

Data on New Intermediate and Accidental Hosts Naturally Infected with Angiostrongylus cantonensis in La Gomera and Gran Canaria (Canary Islands, Spain)
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
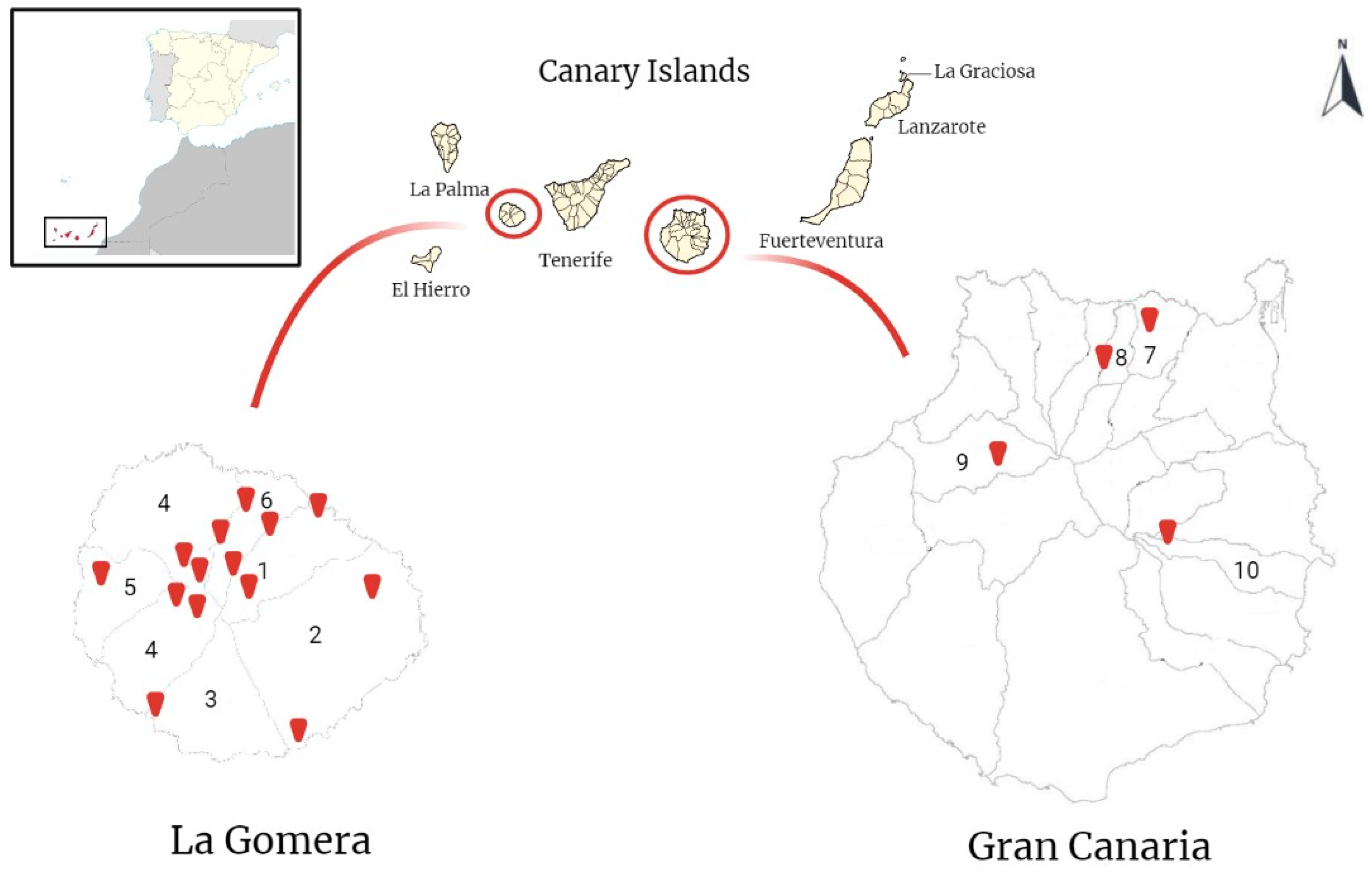
2.1. Samples Collection
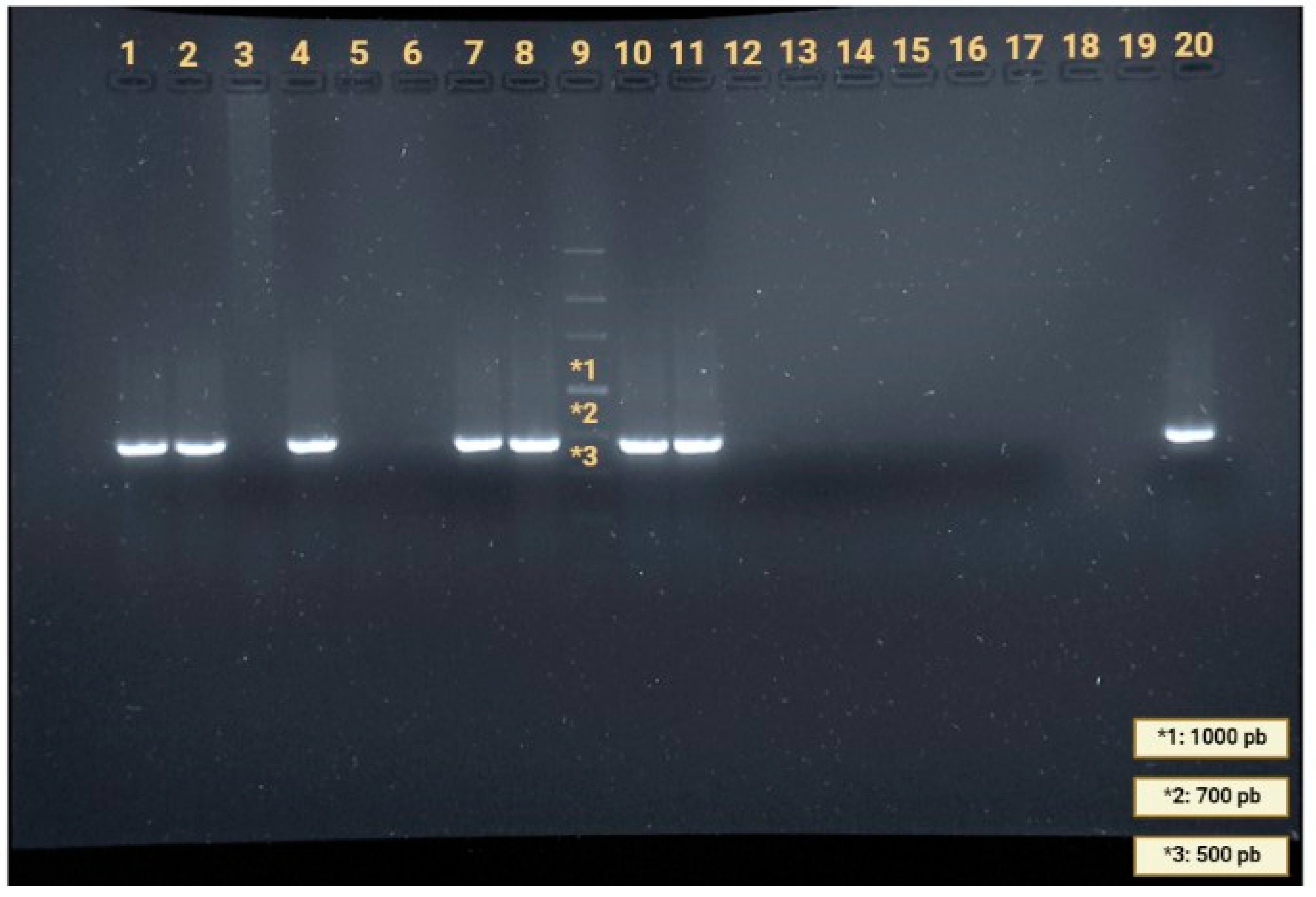
2.2. Search for Angiostrongylus cantonensis and Sample Isolation
2.3. DNA Extraction
2.4. PCR Amplification
2.5. Sequencing and Sequencing Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, D.Y.; Stewart, T.B.; Bauer, R.W.; Mitchell, M. Parastrongylus (=Angiostrongylus) cantonensis now endemic in Louisiana wildlife. J. Parasitol. 2002, 88, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Sullivan, J.J.; Bishop, H.S.; Hollingsworth, R.; da Silva, A.J. PCR-based detection of Angiostrongylus cantonensis in tissue and mucus secretions from molluscan hosts. Appl. Environ. Microbiol. 2007, 73, 1415–1419. [Google Scholar] [CrossRef]
- Chen, H.T. Un nouveau nématode pulmonaire, Pulmonema cantonensis, ng, n. sp. Ann. Parasitol. Hum. Comp. 1935, 13, 312–317. [Google Scholar] [CrossRef]
- Yong, H.S.; Eamsobhana, P. Definitive rodent hosts of the rat lungworm Angiostrongylus cantonensis. Raffles Bull. Zool. 2013, 29, 111–115. [Google Scholar]
- Beaver, P.C.; Rosn, L. Memorandum on the First Report of Angiostrongylus in Man, by Nomura and Lin, 1945. Am. J. Trop. Med. Hyg. 1964, 13, 589–590. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.S.; Johnson, S. Clinical Aspects of Eosinophilic Meningitis and Meningoencephalitis caused by Angiostrongylus cantonensis, the Rat Lungworm. Hawaii J. Med. Public Health 2013, 72, 35. [Google Scholar]
- Turck, H.C.; Fox, M.T.; Cowie, R.H. Paratenic hosts of Angiostrongylus cantonensis and their relation to human neuroangiostrongyliasis globally. One Health 2022, 15, 100426. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lu, L.; She, D.; Wen, Z.; Mo, Z.; Li, J.; Li, H. Eating Centipedes Can Result in Angiostrongylus cantonensis Infection: Two Case Reports and Pathogen Investigation. Am. J. Trop. Med. Hyg. 2018, 99, 743. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-P.; Lai, D.-H.; Zhu, X.-Q.; Chen, X.-G.; Lun, Z.-R. Human angiostrongyliasis. Lancet Infect. Dis. 2008, 8, 621–630. [Google Scholar] [CrossRef]
