

Impact of the 294 bp SINE Insertion in 5′UTR of the GLYATL3 Gene on Gene Expression and Phenotypic Variation
 ,
,  , ,
, ,  ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. DNA and RNA Extraction
2.3. Verification of RIPs and Genotyping Using PCR
2.4. Expression Analysis Using qPCR
2.5. Vector Construction
2.6. Dual-Luciferase Reporter Assay
2.7. Statistical Analysis
3. Results
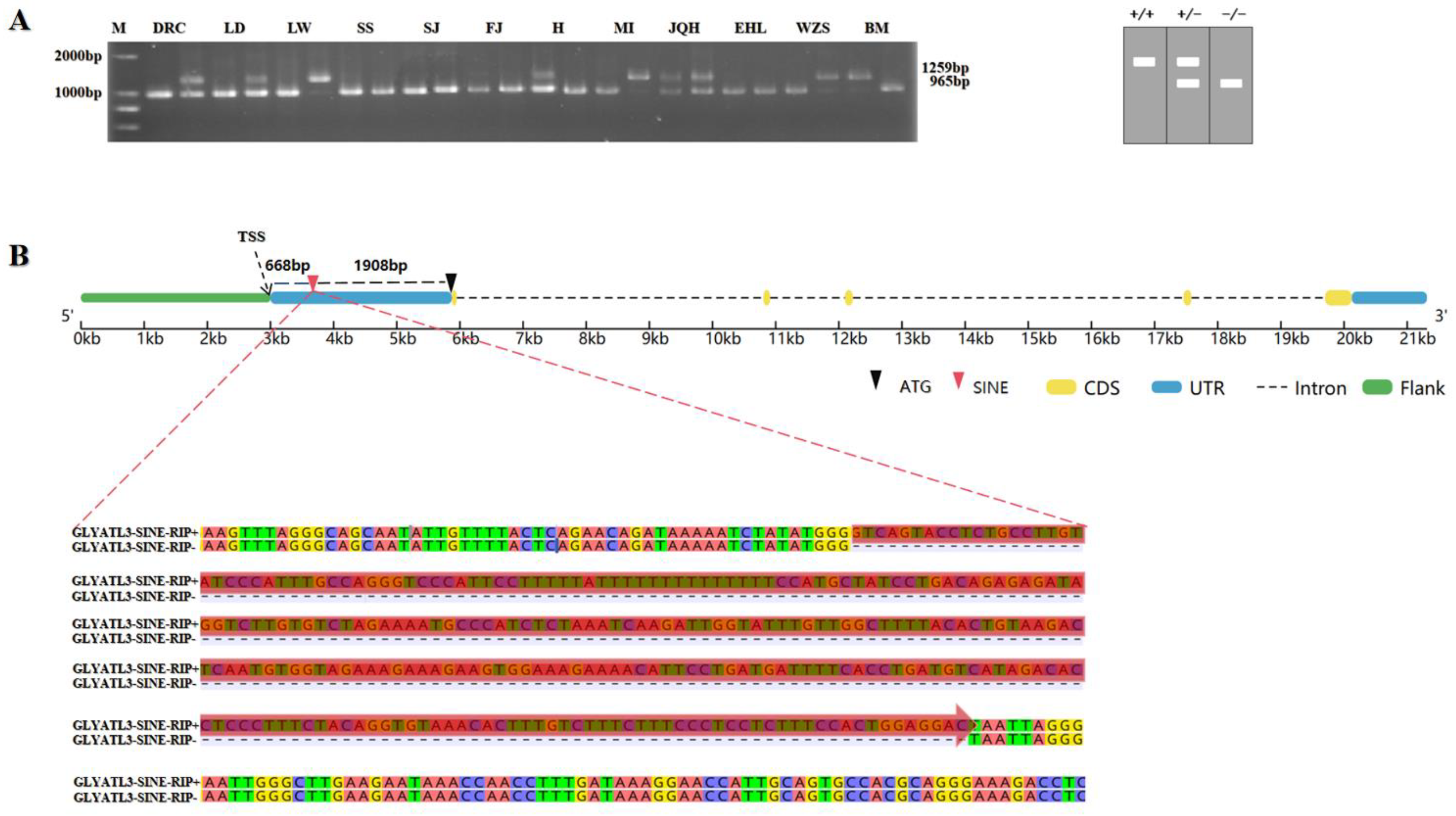
3.1. RIP Generated by SINE Insertion in Pig GLYATL3
3.2. Distribution and Genetic Diversity of GLYATL3-SINE-RIP in Different Pig Breeds
3.3. Correlation Between GLYATL3-SINE-RIP and the Phenotype of Largewhite
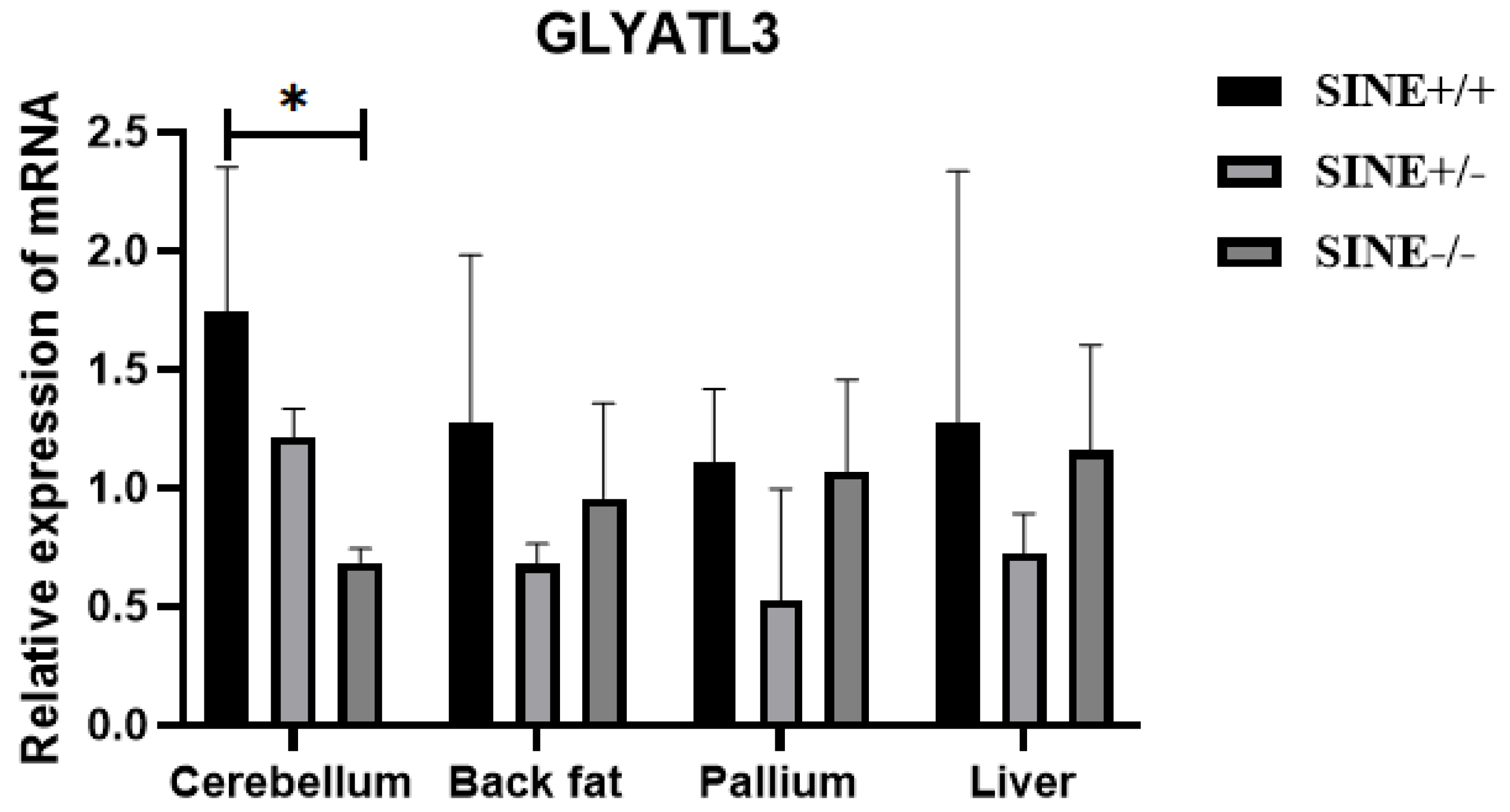
3.4. Effect of GLYATL3-SINE-RIP Insertion on GLYATL3 Expression in Mi Pigs
3.5. Validation of the Enhancer of SINE Insertion in the Pig GLYATL3 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeffries, K.A.; Dempsey, D.R.; Farrell, E.K.; Anderson, R.L.; Garbade, G.J.; Gurina, T.S.; Gruhonjic, I.; Gunderson, C.A.; Merkler, D.J. Glycine N-acyltransferase-like 3 is responsible for long-chain N-acylglycine formation in N18TG2 cells. J. Lipid Res. 2016, 57, 781–790. [Google Scholar] [CrossRef]
- Cheng, W.; Doyle, D.A.; El Arnaout, T. The N-acyltransferase Lnt: Structure-function insights from recent simultaneous studies. Int. J. Biol. Macromol. 2018, 117, 870–877. [Google Scholar] [CrossRef]
- Waluk, D.P.; Schultz, N.; Hunt, M.C. Identification of glycine N-acyltransferase-like 2 (GLYATL2) as a transferase that produces N-acyl glycines in humans. FASEB J. 2010, 24, 2795–2803. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, H.B.; Rimmerman, N.; Hu, S.S.; Burstein, S.; Walker, J.M. Novel endogenous N-acyl glycines identification and characterization. Vitam. Horm. 2009, 81, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Uehiro, N.; Sato, F.; Pu, F.; Tanaka, S.; Kawashima, M.; Kawaguchi, K.; Sugimoto, M.; Saji, S.; Toi, M. Circulating cell-free DNA-based epigenetic assay can detect early breast cancer. Breast Cancer Res. 2016, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, K.A.; Dempsey, D.R.; Behari, A.L.; Anderson, R.L.; Merkler, D.J. Drosophila melanogaster as a model system to study long-chain fatty acid amide metabolism. FEBS Lett. 2014, 588, 1596–1602. [Google Scholar] [CrossRef]
