
Changes in Dynamic Thiol/Disulfide Homeostasis, and Substance P, B-Endorphin and α-Tocopherol Concentrations in the Spinal Cord of Chronically Lame Dairy Cows
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Lameness Assessment
2.3. Spinal Cord Processing, Protein Extraction, and Quantification
2.4. Spinal Dynamic Thiol/Disulfide Homeostasis Assay
2.5. Spinal Substance P and β-Endorphin Immunoassay
2.6. Determination of α-Tocopherol Concentration by High-Performance Liquid Chromatography (HPLC)
2.7. Statistical Analysis
3. Results
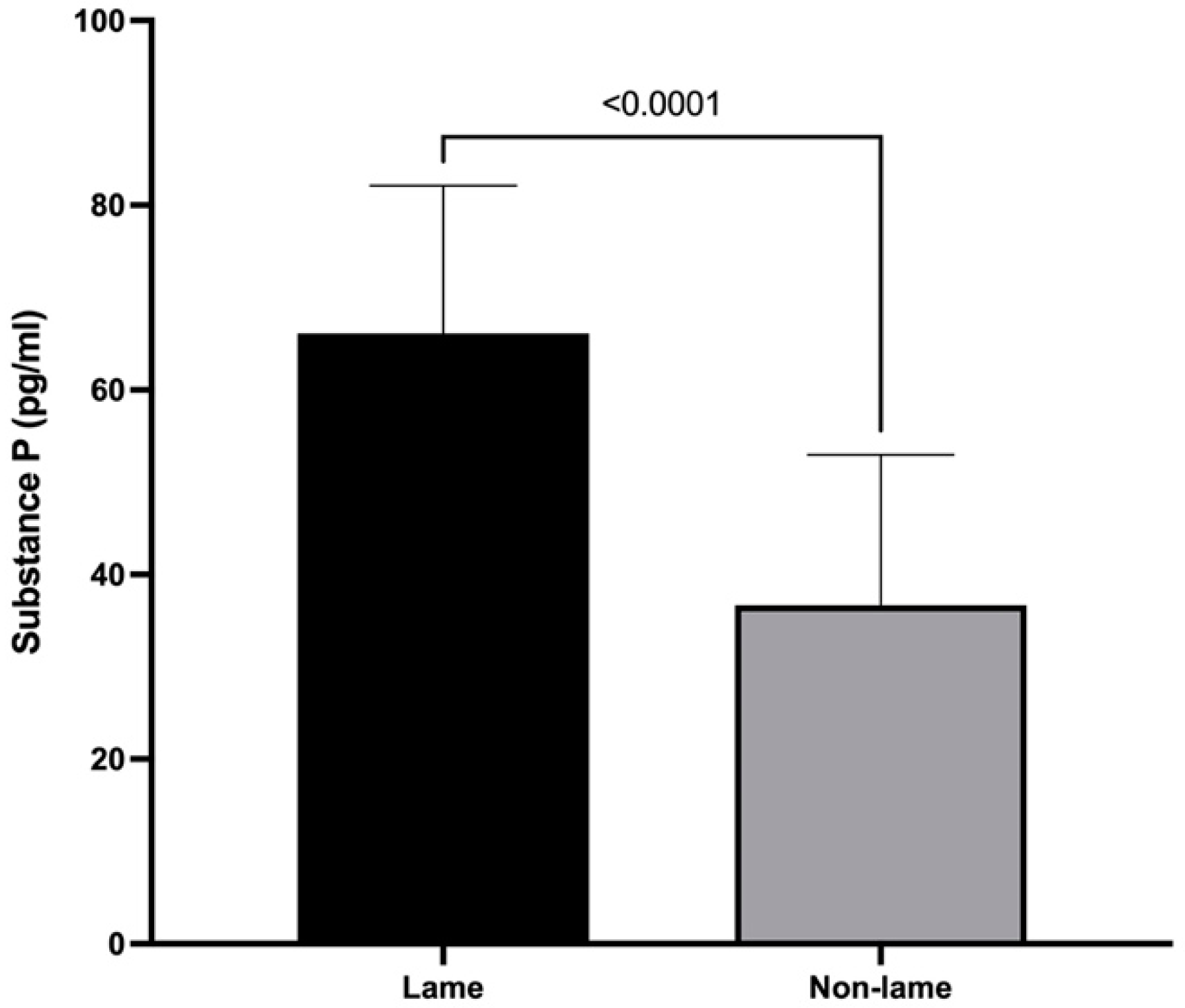
3.1. Substance P Spinal Cord Concentrations
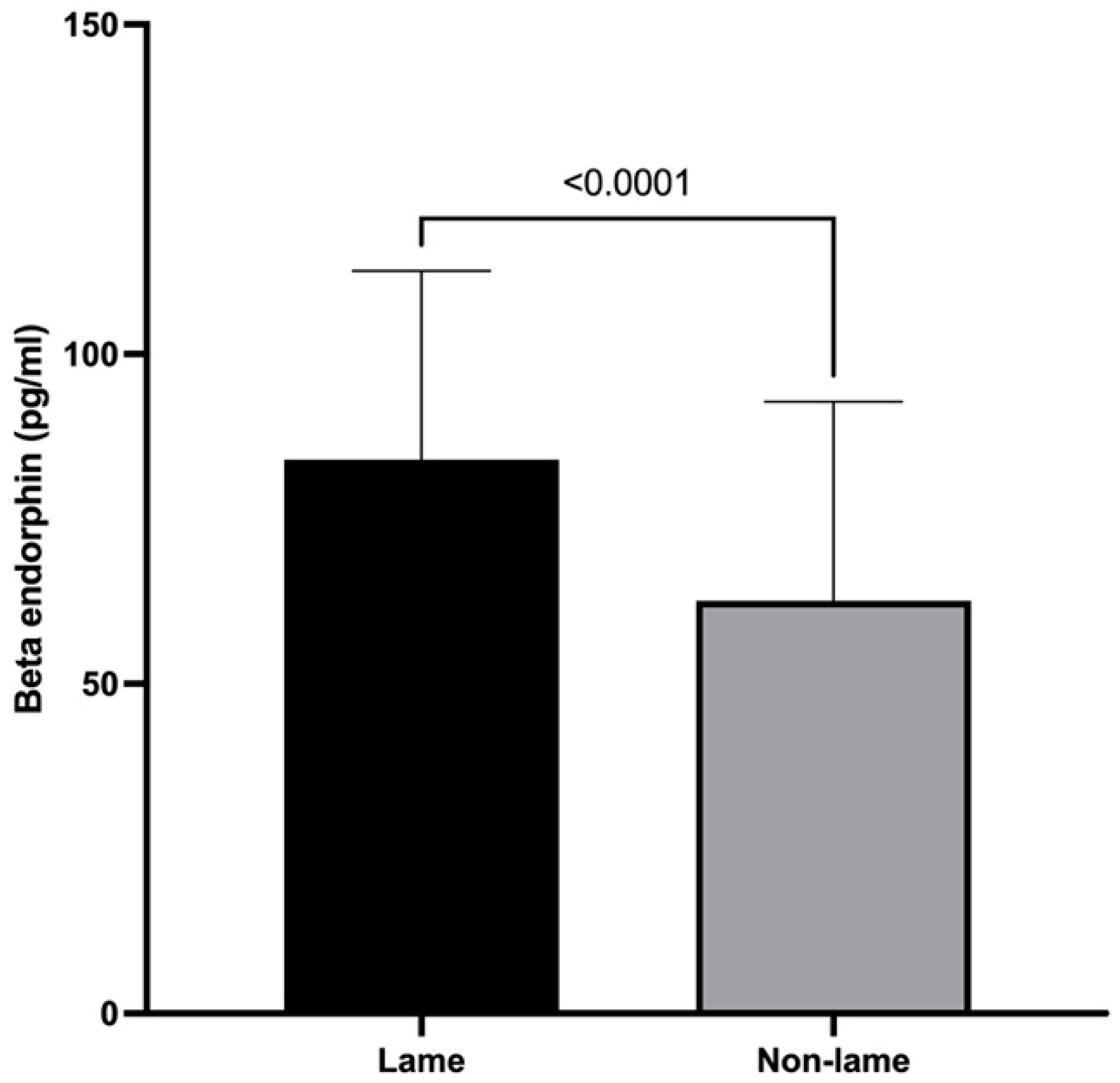
3.2. β-Endorphin Spinal Cord Concentrations
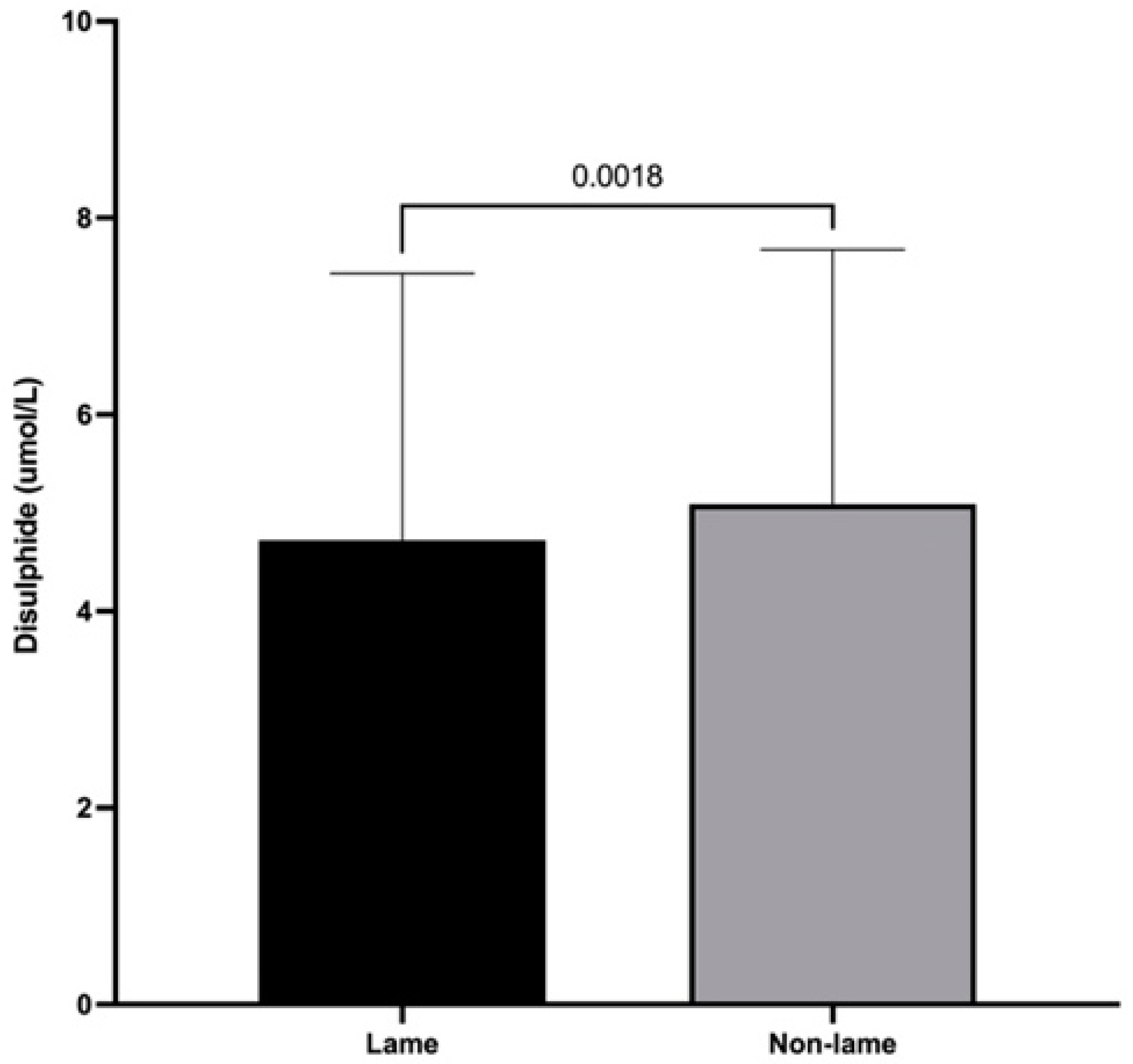
3.3. Dynamic Thiol/Disulfide Homeostasis
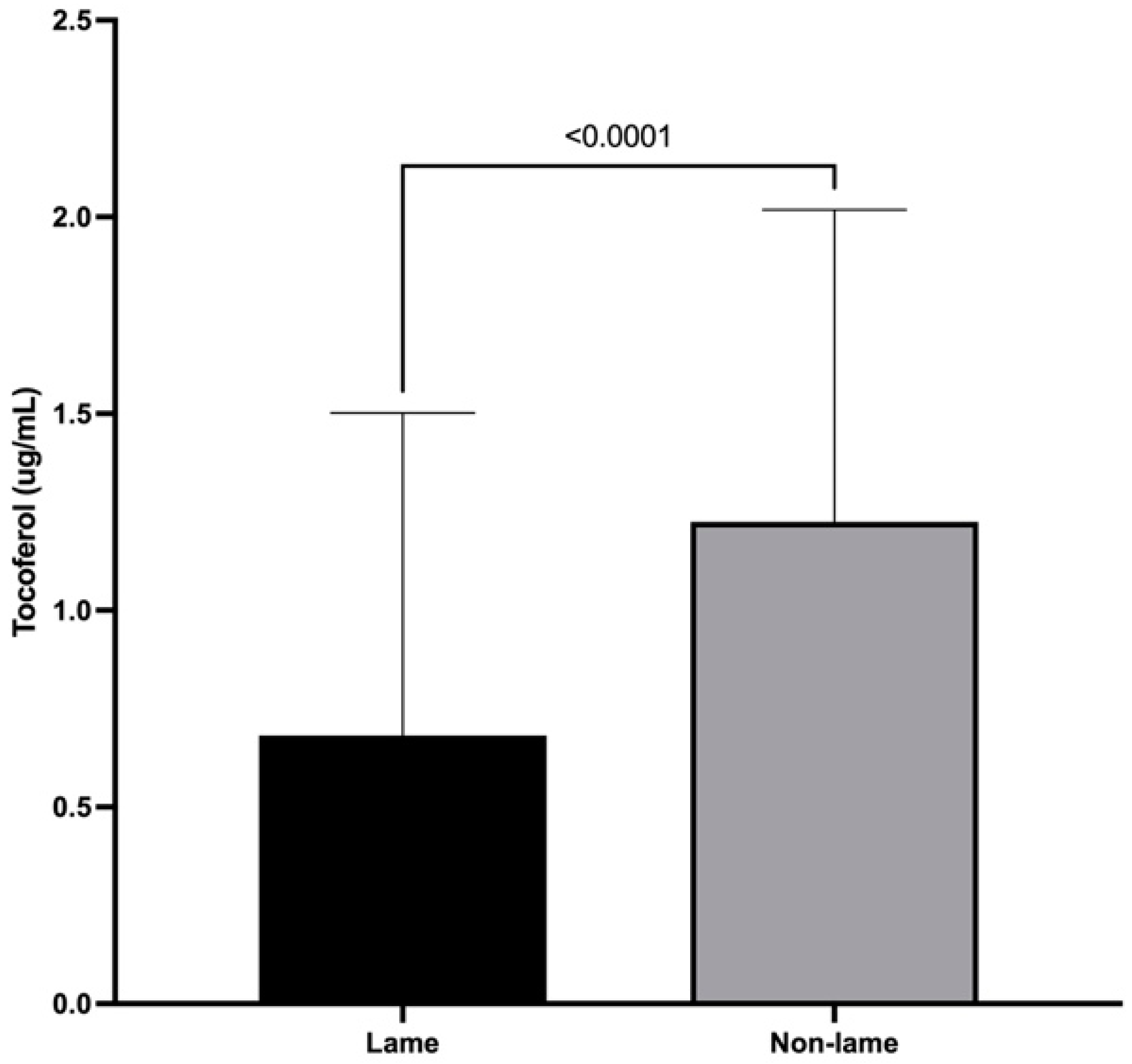
3.4. α-Tocopherol Spinal Cord Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coetzee, J.F.; Shearer, J.K.; Stock, M.L.; Kleinhenz, M.D.; van Amstel, S.R. An Update on the Assessment and Management of Pain Associated with Lameness in Cattle. Vet. Clin. N. Am.—Food Anim. Pract. 2017, 33, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.R.; Herzberg, D.E.; Werner, M.P.; Müller, H.Y.; Bustamante, H.A. Plasma concentration of norepinephrine, β-endorphin, and substance P in lame dairy cows. J. Vet. Res. 2018, 62, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Herzberg, D.; Strobel, P.; Chihuailaf, R.; Ramirez-Reveco, A.; Müller, H.; Werner, M.; Bustamante, H. Spinal reactive oxygen species and oxidative damage mediate chronic pain in lame dairy cows. Animals 2019, 9, 693. [Google Scholar] [CrossRef]