- Slom, T.J.; Cortese, M.M.; Gerber, S.I.; Jones, R.C.; Holtz, T.H.; Lopez, A.S.; Zambrano, C.H.; Sufit, R.L.; Sakilvaree, Y.; Chaicumpa, W.; et al. An outbreak of eosinophilic meningitis caused by Angiostrongylus cantonensis in travelers returning from the Caribbean. N. Engl. J. Med. 2002, 346, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Yii, C.-Y. Clinical Observations on Eosinophilic Meningitis and Meningoencephalitis caused by Angiostrongylus cantonensis on Taiwan. Am. J. Trop. Med. Hyg. 1976, 25, 233–249. [Google Scholar] [CrossRef]
- Bowden, D.K. Eosinophilic Meningitis in the New Hebrides: Two Outbreaks and two Deaths. Am. J. Trop. Med. Hyg. 1981, 30, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Chotmongkol, V.; Sawanyawisuth, K. Clinical manifestations and outcome of patients with severe eosinophilic meningoencephalitis presumably caused by Angiostrongylus cantonensis. Age 2002, 45, 30–61. [Google Scholar]
- Cowie, R.H. Biology, Systematics, Life Cycle, and Distribution of Angiostrongylus cantonensis, the Cause of Rat Lungworm Disease. Hawaii J. Med. Public Health 2013, 72, 6. [Google Scholar] [PubMed]
- Cowie, R.H.; Ansdell, V.; Panosian Dunavan, C.; Rollins, R.L. Neuroangiostrongyliasis: Global Spread of an Emerging Tropical Disease. Am. J. Trop. Med. Hyg. 2022, 107, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Foronda, P.; López-González, M.; Miquel, J.; Torres, J.; Segovia, M.; Abreu-Acosta, N.; Casanova, J.C.; Valladares, B.; Mas-Coma, S.; Bargues, M.D.; et al. Finding of Parastrongylus cantonensis (Chen, 1935) in Rattus rattus in Tenerife, Canary Islands (Spain). Acta Trop. 2010, 114, 123–127. [Google Scholar] [CrossRef]
- Paredes-Esquivel, C.; Sola, J.; Delgado-Serra, S.; Puig Riera, M.; Negre, N.; Miranda, M.Á.; Jurado-Rivera, J.A. Angiostrongylus cantonensis in North African hedgehogs as vertebrate hosts, Mallorca, Spain, October 2018. Eurosurveillance 2019, 24, 1900489. [Google Scholar] [CrossRef]
- Galán-Puchades, M.T.; Gómez-Samblás, M.; Osuna, A.; Sáez-Durán, S.; Bueno-Marí, R.; Fuentes, M.V. Autochthonous Angiostrongylus cantonensis Lungworms in Urban Rats, Valencia, Spain, 2021. Emerg. Infect. Dis. 2022, 28, 2564–2567. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Steinmann, P.; Yang, G.J.; Yang, K.; Zhou, X.N.; Utzinger, J. The emergence of angiostrongyliasis in the People’s Republic of China: The interplay between invasive snails, climate change and transmission dynamics. Fresh Biol. 2011, 56, 717–734. [Google Scholar] [CrossRef]
