
Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Broilers and Sampling
2.2. Detection of Growth Performance
2.3. Detection of Antioxidant Status
2.4. Detection of GSH-Related Enzymes
2.5. Detection of Nrf2 and p38 MAPK Concentrations
2.6. Expression of Nrf2 and p38 MAPK mRNA
2.7. Statistical Analysis
3. Results
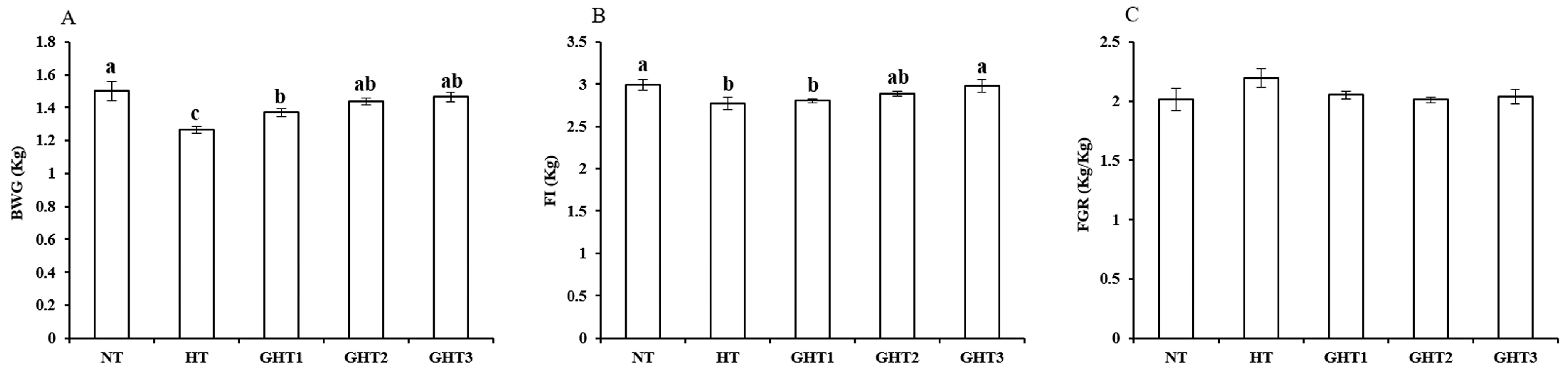
3.1. Effects of Gln Supplement on Growth Performance of Heat-Stressed Broilers
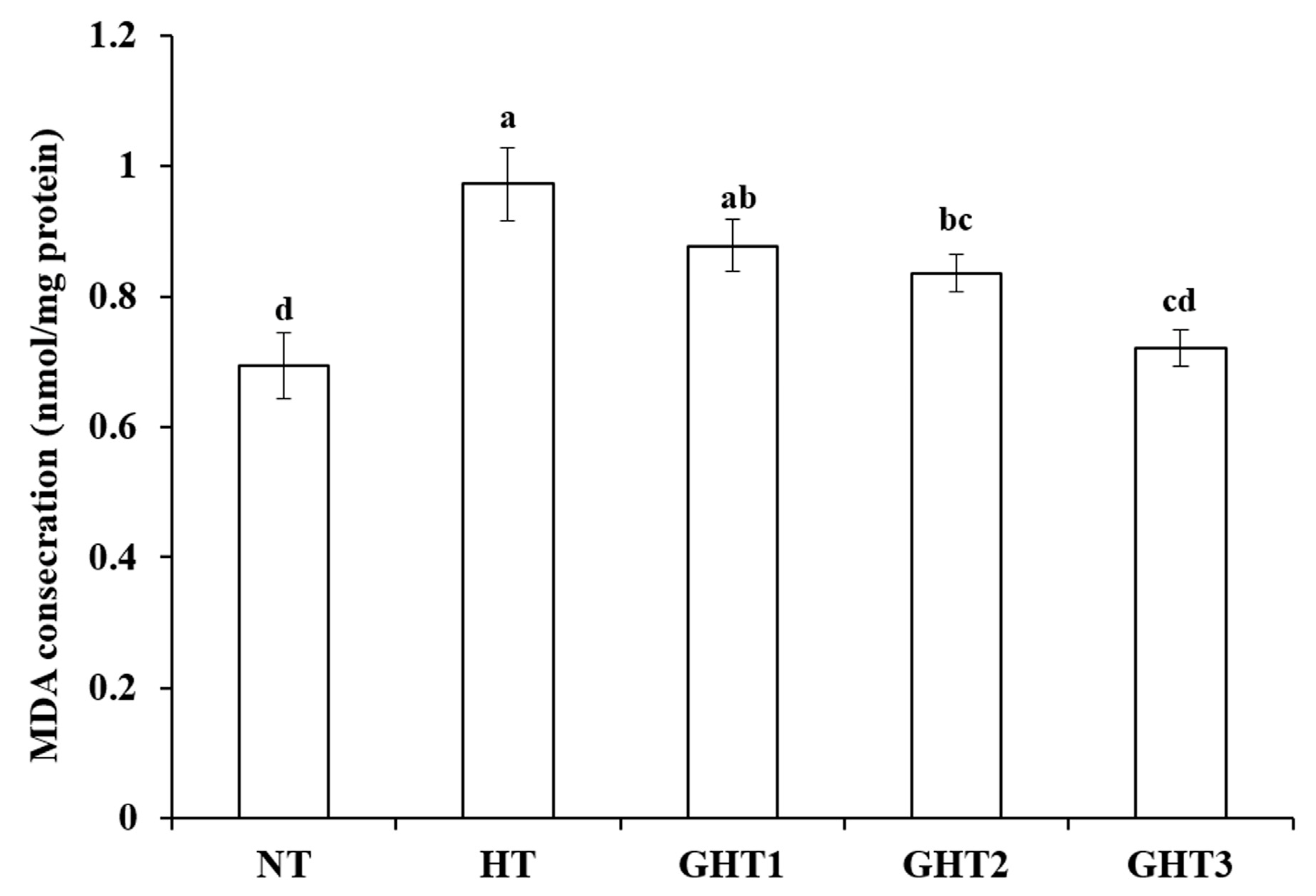
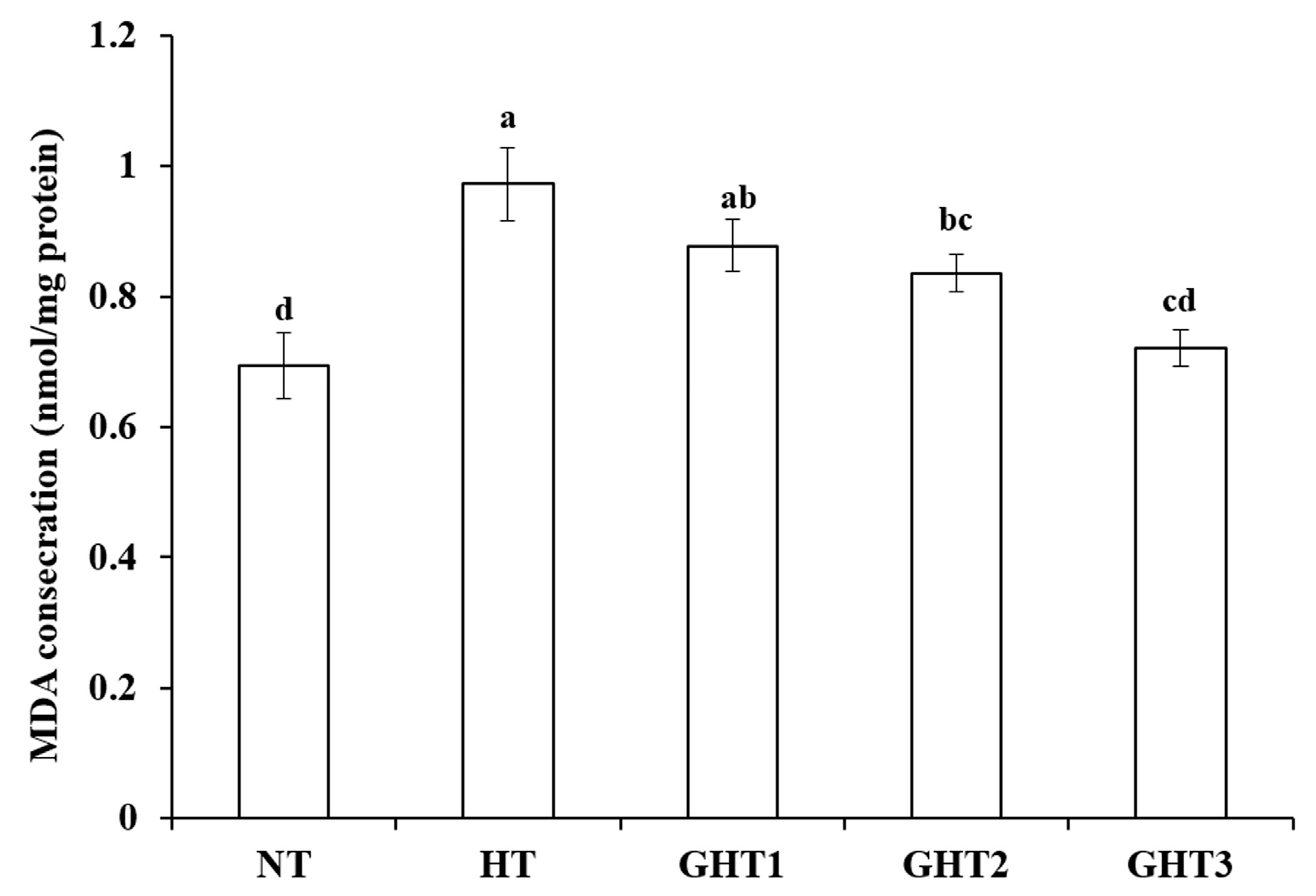
3.2. Effects of Gln Supplement on MDA on the Livers of Heat-Stressed Broilers
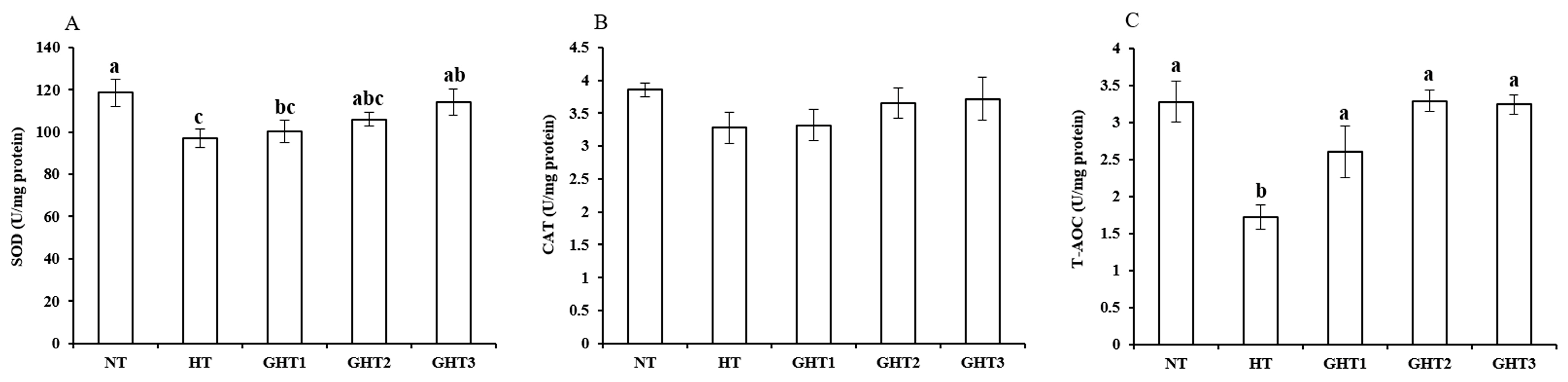
3.3. Effects of Gln Supplement on SOD, CAT, and T-AOC on the Livers of Heat-Stressed Broilers
3.4. Effects of Gln Supplement on GSH-Related Enzymes on the Livers of Heat-Stressed Broilers
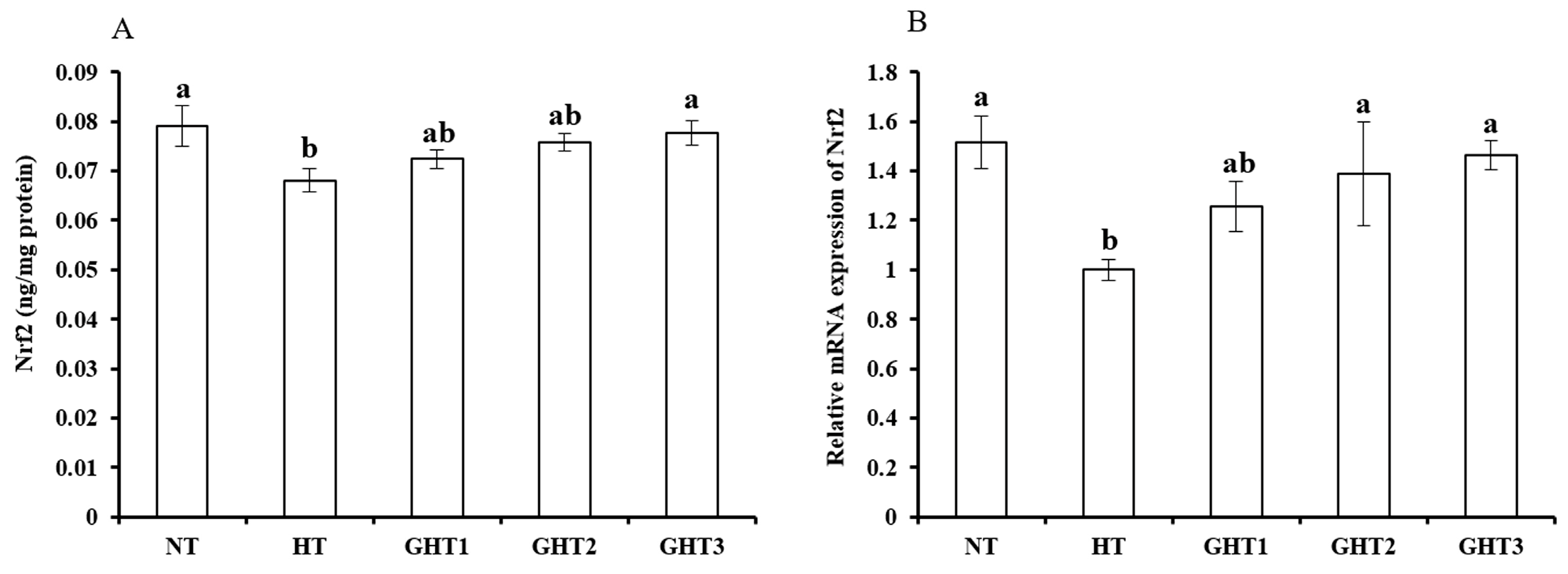
3.5. Effects of Gln Supplement on Nrf2 Protein and mRNA Expression on the Livers of Heat-Stressed Broilers
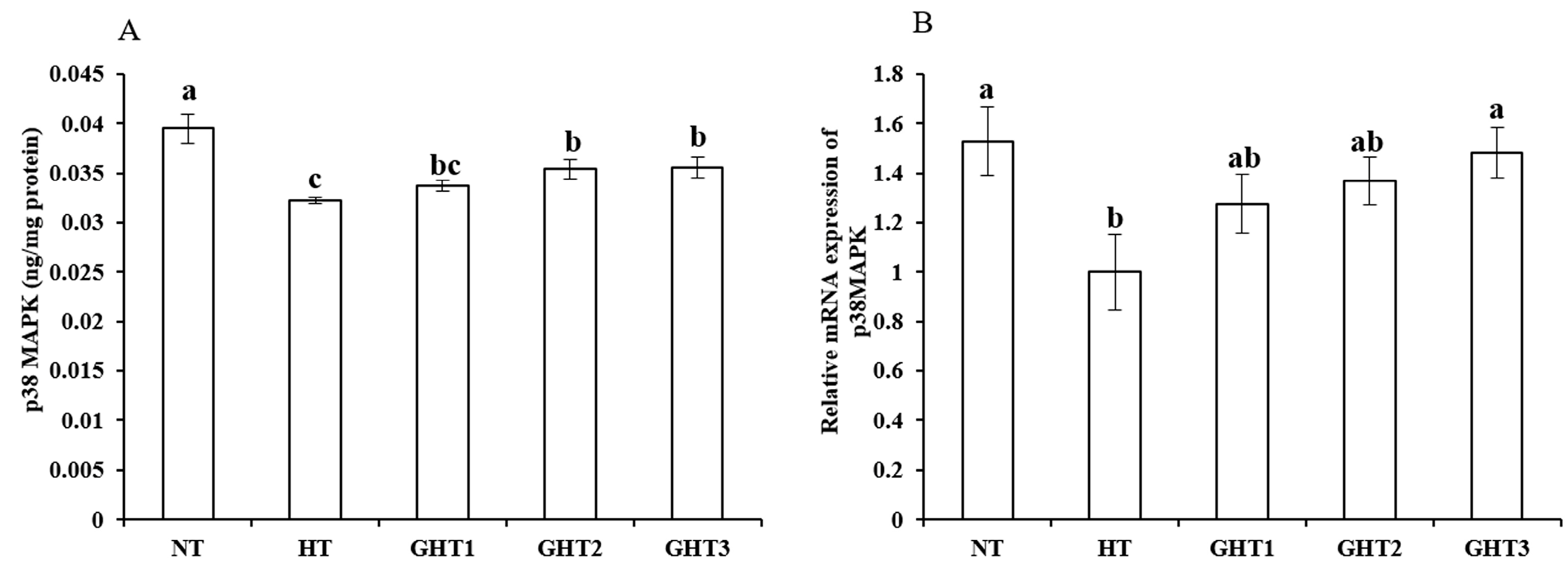
3.6. Effects of Gln Supplement on p38 MAPK Protein and mRNA Expression on the Livers of Heat-Stressed Broilers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Kikusato, M.; Xue, G.; Pastor, A.; Niewold, T.A.; Toyomizu, M. Effects of plant-derived isoquinoline alkaloids on growth performance and intestinal function of broiler chickens under heat stress. Poult. Sci. 2021, 100, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.; Li, Y.; Chang, Q.; Zhao, Z. Dietary chitosan oligosaccharides alleviate heat stress–induced intestinal oxidative stress and inflammatory response in yellow-feather broilers. Poult. Sci. 2020, 99, 6745–6752. [Google Scholar] [CrossRef]