- Lu, G.; Xu, Y.; Zhang, K.; Xiong, Y.; Li, H.; Cui, L.; Wang, X.; Lou, J.; Zhai, Y.; Sun, F.; et al. Crystal structure of E. coli apolipoprotein N-acyl transferase. Nat. Commun. 2017, 8, 15948. [Google Scholar] [CrossRef]
- Kramerov, D.A.; Vassetzky, N.S. Short retroposons in eukaryotic genomes. Int. Rev. Cytol. 2005, 247, 165–221. [Google Scholar] [CrossRef]
- Kramerov, D.A.; Vassetzky, N.S. SINEs. Wiley Interdiscip. Rev. RNA 2011, 2, 772–786. [Google Scholar] [CrossRef]
- Billon, V.; Sanchez-Luque, F.J.; Rasmussen, J.; Bodea, G.O.; Gerhardt, D.J.; Gerdes, P.; Cheetham, S.W.; Schauer, S.N.; Ajjikuttira, P.; Meyer, T.J.; et al. Somatic retrotransposition in the developing rhesus macaque brain. Genome Res. 2022, 32, 1298–1314. [Google Scholar] [CrossRef]
- Kohlrausch, F.B.; Berteli, T.S.; Wang, F.; Navarro, P.A.; Keefe, D.L. Control of LINE-1 Expression Maintains Genome Integrity in Germline and Early Embryo Development. Reprod. Sci. 2022, 29, 328–340. [Google Scholar] [CrossRef]
- Faulkner, G.J. Retrotransposons: Still mobile in humans. Nat. Rev. Genet. 2022, 23, 458–459. [Google Scholar] [CrossRef] [PubMed]
- Lagisquet, J.; Zuber, K.; Gramberg, T. Recognize Yourself-Innate Sensing of Non-LTR Retrotransposons. Viruses 2021, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Göke, J.; Ng, H.H. CTRL+INSERT: Retrotransposons and their contribution to regulation and innovation of the transcriptome. EMBO Rep. 2016, 17, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, W.; Wang, X.; Shen, D.; Wang, S.; Wang, Y.; Gao, B.; Wimmers, K.; Mao, J.; Li, K.; et al. Retrotransposons evolution and impact on lncRNA and protein coding genes in pigs. Mob. DNA 2019, 10, 19. [Google Scholar] [CrossRef]
- Richardson, S.R.; Doucet, A.J.; Kopera, H.C.; Moldovan, J.B.; Garcia-Perez, J.L.; Moran, J.V. The Influence of LINE-1 and SINE Retrotransposons on Mammalian Genomes. Microbiol. Spectr. 2015, 3, Mdna3-0061-2014. [Google Scholar] [CrossRef]
- Burgess, D.J. Population genetics: Mobile elements across human populations. Nat. Rev. Genet. 2013, 14, 370. [Google Scholar] [CrossRef]
- Li, J.; Davis, B.W.; Jern, P.; Dorshorst, B.J.; Siegel, P.B.; Andersson, L. Characterization of the endogenous retrovirus insertion in CYP19A1 associated with henny feathering in chicken. Mob. DNA 2019, 10, 38. [Google Scholar] [CrossRef]