- Stokes, V.L.; Spratt, D.M.; Banks, P.B.; Pech, R.P.; Williams, R.L. Occurrence of Angiostrongylus species (Nematoda) in populations of Rattus rattus and Rattus fuscipes in coastal forests of south-eastern Australia. Aust. J. Zool. 2007, 55, 177–184. [Google Scholar] [CrossRef]
- York, E.M.; Butler, C.J.; Lord, W.D. Global Decline in Suitable Habitat for Angiostrongylus (=Parastrongylus) cantonensis: The Role of Climate Change. PLoS ONE 2014, 9, e103831. [Google Scholar] [CrossRef] [PubMed]
- Barratt, J.; Chan, D.; Sandaradura, I.; Malik, R.; Spielman, D.; Lee, R.; Lee, R.; Marriott, D.; Harkness, J.; Ellis, J.; et al. Angiostrongylus cantonensis: A review of its distribution, molecular biology and clinical significance as a human pathogen. Parasitology 2016, 143, 1087–1118. [Google Scholar] [CrossRef]
- Martín-Carrillo, N.; Feliu, C.; Abreu-Acosta, N.; Izquierdo-Rodriguez, E.; Dorta-Guerra, R.; Miquel, J.; Abreu-Yanes, E.; Martín-Alonso, A.; García-Livia, K.A.; Quispe-Ricalde, M.A.; et al. A peculiar distribution of the emerging nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent introduction or isolation effect? Animals 2021, 11, 1267. [Google Scholar] [CrossRef] [PubMed]
- Instituto Canario de Estadística (ISTAC). Available online: http://www.gobiernodecanarias.org/istac/estadisticas/demografia/poblacion/cifraspadronales/E30245A.html (accessed on 6 March 2023).
- Martin-Alonso, A.; Abreu-Yanes, E.; Feliu, C.; Mas-Coma, S.; Bargues, M.D.; Valladares, B.; Foronda, P. Intermediate Hosts of Angiostrongylus cantonensis in Tenerife, Spain. PLoS ONE 2015, 10, e0120686. [Google Scholar] [CrossRef] [PubMed]
- Anettová, L.; Izquierdo-Rodriguez, E.; Foronda, P.; Baláž, V.; Novotný, L.; Modrý, D. Endemic lizard Gallotia galloti is a paratenic host of invasive Angiostrongylus cantonensis in Tenerife, Spain. Parasitology 2022, 149, 934–939. [Google Scholar] [CrossRef] [PubMed]