- Whay, H.R.; Shearer, J.K. The Impact of Lameness on Welfare of the Dairy Cow. Vet. Clin. N. Am.—Food Anim. Pract. 2017, 33, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Little, J.W.; Doyle, T.; Neumann, W.L. Roles of reactive oxygen and nitrogen species in pain. Free Radic. Biol. Med. 2011, 51, 951–966. [Google Scholar] [CrossRef]
- Haddad, J.J.; Land, S.C. Redox/ROS regulation of lipopolysaccharide-induced mitogen-activated protein kinase (MAPK) activation and MAPK-mediated TNF-a biosynthesis. Br. J. Pharmacol. 2002, 135, 520–536. [Google Scholar] [CrossRef]
- Gao, X.; Kim, H.K.; Chung, J.M.; Chung, K. Reactive oxygen species (ROS) are involved in enhancement of NMDA-receptor phosphorylation in animal models of pain. Pain 2007, 131, 262–271. [Google Scholar] [CrossRef]
- Kaur, U.; Banerjee, P.; Bir, A.; Sinha, M.S.; Biswas, A.; Chakrabarti, S. Reactive Oxygen Species, Redox Signaling and Neuroinflammation in Alzheimer’s Disease: The NF-κB Connection. Curr. Top. Med. Chem. 2015, 15, 446–457. [Google Scholar] [CrossRef]
- Nishio, N.; Taniguchi, W.; Sugimura, Y.; Takiguchi, N.; Yamanaka, M.; Kiyoyuki, Y.; Yamada, H.; Miyazaki, N.; Yoshida, M.; Nakatsuka, T. Reactive oxygen species enhance excitatory synaptic transmission in rat spinal dorsal horn neurons by activating TRPA1 and TRPV1 channels. Neuroscience 2013, 247, 201–212. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Sen, C.K.; Packer, L. Thiol homeostasis and supplements in physical exercise. Am. J. Clin. Nutr. 2000, 72, 653S–669S. [Google Scholar] [CrossRef] [PubMed]
