiniou, N.; Noblet, J.; van Milgen, J.; Dubois, S. Modelling heat production and energy balance in group-housed growing pigs exposed to low or high ambient temperatures. Br. J. Nutr. 2001, 85, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, E. Report of Sub-Committee on Constants and Factors. In Proceedings of the 3rd Symposium on Energy Metabolism of Farm Animals, Troon, UK, May 1964; Blaxter, K.L., Ed.; Academic Press: London, UK, 1965; Volume 11, pp. 441–443. [Google Scholar]
- Rauw, W.M.; de Mercado de la Peña, E.; Gomez-Raya, L.; García-Cortés, L.A.; Ciruelos, J.J.; Gómez-Izquierdo, E. Impact of environmental temperature on production traits in pigs. Sci. Rep. 2020, 10, 2106. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.T.; Aarnink, A.J.; Verstegen, M.W.; Gerrits, W.J.; Heetkamp, M.J.; Kemp, B.; Canh, T.T. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci. 2005, 83, 1385–1396. [Google Scholar] [CrossRef]
- Lachica, M.; Pardo, Z.; Lara, L.; Nieto, R.; Fernández-Fígares, I. Heat Production of Iberian pig exposed to high temperature and effect of dietary supplementation with betaine or zinc. Animals 2024, 14, 2033. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, H.M.; Aarnink, A.J.A. Review on Heat Stress in Pigs on Farm. EU Reference Centre for Animal Welfare (Pigs). Wageningen Livestock Research, The Netherlands. 2023. Available online: https://edepot.wur.nl/587090 (accessed on 10 September 2024).
- Xin, H.; Harmon, J. Livestock Industry Facilities and Environment: Heat Stress Indices for Livestock; Iowa State University: Ames, IA, USA, 1998. [Google Scholar]
- Collin, A.; Van Milgen, J.; Dubois, S.; Noblet, J. Effect of high temperature and feeding level on energy utilization in piglets. J. Anim. Sci. 2001, 79, 1849–1857. [Google Scholar] [CrossRef]
- Hao, Y.; Feng, Y.; Yang, P.; Feng, J.; Lin, H.; Gu, X. Nutritional and physiological responses of finishing pigs exposed to a permanent heat exposure during three weeks. Arch. Anim. Nutr. 2014, 68, 296–308. [Google Scholar] [CrossRef]
- Pearce, S.C.; Gabler, N.K.; Ross, J.W.; Escobar, J.; Patience, J.F.; Rhoads, R.P.; Baumgard, L.H. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J. Anim. Sci. 2013, 91, 2108–2118. [Google Scholar] [CrossRef]
- Close, W.H. The influence of environmental temperature and plane of nutrition on heat losses from individual pigs. Anim. Sci. 1971, 13, 295–302. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L.; Yen, J.T. Thermal environment effects on growing-finishing swine Part I-Growth, feed intake and heat production. Trans. ASAE 1987, 30, 1772–1775. [Google Scholar] [CrossRef]
- Santos Ricalde, R.H.; Lean, I.J. Effect of feed intake during pregnancy on productive performance and grazing behaviour of primiparous sows kept in an outdoor system under tropical conditions. Livest. Prod. Sci. 2002, 77, 13–21. [Google Scholar] [CrossRef]
- Li, Y.; Cao, Y.; Zhou, X.; Wang, F.; Shan, T.; Li, Z.; Xu, W.; Li, C. Effects of zinc sulfate pretreatment on heat tolerance of Bama miniature pig under high ambient temperature. J. Anim. Sci. 2015, 93, 3421–3430. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K.; Schmidt-Nielsen, B.; Jarnum, S.A.; Houpt, T.R. Body temperature of the camel and its relation to water economy. Am. J. Physiol. 1956, 188, 103–112. [Google Scholar] [CrossRef]