- Kratochwil, C.F.; Kautt, A.F.; Nater, A.; Härer, A.; Liang, Y.; Henning, F.; Meyer, A. An intronic transposon insertion associates with a trans-species color polymorphism in Midas cichlid fishes. Nat. Commun. 2022, 13, 296. [Google Scholar] [CrossRef]
- Chang, L.; Xueqin, R.; Xi, N.; Sheng, L.; Jiafu, W.; Qin, Z. Insertion of 275-bp SINE into first intron of PDIA4 gene is associated with litter size in Xiang pigs. Anim. Reprod. Sci. 2018, 195, 16–23. [Google Scholar] [CrossRef]
- Luca Fontanesi, E.S.L.B.S.D.O.; Vincenzo, R. Investigation of a Short Interspersed Nuclear Element Polymorphic Site in the Porcine Vertnin Gene: Allele Frequencies and Association Study With Meat Quality, Carcass and Production Traits in Italian Large White pigs. Ital. J. Anim. Sci. 2014, 13, 3090. [Google Scholar] [CrossRef]
- Chen, C.; Wang, X.; Zong, W.; D’Alessandro, E.; Giosa, D.; Guo, Y.; Mao, J.; Song, C. Genetic Diversity and Population Structures in Chinese Miniature Pigs Revealed by SINE Retrotransposon Insertion Polymorphisms, a New Type of Genetic Markers. Animals 2021, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Flavell, A.J.; Ellis, T.H.N.; Sjakste, T.; Moisy, C.; Schulman, A.H. Analysis of plant diversity with retrotransposon-based molecular markers. Heredity 2011, 106, 520–530. [Google Scholar] [CrossRef]
- Vassetzky, N.S.; Borodulina, O.R.; Ustyantsev, I.G.; Kosushkin, S.A.; Kramerov, D.A. Analysis of SINE Families B2, Dip, and Ves with Special Reference to Polyadenylation Signals and Transcription Terminators. Int. J. Mol. Sci. 2021, 22, 9897. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Peng, J.; Yang, S.; Liu, D.; Gao, S.; Zhu, Y.; Chai, Z.; Lee, B.C.; Wei, R.; Wang, J.; et al. SINE-Associated LncRNA SAWPA Regulates Porcine Zygotic Genome Activation. Adv. Sci. 2024, 11, e2307505. [Google Scholar] [CrossRef]
- Sun, D.; Zhu, Y.; Peng, W.; Zheng, S.; Weng, J.; Dong, S.; Li, J.; Chen, Q.; Ge, C.; Liao, L.; et al. SETDB1 regulates short interspersed nuclear elements and chromatin loop organization in mouse neural precursor cells. Genome Biol. 2024, 25, 175. [Google Scholar] [CrossRef]
- Langevin, M.; Synkova, H.; Jancuskova, T.; Pekova, S. Merle phenotypes in dogs—SILV SINE insertions from Mc to Mh. PLoS ONE 2018, 13, e0198536. [Google Scholar] [CrossRef]