- Ghanima, M.M.A.; Abd El-Hack, M.E.; Othman, S.I.; Taha, A.E.; Allam, A.A.; Abdel-Moneim, A.M.E. Impact of different rearing systems on growth, carcass traits, oxidative stress biomarkers, and humoral immunity of broilers exposed to heat stress. Poult. Sci. 2020, 99, 3070–3078. [Google Scholar] [CrossRef] [PubMed]
- Coqueiro, A.Y.; Rogero, M.M.; Tirapegui, J. Glutamine as an anti-fatigue amino acid in sports nutrition. Nutrients 2019, 1, 863. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liao, S.F. Physiological effects of dietary amino acids on gut health and functions of swine. Front. Vet. Sci. 2019, 6, 169. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.N.; Lopreiato, V.; Alharthi, A.; Loor, J.J. Amino acids and the regulation of oxidative stress and immune function in dairy cattle. J. Anim. Sci. 2020, 98, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, C.; Liao, J.; Hu, N.; Cheng, B.; Ma, Y.; Wang, Y. Effects of Dietary supplementation with glutamine on the immunity and intestinal barrier gene expression in broiler chickens infected with Salmonella enteritidis. Animals 2022, 12, 2168. [Google Scholar] [CrossRef]
- Liu, É.R.; de Freitas Dresch, D.; Carvalho, C.; Medeiros, P.C.; Rosa, T.G.; de Oliveira, C.M.; Martins, L.; Schemitt, E.; Bona, S.; Guma, F.C.R.; et al. Oral glutamine supplementation attenuates inflammation and oxidative stress-mediated skeletal muscle protein content degradation in immobilized rats: Role of 70 kDa heat shock protein. Free Radical Bio. Med. 2019, 145, 87–102. [Google Scholar]