- López, C.; Clemente, S.; Almeida, C.; Brito, A.; Hernández, M. A genetic approach to the origin of Millepora sp. in the eastern Atlantic. Coral Reefs 2015, 34, 631–638. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; da Silva, A.C.A.; Teem, J.L.; Hollingsworth, R.; Bishop, H.; Graeff-Teixeira, C.; da Silva, A.J. Improved Molecular Detection of Angiostrongylus cantonensis in Mollusks and Other Environmental Samples with a Species-Specific Internal Transcribed Spacer 1-Based TaqMan Assay. Appl. Environ. Microbiol. 2010, 76, 5287–5289. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Rodriguez, E.; Anettová, L.; Hrazdilová, K.; Foronda, P.; Modrý, D. Range of metastrongylids (superfamily Metastrongyloidea) of veterinary and public health interest present in livers of the endemic lizard Gallotia galloti from Tenerife, Canary Islands, Spain. Parasites Vectors 2023, 16, 81. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Wang, L.-C.; Chao, D.; Chen, E.-R. Experimental infection routes of Angiostrongylus cantonensis in mice. J. Helminthol. 1991, 65, 296–300. [Google Scholar] [CrossRef]
- Sugaya, H.; Aoki, M.; Abe, T.; Ishida, K.; Yoshimura, K. Cytokine responses in mice infected with Angiostrongylus cantonensis. Parasitol. Res. 1996, 83, 10–15. [Google Scholar] [CrossRef]
- Nogales, M.; Rodríguez Luengo, J.L.; Delgado, G.; Quilis, V.; Trujullo, O. The diet of feral cats (Felis catus) on Alegranza Island (north of Lanzarote, Canary Islands). Folia Zool. 1992, 41, 209–212. [Google Scholar]
- Medina, F.M.; Bonnaud, E.; Vidal, E.; Tershy, B.R.; Zavaleta, E.S.; Donlan, C.J.; Keitt, B.S.; Le Corre, M.; Horwath, S.V.; Nogales, M. A global review of the impacts of invasive cats on insular endangered vertebrates. Glob. Chang. Biol. 2010, 17, 3503–3510. [Google Scholar] [CrossRef]
- Bonnaud, E.; Medina, F.M.; Vidal, E.; Nogales, M.; Tershy, B.; Zavaleta, E.; Donlan, C.J.; Keitt, B.; Le Corre, M.; Horwath, S.V. The diet of feral cats on islands: A review and a call for more studies. Biol. Invasions 2011, 13, 581–603. [Google Scholar] [CrossRef]
- Gobierno de Canarias. Banco de Datos de Biodiversidad de Canarias. Available online: http://www.biodiversidadcanarias.es/biota (accessed on 29 March 2023).
- Kim, J.R.; Hayes, K.A.; Yeung, N.W.; Cowie, R.H. Diverse gastropod hosts of Angiostrongylus cantonensis, the rat lungworm, globally and with a focus on the Hawaiian Islands. PLoS ONE 2014, 9, e94969. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.R.; Jefferies, R.; Krajewski, M.; Ward, P.; Shaw, S.E. Canine pulmonary angiostrongylosis: The influence of climate on parasite distribution. Parasitol. Int. 2009, 58, 406–410. [Google Scholar] [CrossRef]