- Rehder, D.S.; Borges, C.R. Cysteine sulfenic acid as an intermediate in disulfide bond formation and nonenzymatic protein folding. Biochemistry 2010, 49, 7748–7755. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P.; Liang, Y. Measuring the poise of thiol/disulfide couples in vivo. Free Radic. Biol. Med. 2009, 47, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Erel, O.; Neselioglu, S. A novel and automated assay for thiol/disulphide homeostasis. Clin. Biochem. 2014, 47, 326–332. [Google Scholar] [CrossRef]
- Eroglu, S.; Haskul, I.; Aziz, V.; Yurtcu, E.; Karatas, F.; Neşelioğlu, S.; Erel, O. Dynamic thiol/disulphide homeostasis in patients with Uterine Myoma. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 216, 24–26. [Google Scholar] [CrossRef]
- Karatas, G.; Gunduz, R.; Haskul, I.; Ustun, B.; Neselioglu, S.; Karatas, F.; Akyuz, M.; Erel, O. Dynamic thiol and disulphide homoeostasis in fibromyalgia. Arch. Med. Sci. 2020, 16, 597–602. [Google Scholar] [CrossRef]
- Tuzcu, A.; Baykara, R.A.; Alışık, M.; Omma, A.; Acet, G.K.; Dogan, E.; Cure, M.C.; Duygun, F.; Cure, E.; Erel, O. Alteration of Thiol-Disulfide Homeostasis in Fibromyalgia Syndrome. Acta Med. 2019, 62, 12–18. [Google Scholar] [CrossRef]
- Chudinova, V.V.; Alekseev, S.M.; I Zakharova, E.; Evstigneeva, R.P. Lipid peroxidation and mechanism of antioxidant effect of vitamin E. Bioorg. Khim. 1994, 20, 1029–1046. Available online: https://pubmed.ncbi.nlm.nih.gov/7826404/ (accessed on 16 December 2022).
- Herrera, E.; Barbas, C. Vitamin E: Action, metabolism and perspectives. J. Physiol. Biochem. 2001, 57, 43–56. [Google Scholar] [CrossRef]
- Wefers, H.; Sies, H. The protection by ascorbate and glutathione against microsomal lipid peroxidation is dependent on vitamin E. Eur. J. Biochem. 1988, 174, 353–357. [Google Scholar] [CrossRef]