- Cao, Y.; Feng, Z.; Hoos, A.; Klimberg, V.S. Glutamine enhances gut glutathione production. J. Parenter. Enteral Nutr. 1998, 22, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ren, M.; Ren, K.; Jin, Y.; Yan, M. Heat stress impacts on broiler performance: A systematic review and meta-analysis. Poult. Sci. 2020, 99, 6205–6211. [Google Scholar] [CrossRef] [PubMed]
- Alhenaky, A.; Abdelqader, A.; Abuajamieh, M.; Al-Fataftah, A.R. The effect of heat stress on intestinal integrity and Salmonella invasion in broiler birds. J. Therm. Biol. 2017, 70, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Lim, B.; Kim, J.M.; Kil, D.Y. Integrated transcriptome analysis for the hepatic and jejunal mucosa tissues of broiler chickens raised under heat stress conditions. J. Anim. Sci. Biotechnol. 2022, 13, 1–17. [Google Scholar] [CrossRef]
- Habashy, W.S.; Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol. 2019, 63, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Del Vesco, A.P.; Khatlab, A.S.; Goes, E.S.R.; Utsunomiya, K.S.; Vieira, J.S.; Neto, A.O.; Gasparino, E. Age-related oxidative stress and antioxidant capacity in heat-stressed broilers. Animal 2017, 11, 1783–1790. [Google Scholar] [CrossRef]
- Olubodun, J.O.; Zulkifli, I.; Farjam, A.S.; Hair-Bejo, M.; Kasim, A. Glutamine and glutamic acid supplementation enhances performance of broiler chickens under the hot and humid tropical condition. Ital. J. Anim. Sci. 2015, 14, 3263. [Google Scholar] [CrossRef]