- He, J.; Yu, M.; Chi, C.; Du, Z.; Zheng, Y.; Chen, C.; Moawad, A.S.; Song, C.; Wang, X. Insertion of 643bp Retrotransposon Upstream of PPARγ CDS Is Associated with Backfat of Large White Pigs. Animals 2023, 13, 2355. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, C.; Wang, M.; Moawad, A.S.; Wang, X.; Song, C. SINE Insertion in the Pig Carbonic Anhydrase 5B (CA5B) Gene Is Associated with Changes in Gene Expression and Phenotypic Variation. Animals 2023, 13, 1942. [Google Scholar] [CrossRef]
- Platt, R.N., 2nd; Vandewege, M.W.; Ray, D.A. Mammalian transposable elements and their impacts on genome evolution. Chromosome Res. 2018, 26, 25–43. [Google Scholar] [CrossRef]
- Gardner, E.J.; Lam, V.K.; Harris, D.N.; Chuang, N.T.; Scott, E.C.; Pittard, W.S.; Mills, R.E.; Devine, S.E. The Mobile Element Locator Tool (MELT): Population-scale mobile element discovery and biology. Genome Res. 2017, 27, 1916–1929. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Zhao, R.; Wang, X.; Zhou, C.; Wang, J.; Chen, C.; Niu, N.; Zheng, Y.; Chen, L.; Liu, X.; et al. Population genetic analysis based on the polymorphisms mediated by transposons in the genomes of pig. DNA Res. 2024, 31, dsae008. [Google Scholar] [CrossRef] [PubMed]
- Suster, M.L.; Kikuta, H.; Urasaki, A.; Asakawa, K.; Kawakami, K. Transgenesis in zebrafish with the tol2 transposon system. Methods Mol. Biol. 2009, 561, 41–63. [Google Scholar] [CrossRef]
- Serrote, C.M.L.; Reiniger, L.R.S.; Silva, K.B.; Rabaiolli, S.; Stefanel, C.M. Determining the Polymorphism Information Content of a molecular marker. Gene 2020, 726, 144175. [Google Scholar] [CrossRef] [PubMed]
- Singer, D.S.; Parent, L.J.; Ehrlich, R. Identification and DNA sequence of an interspersed repetitive DNA element in the genome of the miniature swine. Nucleic Acids Res. 1987, 15, 2780. [Google Scholar] [CrossRef]
- Wang, X.; Chi, C.; He, J.; Du, Z.; Zheng, Y.; D’Alessandro, E.; Chen, C.; Moawad, A.S.; Asare, E.; Song, C. SINE Insertion May Act as a Repressor to Affect the Expression of Pig LEPROT and Growth Traits. Genes 2022, 13, 1422. [Google Scholar] [CrossRef]