- Schneider, C. Chemistry and biology of vitamin E. Mol. Nutr. Food Res. 2005, 49, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Tasinato, A.; Boscoboinik, D.; Bartoli, G.M.; Maroni, P.; Azzi, A. d-α-tocopherol inhibition of vascular smooth muscle cell proliferation occurs at physiological concentrations, correlates with protein kinase C inhibition, and is independent of its antioxidant properties. Proc. Natl. Acad. Sci. USA 1995, 92, 12190–12194. [Google Scholar] [CrossRef] [PubMed]
- Morani, A.S.; Bodhankar, S.L. Early co-administration of vitamin E acetate and methylcobalamin improves thermal hyperalgesia and motor nerve conduction velocity following sciatic nerve crush injury in rats. Pharmacol. Rep. 2010, 62, 405–409. [Google Scholar] [CrossRef]
- Milligan, E.D.; Watkins, L.R. Pathological and protective roles of glia in chronic pain. Nat. Rev. Neurosci. 2009, 10, 23–36. [Google Scholar] [CrossRef]
- Watkins, L.R.; Maier, S.F. Immune regulation of central nervous system functions: From sickness responses to pathological pain. J. Intern. Med. 2005, 257, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Li, W.W.; Guo, T.Z.; Shi, X.; Sun, Y.; Wei, T.; Clark, D.; Kingery, W. Substance P spinal signaling induces glial activation and nociceptive sensitization after fracture. Neuroscience 2015, 310, 73–90. [Google Scholar] [CrossRef]
- Drew, G.M.; Lau, B.K.; Vaughan, C.W. Substance P drives endocannabinoid-mediated disinhibition in a midbrain descending analgesic pathway. J. Neurosci. 2009, 29, 7220–7229. [Google Scholar] [CrossRef]