- Shakeri, M.; Zulkifli, I.; Soleimani, A.F.; o’Reilly, E.L.; Eckersall, P.D.; Anna, A.A.; Kumari, S.; Abdullah, F.F.J. Response to dietary supplementation of L-glutamine and L-glutamate in broiler chickens reared at different stocking densities under hot, humid tropical conditions. Poult. Sci. 2014, 93, 2700–2708. [Google Scholar] [CrossRef]
- Dai, S.F.; Gao, F.; Zhang, W.H.; Song, S.X.; Xu, X.L.; Zhou, G.H. Effects of dietary glutamine and gamma-aminobutyric acid on performance, carcass characteristics and serum parameters in broilers under circular heat stress. Anim. Feed Sci. Technol. 2011, 168, 51–60. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Hu, H.; Chen, L.; Dai, S.; Li, J.; Bai, X. Effect of glutamine on antioxidant capacity and lipid peroxidation in the breast muscle of heat-stressed broilers via antioxidant genes and HSP70 pathway. Animals 2020, 10, 404. [Google Scholar] [CrossRef]
- Qaid, M.M.; Al-Garadi, M.A. Protein and amino acid metabolism in poultry during and after heat stress: A review. Animals 2021, 11, 1167. [Google Scholar] [CrossRef]
- Hu, H.; Bai, X.; Xu, K.; Zhang, C.; Chen, L. Effect of phloretin on growth performance, serum biochemical parameters and antioxidant profile in heat-stressed broilers. Poult. Sci. 2021, 100, 101217. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Bai, X.; Zhang, D.; Hu, H.; Wu, X.; Wen, A.; He, S.J.; Zhao, L. Dietary glutamine improves meat quality, skeletal muscle antioxidant capacity and glutamine metabolism in broilers under acute heat stress. J. Appl. Anim. Res. 2018, 46, 1412–1417. [Google Scholar]