- Collin, A.; Lebreton, Y.; Fillaut, M.; Vincent, A.; Thomas, F.; Herpin, P. Effects of exposure to high temperature and feeding level on regional blood flow and oxidative capacity of tissues in piglets. Exp. Physiol. 2001, 86, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.T.; Nienaber, D.A.H.; Pond, W.G. Oxygen consumption by portal vein-drained organs and by whole animal in consciuos growing swine. Proc. Soc. Exp. Biol. Med. 1989, 190, 2393–2395. [Google Scholar] [CrossRef]
- González-Valero, L.; Rodríguez-López, J.M.; Lachica, M.; Fernández-Fígares, I. Contribution of portal-drained viscera to heat production in Iberian gilts fed a low-protein diet: Comparison to Landrace. J. Sci. Food Agric. 2016, 96, 1202–1208. [Google Scholar] [CrossRef]
- Saxton, C. Effects of severe heat stress on respiration and metabolic rate in resting man. Aviat. Space Environ. Med. 1981, 52, 281–286. [Google Scholar]
- Sundstøl, F.; Standal, N.; Vangen, O. Energy metabolismin lines of pigs selected for thickness of backfat and rate of gain. Acta Agric. Scand. 1979, 29, 337–345. [Google Scholar] [CrossRef]
- Tess, M.W.; Dickerson, G.E.; Nienaber, J.A.; Ferrell, C.L. The effects of body composition on fasting heat production in pigs. J. Anim. Sci. 1984, 58, 99–110. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L.; Eigenberg, R.A.; Korthals, R.L.; Yen, J.T.; Harris, D.L. Genetic and heat stress interaction effects on finishing swine. In Proceedings of the 5th International Livestock Environment Symposium, Bloomington, MN, USA, 29–31 May 1997; pp. 1017–1023. [Google Scholar]
- Moreira, V.E.; Veroneze, R.; Teixeira, A.D.R.; Campos, L.D.; Lino, L.F.L.; Santos, G.A.; Silva, B.A.N.; Campos, P.H.R.F. Effects of ambient temperature on the performance and thermoregulatory responses of commercial and crossbred (Brazilian Piau purebred sires × commercial dams) growing-finishing pigs. Animals 2021, 11, 3303. [Google Scholar] [CrossRef] [PubMed]
- Brown-Brandl, T.M.; Nienaber, J.A.; Xin, H.; Gates, R.S. A literature review of swine heat production. Trans. ASAE 2004, 47, 259–270. [Google Scholar] [CrossRef]
- Le Bellego, L.; Van Milgen, J.; Noblet, J. Effect of high ambient temperature on protein and lipid deposition and energy utilization in growing pigs. Anim. Sci. 2002, 75, 85–96. [Google Scholar] [CrossRef]
- Morales, A.; Hernández, L.; Buenabad, L.; Avelar, E.; Bernal, H.; Baumgard, L.H.; Cervantes, M. Effect of heat stress on the endogenous intestinal loss of amino acids in growing pigs. J. Anim. Sci. 2016, 94, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Patience, J.F.; Umboh, J.F.; Chaplin, R.K.; Nyachoti, C.M. Nutritional and physiological responses of growing pigs exposed to a diurnal pattern of heat stress. Livest. Prod. Sci. 2005, 96, 205–214. [Google Scholar] [CrossRef]
- Lykhach, A.; Lykhach, V.; Mylostyvyi, R.; Barkar, Y.; Shpetny, M.; Izhboldina, O. Influence of housing air temperature on the behavioural acts, physiological parameters, and performance responses of fattening pigs. J. Anim. Behav. Biometeorol. 2022, 10, 2226. [Google Scholar] [CrossRef]
- Rinaldo, D.; Le Dividich, J. Assessment of optimal temperature for performance and chemical body composition of growing pigs. Livest. Prod. Sci. 1991, 29, 61–75. [Google Scholar] [CrossRef]