- Bán, E.G.; Brassai, A.; Vizi, E.S. The role of the endogenous neurotransmitters associated with neuropathic pain and in the opioid crisis: The innate pain-relieving system. Brain Res. Bull. 2020, 155, 129–136. [Google Scholar] [CrossRef]
- De Felipe, C.; Herrero, J.F.; O’Brien, J.A.; Palmer, J.A.; Doyle, C.A.; Smith, A.J.H.; Laird, J.M.A.; Belmonte, C.; Cervero, F.; Hunt, S.P. Altered nociception, analgesia and aggression in mice lacking the receptor for substance P. Nature 1998, 392, 394–397. [Google Scholar] [CrossRef]
- Holden, J.E.; Jeong, Y.; Forrest, J.M. The endogenous opioid system and clinical pain management. AACN Clin. Issues 2005, 16, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Cesselin, F.; Benoliel, J.; Bourgoin, S.; Collin, E.; Pohl, M.; Hamon, M. Spinal mechanisms of opioid analgesia. In Opioids in Pain Control: Basic and Clinical Aspects; Stein, C., Ed.; Cambridge University Press: Cambridge, UK, 1990; pp. 70–95. [Google Scholar]
- Niikura, K.; Narita, M.; Butelman, E.R.; Kreek, M.J.; Suzuki, T. Neuropathic and chronic pain stimuli downregulate central μ-opioid and dopaminergic transmission. Trends Pharmacol. Sci. 2010, 31, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Veening, J.G.; Gerrits, P.O.; Barendregt, H.P. Volume transmission of beta-endorphin via the cerebrospinal fluid; a review. Fluids Barriers CNS 2012, 9, 16. [Google Scholar] [CrossRef]
- Reader, J.D.; Green, M.J.; Kaler, J.; Mason, S.A.; Green, L.E. Effect of mobility score on milk yield and activity in dairy cattle. J. Dairy Sci. 2011, 94, 5045–5052. [Google Scholar] [CrossRef] [PubMed]