- Bai, X.; Dai, S.; Li, J.; Xiao, S.; Wen, A.; Hu, H. Glutamine improves the growth performance, serum biochemical profile and antioxidant status in broilers under medium-term chronic heat stress. J. Appl. Poult. Res. 2019, 28, 1248–1254. [Google Scholar] [CrossRef]
- Mazzoni, M.; Zampiga, M.; Clavenzani, P.; Lattanzio, G.; Tagliavia, C.; Sirri, F. Effect of chronic heat stress on gastrointestinal histology and expression of feed intake-regulatory hormones in broiler chickens. Animal 2022, 16, 100600. [Google Scholar] [CrossRef]
- Khan, R.U.; Naz, S.; Ullah, H.; Ullah, Q.; Laudadio, V.; Qudratullah.; Bozzo, G.; Tufarelli, V. Physiological dynamics in broiler chickens under heat stress and possible mitigation strategies. Anim. Biotechnol. 2021, 2, 1–10. [Google Scholar] [CrossRef]
- Saracila, M.; Panaite, T.D.; Papuc, C.P.; Criste, R.D. Heat stress in broiler chickens and the effect of dietary polyphenols, with special reference to Willow (Salix spp.) bark supplements—A review. Antioxidants 2021, 10, 686. [Google Scholar] [CrossRef]
- Emami, N.K.; Jung, U.; Voy, B.; Dridi, S. Radical response: Effects of heat stress-induced oxidative stress on lipid metabolism in the avian liver. Antioxidants 2020, 10, 35. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.P.; Liu, Y.L.; Zhang, J.X.; Ding, K.N.; Lu, M.H.; He, Y.M. Heat stress in broilers of liver injury effects of heat stress on oxidative stress and autophagy in liver of broilers. Poult. Sci. 2022, 101, 102085. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Tsai, P.H.; Liu, J.J.; Yeh, C.L.; Chiu, W.C.; Yeh, S.L. Effects of glutamine supplementation on oxidative stress-related gene expression and antioxidant properties in rats with streptozotocin-induced type 2 diabetes. Br. J. Nutr. 2012, 107, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Bai, K.W.; Su, W.P.; Wang, A.A.; Zhang, L.L.; Huang, K.H.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poul. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Chattopadhyay, A. Nrf2–ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef]
- Gan, L.; Johnson, J.A. Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. BBA Mol. Basis Dis. 2014, 1842, 1208–1218. [Google Scholar] [CrossRef]
- Cheng, L.; Jin, Z.; Zhao, R.; Ren, K.; Deng, C.; Yu, S. Resveratrol attenuates inflammation and oxidative stress induced by myocardial ischemia-reperfusion injury: Role of Nrf2/ARE pathway. Int. J. Clin. Exp. Med. 2015, 8, 10420. [Google Scholar] [PubMed]
- Moine, L.; de Barboza, G.D.; Pérez, A.; Benedetto, M.; de Talamoni, N.T. Glutamine protects intestinal calcium absorption against oxidative stress and apoptosis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 212, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Dai, S.; Li, J.; Wen, A.; Bai, X. Glutamine improves heat stress–induced oxidative damage in the broiler thigh muscle by activating the nuclear factor erythroid 2–related 2/Kelch-like ECH-associated protein 1 signaling pathway. Poult. Sci. 2020, 99, 1454–1461. [Google Scholar] [CrossRef]