- Kutz, S.J.; Hoberg, E.P.; Polley, L.; Jenkins, E.J. Global warming is changing the dynamics of Arctic host–parasite systems. Proc. R. Soc. Lond. Biol. 2005, 272, 2571–2576. [Google Scholar] [CrossRef] [PubMed]
- Salb, A.L.; Barkema, H.W.; Elkin, B.T.; Thompson, R.C.A.; Whiteside, D.P.; Black, S.R.; Dubey, J.P.; Kutz, S.J. Dogs as Sources and Sentinels of Parasites in Humans and Wildlife, Northern Canada. Emerg. Infect. Dis. 2008, 14, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Eamsobhana, P. Review paper eosinophilic meningitis caused by Angiostrongylus cantonensis a neglected disease with escalating importance. Trop. Biomed 2014, 31, 569–578. [Google Scholar] [PubMed]
- Nguyen, Y.; Rossi, B.; Argy, N.; Baker, C.; Nickel, B.; Marti, H.; Zarrouk, V.; Houzé, S.; Fantin, B.; Lefort, A. Autochthonous case of eosinophilic meningitis caused by Angiostrongylus cantonensis, France, 2016. Emerg. Infect. Dis. 2017, 23, 1045. [Google Scholar] [CrossRef] [PubMed]
- Halliday, J.E.; Meredith, A.L.; Knobel, D.L.; Shaw, D.J.; Bronsvoort, B.M.D.C.; Cleaveland, S. A framework for evaluating animals as sentinels for infectious disease surveillance. J. R. Soc. Interface 2007, 4, 973–984. [Google Scholar] [CrossRef]
- Morgan, E.R.; Modry, D.; Paredes-Esquivel, C.; Foronda, P.; Traversa, D. Angiostrongylosis in Animals and Humans in Europe. Pathogens 2021, 10, 1236. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | Island | Sample Size | Total |
|---|---|---|---|
| Felis catus | La Gomera | 40 | 40 |
| Mus musculus | La Gomera | 34 | 60 |
| Gran Canaria | 26 | ||
| Rattus norvegicus | La Gomera | 4 | 4 |
| Rattus rattus | La Gomera | 33 | 40 |
| Gran Canaria | 7 | ||
| Insulivitrina oromii | La Gomera | 8 | 8 |
| Insulivitrina emmersoni | La Gomera | 28 | 28 |
| Limacus flavus | La Gomera | 10 | 10 |
| Milax gagates | Gran Canaria | 24 | 24 |
| Total | 214 |
| Island | Location | Host Species | Prevalence of Angiostrongylus cantonensis P (%) * (+/n) | Total P (%) * (+/n) |
|---|---|---|---|---|
| La Gomera | Agulo | Felis catus | 0 (0/2) | 11.11 (1/9) |
| Mus musculus | 0 (0/5) | |||
| Insulivitrina oromii | 50 (1/2) | |||
| Alajeró | Felis catus | 0 (0/1) | 0 (0/6) | |
| Mus musculus | 0 (0/3) | |||
| Insulivitrina oromii | 0 (0/2) | |||
| Hermigua | Felis catus | 12.5 (1/8) | 25% (9/36) | |
| Mus muculus | 0 (0/15) | |||