- Guo, Y.F.; Zhang, L.S.; Liu, Y.J.; Hu, H.G.; Li, J.; Tian, Q.; Yu, P.; Zhang, F.; Yang, T.L.; Guo, Y.; et al. Suggestion of GLYAT gene underlying variation of bone size and body lean mass as revealed by a bivariate genome-wide association study. Hum. Genet. 2013, 132, 189–199. [Google Scholar] [CrossRef]
- Cortes-Araya, Y.; Cheung, S.; Ho, W.; Stenhouse, C.; Ashworth, C.J.; Esteves, C.L.; Donadeu, F.X. Effects of foetal size, sex and developmental stage on adaptive transcriptional responses of skeletal muscle to intrauterine growth restriction in pigs. Sci. Rep. 2024, 14, 8500. [Google Scholar] [CrossRef]
- Dwyer, C.M.; Fletcher, J.M.; Stickland, N.C. Muscle cellularity and postnatal growth in the pig. J. Anim. Sci. 1993, 71, 3339–3343. [Google Scholar] [CrossRef]
- Reiland, S. Growth and skeletal development of the pig. Acta Radiol. Suppl. 1978, 358, 15–22. [Google Scholar]
- Robison, O.W. Growth patterns in swine. J. Anim. Sci. 1976, 42, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Estécio, M.R.; Gallegos, J.; Dekmezian, M.; Lu, Y.; Liang, S.; Issa, J.P. SINE retrotransposons cause epigenetic reprogramming of adjacent gene promoters. Mol. Cancer Res. 2012, 10, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Yeom, Y.I.; Ha, H.S.; Balling, R.; Schöler, H.R.; Artzt, K. Structure, expression and chromosomal location of the Oct-4 gene. Mech. Dev. 1991, 35, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Schöler, H.R.; Balling, R.; Hatzopoulos, A.K.; Suzuki, N.; Gruss, P. Octamer binding proteins confer transcriptional activity in early mouse embryogenesis. Embo J. 1989, 8, 2551–2557. [Google Scholar] [CrossRef]
- Dhadi, S.R.; Xu, Z.; Shaik, R.; Driscoll, K.; Ramakrishna, W. Differential regulation of genes by retrotransposons in rice promoters. Plant Mol. Biol. 2015, 87, 603–613. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| RIP | Breeds | Number | +/+ | +/− | −/− | + | − | p Value | PIC |
|---|---|---|---|---|---|---|---|---|---|
| GLYATL3-SINE-RIP | DRC | 24 | 0.25 | 0.58 | 0.17 | 0.54 | 0.46 | 0.39 | 0.37 |
| LD | 23 | 0.17 | 0.61 | 0.22 | 0.49 | 0.51 | 0.29 | 0.38 | |
| LW | 458 | 0.48 | 0.41 | 0.11 | 0.67 | 0.33 | 0.47 | 0.34 | |
| SS | 24 | 0 | 0.08 | 0.92 | 0.04 | 0.96 | 0.83 | 0.08 | |
| SJ | 24 | 0 | 0.08 | 0.92 | 0.04 | 0.96 | 0.83 | 0.08 | |
| EHL | 24 | 0 | 0.17 | 0.83 | 0.08 | 0.92 | 0.66 | 0.14 | |