- Chihuailaf, R.H.; González, C.S.; Wittwer, F.; Contreras, P.A. Plasma retinol concentration in grazing heifers: First data obtained from a dairy herd in the south of Chile. Concentraciones plasmáticas de retinol en vaquillas a pastoreo: Primeros valores obtenidos en un rebaño del sur de Chile. Arch. Med. Vet. 2008, 68, 65–68. [Google Scholar]
- Bekhbat, M.; Rowson, S.A.; Neigh, G.N. Checks and balances: The glucocorticoid receptor and NFĸB in good times and bad. Front. Neuroendocrinol. 2017, 46, 15–31. [Google Scholar] [CrossRef]
- Yang, K.; Takeuchi, K.; Wei, F.; Dubner, R.; Ren, K. Activation of group i mGlu receptors contributes to facilitation of NMDA receptor membrane current in spinal dorsal horn neurons after hind paw inflammation in rats. Eur. J. Pharmacol. 2011, 670, 509–518. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central modulation of pain. J. Clin. Investig. 2010, 120, 3779–3787. [Google Scholar] [CrossRef]
- Whay, H.; Waterman, A.; Webster, A.; O’Brien, J. The influence of lesion type on the duration of hyperalgesia associated with hindlimb lameness in dairy cattle. Vet. J. 1998, 156, 23–29. [Google Scholar] [CrossRef]
- Tadich, N.; Flor, E.; Green, L. Associations between hoof lesions and locomotion score in 1098 unsound dairy cows. Vet. J. 2010, 184, 60–65. [Google Scholar] [CrossRef]
- Costigan, M.; Scholz, J.; Woolf, C.J. Neuropathic Pain: A Maladaptive Response of the Nervous System to Damage. Annu. Rev. Neurosci. 2009, 32, 1–32. [Google Scholar] [CrossRef]
- Niikura, K.; Narita, M.; Narita, M.; Nakamura, A.; Okutsu, D.; Ozeki, A.; Kurahashi, K.; Kobayashi, Y.; Suzuki, M.; Suzuki, T. Direct evidence for the involvement of endogenous β-endorphin in the suppression of the morphine-induced rewarding effect under a neuropathic pain-like state. Neurosci. Lett. 2008, 435, 257–262. [Google Scholar] [CrossRef]