| Insulivitrina oromii | 66.06 (2/3) | |||
| Limacus flavus | 60 (6/10) | |||
| San Sebastián de la Gomera | Mus musculus | 0 (0/6) | 0 (0/6) | |
| Valle Gran Rey | Felis catus | 0 (0/6) | 0 (0/7) | |
| Insulivitrina oromii | 0 (0/1) | |||
| Vallehermoso | Felis catus | 17.39 (4/23) | 23.21 (13/56) | |
| Mus musculus | 0 (0/5) | |||
| Insulivitrina emmersoni | 32.14 (9/28) | |||
| Total of La Gomera | 19.16 (23/120) | |||
| Gran Canaria | Artenara | Mus musculus | 0 (0/2) | 0 (0/2) |
| Arucas | Mus musculus | 16.66 (1/6) | 16.66 (1/6) | |
| Firgas | Mus musculus | 100 (1/1) | 8 (2/25) | |
| Milax gagates | 4.16 (1/24) | |||
| Ingenio | Mus musculus | 41.17 (7/17) | 41.17 (7/17) | |
| Total of Gran Canaria | 20 (10/50) | |||
| Total | 13.52% (33/170) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Carrillo, N.; Baz-González, E.; García-Livia, K.; Amaro-Ramos, V.; Abreu-Acosta, N.; Miquel, J.; Abreu-Yanes, E.; Pino-Vera, R.; Feliu, C.; Foronda, P. Data on New Intermediate and Accidental Hosts Naturally Infected with Angiostrongylus cantonensis in La Gomera and Gran Canaria (Canary Islands, Spain). Animals 2023, 13, 1969. https://doi.org/10.3390/ani13121969
Martin-Carrillo N, Baz-González E, García-Livia K, Amaro-Ramos V, Abreu-Acosta N, Miquel J, Abreu-Yanes E, Pino-Vera R, Feliu C, Foronda P. Data on New Intermediate and Accidental Hosts Naturally Infected with Angiostrongylus cantonensis in La Gomera and Gran Canaria (Canary Islands, Spain). Animals. 2023; 13(12):1969. https://doi.org/10.3390/ani13121969
Chicago/Turabian StyleMartin-Carrillo, Natalia, Edgar Baz-González, Katherine García-Livia, Virginia Amaro-Ramos, Néstor Abreu-Acosta, Jordi Miquel, Estefanía Abreu-Yanes, Román Pino-Vera, Carlos Feliu, and Pilar Foronda. 2023. "Data on New Intermediate and Accidental Hosts Naturally Infected with Angiostrongylus cantonensis in La Gomera and Gran Canaria (Canary Islands, Spain)" Animals 13, no. 12: 1969. https://doi.org/10.3390/ani13121969
APA StyleMartin-Carrillo, N., Baz-González, E., García-Livia, K., Amaro-Ramos, V., Abreu-Acosta, N., Miquel, J., Abreu-Yanes, E., Pino-Vera, R., Feliu, C., & Foronda, P. (2023). Data on New Intermediate and Accidental Hosts Naturally Infected with Angiostrongylus cantonensis in La Gomera and Gran Canaria (Canary Islands, Spain). Animals, 13(12), 1969. https://doi.org/10.3390/ani13121969