| JQH | 23 | 0.48 | 0.52 | 0 | 0.74 | 0.26 | 0.10 | 0.31 | |
| MI | 29 | 0.14 | 0.69 | 0.17 | 0.49 | 0.51 | 0.04 | 0.38 | |
| MS | 24 | 0.5 | 0.29 | 0.21 | 0.64 | 0.36 | 0.08 | 0.36 | |
| MZ | 24 | 0.17 | 0.29 | 0.54 | 0.31 | 0.69 | 0.16 | 0.34 | |
| RC | 24 | 0 | 0.33 | 0.67 | 0.17 | 0.83 | 0.33 | 0.24 | |
| TB | 24 | 0.13 | 0.29 | 0.58 | 0.27 | 0.73 | 0.20 | 0.32 | |
| JH | 24 | 0.29 | 0.38 | 0.33 | 0.48 | 0.52 | 0.22 | 0.38 | |
| WZS | 23 | 0.29 | 0.17 | 0.54 | 0.35 | 0.65 | <0.01 | 0.35 | |
| BM | 24 | 0.33 | 0.25 | 0.42 | 0.46 | 0.54 | 0.02 | 0.37 |
| Genotype | Number | Body Weight Before Slaughter (kg) | Thickness of Back Fat (mm) | Age at 30 kg Body Weight (Days) | Age at 100 kg Body Weight (Days) | Back-Fat Thickness at 100 kg Body Weight (mm) |
|---|---|---|---|---|---|---|
| GLYATL3-SINE+/+ | 220 | 100.52 ± 9.11 | 10.79 ± 2.83 | 74.57 ± 7.63 a | 162.37 ± 7.94 | 10.70 ± 2.55 |
| GLYATL3-SINE+/− | 190 | 100.04 ± 8.97 | 10.83 ± 2.94 | 74.74 ± 7.25 ab | 163.19 ± 8.51 | 10.80 ± 2.71 |
| GLYATL3-SINE−/− | 48 | 99.07 ± 10.19 | 11.17 ± 3.31 | 77.39 ± 8.68 b | 163.29 ± 7.95 | 11.20 ± 2.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Qiao, S.; Zheng, Y.; Yu, M.; Chen, H.; Chen, C.; Moawad, A.S.; Gao, B.; Song, C.; Wang, X. Impact of the 294 bp SINE Insertion in 5′UTR of the GLYATL3 Gene on Gene Expression and Phenotypic Variation. Animals 2025, 15, 1375. https://doi.org/10.3390/ani15101375
Zhou C, Qiao S, Zheng Y, Yu M, Chen H, Chen C, Moawad AS, Gao B, Song C, Wang X. Impact of the 294 bp SINE Insertion in 5′UTR of the GLYATL3 Gene on Gene Expression and Phenotypic Variation. Animals. 2025; 15(10):1375. https://doi.org/10.3390/ani15101375
Chicago/Turabian StyleZhou, Chenyu, Suwei Qiao, Yao Zheng, Miao Yu, Hong Chen, Cai Chen, Ali Shoaib Moawad, Bo Gao, Chengyi Song, and Xiaoyan Wang. 2025. "Impact of the 294 bp SINE Insertion in 5′UTR of the GLYATL3 Gene on Gene Expression and Phenotypic Variation" Animals 15, no. 10: 1375. https://doi.org/10.3390/ani15101375
APA StyleZhou, C., Qiao, S., Zheng, Y., Yu, M., Chen, H., Chen, C., Moawad, A. S., Gao, B., Song, C., & Wang, X. (2025). Impact of the 294 bp SINE Insertion in 5′UTR of the GLYATL3 Gene on Gene Expression and Phenotypic Variation. Animals, 15(10), 1375. https://doi.org/10.3390/ani15101375