- Le Roy, C.; Laboureyras, E.; Gavello-Baudy, S.; Chateauraynaud, J.; Laulin, J.P.; Simonnet, G. Endogenous opioids released during non-nociceptive environmental stress induce latent pain sensitization via a NMDA-dependent process. J. Pain 2011, 12, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Fidan, F.; Alkan, B.M.; Uğurlu, F.G.; Bozkurt, S.; Sezer, N.; Biçer, C.; Erel, Ö.; Ardiçoğlu, Ö.; Akkuş, S. Dynamic thiol/disulphide homeostasis in patients with fibromyalgia. Arch. Rheumatol. 2017, 32, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Matteucci, E.; Giampietro, O. Thiol signalling network with an eye to diabetes. Molecules 2010, 15, 8890–8903. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, A.; Sarcar, B.; Kahali, S.; Yuan, Z.; Johnson, J.J.; Adam, K.-P.; Kensicki, E.; Chinnaiyan, P. Cysteine catabolism: A novel metabolic pathway contributing to glioblastoma growth. Cancer Res. 2014, 74, 787–796. [Google Scholar] [CrossRef]
- Smeyne, M.; Smeyne, R.J. Glutathione metabolism and Parkinson’s disease. Free Radic. Biol. Med. 2013, 62, 13–25. [Google Scholar] [CrossRef]
- Lemke, M.; Frei, B.; Ames, B.N.; Faden, A.I. Decreases in tissue levels of ubiquinol-9 and -10, ascorbate and α-tocopherol following spinal cord impact trauma in rats. Neurosci. Lett. 1990, 108, 201–206. [Google Scholar] [CrossRef]
- Riffel, A.P.K.; Santos, M.; De Souza, J.; Scheid, T.; Horst, A.; Kolberg, C.; Bello-Klein, A.; Partata, W. Treatment with ascorbic acid and α-tocopherol modulates oxidative-stress markers in the spinal cord of rats with neuropathic pain. Braz. J. Med. Biol. Res. 2018, 51, e7079. [Google Scholar] [CrossRef]