- Tolba, S.A.; Magnuson, A.D.; Sun, T.; Lei, X.G. Dietary supplemental microalgal astaxanthin modulates molecular profiles of stress, inflammation, and lipid metabolism in broiler chickens and laying hens under high ambient temperatures. Poult. Sci. 2020, 99, 4853–4860. [Google Scholar] [CrossRef]
- Hu, Y.; Lin, L.; Liu, K.; Liu, E.; Han, S.; Gong, Z.; Xiao, W. l-Theanine alleviates heat stress-induced impairment of immune function by regulating the p38 MAPK signalling pathway in mice. Food Funct. 2023, 14, 335–343. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhou, B.; Chen, H.; Lu, K.; Wang, Y. Oxidative stress activates the Nrf2-mediated antioxidant response and P38 MAPK pathway: A possible apoptotic mechanism induced by BDE-47 in rainbow trout (Oncorhynchus mykiss) gonadal RTG-2 cells. Environ. Pollut. 2021, 287, 117341. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Wang, K.; Khan, R.U.; Zhang, C.; Hu, H. Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers. Animals 2023, 13, 652. https://doi.org/10.3390/ani13040652
Bai X, Wang K, Khan RU, Zhang C, Hu H. Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers. Animals. 2023; 13(4):652. https://doi.org/10.3390/ani13040652
Chicago/Turabian StyleBai, Xi, Kunping Wang, Rifat Ullah Khan, Cheng Zhang, and Hong Hu. 2023. "Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers" Animals 13, no. 4: 652. https://doi.org/10.3390/ani13040652
APA StyleBai, X., Wang, K., Khan, R. U., Zhang, C., & Hu, H. (2023). Effect of Glutamine on the Growth Performance, Oxidative Stress, and Nrf2/p38 MAPK Expression in the Livers of Heat-Stressed Broilers. Animals, 13(4), 652. https://doi.org/10.3390/ani13040652