- Quiniou, N.; Noblet, J. Influence of high ambient temperatures on performance of multiparous lactating sows. J. Anim. Sci. 1999, 77, 2124–2134. [Google Scholar] [CrossRef]
- Campos, P.H.; Labussière, E.; Hernández-García, J.; Dubois, S.; Renaudeau, D.; Noblet, J. Effects of ambient temperature on energy and nitrogen utilization in lipopolysaccharide-challenged growing pigs. J. Anim. Sci. 2014, 92, 4909–4920. [Google Scholar] [CrossRef]
- Van der Waaij, E.H. A resource allocation model describing consequences of artificial selection under metabolic stress. J. Anim. Sci. 2004, 82, 973–981. [Google Scholar] [CrossRef]
- Bloemhof, S.; Van der Waaij, E.H.; Merks, J.W.M.; Knol, E.F. Sow line differences in heat stress tolerance expressed in reproductive performance traits. J. Anim. Sci. 2008, 86, 3330–3337. [Google Scholar] [CrossRef]
- Qu, H.; Yan, H.; Lu, H.; Donkin, S.S.; Ajuwon, K.M. Heat stress in pigs is accompanied by adipose tissue-specific responses that favor increased triglyceride storage. J. Anim. Sci. 2016, 94, 1884–1896. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Ajuwon, K.M. Adipose tissue-specific responses reveal an important role of lipogenesis during heat stress adaptation in pigs. J. Anim. Sci. 2018, 96, 975–989. [Google Scholar] [CrossRef]
- Johnson, J.S.; Sanz Fernandez, M.V.; Gutierrez, N.A.; Patience, J.F.; Ross, J.W.; Gabler, N.K.; Lucy, M.C.; Safranski, T.J.; Rhoads, R.P.; Baumgard, L.H. Effects of in utero heat stress on postnatal body composition in pigs: I. Growing phase. J. Anim. Sci. 2015, 93, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Kouba, M.; Hermier, D.; Le Dividich, J. Influence of a high ambient temperature on stearoyl-CoA-desaturase activity in the growing pig. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1999, 124, 7–13. [Google Scholar] [CrossRef]
- Vernon, R.G. Effects of diet on lipolysis and its regulation. Proc. Nutr. Soc. 1992, 51, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Sanz Fernandez, M.; Johnson, J.S.; Abuajamieh, M.; Stoakes, S.K.; Seibert, J.T.; Cox, L.; Kahl, S.; Elsasser, T.H.; Ross, J.W.; Isom, S.C.; et al. Effects of heat stress on carbohydrate and lipid metabolism in growing pigs. Physiol. Rep. 2015, 3, e12315. [Google Scholar] [CrossRef]
- Le Dividich, J.; Vermorel, M.; Noblet, J.; Bouvier, J.C.; Aumaitre, A. Effects of environmental temperature on heat production, energy retention, protein and fat gain in early-weaned piglets. Br. J. Nutr. 1980, 44, 313–323. [Google Scholar] [CrossRef]
- Oresanya, T.F.; Beaulieu, A.D.; Patience, J.F. Investigations of energy metabolism in weanling barrows: The interaction of dietary energy concentration and daily feed (energy) intake. J. Anim. Sci. 2008, 86, 348–363. [Google Scholar] [CrossRef]
- Wu, X.; Li, Z.-Y.; Jia, A.-F.; Su, H.-G.; Hu, C.-H.; Zhang, M.-H.; Feng, J.-H. Effects of high ambient temperature on lipid metabolism in finishing pigs. J. Integr. Agric. 2016, 15, 391–396. [Google Scholar] [CrossRef]
- Katsumata, M.; Yano, H.; Ishida, N.; Miyazaki, A. Influence of a high ambient temperature and administration of clenbuterol on body composition in rats. J. Nutr. Sci. Vitaminol. 1990, 36, 569–578. [Google Scholar] [CrossRef]