- Park, E.S.; Gao, X.; Chung, J.M.; Chung, K. Levels of mitochondrial reactive oxygen species increase in rat neuropathic spinal dorsal horn neurons. Neurosci. Lett. 2006, 391, 108–111. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, H.; Herzberg, D.; Chihuailaf, R.; Strobel, P.; Werner, M.; Bustamante, H. Changes in Dynamic Thiol/Disulfide Homeostasis, and Substance P, B-Endorphin and α-Tocopherol Concentrations in the Spinal Cord of Chronically Lame Dairy Cows. Animals 2023, 13, 1620. https://doi.org/10.3390/ani13101620
Müller H, Herzberg D, Chihuailaf R, Strobel P, Werner M, Bustamante H. Changes in Dynamic Thiol/Disulfide Homeostasis, and Substance P, B-Endorphin and α-Tocopherol Concentrations in the Spinal Cord of Chronically Lame Dairy Cows. Animals. 2023; 13(10):1620. https://doi.org/10.3390/ani13101620
Chicago/Turabian StyleMüller, Heine, Daniel Herzberg, Ricardo Chihuailaf, Pablo Strobel, Marianne Werner, and Hedie Bustamante. 2023. "Changes in Dynamic Thiol/Disulfide Homeostasis, and Substance P, B-Endorphin and α-Tocopherol Concentrations in the Spinal Cord of Chronically Lame Dairy Cows" Animals 13, no. 10: 1620. https://doi.org/10.3390/ani13101620
APA StyleMüller, H., Herzberg, D., Chihuailaf, R., Strobel, P., Werner, M., & Bustamante, H. (2023). Changes in Dynamic Thiol/Disulfide Homeostasis, and Substance P, B-Endorphin and α-Tocopherol Concentrations in the Spinal Cord of Chronically Lame Dairy Cows. Animals, 13(10), 1620. https://doi.org/10.3390/ani13101620