- Kouba, M.; Hermier, D.; Le Dividich, J. Influence of high ambient temperature on lipid metabolism in the growing pig. J. Anim. Sci. 2001, 79, 81–87. [Google Scholar] [CrossRef]
- Mujahid, A.; Furuse, M. Central administration of corticotropin-releasing factor induces thermogenesis by changes in mitochondrial bioenergetics in neonatal chicks. Neuroscience 2008, 155, 845–851. [Google Scholar] [CrossRef]
- Rhoads, R.P.; Baumgard, L.H.; Suagee, J.K. 2011 and 2012 Early Careers Achievement Awards: Metabolic priorities during heat stress with an emphasis on skeletal muscle. J. Anim. Sci. 2013, 91, 2492–2503. [Google Scholar] [CrossRef]
- Black, J.L.; Mullan, B.P.; Lorschy, M.L.; Giles, L.R. Lactation in the sow during heat stress. Livest. Prod. Sci. 1993, 35, 153–170. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Ingredients | |
|---|---|
| Barley grain | 359 |
| Corn | 150 |
| Soft wheat | 300 |
| Soybean meal | 100 |
| Beet pulp | 30 |
| Lard | 32 |
| Monocalcium phosphate | 2.0 |
| Calcium carbonate | 10 |
| Sodium chloride | 4.0 |
| L-Lysine HCl (98%) | 1.5 |
| Vitamins and minerals 1 | 12 |
| Chemical analysis | |
| Dry matter | 905 |
| Ash | 48.0 |
| Ether extract | 49.9 |
| Crude protein | 136.1 |
| Crude fiber | 36 |
| Metabolizable energy (MJ/kg) 2 | 13.1 |
| 24 °C | 26 °C | 28 °C | 30 °C | 32 °C | SEM | p-Value | |
|---|---|---|---|---|---|---|---|
| T | 23.9 a | 26.0 b | 27.9 c | 30.0 d | 31.9 e | 0.005 | <0.001 |
| RH | 75.5 a | 77.1 b | 78.0 c | 77.5 bc | 75.3 a | 0.223 | <0.001 |
| THI | 72.8 a | 76.1 b | 79.3 c | 82.4 d | 85.1 e | 0.031 | <0.001 |
| 24 °C | 26 °C | 28 °C | 30 °C | 32 °C | SEM | p-Value | |
|---|---|---|---|---|---|---|---|
| BW | 100.9 | 101.2 | 101.6 | 101.9 | 102.2 | 1.50 | 0.9763 |
| VFI | 3259.4 a | 2967.3 ab | 2783.6 ab | 2293.1 bc | 1946.2 c | 188.47 | 0.0001 |
| BR | 28.5 a | 55.0 ab | 74.0 bc | 99.1 c | 106.0 c | 8.89 | <0.0001 |
| HR | 102.6 | 99.6 | 98.0 | 95.8 | 92.6 | 3.10 | 0.2225 |
| ST | 37.2 a | 37.4 ab | 37.5 ab | 38.0 ab | 38.1 b | 0.20 | 0.0146 |
| RT | 39.6 | 39.7 | 39.6 | 39.8 | 39.8 | 0.13 | 0.6023 |
| ME intake | 1477.4 a | 1341.2 ab | 1256.3 ab | 1033.6 bc | 872.5 c | 82.42 | 0.0001 |
| HP | 743.5 a | 769.2 a | 724.5 ab | 666.3 ab | 621.4 b | 28.35 | 0.0061 |
| RE 1 | 734.0 a | 607.4 ab | 531.8 ab | 324.3 bc | 251.1 c | 68.37 | 0.0001 |
| HP/ME intake | 0.509 a | 0.554 ab | 0.592 ab | 0.662 b | 0.701 b | 0.037 | 0.0066 |
| RE/ME intake | 0.491 a | 0.446 ab | 0.408 ab | 0.338 b | 0.299 b | 0.037 | 0.0066 |
| RQ | 1.14 a | 1.13 a | 1.11 a | 1.08 ab | 1.02 b | 0.016 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachica, M.; Román, A.; Fernández-Fígares, I.; Nieto, R. Upper Critical Temperature of Iberian Pigs. Animals 2025, 15, 1374. https://doi.org/10.3390/ani15101374
Lachica M, Román A, Fernández-Fígares I, Nieto R. Upper Critical Temperature of Iberian Pigs. Animals. 2025; 15(10):1374. https://doi.org/10.3390/ani15101374
Chicago/Turabian StyleLachica, Manuel, Andreea Román, Ignacio Fernández-Fígares, and Rosa Nieto. 2025. "Upper Critical Temperature of Iberian Pigs" Animals 15, no. 10: 1374. https://doi.org/10.3390/ani15101374
APA StyleLachica, M., Román, A., Fernández-Fígares, I., & Nieto, R. (2025). Upper Critical Temperature of Iberian Pigs. Animals, 15(10), 1374. https://doi.org/10.3390/ani15101374